
Correspondence between the Compositional and Aromatic Diversity of Leaf and Fruit Essential Oils and the Pomological Diversity of 43 Sweet Oranges (Citrus x aurantium var sinensis L.)
 ,
,  and
and
Abstract
:1. Introduction
2. Results
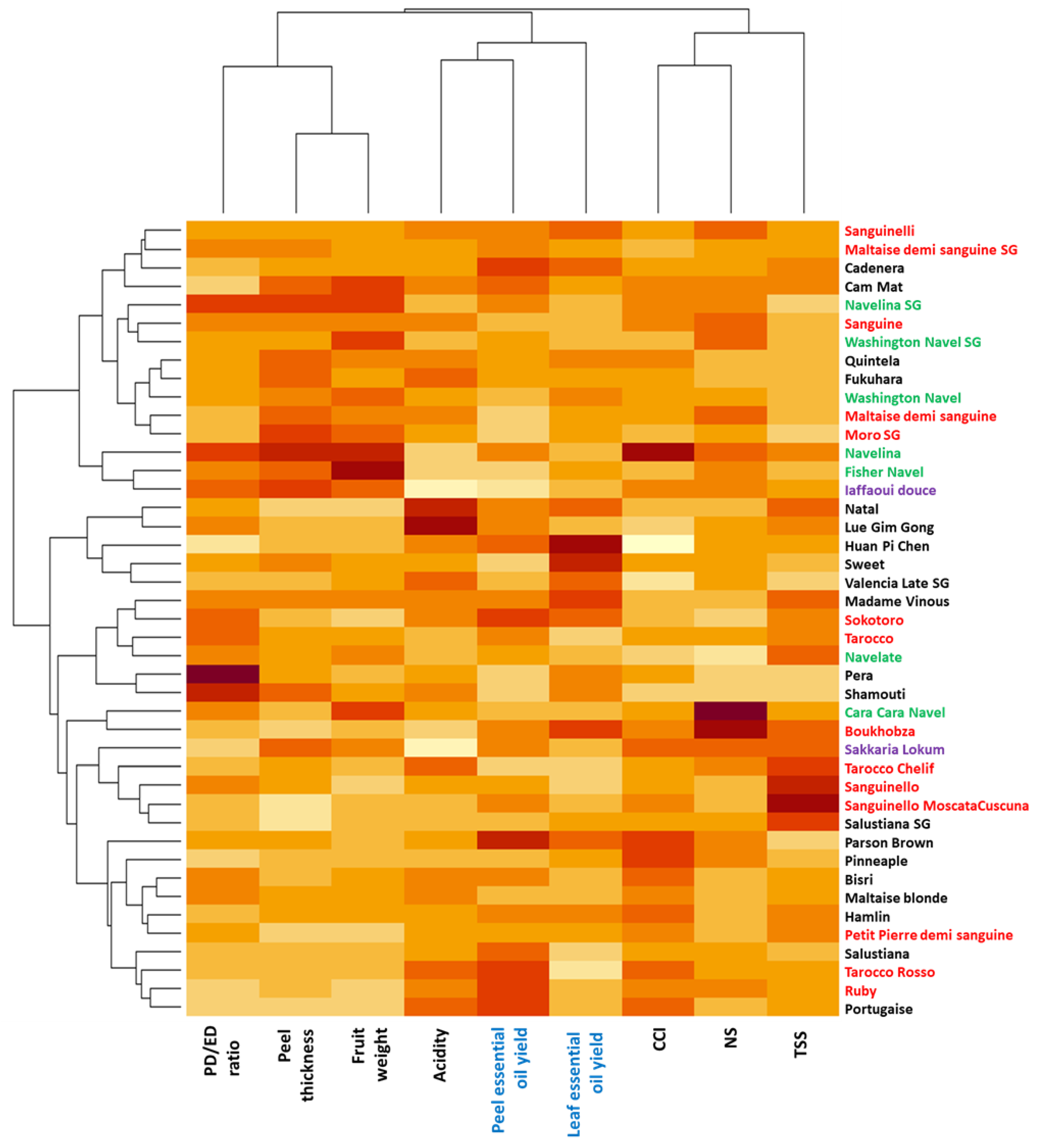
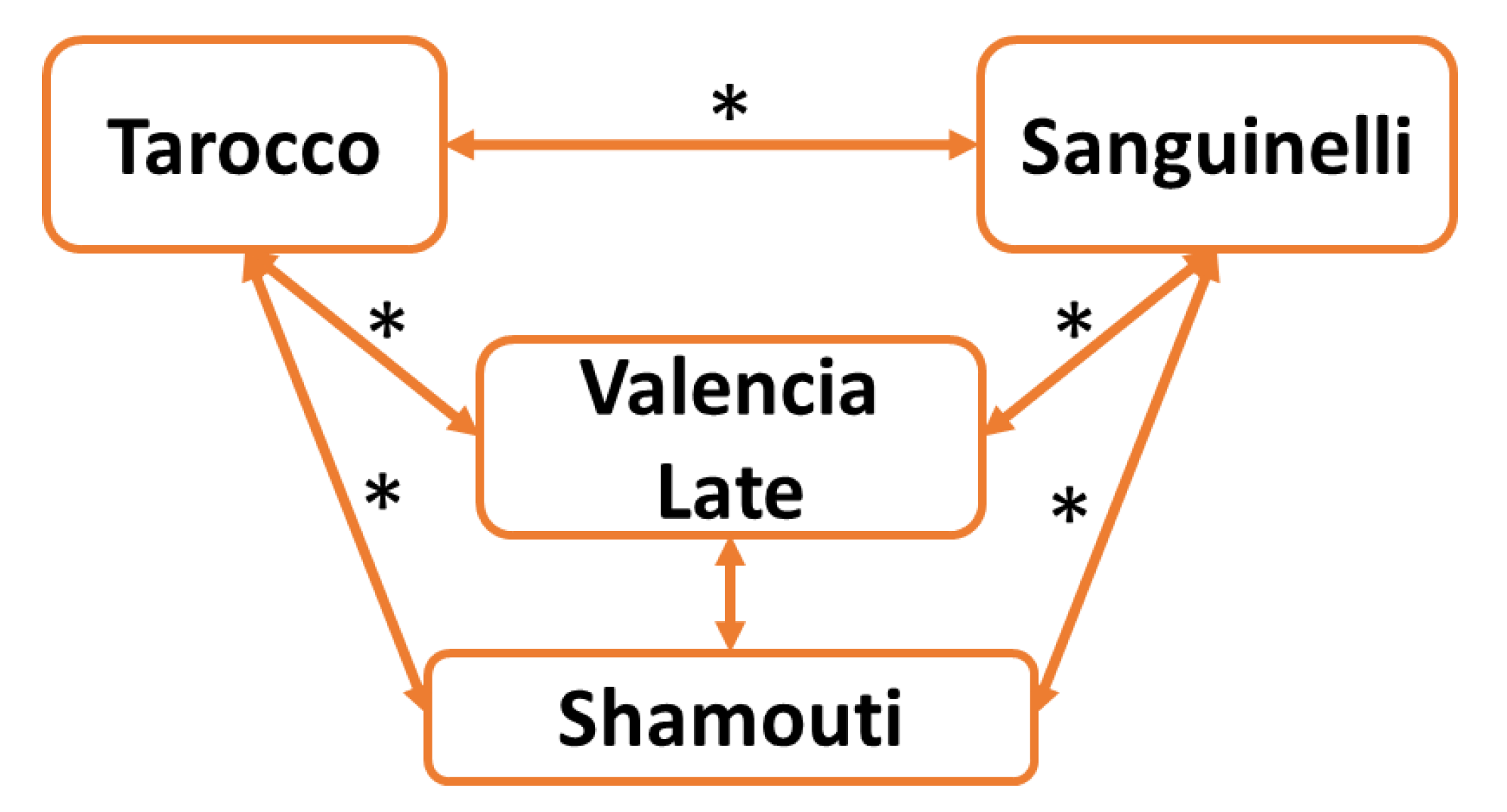
2.1. Genetic Diversity of Oranges
