

Heat-Stress-Induced Changes in Physio-Biochemical Parameters of Mustard Cultivars and Their Role in Heat Stress Tolerance at the Seedling Stage

 , ,
, ,  ,
, .jpg) and
and
Abstract
:1. Introduction
2. Results
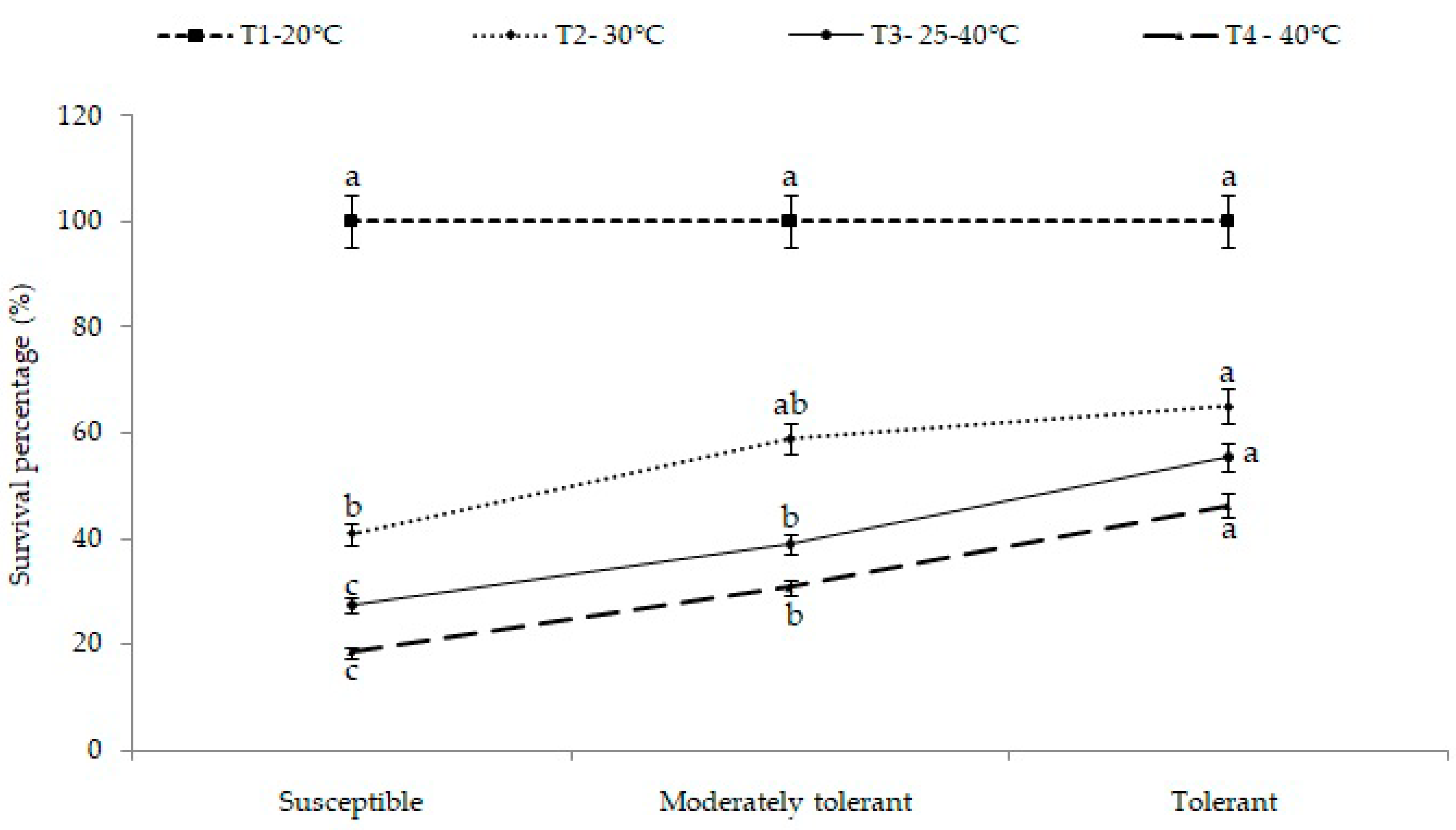
2.1. Survival (Percentage)
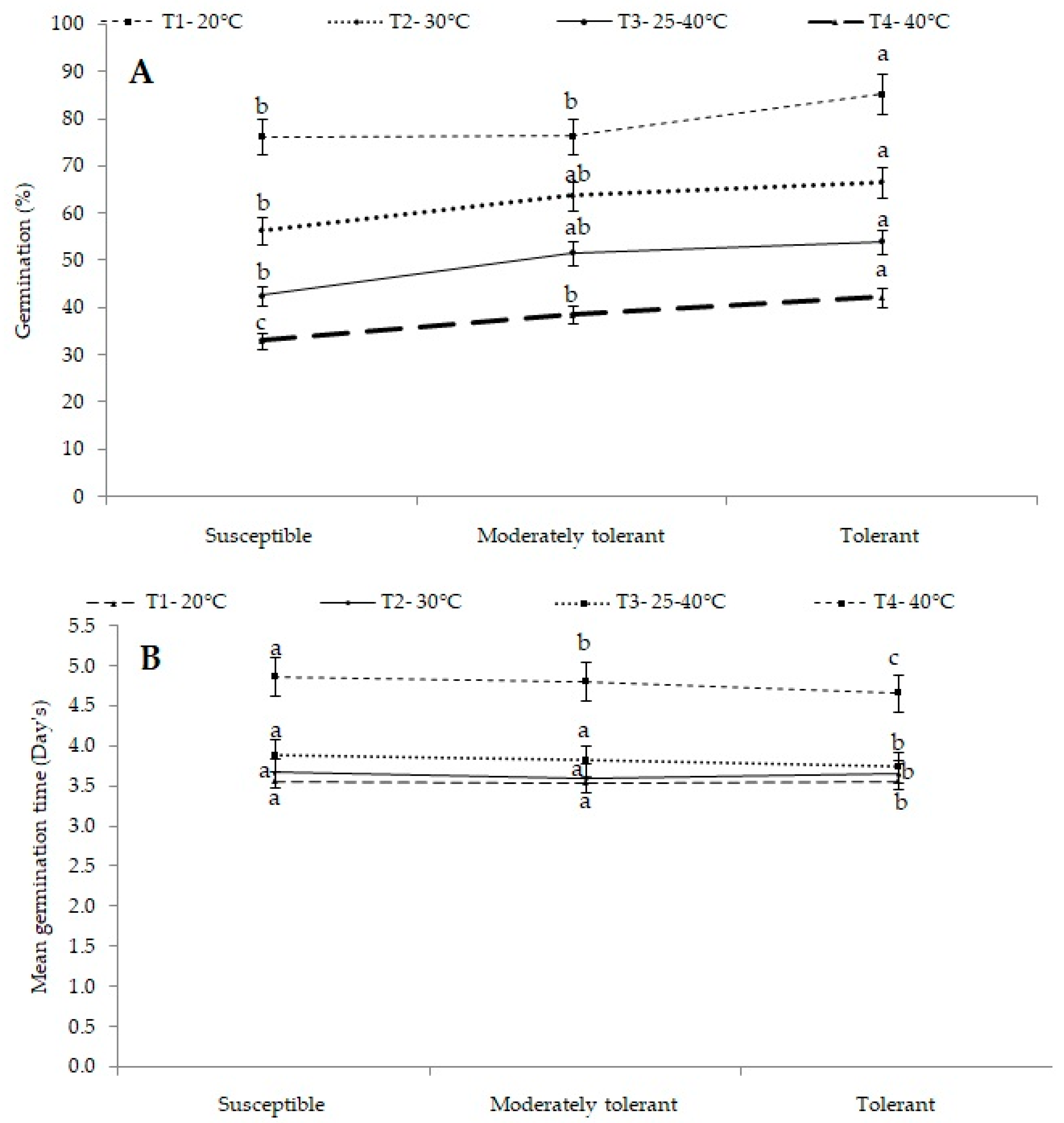
2.2. Germination Percentage and Mean Germination Time (Days)
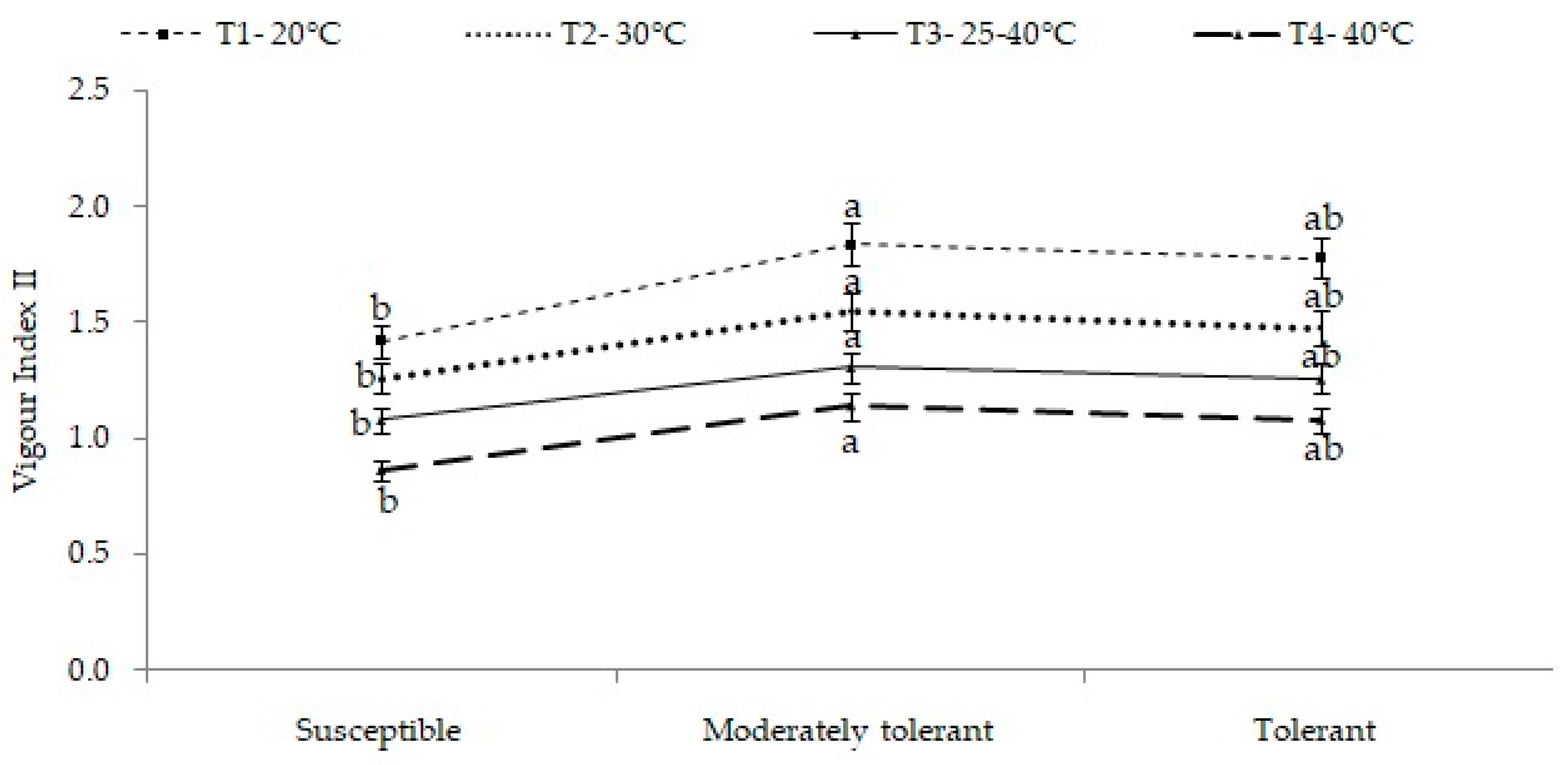
2.3. Seedling Vigor Index II (SVI-II)
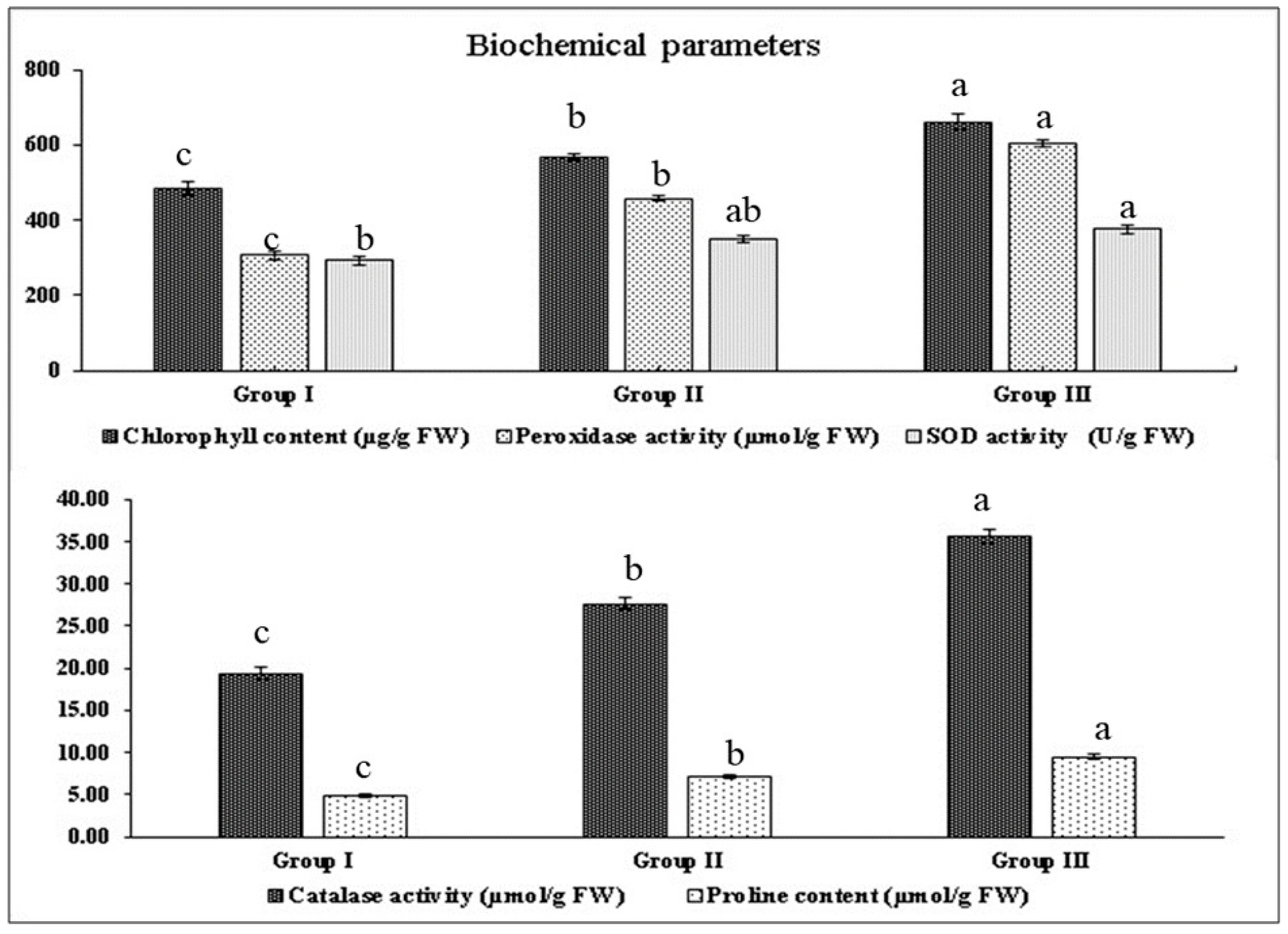
2.4. Biochemical Parameters
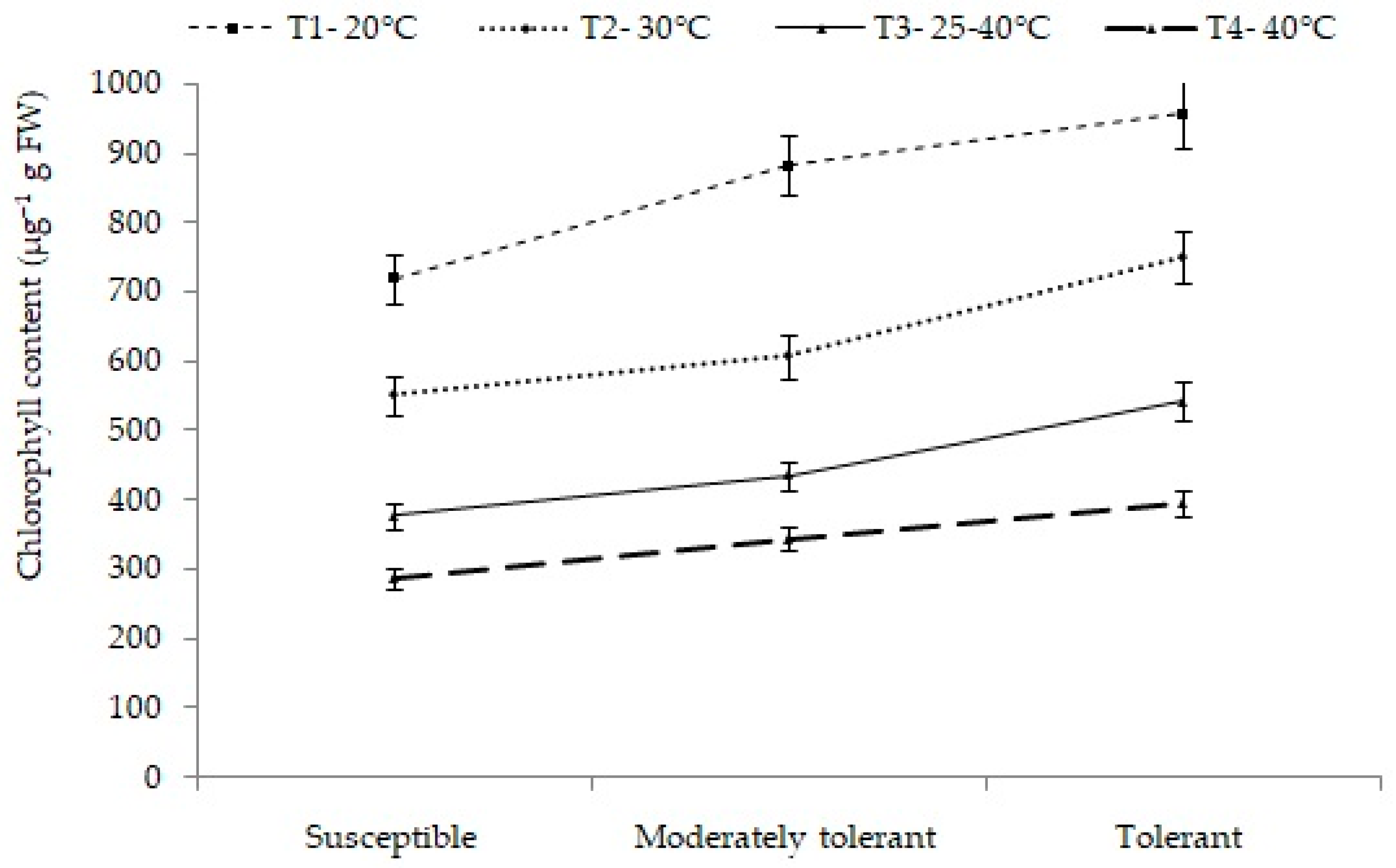
2.4.1. Chlorophyll Content (µg/g FW)
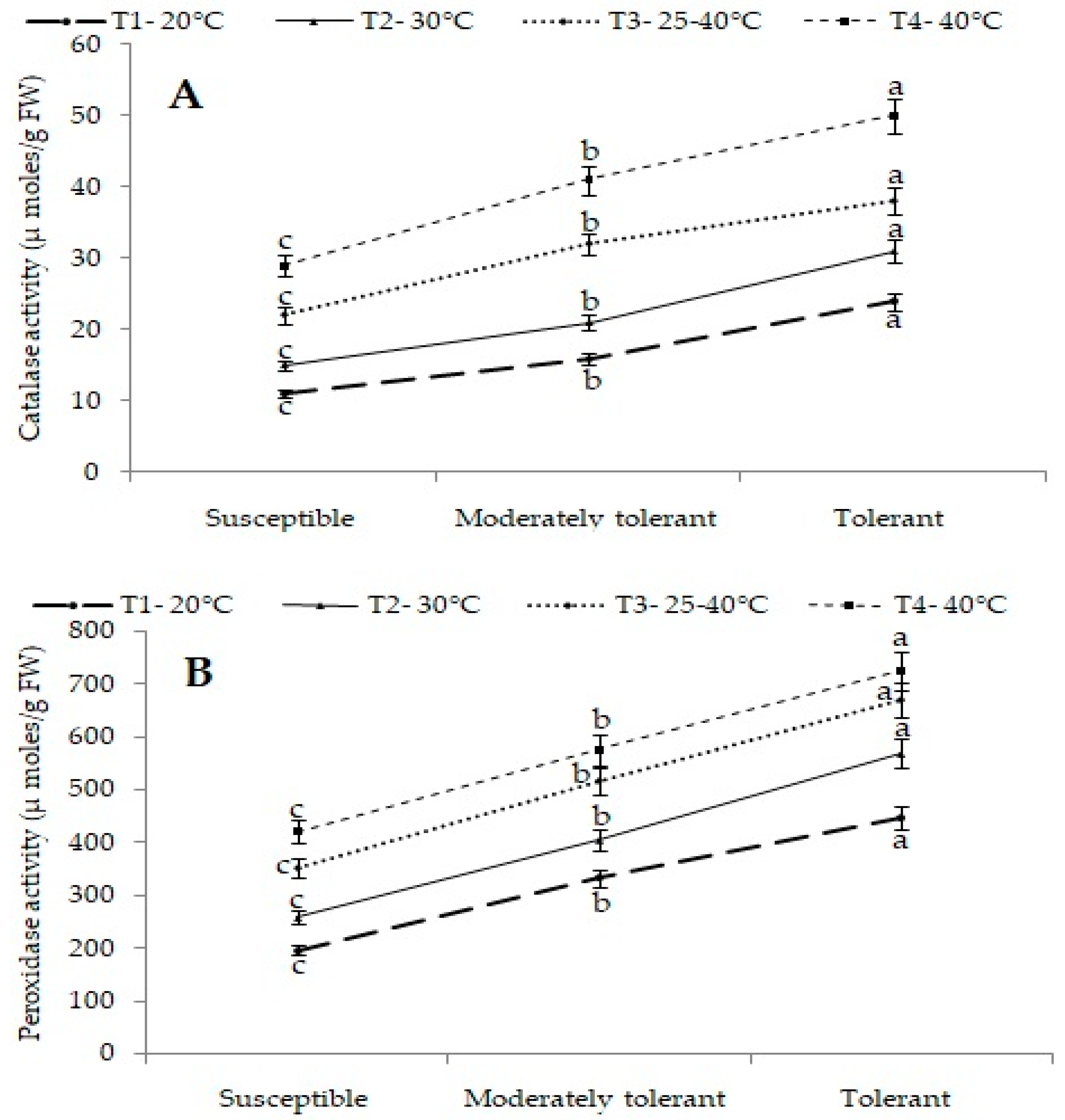
2.4.2. Antioxidant Enzyme Activities
Catalase and Peroxidase Activity (µmol/g FW)
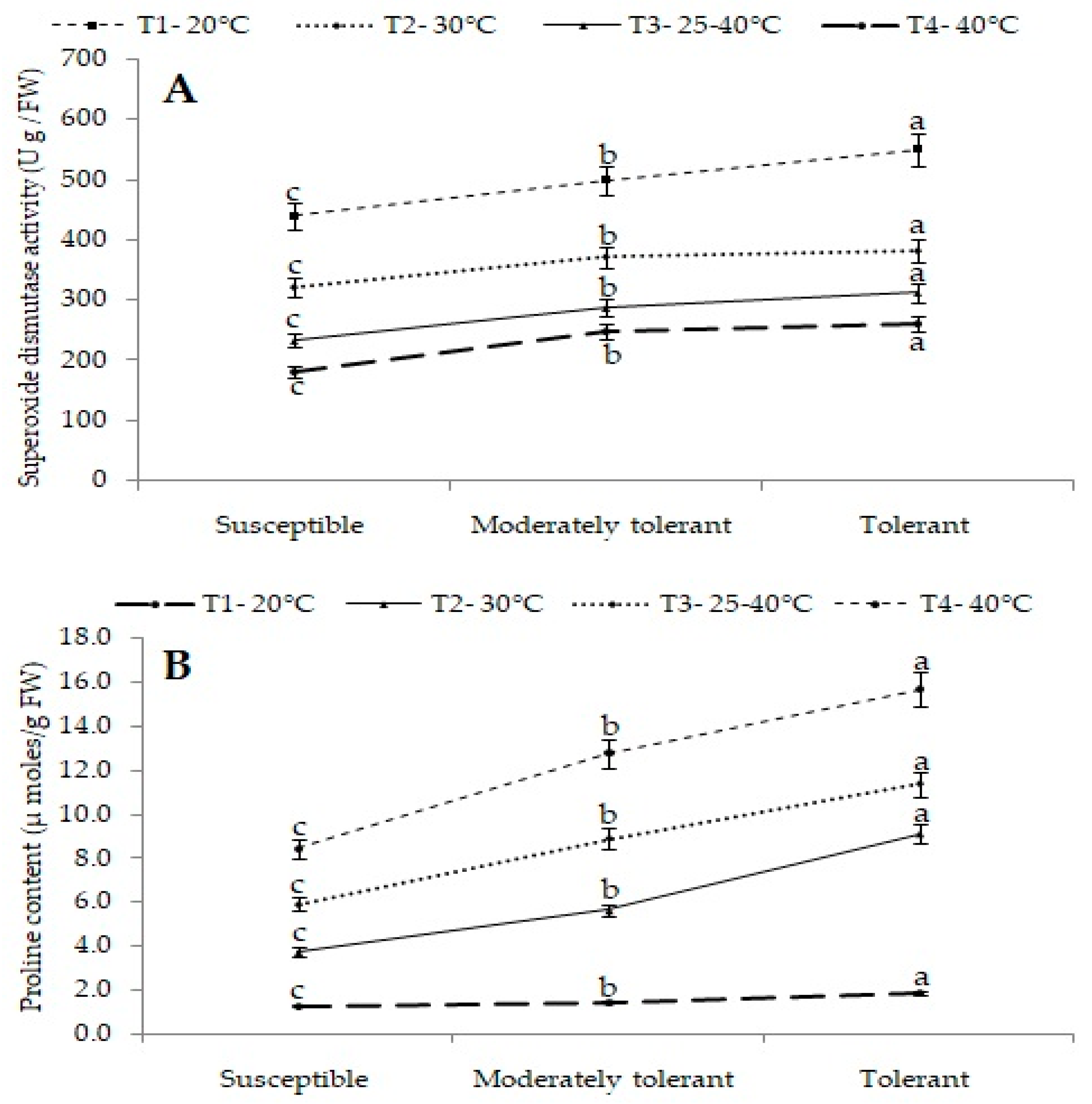
Superoxide Dismutase Activity (U/g FW)
2.4.3. Proline Content (µmol/g FW)
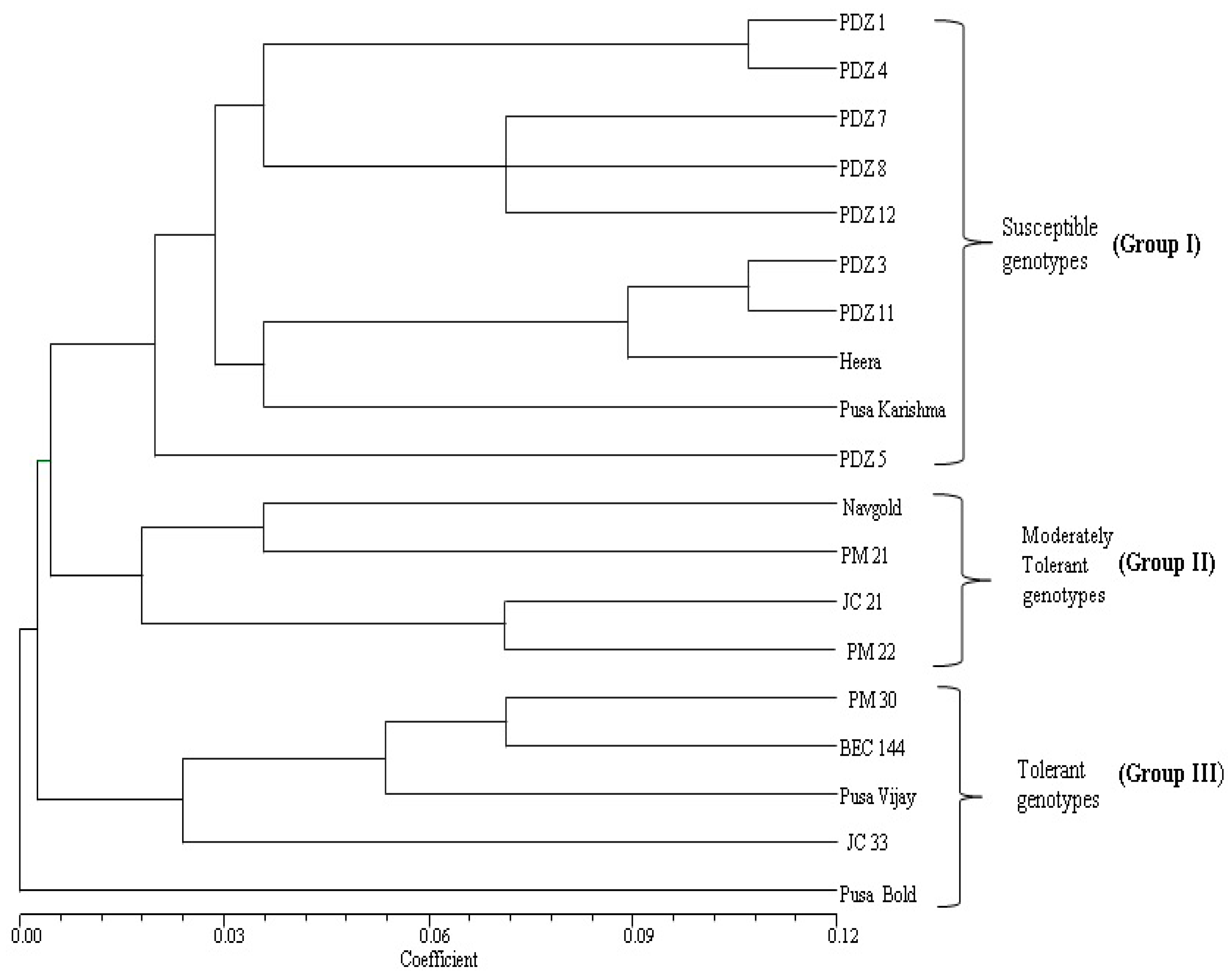
2.5. Correlation Analysis between Different Parameters
2.6. Seed Yield Attributing Traits
3. Discussion
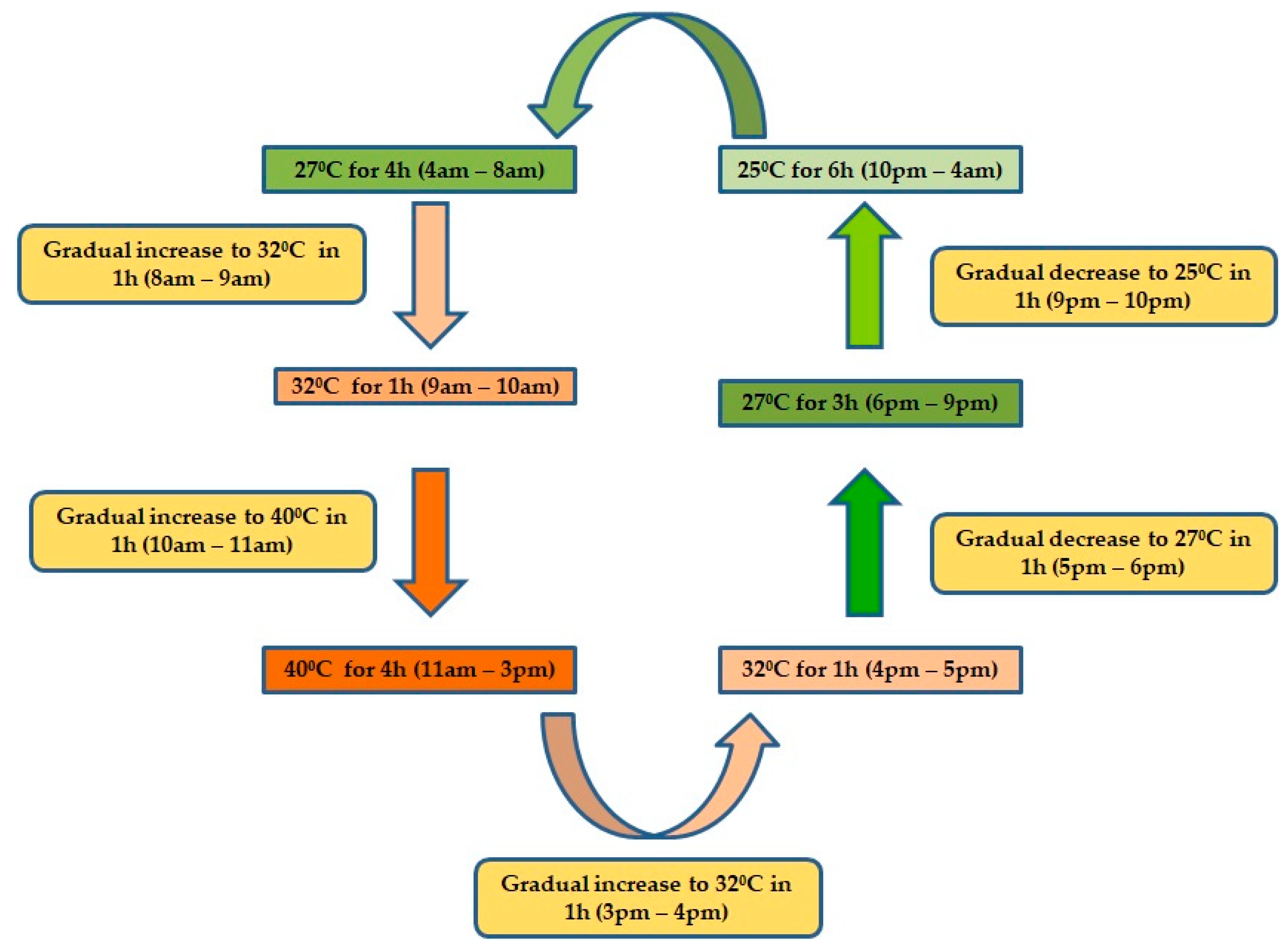
4. Materials and Methods
4.1. Experimental Materials
4.2. Methodology
4.2.1. Survival Percentage
