
Avenanthramides, Distinctive Hydroxycinnamoyl Conjugates of Oat, Avena sativa L.: An Update on the Biosynthesis, Chemistry, and Bioactivities


Abstract
:1. Introduction
2. Avenanthramides as Distinctive Oat Phytochemicals
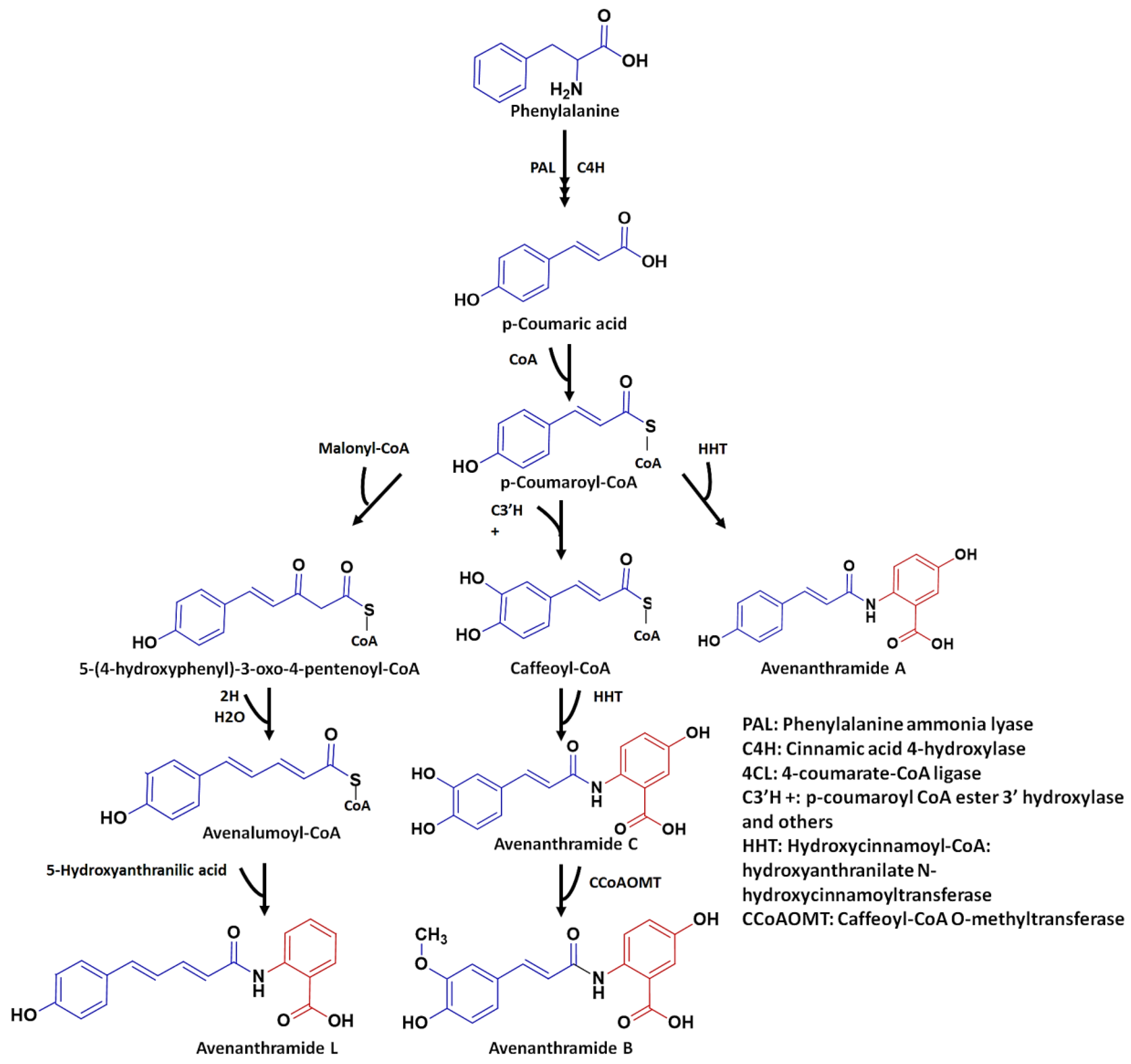
3. Biosynthesis of Avenanthramides
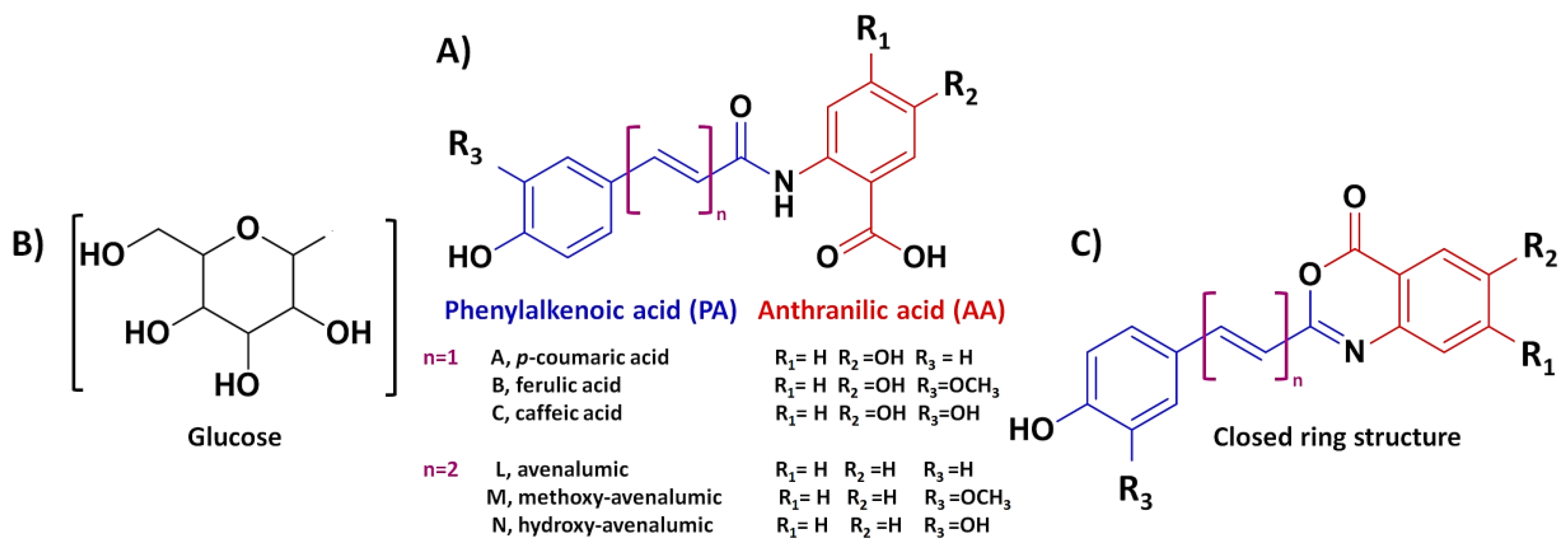
4. Diversity of Avenanthramide Structures and Associated Nomenclature
5. Avenanthramide Extraction and Detection
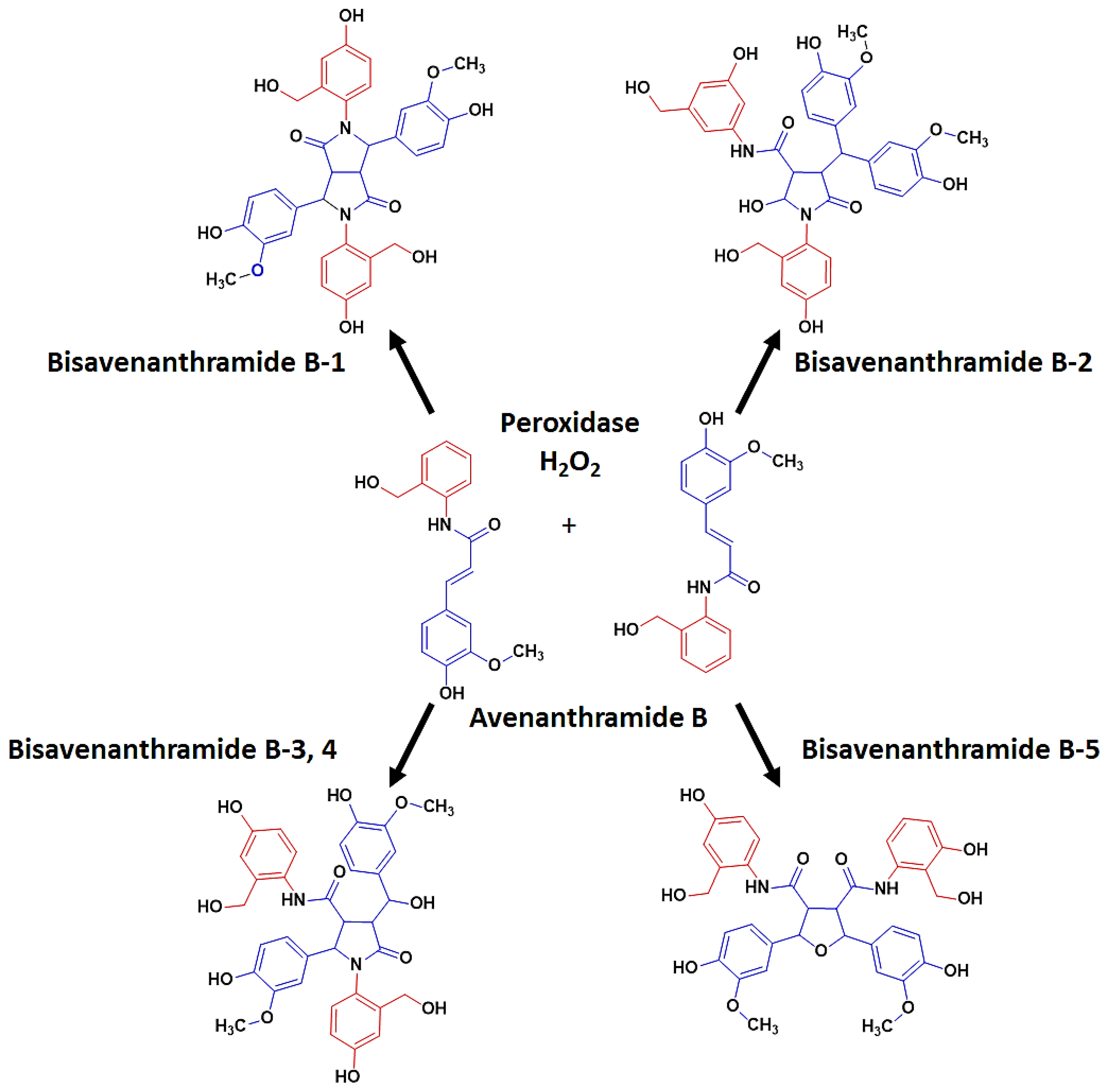
6. Post-Synthesis Alterations: Isomerisation and Dimerisation
7. In Planta Bioactivities of Avenanthramides
8. Health-Beneficial Properties of Avenanthramides
9. Biotechnological Approaches for Avenanthramide Synthesis
10. Concluding Remarks and Future Perspectives
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Méteignier, L.-V.; Nützmann, H.-W.; Papon, N.; Osbourn, A.; Courdavault, V. Emerging mechanistic insights into the regulation of specialized metabolism in plants. Nat. Plants 2023, 9, 22–30. [Google Scholar] [CrossRef] [PubMed]
- D’Auria, J.C. Acyltransferases in plants: A good time to be BAHD. Curr. Opin. Plant Biol. 2006, 9, 331–340. [Google Scholar] [CrossRef]
- Petersen, M. Hydroxycinnamoyltransferases in plant metabolism. Phytochem. Rev. 2016, 15, 699–727. [Google Scholar] [CrossRef]
- Zeiss, D.R.; Piater, L.A.; Dubery, I.A. Hydroxycinnamate amides: An intriguing combination of plant protective metabolites. Trends Plant Sci. 2021, 26, 184. [Google Scholar] [CrossRef] [PubMed]
- Bontpart, T.; Cheynier, V.; Ageorges, A.; Terrier, N. BAHD or SCPL acyltransferase? What a dilemma for acylation in the world of plant phenolic compounds. New Phytol. 2015, 208, 695–707. [Google Scholar] [CrossRef]
- Boz, H. Phenolic amides (avenanthramides) in oats—A review. Czech J. Food Sci. 2015, 33, 399–404. [Google Scholar] [CrossRef] [Green Version]
- Sang, S.; Chu, Y. Whole grain oats, more than just a fiber: Role of unique phytochemicals. Molec. Nutr. Food Res. 2017, 61, 1600715. [Google Scholar] [CrossRef]
- Leonova, A.; Gnutikov, I.; Loskutov, E.; Blinova, K.-E.; Gustafsson, O.; Olsson, O. Diversity of avenanthramide content in wild and cultivated oats. Proc. Appl. Bot. Genet. Breed. 2020, 181, 30–47. [Google Scholar] [CrossRef]