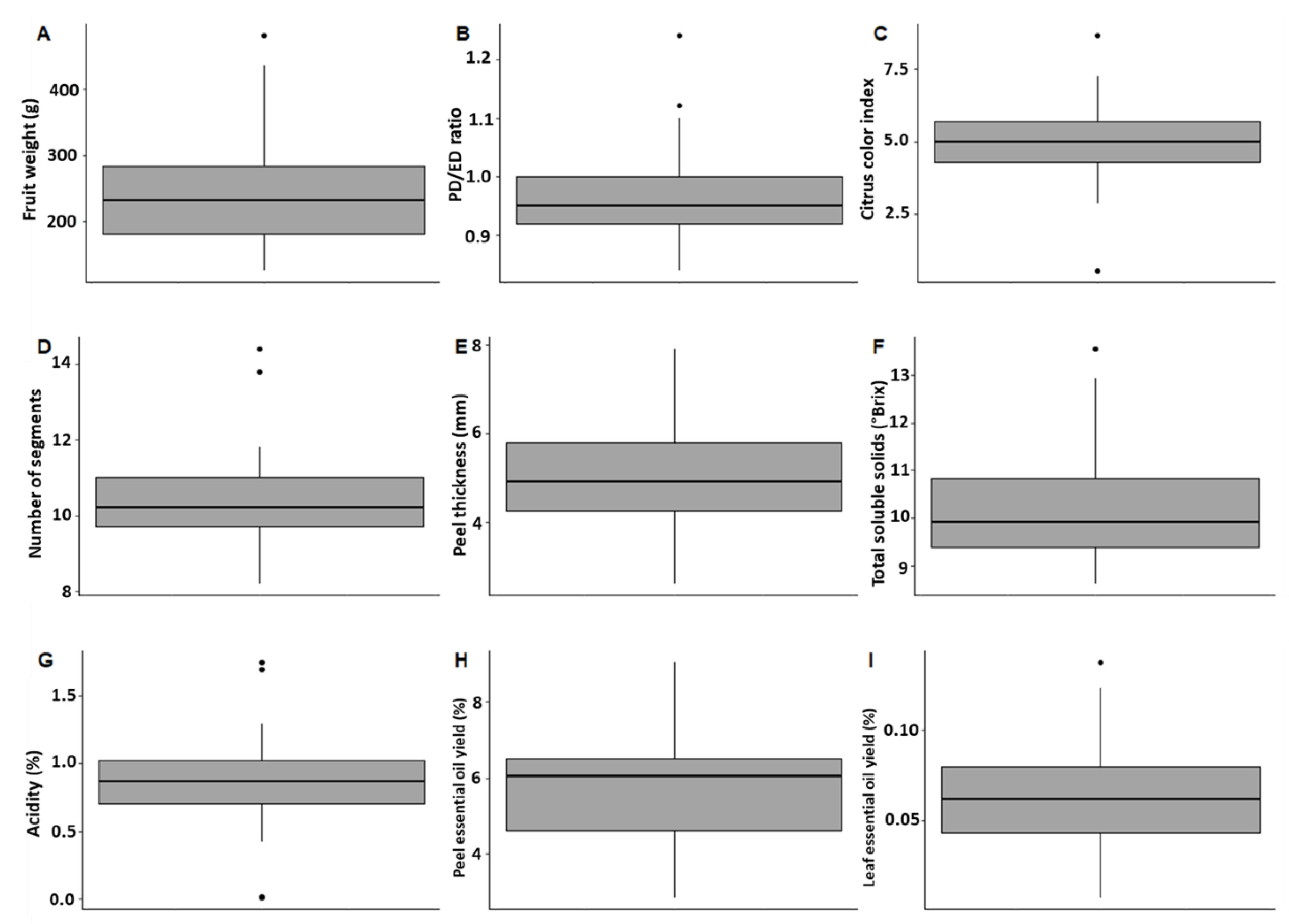
2.2. Pomological Diversity of Oranges
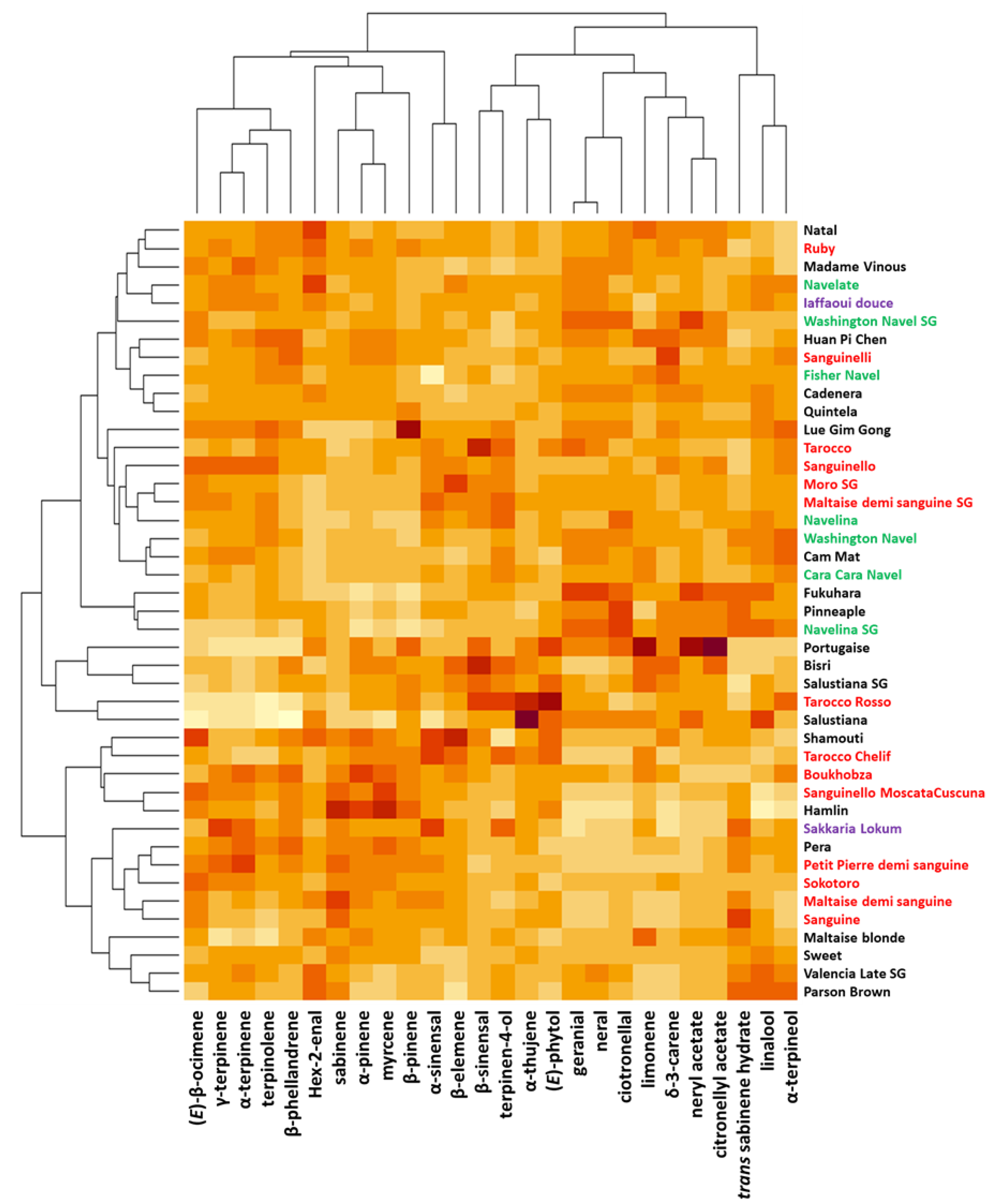
2.3. Diversity of LEO Composition
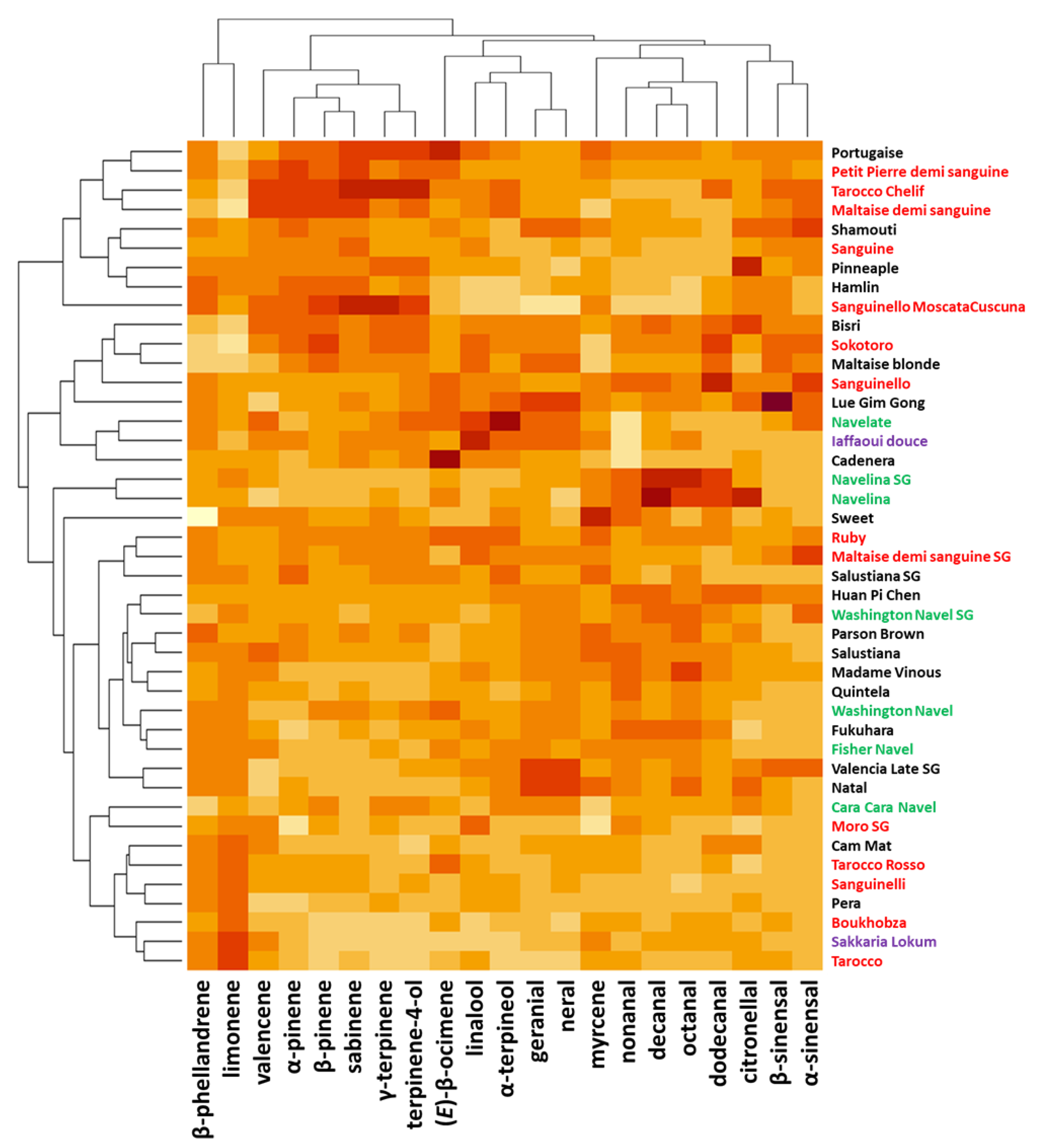
2.4. Compositional Diversity of Peel Oil Diversity of PEO Composition
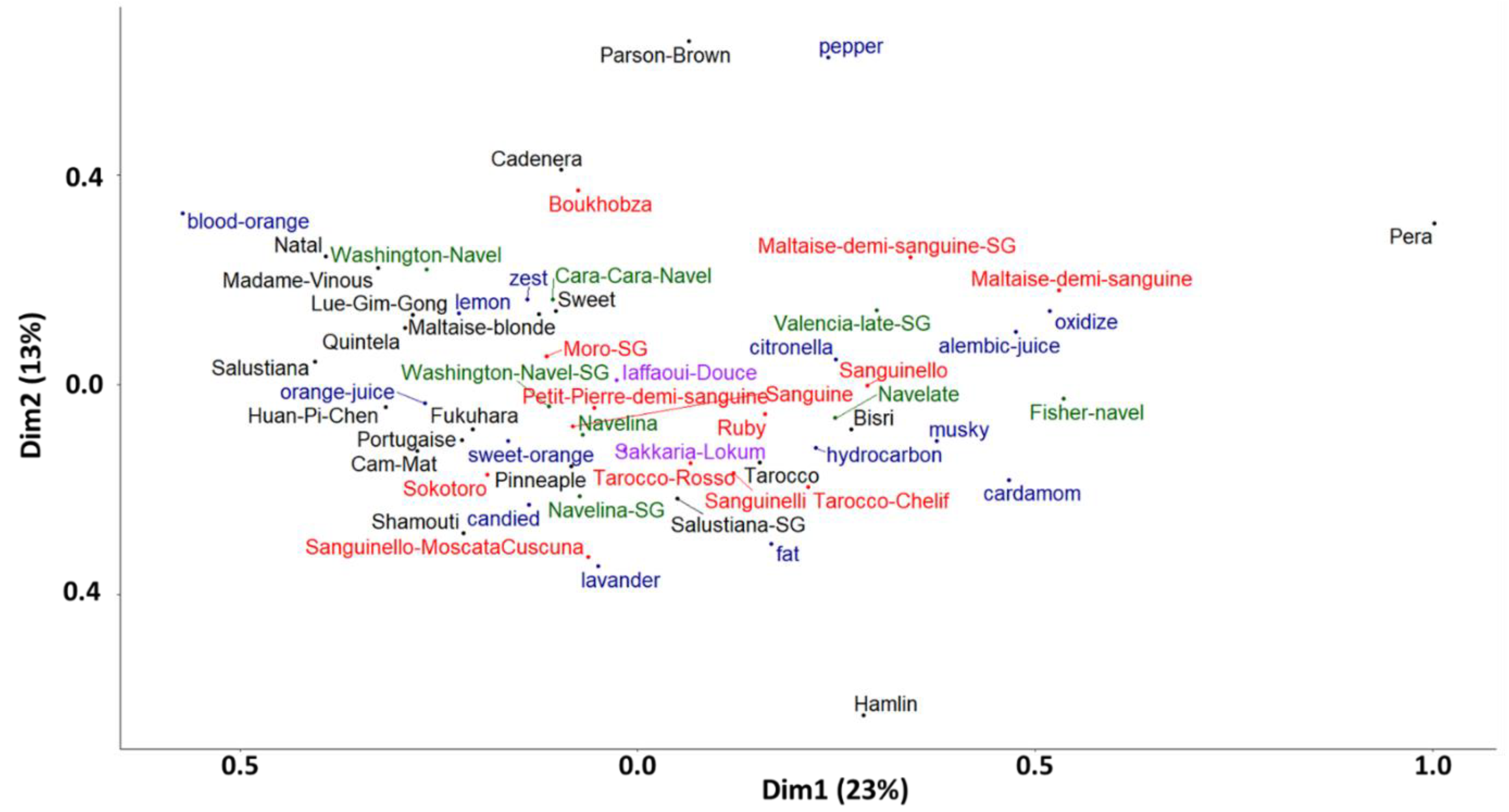
2.5. Aromatic Diversity of the Peel Oil
3. Discussion
4. Materials and Methods
4.1. Biological Materials
4.2. Phenotypic Description
4.2.1. Data Acquisition
4.2.2. Statistical Analysis
4.3. Essential Oil Analysis
4.3.1. Raw Materials
4.3.2. Hydrodistillation
4.3.3. Essential Oil Analysis Using Gas Chromatography and Gas Chromatography-Mass Spectrometry
4.3.4. Statistical Analysis
4.4. Sensorial Analysis of Sweet Orange Peel Oil
4.4.1. Tests, Panelists, and Descriptors
4.4.2. Statistical Analysis
4.5. Genetic Diversity Analysis
4.5.1. DNA Extraction
4.5.2. SSR Genotyping
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Food and Agriculture Organization of the United Nations. FAOSTAT Statistical Database. Available online: https://www.fao.org/ (accessed on 11 May 2022).