4.2.2. Germination Percentage and Mean Germination Time (d)
4.2.3. Seedling Dry Weight (g)
4.2.4. Seedling Vigor Index II
4.2.5. Chlorophyll Content (µg/g FW)
4.2.6. Antioxidant Enzyme Activities
Catalase (CAT) Assay (µmol/g FW)
Peroxidase (POD) Assay (µmol/g FW)
Superoxide Dismutase Activity (SOD) Assay (U/g FW)
4.2.7. Proline Content (µmol/g FW)
4.2.8. Yield Parameters
No. of Primary Branches per Plant
No. of Secondary Branches per Plant
Main Shoot Length (cm)
Siliqua Length (cm)
Number of Siliquae on the Main Shoot
Siliqua Density on Main Shoot
The Number of Seeds per Siliqua
Seed Yield per Plant (g)
4.3. Experimental Design and Statistical Analyses
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tomlinson, I. Doubling food production to feed the 9 billion: A critical perspective on a key discourse of food security in the UK. J. Rural Stud. 2013, 29, 81–90. [Google Scholar] [CrossRef]
- Stocher, T.F.; Plattner, G.K.; Tignor, M.M.B.; Allen, S.K.; Boschung, J.; Nauel, A.; Xia, Y.; Bex, V.; Midgley, P. Climate Change 2013: The Physical Science Basis: Working Group I Contribution to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2013. [Google Scholar]