- Matilla, P.; Pihlava, J.-M.; Hellstrom, J. Contents of phenolic acids, alkyl- and alkenylresorcinols, and avenanthramides in commercial grain products. J. Agric. Food Chem. 2005, 53, 8290–8295. [Google Scholar] [CrossRef]
- Perrelli, A.; Goitre, l.; Salzano, A.M.; Moglia, A.; Scaloni, A.; Retta, S.F. Biological activities, health benefits, and therapeutic properties of avenanthramides: From skin protection to prevention and treatment of cerebrovascular diseases. Oxid. Med. Cell. Longev. 2018, 2018, 6015351. [Google Scholar] [CrossRef]
- Dvořáček, V.; Jágr, M.; Kotrbová Kozak, A.; Capouchová, I.; Konvalina, P.; Faměra, O.; Hlásná Čepková, P. Avenanthramides: Unique bioactive substances of oat grain in the context of cultivar, cropping system, weather conditions and other grain parameters. Plants 2021, 10, 2485. [Google Scholar] [CrossRef] [PubMed]
- Collins, F.; Mullin, W. High-performance liquid chromatographic determination of avenanthramides, n-aroylanthranilic acid alkaloids from oats. J. Chrom. A 1988, 445, 363–370. [Google Scholar] [CrossRef]
- Woolman, M.; Liu, K. Simplified analysis and expanded profiles of avenanthramides in oat grains. Foods 2022, 16, 560. [Google Scholar] [CrossRef] [PubMed]
- Wise, M.L. Avenanthramides: Chemistry and biosynthesis. In Oats Nutrition and Technology; Chu, Y.F., Ed.; John Wiley: New York, NY, USA, 2013; pp. 195–226. [Google Scholar]
- Matsukawa, T.; Isobe, T.; Ishihara, A.; Iwamura, H. Occurrence of avenanthramides and hydroxycinnamoyl-CoA:hydroxyanthranilate N-hydroxycinnamoyltransferase activity in oat seeds. Z. Naturforsch. 2000, 55, 30–36. [Google Scholar] [CrossRef]
- de Bruijn, W.J.; van Dinteren, S.; Gruppen, H.; Vincken, J.P. Mass spectrometric characterisation of avenanthramides and enhancing their production by germination of oat (Avena sativa). Food Chem. 2019, 277, 682–690. [Google Scholar] [CrossRef]
- Wise, M.L.; Sreenath, H.K.; Skadsen, R.W.; Kaeppler, H.F. Biosynthesis of avenanthramides in suspension cultures of oat (Avena sativa). Plant Cell Tissue Organ Cult. 2009, 97, 81–90. [Google Scholar] [CrossRef]