- United Stats Department of Agriculture. Citrus World Markets and Trade. Available online: https://www.fas.usda.gov/data/citrus-world-markets-and-trade (accessed on 11 May 2022).
- Zema, D.A.; Calabrò, P.S.; Folino, A.; Tamburino, V.; Zappia, G.; Zimbone, S.M. Valorisation of Citrus Processing Waste: A Review. Waste Manag. 2018, 80, 252–273. [Google Scholar] [CrossRef] [PubMed]
- Teigiserova, D.A.; Tiruta-Barna, L.; Ahmadi, A.; Hamelin, L.; Thomsen, M. A Step Closer to Circular Bioeconomy for Citrus Peel Waste: A Review of Yields and Technologies for Sustainable Management of Essential Oils. J. Environ. Manag. 2021, 280, 111832. [Google Scholar] [CrossRef] [PubMed]
- Peyron, L. Citrus: The Genus Citrus; Dugo, G., Di Giacomo, A., Eds.; Taylor & Francis, CRC Press: London, UK, 2002; Volume 26, pp. 148–152. ISBN 978-0-415-28491-2. [Google Scholar]
- Dugo, G.; Cotroneo, A.; Bonaccorsi, I.; Trozzi, A. Citrus Oils: Composition, Advanced Analytical Techniques, Contaminants, and Biological Activity, 1st ed.; Dugo, G., Modello, L., Eds.; CRC Press: Boca Raton, FL, USA, 2010; pp. 10–20. ISBN 978-1-4398-0029-4. [Google Scholar]
- González-Mas, M.C.; Rambla, J.L.; López-Gresa, M.P.; Blázquez, M.A.; Granell, A. Volatile Compounds in Citrus Essential Oils: A Comprehensive Review. Front. Plant Sci. 2019, 10, 12. [Google Scholar] [CrossRef] [PubMed]
- Sugisawa, H.; Takeda, M.; Yang, R.H.; Takagi, N. The Comparison of Odor Quality of Volatiles in Peel Oils of Five Kinds of Navel Oranges. Nippon. Shokuhin Kogyo 1991, 38, 668–674. [Google Scholar] [CrossRef]
- Gaffney, B.M.; Havekotte, M.; Jacobs, B.; Costa, L.; Ho, C.-T.H. CharmAnalysis of Two Citrus Sinensis Peel Oil Volatiles. Perfum. Flavorist 1996, 21, 1–4. [Google Scholar]
- Högnadóttir, Á.; Rouseff, R.L. Identification of Aroma Active Compounds in Orange Essence Oil Using Gas Chromatography–Olfactometry and Gas Chromatography–Mass Spectrometry. J. Chromatogr. A 2003, 998, 201–211. [Google Scholar] [CrossRef]
- Widder, S.; Eggers, M.; Looft, J.; Vössing, T.; Pickenhagen, W. Handbook of Flavor Characterization: Sensory Analysis, Chemistry, and Physiology; Deibler, K.D., Delwiche, J., Eds.; Food Science and Technology; M. Dekker: New York, NY, USA, 2004; Chapter 14; ISBN 978-0-8247-4703-9. [Google Scholar]
- Qiao, Y.; Xie, B.; Zhang, Y.; Zhang, Y.; Fan, G.; Yao, X.; Pan, S. Characterization of Aroma Active Compounds in Fruit Juice and Peel Oil of Jinchen Sweet Orange Fruit (Citrus sinensis (L.) Osbeck) by GC-MS and GC-O. Molecules 2008, 13, 1333–1344. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Z.; Ma, S.; Niu, Y.; Chen, F.; Yu, D. Characterization of Odour-Active Compounds of Sweet Orange Essential Oils of Different Regions by Gas Chromatography-Mass Spectrometry, Gas Chromatography-Olfactometry and Their Correlation with Sensory Attributes: Characterization of Odor-Active Compounds of Sweet Orange Oils. Flavour Fragr. J. 2016, 31, 41–50. [Google Scholar] [CrossRef]
- Gonçalves, D.; Costa, P.; Rodrigues, C.E.C.; Rodrigues, A.E. Effect of Citrus sinensis Essential Oil Deterpenation on the Aroma Profile of the Phases Obtained by Solvent Extraction. J. Chem. Thermodyn. 2018, 116, 166–175. [Google Scholar] [CrossRef]
- Gaff, M.; Esteban-Decloux, M.; Giampaoli, P. Bitter Orange Peel Essential Oil: A Review of the Different Factors and Chemical Reactions Influencing Its Composition. Flavour Fragr. J. 2020, 35, 247–269. [Google Scholar] [CrossRef]
- Luro, F.; Garcia Neves, C.; Costantino, G.; da Silva Gesteira, A.; Paoli, M.; Ollitrault, P.; Tomi, F.; Micheli, F.; Gibernau, M. Effect of Environmental Conditions on the Yield of Peel and Composition of Essential Oils from Citrus Cultivated in Bahia (Brazil) and Corsica (France). Agronomy 2020, 10, 1256. [Google Scholar] [CrossRef]
- Gioffrè, G.; Ursino, D.; Labate, M.L.C.; Giuffrè, A.M. The Peel Essential Oil Composition of Bergamot Fruit (Citrus bergamia, Risso) of Reggio Calabria (Italy): A Review. Emir. J. Food Agric. 2020, 32, 835–845. [Google Scholar] [CrossRef]
- Rowshan, V.; Najafian, S. Changes of Peel Essential Oil Composition of Citrus aurantium L. During Fruit Maturation in Iran. J. Essent. Oil Bear. Plants 2015, 18, 1006–1012. [Google Scholar] [CrossRef]
- Ferrer, V.; Paymal, N.; Quinton, C.; Tomi, F.; Luro, F. Investigations of the Chemical Composition and Aromatic Properties of Peel Essential Oils throughout the Complete Phase of Fruit Development for Two Cultivars of Sweet Orange (Citrus sinensis (L.) Osb.). Plants 2022, 11, 2747. [Google Scholar] [CrossRef]