- Begcy, K.; Dresselhaus, T. Epigenetic responses to abiotic stresses during reproductive development in cereals. Plant Reprod. 2018, 31, 343–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sehgal, A.; Kumari, S.; Siddique, K.H.M.; Kumar, R.; Bhogireddy, S.; Varshney, R.K.; Rao, B.H.; Nair, R.M.; Prasad, P.V.V.; Nayyar, H. Drought or/and heat-stress effects on seed filling in food crops: Impacts on functional biochemistry, seed yields, and nutritional quality. Front. Plant Sci. 2018, 9, 1705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Folsom, J.J.; Begcy, K.; Hao, X.; Wang, D.; Walia, H. Rice fertilization-Independent Endosperm1 regulates seed size under high temperature stress by controlling early endosperm development. Plant Physiol. 2014, 165, 238–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hameed, A.; Goher, M.; Iqbal, N. Heat stress-induced cell death, changes in antioxidants, lipid peroxidation, and protease activity in wheat leaves. J. Plant Growth Regul. 2012, 31, 283–291. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Ort, D.R. How do we improve crop production in a warming world? Plant Physiol. 2010, 154, 526–530. [Google Scholar] [CrossRef] [Green Version]
- Biswal, B.; Joshi, P.N.; Raval, M.K.; Biswal, U.C. Photosynthesis, a global sensor of environmental stress in green plants: Stress signalling and adaptation. Curr. Sci. 2011, 101, 47–56. [Google Scholar]
- Gupta, N.K.; Agarwal, S.; Agarwal, V.P.; Nathawat, N.S.; Gupta, S.; Singh, G. Effect of short-term high temperature stress on growth, physiology and antioxidative defence system in wheat seedlings. Acta Physiol. Plant. 2013, 35, 1837–1842. [Google Scholar] [CrossRef]
- Erdal, S. Androsterone-induced molecular and physiological changes in maize seedlings in response to chilling stress. Plant Physiol. Biochem. 2012, 57, 1–7. [Google Scholar] [CrossRef]
- Balestrasse, K.B.; Tomaro, M.L.; Batlle, A.; Noriega, G.O. The role of 5-aminolevulinic acid in the response to cold stress in soybean plants. Phytochemistry 2010, 71, 2038–2045. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 2012, 217037. [Google Scholar] [CrossRef] [Green Version]