- Yuan, Z.; Yang, H.; Pan, L.; Zhao, W.; Liang, L.; Gatera, A.; Tucker, M.R.; Xu, D. Systematic identification and expression profiles of the BAHD superfamily acyltransferases in barley (Hordeum vulgare). Sci. Rep. 2022, 12, 5063. [Google Scholar] [CrossRef]
- Yang, Q.; Reinhard, K.; Schiltz, E.; Matern, U. Characterization and heterologous expression of hydroxycinnamoyl/benzoyl-CoA:anthranilate N-hydroxycinnamoyl/benzoyltransferase from elicited cell cultures of carnation, Dianthus caryophyllus L. Plant Molec. Biol. 1997, 35, 777–789. [Google Scholar] [CrossRef]
- Kruse, L.H.; Weigle, A.T.; Irfan, M.; Martínez-Gómez, J.; Chobirko, J.D.; Schaffer, J.E.; Bennett, A.A.; Specht, C.D.; Jez, J.M.; Shukla, D.; et al. Multiple routes of functional diversification of the plant BAHD acyltransferase family revealed by comparative biochemical and genomic analyses. bioRxiv 2021. [Google Scholar] [CrossRef]
- Weng, J.-K.; Lynch, J.H.; Matos, J.O.; Dudareva, N. Adaptive mechanisms of plant specialized metabolism connecting chemistry to function. Nat. Chem. Biol. 2021, 17, 1037–1045. [Google Scholar] [CrossRef]
- Yang, Q.; Trinh, H.X.; Imai, S.; Ishihara, A.; Zhang, L.; Nakayashiki, H.; Tosa, Y.; Mayama, S. Analysis of the involvement of hydroxyanthranilate hydroxycinnamoyltransferase and caffeoyl-CoA 3-O-methyltransferase in phytoalexin biosynthesis in oat. Molec. Plant Micr. Interact. 2004, 17, 81–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.J.; Sim, G.Y.; Kang, H.; Yeo, W.S.; Kim, B.G.; Ahn, J.H. Synthesis of avenanthramides using engineered Escherichia coli. Micr. Cell Factories 2018, 17, 46. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Chen, Y.; Meesapyodsuk, D.; Qiu, X. The biosynthetic pathway of major avenanthramides in oat. Metabolites 2019, 9, 163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishihara, A.; Ohtsu, Y.; Iwamura, H. Induction of biosynthetic enzymes for avenanthramides in elicitor-treated oat leaves. Planta 1999, 208, 519–527. [Google Scholar] [CrossRef]
- Collins, F.W.; McLachlan, D.C.; Blackwell, B.A. Oat phenolics: Avenalumic acid, a new group of bound phenolic acids from oat groats and hulls. Cereal Chem. 1991, 68, 184–189. [Google Scholar]