- Ferrer, V.; Paymal, N.; Quinton, C.; Costantino, G.; Paoli, M.; Froelicher, Y.; Ollitrault, P.; Tomi, F.; Luro, F. Influence of the Rootstock and the Ploidy Level of the Scion and the Rootstock on Sweet Orange (Citrus sinensis) Peel Essential Oil Yield, Composition and Aromatic Properties. Agriculture 2022, 12, 214. [Google Scholar] [CrossRef]
- Benjamin, G.; Tietel, Z.; Porat, R. Effects of Rootstock/Scion Combinations on the Flavor of Citrus Fruit. J. Agric. Food Chem. 2013, 61, 11286–11294. [Google Scholar] [CrossRef]
- Ferhat, M.A.; Boukhatem, M.N.; Hazzit, M.; Meklati, B.Y.; Chemat, F. Cold Pressing, Hydrodistillation and Microwave Dry Distillation of Citrus Essential Oil from Algeria: A Comparative Study. Electron. J. Biol. 2016, 1, 30–41. [Google Scholar]
- Farahmandfar, R.; Tirgarian, B.; Dehghan, B.; Nemati, A. Changes in Chemical Composition and Biological Activity of Essential Oil from Thomson Navel Orange (Citrus sinensis L. Osbeck) Peel under Freezing, Convective, Vacuum, and Microwave Drying Methods. Food Sci. Nutr. 2020, 8, 124–138. [Google Scholar] [CrossRef] [Green Version]
- Usai, M.; Arras, G.; Fronteddu, F. Effects of Cold Storage on Essential Oils of Peel of Thompson Navel Oranges. J. Agric. Food Chem. 1992, 40, 271–275. [Google Scholar] [CrossRef]
- Zouaghi, G.; Najar, A.; Aydi, A.; Claumann, C.A.; Zibetti, A.W.; Ben Mahmoud, K.; Jemmali, A.; Bleton, J.; Moussa, F.; Abderrabba, M.; et al. Essential Oil Components of Citrus Cultivar ‘Maltaise Demi Sanguine’ (Citrus sinensis) as Affected by the Effects of Rootstocks and Viroid Infection. Int. J. Food Prop. 2019, 22, 438–448. [Google Scholar] [CrossRef] [Green Version]
- Xu, B.M.; Baker, G.L.; Sarnoski, P.J.; Goodrich-Schneider, R.M. A Comparison of the Volatile Components of Cold Pressed Hamlin and Valencia (Citrus sinensis (L.) Osbeck) Orange Oils Affected by Huanglongbing. J. Food Qual. 2017, 2017, 6793986. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Yang, H.; Zhao, W.; Bourcier, E.; Baldwin, E.A.; Plotto, A.; Irey, M.; Bai, J. Huanglongbing and Foliar Spray Programs Affect the Chemical Profile of “Valencia” Orange Peel Oil. Front. Plant Sci. 2021, 12, 611449. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.A.; Prochnik, S.; Jenkins, J.; Salse, J.; Hellsten, U.; Murat, F.; Perrier, X.; Ruiz, M.; Scalabrin, S.; Terol, J.; et al. Sequencing of Diverse Mandarin, Pummelo and Orange Genomes Reveals Complex History of Admixture during Citrus Domestication. Nat. Biotechnol. 2014, 32, 656–662. [Google Scholar] [CrossRef] [PubMed]
- Barkley, N.A.; Roose, M.L.; Krueger, R.R.; Federici, C.T. Assessing Genetic Diversity and Population Structure in a Citrus Germplasm Collection Utilizing Simple Sequence Repeat Markers (SSRs). Theor. Appl. Genet. 2006, 112, 1519–1531. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, T.; Kitajima, A.; Nonaka, K.; Yoshioka, T.; Ohta, S.; Goto, S.; Toyoda, A.; Fujiyama, A.; Mochizuki, T.; Nagasaki, H.; et al. Hybrid Origins of Citrus Varieties Inferred from DNA Marker Analysis of Nuclear and Organelle Genomes. PLoS ONE 2016, 11, e0166969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, G.A.; Terol, J.; Ibanez, V.; López-García, A.; Pérez-Román, E.; Borredá, C.; Domingo, C.; Tadeo, F.R.; Carbonell-Caballero, J.; Alonso, R.; et al. Genomics of the Origin and Evolution of Citrus. Nature 2018, 554, 311–316. [Google Scholar] [CrossRef] [Green Version]
- Arena, E.; Guarrera, N.; Campisi, S.; Nicolosiasmundo, C. Comparison of Odour Active Compounds Detected by Gas-Chromatography–Olfactometry between Hand-Squeezed Juices from Different Orange Varieties. Food Chem. 2006, 98, 59–63. [Google Scholar] [CrossRef]
- Debbabi, O.S.; Bouhlal, R.; Abdelaali, N.; Mnasri, S.; Mars, M. Pomological Study of Sweet Orange (Citrus sinensis L. Osbeck) Cultivars From Tunisia. Int. J. Fruit Sci. 2013, 13, 274–284. [Google Scholar] [CrossRef]
- Dubey, A.K.; Srivastav, M.; Kaur, C. Variation of Sweet Orange (Citrus sinensis L. Osbeck) Accessions in India. Indian J. Hortic. 2014, 71, 321–325. [Google Scholar]
- Caruso, M.; Ferlito, F.; Licciardello, C.; Allegra, M.; Strano, M.C.; Di Silvestro, S.; Russo, M.P.; Pietro Paolo, D.; Caruso, P.; Las Casas, G.; et al. Pomological Diversity of the Italian Blood Orange Germplasm. Sci. Hortic. 2016, 213, 331–339. [Google Scholar] [CrossRef]
- Butelli, E.; Licciardello, C.; Zhang, Y.; Liu, J.; Mackay, S.; Bailey, P.; Reforgiato-Recupero, G.; Martin, C. Retrotransposons Control Fruit-Specific, Cold-Dependent Accumulation of Anthocyanins in Blood Oranges. Plant Cell 2012, 24, 1242–1255. [Google Scholar] [CrossRef] [Green Version]