- Bhuyan, M.B.; Hasanuzzaman, M.; Parvin, K.; Mohsin, S.M.; Al Mahmud, J.; Nahar, K.; Fujita, M. Nitric oxide and hydrogen sulfide: Two intimate collaborators regulating plant defense against abiotic stress. Plant Growth Regul. 2020, 90, 409–424. [Google Scholar] [CrossRef]
- Szepesi, Á.; Szőllősi, R. Mechanism of proline biosynthesis and role of proline metabolism enzymes under environmental stress in plants. In Plant Metabolites and Regulation under Environmental Stress; Academic Press: Cambridge, MA, USA, 2018; pp. 337–353. [Google Scholar]
- Slama, I.; Abdelly, C.; Bouchereau, A.; Flowers, T.; Savouré, A. Diversity, distribution and roles of osmoprotective compounds accumulated in halophytes under abiotic stress. Ann. Bot. 2015, 115, 433–447. [Google Scholar] [CrossRef] [Green Version]
- Iwaniuk, P.; Lozowicka, B. Biochemical compounds and stress markers in lettuce upon exposure to pathogenic Botrytis cinerea and fungicides inhibiting oxidative phosphorylation. Planta 2022, 255, 61. [Google Scholar] [CrossRef]
- DAFW Department of Agriculture & Farmers Welfare Ministry of Agriculture & Farmers Welfare, Government of India. 2020. Available online: https://agricoop.nic.in/en (accessed on 23 December 2020).