- Kamal, N.; Tsardakas Renhuldt, N.; Bentzer, J.; Gundlach, H.; Haberer, G.; Juhász, A.; Lux, T.; Bose, U.; Tye-Din, J.A.; Lang, D.; et al. The mosaic oat genome gives insights into a uniquely healthy cereal crop. Nature 2022, 606, 113. [Google Scholar] [CrossRef]
- Yang, Z.; Wang, K.; Aziz, U.; Zhao, C.; Zhang, M. Evaluation of duplicated reference genes for quantitative real-time PCR analysis in genome unknown hexaploid oat (Avena sativa L.). Plant Meth. 2020, 16, 138. [Google Scholar] [CrossRef]
- Ishihara, A.; Kojima, K.; Fujita, T.; Yamamoto, Y.; Nakajima, H. New series of avenanthramides in oat seed. Biosci. Biotech. Biochem. 2014, 78, 1975–1983. [Google Scholar] [CrossRef]
- Wu, W.; Tang, Y.; Yang, J.; Idehen, E.; Sang, S. Avenanthramide aglycones and glucosides in oat bran: Chemical profile, levels in commercial oat products, and cytotoxicity to human colon cancer cells. J. Agric. Food Chem. 2018, 66, 8005–8014. [Google Scholar] [CrossRef]
- Mayama, S.; Matsuura, H.; Iida, H.; Tani, T. The role of avenalumin in the resistance of oat to crown rust, Puccinia coronata f. sp. avenae. Physiol. Plant Pathol. 1982, 20, 189–199. [Google Scholar] [CrossRef]
- Collins, F.W. Oat phenolics: Avenanthramides, novel substituted N-cinnamoylanthranilate alkaloids from oat groats and hulls. J. Agric. Food Chem. 1989, 37, 60–66. [Google Scholar] [CrossRef]
- Dimberg, L.H.; Theander, O.; Lingnert, H. Avenanthramides—A group of phenolic antioxidants in oats. Cereal Chem. 1993, 70, 637. [Google Scholar]
- Dimberg, L.H.; Sunnerheim, K.; Sundberg, B.; Walsh, K. Stability of oat avenanthramides. Cereal Chem. 2001, 78, 278–281. [Google Scholar] [CrossRef]
- Jágr, M.; Dvořáček, V.; Hlásná Čepková, P.; Doležalová, J. Comprehensive analysis of oat avenanthramides using hybrid quadrupole-Orbitrap mass spectrometry: Possible detection of new compounds. Rapid Commun. Mass Spectrom. 2020, 34, e8718. [Google Scholar] [CrossRef] [PubMed]
- Maliarova, M.; Mrazova, V.; Havrlentova, M.; Sokol, J. Optimization of parameters for extraction of avenanthramides from oat (Avena sativa L.) grain using response surface methodology (RSM). J. Brazilian Chem. Soc. 2015, 26, 2369–2378. [Google Scholar]