- Kesterson, J.W.; Braddock, R.J. Total peel oil content of the major Florida citrus cultivars. J. Food Sci. 1975, 40, 931–933. [Google Scholar] [CrossRef]
- Dugo, G.; Verzera, A.; D’Alcontres, I.S.; Cotroneo, A.; Trozzi, A.; Mondello, L. On the Genuineness of Citrus Essential Oils. Part XLIII. The Composition of the Volatile Fraction of Italian Sweet Orange Oils (Citrus sinensis (L.) Osbeck). J. Essent. Oil Res. 1994, 6, 101–137. [Google Scholar] [CrossRef]
- Verzera, A.; Trozzi, A.; D’Alcontres, I.S.; Cotroneo, A. On the Genuineness of Citrus Essential Oils. Part XLVIII. The Composition of Volatile Fraction of Some Varieties of Sweet Orange Oils. J. Essent. Oil Res. 1996, 8, 159–170. [Google Scholar] [CrossRef]
- Trozzi, A.; Verzera, A.; Lamonica, G. Essential Oil Composition of Citrus sinensis (L.) Osbeck Cv. Maltese. J. Essent. Oil Res. 1999, 11, 482–488. [Google Scholar] [CrossRef]
- García-Lor, A.; Luro, F.; Navarro, L.; Ollitrault, P. Comparative Use of InDel and SSR Markers in Deciphering the Interspecific Structure of Cultivated Citrus Genetic Diversity: A Perspective for Genetic Association Studies. Mol. Genet. Genom. 2012, 287, 77–94. [Google Scholar] [CrossRef]
- Rao, M.J.; Zuo, H.; Xu, Q. Genomic Insights into Citrus Domestication and Its Important Agronomic Traits. Plant Commun. 2021, 2, 100138. [Google Scholar] [CrossRef]
- Butelli, E.; Garcia-Lor, A.; Licciardello, C.; Las Casas, G.; Hill, L.; Recupero, G.R.; Keremane, M.L.; Ramadugu, C.; Krueger, R.; Xu, Q.; et al. Changes in Anthocyanin Production during Domestication of Citrus. Plant Physiol. 2017, 173, 2225–2242. [Google Scholar] [CrossRef] [Green Version]
- Butelli, E.; Licciardello, C.; Ramadugu, C.; Durand-Hulak, M.; Celant, A.; Reforgiato Recupero, G.; Froelicher, Y.; Martin, C. Noemi Controls Production of Flavonoid Pigments and Fruit Acidity and Illustrates the Domestication Routes of Modern Citrus Varieties. Curr. Biol. 2019, 29, 158–164.e2. [Google Scholar] [CrossRef] [Green Version]
- Baaliouamer, A.; Meklati, B.Y.; Fraisse, D.; Scharff, C. Analysis of Leaf Oils from Four Varieties of Sweet Orange by Combined Gas Chromatography-Mass Spectrometry. Flavour Fragr. J. 1988, 3, 47–52. [Google Scholar] [CrossRef]
- Fleisher, Z.; Fleisher, A. Sweet Orange Leaf Oil (Citrus sinensis (L.) Osbeck).: Aromatic Plants of the Holy Land and the Sinai. Part I. J. Essent. Oil Res. 1990, 2, 203–205. [Google Scholar] [CrossRef]
- Eldahshan, O.A.; Halim, A.F. Comparison of the Composition and Antimicrobial Activities of the Essential Oils of Green Branches and Leaves of Egyptian Navel Orange (Citrus sinensis (L.) Osbeck Var. Malesy). Chem. Biodivers. 2016, 13, 681–685. [Google Scholar] [CrossRef]
- Khalid, K.A.; Ahmed, A.M.A.; El-Gohary, A.E. Effect of Growing Seasons on the Leaf Essential Oil Composition of Citrus Species That Are Cultivated in Egypt. J. Essent. Oil Res. 2020, 32, 296–307. [Google Scholar] [CrossRef]
- Näf, R.; Velluz, A.; Meyer, A.P. Volatile Constituents of Blood and Blond Orange Juices: A Comparison. J. Essent. Oil Res. 1996, 8, 587–595. [Google Scholar] [CrossRef]
- Perez-Cacho, P.R.; Rouseff, R.L. Fresh Squeezed Orange Juice Odor: A Review. Crit. Rev. Food Sci. Nutr. 2008, 48, 681–695. [Google Scholar] [CrossRef] [PubMed]
- Porat, R.; Deterre, S.; Giampaoli, P.; Plotto, A. Biotechnology in Flavor Production, 2nd ed.; WILEY/Blackwell: Chichester, UK, 2016; Chapter 1; ISBN 978-1-118-35406-3. [Google Scholar]
- Rodríguez, A.; Peris, J.E.; Redondo, A.; Shimada, T.; Costell, E.; Carbonell, I.; Rojas, C.; Peña, L. Impact of D-Limonene Synthase up- or down-Regulation on Sweet Orange Fruit and Juice Odor Perception. Food Chem. 2017, 217, 139–150. [Google Scholar] [CrossRef]
- Luro, F.; Bloquel, E.; Tomu, B.; Costantino, G.; Tur, I.; Riolacci, S.; Varamo, F.; Ollitrault, P.; Froelicher, Y.; Curk, F.; et al. The Inracirad Citrus Germplasm Collection of San Giuliano, Corsica; Publications du centre Jean Bérard: Naples, Italy, 2017; pp. 243–261. [Google Scholar]
- Jimenez-Cuesta, M.; Cuquerella, J.; Martinez-Javaga, J.M. Determination of a color index for citrus fruit de-greening. In Proceedings of the International Society of Citriculture/International Citrus Congres, Tokyo, Japan, 9–12 November 1982. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis, 2nd ed.; Springer International Publishing: New York, NY, USA, 2016; pp. 3–253. ISBN 978-3-319-24275-0. [Google Scholar]
- Gomez, K.A.; Gomez, A.A. Statistical Procedures for Agricultural Research, 2nd ed.; John Wiley & Sons: New York, NY, USA, 1984; pp. 3–700. ISBN 978-0-471-87092-0. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 11 May 2022).