- Shekhawat, K.; Rathore, S.S.; Premi, O.P.; Kandpal, B.K.; Chauhan, J.S. Advances in agronomic management of Indian mustard (Brassica juncea (L.) Czern&Coss): An overview. Int. J. Agron. 2012, 2012, 408284. [Google Scholar]
- Niwas, R.; Khichar, M.L. Managing impact of climatic vagaries on the productivity of wheat and mustard in India. Mausam 2016, 67, 205–222. [Google Scholar] [CrossRef]
- Kateřina, M.; Prabhullachandran, U.; Štefková, M.; Spyroglou, I.; Pěnčík, A.; Endlová, L.; Novak, O.; Robert, H.S. Long-term high-temperature stress impacts on embryo and seed development in Brassica napus. Front. Plant Sci. 2022, 13, 844292. [Google Scholar]
- Azharudheen, T.P.M.; Yadava, D.K.; Singh, N.; Vasudev, S.; Singh, R.; Prabhu, K.V. A study on the thermo-tolerance at germination and seedling stage in Indian mustard [Brassica juncea (L.) Czern&Coss]. Int. J. Agric. Food Sci. 2013, 4, 589–594. [Google Scholar]
- Zhang, J.; Jiang, F.; Yang, P.; Li, J.; Yan, G.; Hu, L. Responses of canola (Brassica napus L.) cultivars under contrasting temperature regimes during early seedling growth stage as revealed by multiple physiological criteria. Acta Physiol. Plant. 2015, 37, 7. [Google Scholar] [CrossRef]
- Lietzow, J. Biologically active compounds in mustard seeds: A toxicological perspective. Foods 2021, 10, 2089. [Google Scholar] [CrossRef]
- Swami, S.; Yadav, S.; Yadav, S.K.; Dahuja, A.; Yadava, D.K. Imbibition behaviour and germination response in conventional and quality of Indian mustard (Brassica juncea) seeds. Indian J. Agric. Sci. 2016, 86, 1625–1629. [Google Scholar]
- Biswas, N.; Yadav, S.; Yadav, S.; Choudhary, R.; Saini, N.; Dahuja, A.; Yadav, D.K. Vigor difference during storage and germination in Indian mustard explained by reactive oxygen species and antioxidant enzymes. Turk. J. Agric. 2020, 44, 577–588. [Google Scholar] [CrossRef]
- Singh, V.; Pant, U.; Bhajan, R. Physiological traits versus seed yield derived parameters based high temperature stress study in Indian mustard. SABRAO J. Anim. Breed. Genet. 2016, 48, 127–135. [Google Scholar]
- Liu, Y.; Li, J.; Zhu, Y.; Jones, A.; Rose, R.J.; Song, Y. Heat stress in legume seed setting: Effects, causes, and future prospects. Front. Plant Sci. 2019, 10, 938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- IPCC. Climate Change 2007: Impacts, Adaptation and Vulnerability. In Contribution of Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Parry, M.L., Canziani, O.F., Palutikof, J.P., van der Linden, P.J., Hanson, C.E., Eds.; Cambridge University Press: Cambridge, UK, 2007; 976p. [Google Scholar]
- Raza, A.; Razzaq, A.; Mehmood, S.S.; Zou, X.; Zhang, X.; Lv, Y.; Xu, J. Impact of climate change on crops adaptation and strategies to tackle its outcome: A review. Plants 2019, 8, 34. [Google Scholar] [CrossRef] [Green Version]
- Hossain, A.; Sarker, M.A.Z.; Saifuzzaman, M.; Teixeira da Silva, J.A.; Lozovskaya, M.V.; Akhter, M.M. Evaluation of growth, yield, relative performance and heat susceptibility of eight wheat (Triticum aestivum L.) cultivars grown under heat stress. Int. J. Plant Prod. 2013, 7, 615–636. [Google Scholar]
- Sharma, S.; Singh, V.; Tanwar, H.; Mor, V.S.; Kumar, M.; Punia, R.C.; Singh, J. Impact of high temperature on germination, seedling growth and enzymatic activity of wheat. Agriculture 2022, 12, 1500. [Google Scholar] [CrossRef]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S.; et al. Crop production under drought and heat stress: Plant responses and management options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef] [Green Version]
- Mondal, S.; Bose, B. Seed priming: An interlinking technology between seeds, seed germination and seedling establishment. In Plant Reproductive Ecology-Recent Advances; IntechOpen: London, UK, 2021. [Google Scholar] [CrossRef]
- Soltani, E.; Ghaderi-Far, F.; Baskin, C.C.; Baskin, J.M. Problems with using mean germination time to calculate rate of seed germination. Aust. J. Bot. 2015, 63, 631–635. [Google Scholar] [CrossRef]
- Toh, S.; Imamura, A.; Watanabe, A.; Nakabayashi, K.; Okamoto, M.; Jikumaru, Y. High temperature-induced abscisic acid biosynthesis and its role in the inhibition of gibberellin action in Arabidopsis seeds. Plant Physiol. 2008, 146, 1368–1385. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, N.; Mittler, R. Reactive oxygen species and temperature stresses: A delicate balance between signalling and destruction. Physiol. Plant. 2006, 126, 45–51. [Google Scholar] [CrossRef]
- Han, F.; Chen, H.; Li, X.J.; Yang, M.F.; Liu, G.S.; Shen, S.H. A comparative proteomic analysis of rice seedlings under various high-temperature stresses. BiochimBiophys Acta 2009, 1794, 1625–1634. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.M.; Roychowdhury, R.; Fujita, M. Physiological, biochemical, and molecular mechanisms of heat stress tolerance in plants. Int. J. Mol. Sci. 2013, 14, 9643–9684. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Ding, Y.; Zhu, C. Sensitivity and responses of chloroplasts to high temperature stress in plants. Front. Plant Sci. 2020, 11, 375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wassie, M.; Zhang, W.; Zhang, Q.; Ji, K.; Chen, L. Effect of heat stress on growth and physiological traits of alfalfa (Medicago sativa L.) and a comprehensive evaluation for heat tolerance. Agronomy 2019, 9, 597. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Gupta, D.; Nayyar, H. Comparative response of maize and rice cultivars to heat stress: Status of oxidative stress and antioxidants. Acta Physiol. Plant. 2012, 34, 75–86. [Google Scholar] [CrossRef]
- Gosavi, G.U.; Jadhav, A.S.; Kale, A.A.; Gadakh, S.R.; Pawar, B.D.; Chimote, V.P. Effect of heat stress on proline, chlorophyll content, heat shock proteins and antioxidant enzyme activity in sorghum (Sorghum bicolor) at seedlings stage. Indian J. Biotechnol. 2014, 13, 356–363. [Google Scholar]
- Berjak, P.; Pammenter, N. Implications of the lack of desiccation tolerance in recalcitrant seeds. Front. Plant Sci. 2013, 4, 478. [Google Scholar] [CrossRef] [Green Version]