- Fontes-Candia, C.; Ramos-Sanchez, V.; Chavez-Flores, D.; Salmeron, I.; Perez-Vega, S. Extraction of different phenolic groups from oats at a nonthermal pilot scale: Effect of solvent composition and cycles. J. Food Process Eng. 2018, 41, e12651. [Google Scholar] [CrossRef]
- Walters, M.; Ribeiro, A.P.L.; Hosseinian, F.; Tsopmo, A. Phenolic acids, avenanthramides, and antioxidant activity of oats defatted with hexane or supercritical fluid. J. Cereal Sci. 2018, 79, 21–26. [Google Scholar] [CrossRef]
- Xochitl, A.F.; Rosalía, R.C.; Minerva, R.G.; Mendoza-Sánchez, M.; Mora, O.; Pérez-Ramírez, I.F. Polyphenols and avenanthramides extracted from oat (Avena sativa L.) grains and sprouts modulate genes involved in glucose and lipid metabolisms in 3T3 L1 adipocytes. J. Food Biochem. 2021, 45, e13738. [Google Scholar] [CrossRef]
- Feng, Y.; Suo, D.; Guan, X.; Wang, S.; Xiao, Z.; Li, Y.; Liu, X.; Fan, X. Effect of germination on the avenanthramide content of oats and their in vitro antisensitivity activities. Molecules 2022, 27, 6167. [Google Scholar] [CrossRef]
- Taofiq, O.; González-Paramás, A.M.; Barreiro, M.F.; Ferreira, I.C. Hydroxycinnamic acids and their derivatives: Cosmeceutical significance, challenges and future perspectives, a review. Molecules 2017, 22, 281. [Google Scholar] [CrossRef]
- Ube, N.; Yabuta, Y.; Tohnooka, T.; Ueno, K.; Taketa, S.; Ishihara, A. Biosynthesis of phenylamide phytoalexins in pathogen-infected barley. Int. J. Molec. Sci. 2019, 20, 5541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishihara, A. Defense mechanisms involving secondary metabolism in the grass family. J. Pest. Sci. 2021, 46, 382–392. [Google Scholar] [CrossRef] [PubMed]
- Okazaki, Y.; Ishizuka, A.; Ishihara, A.; Nishioka, T.; Iwamura, H. New dimeric compounds of avenanthramide phytoalexin in oats. J. Org. Chem. 2007, 72, 3830–3839. [Google Scholar] [CrossRef] [PubMed]
- Okazaki, Y.; Isobe, T.; Iwata, Y.; Matsukawa, T.; Matsuda, F.; Miyagawa, H.; Ishihara, A.; Nishioka, T.; Iwamura, H. Metabolism of avenanthramide phytoalexins in oats. Plant Cell 2004, 39, 560–572. [Google Scholar] [CrossRef] [PubMed]