- Luro, F.; Viglietti, G.; Marchi, E.; Costantino, G.; Scarpa, G.M.; Tomi, F.; Paoli, M.; Curk, F.; Ollitrault, P. Genetic, Morphological and Chemical Investigations Reveal the Genetic Origin of Pompia (C. Medica Tuberosa Risso & Poiteau)—An Old Endemic Sardinian Citrus Fruit. Phytochemistry 2019, 168, 112083. [Google Scholar] [CrossRef] [Green Version]
- Christensen, R.H.B.; Brockhoff, P.B.; Kuznetsova, A.; Birot, S.; Stachlewska, K.A. Thurstonian Models for Sensory Discrimination. R Package Version 1.5-2. Available online: https://github.com/perbrock/sensR (accessed on 22 April 2022).
- Hervé, M. RVAideMemoire: Testing and Plotting Procedures for Biostatistics. R Package Version 0.9-79. 2020. Available online: https://CRAN.R-project.org/package=RVAideMemoire (accessed on 21 April 2022).
- Luro, F.L.; Costantino, G.; Terol, J.; Argout, X.; Allario, T.; Wincker, P.; Talon, M.; Ollitrault, P.; Morillon, R. Transferability of the EST-SSRs Developed on Nules Clementine (Citrus clementina Hort Ex Tan) to Other Citrus Species and Their Effectiveness for Genetic Mapping. BMC Genom. 2008, 9, 287. [Google Scholar] [CrossRef]
- Ferrer, V.; Costantino, G.; Paoli, M.; Paymal, N.; Quinton, C.; Ollitrault, P.; Tomi, F.; Luro, F. Intercultivar Diversity of Sour Orange (Citrus aurantium L.) Based on Genetic Markers, Phenotypic Characteristics, Aromatic Compounds and Sensorial Analysis. Agronomy 2021, 11, 1084. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivars | Blond | Blood | Navel |
|---|---|---|---|
| Monoterpene aldehydes | 5.51 ± 2.48 ab | 3.72 ± 1.49 b | 7.35 ± 1.94 a |
| Monoterpene alcohols | 20.94 ± 4.72 a | 20.43 ± 4.52 a | 23.35 ± 3.49 a |
| Monoterpene esters | 0.69 ± 0.45 a | 0.42 ± 0.18 a | 0.76 ± 0.30 a |
| Oxygenated sesquiterpenes | 2.60 ± 0.76 a | 2.84 ± 0.87 a | 2.43 ± 0.40 a |
| Aliphatic aldehydes | 0.75 ± 0.22 a | 0.57 ± 0.14 a | 0.64 ± 0.30 a |
| Cultivars | Blond | Blood | Navel |
|---|---|---|---|
| Monoterpene aldehydes | 0.15 ± 0.05 a | 0.10 ± 0.04 b | 0.15 ± 0.03 ab |
| Monoterpene alcohols | 0.92 ± 0.30 a | 0.96 ± 0.32 a | 0.77 ± 0.37 a |
| Oxygenated sesquiterpenes | 0.03 ± 0.04 a | 0.04 ± 0.03 a | 0.01 ± 0.01 a |
| Aliphatic aldehydes | 0.54 ± 0.16 ab | 0.41 ± 0.15 b | 0.70 ± 0.30 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferrer, V.; Paymal, N.; Costantino, G.; Paoli, M.; Quinton, C.; Tomi, F.; Luro, F. Correspondence between the Compositional and Aromatic Diversity of Leaf and Fruit Essential Oils and the Pomological Diversity of 43 Sweet Oranges (Citrus x aurantium var sinensis L.). Plants 2023, 12, 990. https://doi.org/10.3390/plants12050990
Ferrer V, Paymal N, Costantino G, Paoli M, Quinton C, Tomi F, Luro F. Correspondence between the Compositional and Aromatic Diversity of Leaf and Fruit Essential Oils and the Pomological Diversity of 43 Sweet Oranges (Citrus x aurantium var sinensis L.). Plants. 2023; 12(5):990. https://doi.org/10.3390/plants12050990
Chicago/Turabian StyleFerrer, Vincent, Noémie Paymal, Gilles Costantino, Mathieu Paoli, Carole Quinton, Félix Tomi, and François Luro. 2023. "Correspondence between the Compositional and Aromatic Diversity of Leaf and Fruit Essential Oils and the Pomological Diversity of 43 Sweet Oranges (Citrus x aurantium var sinensis L.)" Plants 12, no. 5: 990. https://doi.org/10.3390/plants12050990
APA StyleFerrer, V., Paymal, N., Costantino, G., Paoli, M., Quinton, C., Tomi, F., & Luro, F. (2023). Correspondence between the Compositional and Aromatic Diversity of Leaf and Fruit Essential Oils and the Pomological Diversity of 43 Sweet Oranges (Citrus x aurantium var sinensis L.). Plants, 12(5), 990. https://doi.org/10.3390/plants12050990