- Hasanuzzaman, M.; Bhuyan, M.H.M.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Mahmud, J.A.; Fotopoulos, V. Reactive oxygen species and antioxidant defense in plants under abiotic stress: Revisiting the crucial role of a universal defense regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef]
- Iwaniuk, P.; Borusiewicz, A.; Lozowicka, B. Fluazinam and its mixtures induce diversified changes of crucial biochemical and antioxidant profile in leafy vegetable. Sci. Hortic. 2022, 298, 110988. [Google Scholar] [CrossRef]
- Harsh, A.; Sharma, Y.K.; Joshi, U.; Rampuria, S.; Singh, G.; Kumar, S.; Sharma, R. Effect of short-term high temperature stress on total sugars, proline and some antioxidant enzymes in moth bean (Vigna aconitifolia). Ann. Agric. Sci. 2016, 61, 57–64. [Google Scholar] [CrossRef] [Green Version]
- Caverzan, A.; Casassola, A.; Brammer, S.P. Antioxidant responses of wheat plants under stress. Genet. Mol. Biol. 2016, 39, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kibinza, S.; Vinel, D.; Come, D.; Bailly, C.; Corbineau, F. Sunflower seed deterioration as related to moisture content during ageing, energy metabolism and active oxygen species scavenging. Physiol. Plant 2006, 128, 496–506. [Google Scholar] [CrossRef]
- Tabatabaei, S.; Ehsanzadeh, P. Photosynthetic pigments, ionic and antioxidative behaviour of hulled tetraploid wheat in response to NaCl. Photosynthetic 2016, 54, 340–350. [Google Scholar] [CrossRef]
- Dar, M.I.; Naikoo, M.I.; Rehman, F.; Naushin, F.; Khan, F.A. Proline accumulation in plants: Roles in stress tolerance and plant development. In Osmolytes and Plants Acclimation to Changing Environment: Emerging Omics Technologies; Springer: Berlin/Heidelberg, Germany, 2016; pp. 155–166. [Google Scholar]
- Siddique, A.; Kandpal, G.; Kumar, P. Proline accumulation and its defensive role under diverse stress condition in plants: An overview. J. Pure Appl. Microbiol. 2018, 12, 1655–1659. [Google Scholar] [CrossRef]
- Kishor, P.K.; Suravajhala, P.; Rathnagiri, P.; Sreenivasulu, N. Intriguing role of proline in redox potential conferring high temperature stress tolerance. Front. Plant Sci. 2022, 13, 867531. [Google Scholar] [CrossRef] [PubMed]
- Chaplot, P.C.; Vandeep, A.; Kumar, R. Effect of balanced fertilization and agrochemical on growth, yield attributes and yields of mustard varieties. Ext. Summ. 2012, 3, 26–30. [Google Scholar]
- Amutha, R.S.; Muthulaksmi, W.; Baby Rani, K.; Mareeswari, P.I. Studies on biochemical basis of heat tolerance in sunflower (Helianthus annuus L.). J. Agric. Biol. Sci. 2007, 3, 234–238. [Google Scholar]
- Saxena, M.C.; Saxena, N.P.; Mohamed, A.K. High temperature stress. In World Crops: Cool Season Food Legumes, Current Plant Science and Biotechnology in Agriculture; Summerfeld, R.J., Ed.; Springer: Dordrecht, The Netherlands, 1988; Volume 5. [Google Scholar]
- Bita, C.E.; Gerats, T. Plant tolerance to high temperature in a changing environment: Scientific fundamentals and production of heat stress-tolerant crops. Front. Plant Sci. 2013, 4, 273. [Google Scholar] [CrossRef] [Green Version]
- Sarsu, F. Screening protocols for heat tolerance in rice at the seedling and reproductive stages. In Pre-Field Screening Protocols for Heat-Tolerant Mutants in Rice; Springer: Berlin/Heidelberg, Germany, 2018; pp. 9–24. [Google Scholar] [CrossRef] [Green Version]
- ISTA. International Rules for Seed Testing; International Seed Testing Association: Bassersdorf, Switzerland, 2020. [Google Scholar]
- Nicholas, M.A.; Heydecker, W. Two approaches to the study of germination data. Proc. Int. Seed Test. Assoc. 1968, 33, 531–540. [Google Scholar]
- Abdul-Baki, A.A.; Anderson, J.D. Vigour determination in soybean seed by multiple criteria. Crop Sci. 1973, 13, 630–633. [Google Scholar] [CrossRef]
- Arnon, D. Copper enzymes isolated chloroplasts, polyphenol oxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raza, S.H.; Athar, H.R.; Ashraf, M.; Hameed, A. Glycine betaine induced modulation of antioxidant enzymes activities and ion accumulation in two wheat cultivars differing in salt tolerance. Environ. Exp. Bot. 2007, 60, 368–376. [Google Scholar] [CrossRef]
- Aebi, H.E. Catalase. In Methods of Enzymatic Analysis, 3rd ed.; Bergmeyer, H.U., Ed.; Verlag Chemie: Weinheim, Germany, 1983; pp. 273–286. [Google Scholar]
- Gao, J.; Xiao, Q.; Ding, L.; Chen, M.; Yin, L.; Li, J.; Zhou, S.; He, G. Differential responses of lipid peroxidation and antioxidants in Alternanthera philoxeroides and Oryza sativa subjected to drought stress. Plant Growth Regul. 2008, 56, 89–95. [Google Scholar] [CrossRef]
- Donahue, J.L.; Okpodu, C.M.; Cramer, C.; Grabau, E.A.; Alscher, R.G. Responses of antioxidants to paraquat in pea leaves (relationships to resistance). Plant Physiol. 1997, 113, 249–257. [Google Scholar] [CrossRef] [Green Version]
- Bates, L.S. Rapid determination of free proline for water-stress studies. Plant Soil. 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Rohlf, F.J. NTSYS 2.1: Numerical Taxonomic and Multivariate Analysis System; Exeter Software: New York, NY, USA, 2000. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Susceptible Cultivars | Moderately Tolerant Cultivars | Tolerant Cultivars | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Temperatures | PDZ-1 | PDZ-3 | PDZ-4 | PDZ-5 | PDZ-7 | PDZ-8 | PDZ-11 | PDZ-12 | Pusa Karishma | Heera | JC-21 | PM-21 | PM-22 | Navgold | PM-30 | Pusa Bold | Pusa Vijay | JC-33 | BEC-144 | Mean |
| T1 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 a (±0.5) |
| T2 | 34.1 | 41.2 | 37.1 | 38.1 | 32.3 | 38.2 | 40.3 | 48.2 | 43.3 | 56.1 | 55.3 | 53.2 | 65.2 | 59.1 | 70.2 | 75.1 | 65.3 | 56.5 | 58.4 | 50.9 b (±3.7) |
| T3 | 30.0 | 28.2 | 26.4 | 27.3 | 28.1 | 32.1 | 22.3 | 29.1 | 22.3 | 27.1 | 42.1 | 37.2 | 41.1 | 35.1 | 65.1 | 74.2 | 49.2 | 48.1 | 41.2 | 37.8 c (±3.8) |
| T4 | 22.4 | 21.5 | 19.2 | 15.2 | 20.1 | 19.3 | 18.3 | 16 | 17.1 | 15.3 | 35.3 | 28.2 | 31.3 | 28.1 | 53.2 | 53.5 | 39.1 | 38.2 | 48.3 | 28.4 d (±3.4) |