- Schendel, R.R. Phenol content in sprouted grains. In Sprouted Grains: Nutritional Value, Production, and Applications; Feng, H., Nemzer, B., Devries, J.W., Eds.; Woodhead Publishing, Elsevier: Cambridge, MA, USA, 2019; pp. 248–315. [Google Scholar] [CrossRef]
- Wise, M.L. Effect of chemical systemic acquired resistance elicitors on avenanthramide biosynthesis in oat (Avena sativa). J. Agric. Food Chem. 2011, 59, 7028–7038. [Google Scholar] [CrossRef]
- Shakeel, Q.; Raheel, M.; Bajwa, R.T.; Rashid, I.; Raza, H.Y.; Saleem, S.R. Etiology and Management of Economically Significant Diseases of Avena sativa. In Sustainable Winter Fodder; CRC Press: Boca Raton, FL, USA, 2021; pp. 131–163. [Google Scholar] [CrossRef]
- Desmedt, W.; Mangelinckx, S.; Kyndt, T.; Vanholme, B. A phytochemical perspective on plant defense against nematodes. Front. Plant Sci. 2020, 11, 602079. [Google Scholar] [CrossRef]
- Pretorius, C.J.; Tugizimana, F.; Steenkamp, P.A.; Piater, L.A.; Dubery, I.A. Metabolomics for biomarker discovery: Key signatory metabolic profiles for the identification and discrimination of oat cultivars. Metabolites 2021, 11, 165. [Google Scholar] [CrossRef]
- Pretorius, C.J.; Tugizimana, F.; Steenkamp, P.A.; Piater, L.A.; Dubery, I.A. Metabolomic characterisation of discriminatory metabolites involved in halo blight disease in oat cultivars caused by Pseudomonas syringae pv. coronafaciens. Metabolites 2022, 12, 248. [Google Scholar] [CrossRef]
- Ramaroson, M.-L.; Koutouan, C.; Helesbeux, J.-J.; Le Clerc, V.; Hamama, L.; Geoffriau, E.; Briard, M. Role of phenylpropanoids and flavonoids in plant resistance to pests and diseases. Molecules 2022, 27, 8371. [Google Scholar] [CrossRef]
- Shahidi, F.; Chandrasekara, A. Hydroxycinnamates and their in vitro and in vivo antioxidant activities. Phytochem. Rev. 2010, 9, 147–170. [Google Scholar] [CrossRef]
- Xue, Y.; Teng, Y.; Chen, M.; Li, Z.; Wang, G. Antioxidant activity and mechanism of avenanthramides: Double H+/e− processes and role of the catechol, guaiacyl, and carboxyl groups. J. Agric. Food Chem. 2021, 69, 7178–7189. [Google Scholar] [CrossRef] [PubMed]
- Spencer, J.; Soycan, G.; Corona, G.; Johnson, J.; Chu, Y.; Shewry, P.; Lovegrove, A. Chronic vascular effects of oat phenolic acids and avenanthramides in pre-or stage 1 hypertensive adults. Curr. Dev. Nutr. 2020, 4, 478. [Google Scholar] [CrossRef]
- Zhouyao, H.; Malunga, L.N.; Chu, Y.F.; Eck, P.; Ames, N.; Thandapilly, S.J. The inhibition of intestinal glucose absorption by oat-derived avenanthramides. J. Food Biochem. 2022, 46, e14324. [Google Scholar] [CrossRef]
- Thies, F.; Masson, L.F.; Boffetta, P.; Kris-Etherton, P. Oats and bowel disease: A systematic literature review. Br. J. Nutr. 2014, 112 (Suppl. S2), S31–S43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turrini, E.; Maffei, F.; Milelli, A.; Calcabrini, C.; Fimognari, C. Overview of the anticancer profile of avenanthramides from oat. Int. J. Molec. Sci. 2019, 20, 4536. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Ni, T.; Zhang, D.; Liu, H.; Wang, J.; Sun, B. Consumption of avenanthramides extracted from oats reduces weight gain, oxidative stress, inflammation and regulates intestinal microflora in high fat diet-induced mice. J. Funct. Foods 2020, 65, 103774. [Google Scholar] [CrossRef]
- Gilissen, L.J.; Van der Meer, I.M.; Smulders, M.J. Why oats are safe and healthy for celiac disease patients. Med. Sci. 2016, 4, 21. [Google Scholar] [CrossRef] [Green Version]
- Sur, R.; Nigam, A.; Grote, D.; Liebel, F.; Southall, M.D. Avenanthramides, polyphenols from oats, exhibit anti-inflammatory and anti-itch activity. Arch. Derm. Res. 2008, 300, 569–574. [Google Scholar] [CrossRef]
- Darakhshan, S.; Pour, A.B. Tranilast: A review of its therapeutic applications. Pharm. Res. 2015, 91, 15–28. [Google Scholar] [CrossRef]
- Lopatin, U. Drugs in the pipeline for HBV. Clin. Liver Dis. 2019, 23, 535–555. [Google Scholar] [CrossRef]
- Prasher, P.; Sharma, M. Medicinal chemistry of anthranilic acid derivatives: A mini review. Drug Dev. Res. 2021, 82, 945–958. [Google Scholar] [CrossRef] [PubMed]
- Nomura, T.; Ogita, S.; Kato, Y. Rational metabolic-flow switching for the production of exogenous secondary metabolites in bamboo suspension cells. Sci. Rep. 2018, 8, 13203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eudes, A.; Juminaga, D.; Baidoo, E.E.; Collins, F.W.; Keasling, J.D.; Loqué, D. Production of hydroxycinnamoyl anthranilates from glucose in Escherichia coli. Micr. Cell Factories 2013, 12, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Collins [32] Nomenclature | Dimberg [33] Nomenclature | n | R1 | R2 | R3 |