| Mean (Temp.) | 46.6 gh (±6.7) | 47.7 g (±6.7) | 45.7 h (±6.9) | 45.1 h (±7.3) | 45.1 h (±6.7) | 47.4 g (±6.7) | 45.2 h (±7.0) | 48.3 f (±6.9) | 45.7 h (±7.0) | 49.6 f (±7.0) | 58.2 de (±5.4) | 54.7 e (±5.7) | 59.4 d (±5.2) | 55.6 e (±6.0) | 72.1 b (±3.7) | 75.7 a (±3.6) | 63.4 c (±5.1) | 60.7 d (±6.0) | 62.0 cd (±5.0) | 54.11 |
| CD (p = 0.05); G = 1.04 **; T = 0.48 **; G × T = 2.09 ** | ||||||||||||||||||||
| Groups | Cultivars | Chlorophyll Content | Catalase Activity | Peroxidase Activity | Superoxide Dismutase | Proline Content |
|---|---|---|---|---|---|---|
| (µg/g FW) | (µmol/g FW) | (µmol/g FW) | Activity(U/g FW) | (µmol/g FW) | ||
| Susceptible cultivars | PDZ-1 | 437 ef (±3.7) | 21.9 d (±0.3) | 338 d (± 4.8) | 297 c (±2.8) | 4.86 de (±0.05) |
| PDZ-3 | 483 e (±3.0) | 19.3 e (±0.1) | 272 e (±1.9) | 291 c (±1.9) | 4.51 e (±0.05) | |
| PDZ-4 | 399 f (±5.2) | 19.6 e (±0.2) | 328 d (±2.8) | 284 cd (±1.6) | 4.45 f (±0.05) | |
| PDZ-5 | 395 f (±6.9) | 17.3 f (±0.2) | 298 de (±4.3) | 276 d (±1.4) | 4.55 e (±0.07) | |
| PDZ-7 | 496 e (±7.0) | 16.0 f (±0.1) | 310 d (±2.5) | 285 cd( ±1.8) | 4.88 de (±0.06) | |
| PDZ-8 | 545 d (±2.9) | 20.0 d (±0.1) | 346 d (±3.5) | 306 c (±1.3) | 5.04 d (±0.05) | |
| PDZ-11 | 498 e (±5.3) | 17.7 f (±0.2) | 266 e (±2.0) | 277 d (±1.9) | 4.70 de (±0.04) | |
| PDZ-12 | 565 cd (±5.4) | 22.9 d (±0.2) | 333 d (±2.7) | 311 bc (±1.8) | 6.04 c (±0.05) | |
| Pusa Karishma | 493 e (±4.4) | 20.2 d (±0.1) | 318 de (±3.8) | 285 cd (±1.5) | 4.52 e (±0.02) | |
| Heera | 530 de (±6.7) | 19.4 e (±0.2) | 265 e (±2.7) | 312 bc (±1.8) | 4.68 de (±0.01) | |
| Moderately tolerant cultivars | JC-21 | 487 e (±4.0) | 29.7 c (±0.3) | 469 c (±2.5) | 333 b (±1.3) | 5.06 c (±0.07) |
| PM-21 | 589 c (±5.7) | 28.5 c (±0.3) | 418 cd (±4.5) | 355 ab (±1.4) | 6.89 c (±0.14) | |
| PM-22 | 659 b (±3.7) | 26.7 cd (±0.4) | 498 c (±4.8) | 392 a (±2.5) | 9.98 a (±0.17) | |
| Navgold | 534 de (±4.6) | 25.7 cd (± 0.1) | 449 c (±4.4) | 324 bc (±4.5) | 6.79 c (±0.05) | |
| Tolerant cultivars | PM-30 | 749 a (±5.9) | 36.4 a (±0.3) | 633 a (±5.4) | 387 a (±1.4) | 10.39 a (±0.10) |
| Pusa Bold | 739 a (±10.5) | 37.3 a (±0.2) | 633 a (±5.2) | 404 a (±2.1) | 10.12 a (± 0.09) | |
| Pusa Vijay | 689 b (±5.9) | 35.6 ab (± 0.1) | 606 ab (±4.2) | 360 b (±2.5) | 9.14 ab (±0.16) | |
| JC-33 | 544 d (±5.8) | 33.8 b (±0.4) | 603 ab (±3.6) | 374 ab (±1.9) | 9.44 a (±0.08) | |
| BEC-144 | 585 c (±8.8) | 35.1 ab (±0.6) | 543 b (±1.6) | 352 b (±2.1) | 8.40 b (±0.07) | |
| Grand Mean | 548 | 25.4 | 417 | 327 | 6.55 | |
| CD (p =0.05) | 7.97 ** | 0.33 ** | 4.08 ** | 4.25 ** | 0.11 ** | |
| Germination % | Survival Percentage | MGT | Vigor Index II | Chlorophyll Content | Catalase Activity | Peroxidase Activity | SOD Activity | Proline Content | |
|---|---|---|---|---|---|---|---|---|---|
| Germination % | 1.000 | ||||||||
| Survival percentage | 0.836 ** | 1.000 | |||||||
| MGT | −0.734 ** | −0.575 ** | 1.000 | ||||||
| Vigor index II | 0.837 ** | 0.756 ** | −0.665 ** | 1.000 | |||||
| Chlorophyll content | 0.800 ** | 0.858 ** | −0.636 ** | 0.816 ** | 1.000 | ||||
| Catalase activity | −0.499 ** | −0.402 ** | 0.598 ** | −0.291 ** | −0.430 ** | 1.000 | |||
| Peroxidase activity | −0.363 ** | −0.226 ** | 0.448 ** | −0.100 ** | −0.198 ** | 0.905 ** | 1.000 | ||
| SOD activity | 0.848 ** | 0.923 ** | −0.666 ** | 0.832 ** | 0.925 ** | −0.428 ** | −0.212 ** | 1.000 | |
| Proline content | −0.590 ** | −0.544 ** | 0.647 ** | −0.439 ** | −0.559 ** | 0.909 ** | 0.824** | −0.576 ** | 1.000 |
| Cultivars | No. of Primary Branches | No. of Secondary Branches | Main Shoot Length (cm) | No. of Siliquae on Main Shoot | Siliqua Length (cm) | No. of Seeds per Siliqua | Siliqua Density on Main Shoot | Seed Yield per Plant (g) |
|---|---|---|---|---|---|---|---|---|
| Susceptible | 8.5 c (±0.26) | 22.8 c (±4.67) | 50.6 c (±1.06) | 31.6 c (±2.62) | 3.37 c (±0.19) | 13.7 c (±0.37) | 0.74 c (±0.04) | 31.9 c (±1.31) |
| Moderately tolerant | 9.1 b (±0.09) | 24.2 b (±1.20) | 57.1 b (±2.22) | 33.6 b (±1.13) | 3.87 b (±0.26) | 13.5 b (±1.65) | 0.62 b (±0.08) | 34.8 b (±1.66) |
| Tolerant | 10.8 a (±0.26) | 29.0 a (±0.88) | 69.5 a (±2.83) | 39.3 a (±0.75) | 5.01 a (±0.11) | 13.7 a (±1.61) | 0.68 a (±0.06) | 40.9 a (±2.39) |
| Mean | 9.5 | 25.3 | 59.1 | 34.8 | 4.08 | 13.6 | 0.68 | 35.8 |
| CD (p =0.05) | 0.54 ** | 6.88 * | 5.28 ** | 4.15 ** | 0.48 ** | NS | NS | 4.49 ** |
| Months | Tmax (°C) | Tmin (°C) | Relative Humidity (%) | Rainfall (mm) | Sunshine (Hrs) |
|---|---|---|---|---|---|
| October | 32.3 | 19.3 | 74.1 | 4.2 | 7.5 |
| November | 27.12 | 10.59 | 71.78 | 0 | 4.5 |
| December | 21.88 | 7.3 | 77.4 | 6 | 3.73 |
| January | 17.37 | 7.45 | 84 | 0 | 2.45 |
| February | 22.82 | 8.54 | 71.61 | 0 | 6.5 |
| March | 31.62 | 14.97 | 61.21 | 0 | 8.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sakpal, A.; Yadav, S.; Choudhary, R.; Saini, N.; Vasudev, S.; Yadava, D.K.; Ercişli, S.; Marc, R.A.; Yadav, S.K. Heat-Stress-Induced Changes in Physio-Biochemical Parameters of Mustard Cultivars and Their Role in Heat Stress Tolerance at the Seedling Stage. Plants 2023, 12, 1400. https://doi.org/10.3390/plants12061400
Sakpal A, Yadav S, Choudhary R, Saini N, Vasudev S, Yadava DK, Ercişli S, Marc RA, Yadav SK. Heat-Stress-Induced Changes in Physio-Biochemical Parameters of Mustard Cultivars and Their Role in Heat Stress Tolerance at the Seedling Stage. Plants. 2023; 12(6):1400. https://doi.org/10.3390/plants12061400
Chicago/Turabian StyleSakpal, Ashwini, Sangita Yadav, Ravish Choudhary, Navinder Saini, Sujata Vasudev, Devendra K. Yadava, Sezai Ercişli, Romina Alina Marc, and Shiv K. Yadav. 2023. "Heat-Stress-Induced Changes in Physio-Biochemical Parameters of Mustard Cultivars and Their Role in Heat Stress Tolerance at the Seedling Stage" Plants 12, no. 6: 1400. https://doi.org/10.3390/plants12061400
APA StyleSakpal, A., Yadav, S., Choudhary, R., Saini, N., Vasudev, S., Yadava, D. K., Ercişli, S., Marc, R. A., & Yadav, S. K. (2023). Heat-Stress-Induced Changes in Physio-Biochemical Parameters of Mustard Cultivars and Their Role in Heat Stress Tolerance at the Seedling Stage. Plants, 12(6), 1400. https://doi.org/10.3390/plants12061400