|---|---|---|---|---|---|
| A | 2p | 1 | H | OH | H |
| B | 2f | 1 | H | OH | OCH3 |
| C * | 2c | 1 | H | OH | OH |
| L | 1pd | 2 | H | H | H |
| D | 1p | 1 | H | H | H |
| F | 1c | 1 | H | H | OH |
| E | 1f | 1 | H | H | OCH3 |
| N | 1cd | 2 | H | H | OH |
| M | 1fd | 2 | H | H | OCH3 |
| O | 2pd | 2 | H | OH | H |
| Q | 2cd | 2 | H | OH | OH |
| P | 2fd | 2 | H | OH | OCH3 |
| X | 3p | 1 | OCH3 | OH | H |
| Z | 3c | 1 | OCH3 | OH | OH |
| Y | 3f | 1 | OCH3 | OH | OCH3 |
| U | 3pd | 2 | OCH3 | OH | H |
| W | 3cd | 2 | OCH3 | OH | OH |
| V | 3fd | 2 | OCH3 | OH | OCH3 |
| G | 4p | 1 | OH | H | H |
| K | 4c | 1 | OH | H | OH |
| H | 4f | 1 | OH | H | OCH3 |
| R | 4pd | 2 | OH | H | H |
| T | 4cd | 2 | OH | H | OH |
| S | 4fd | 2 | OH | H | OCH3 |
| AA | 5p | 1 | OH | OH | H |
| BB | 5f | 1 | OH | OH | OCH3 |
| CC | 5c | 1 | OH | OH | OH |
| OO | 5pd | 2 | OH | OH | H |
| 5cd | 2 | OH | OH | OH | |
| PP | 5fd | 2 | OH | OH | OCH3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pretorius, C.J.; Dubery, I.A. Avenanthramides, Distinctive Hydroxycinnamoyl Conjugates of Oat, Avena sativa L.: An Update on the Biosynthesis, Chemistry, and Bioactivities. Plants 2023, 12, 1388. https://doi.org/10.3390/plants12061388
Pretorius CJ, Dubery IA. Avenanthramides, Distinctive Hydroxycinnamoyl Conjugates of Oat, Avena sativa L.: An Update on the Biosynthesis, Chemistry, and Bioactivities. Plants. 2023; 12(6):1388. https://doi.org/10.3390/plants12061388
Chicago/Turabian StylePretorius, Chanel J., and Ian A. Dubery. 2023. "Avenanthramides, Distinctive Hydroxycinnamoyl Conjugates of Oat, Avena sativa L.: An Update on the Biosynthesis, Chemistry, and Bioactivities" Plants 12, no. 6: 1388. https://doi.org/10.3390/plants12061388
APA StylePretorius, C. J., & Dubery, I. A. (2023). Avenanthramides, Distinctive Hydroxycinnamoyl Conjugates of Oat, Avena sativa L.: An Update on the Biosynthesis, Chemistry, and Bioactivities. Plants, 12(6), 1388. https://doi.org/10.3390/plants12061388