
Metabolomic Study of Flavonoids in Camellia drupifera under Aluminum Stress by UPLC-MS/MS

 , , and
, , and 
Abstract
:1. Introduction
2. Results
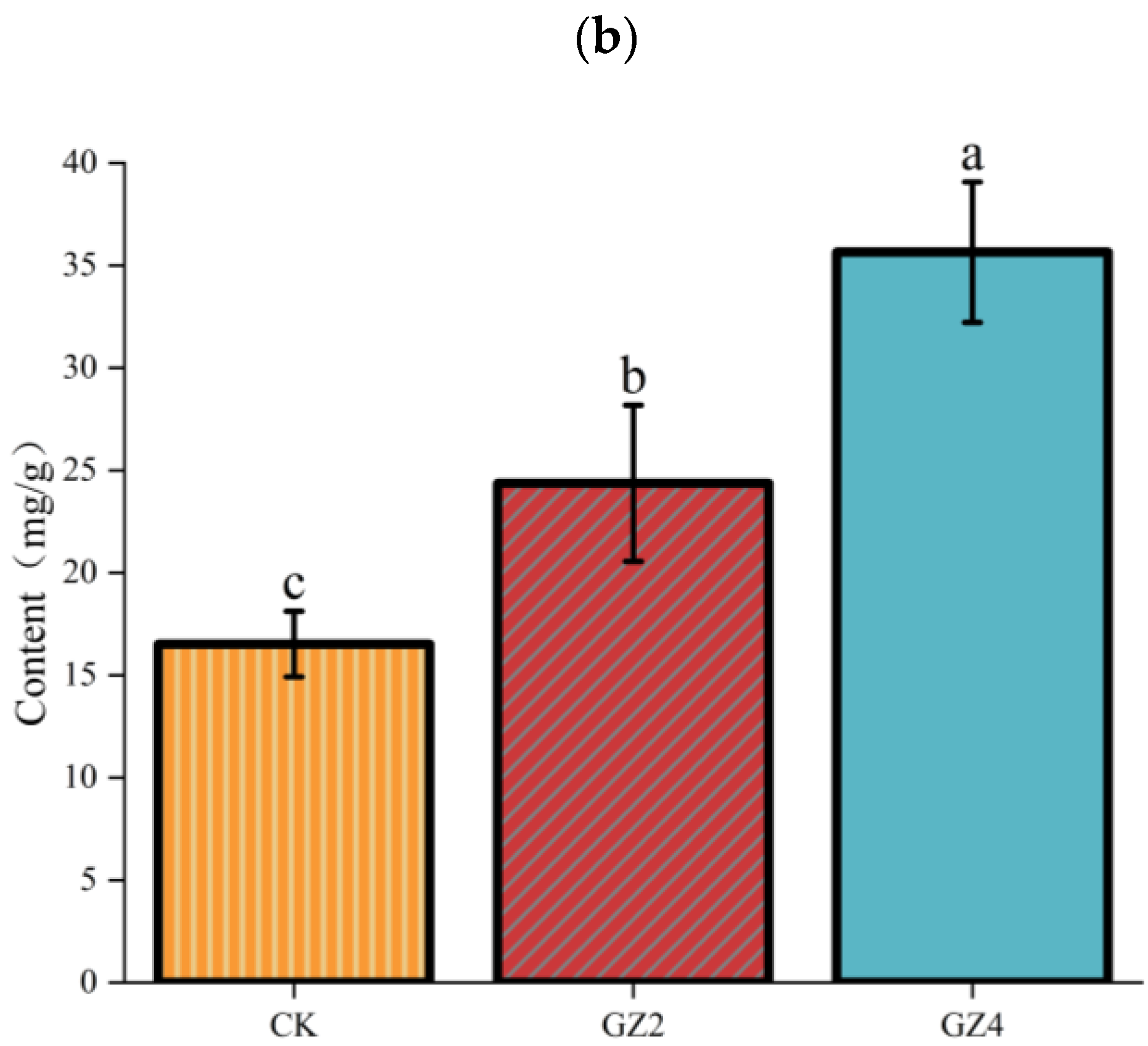
2.1. Morphological Characteristics and Total Flavonoid Content under Different Concentrations of Aluminum in C. drupifera
2.2. Comprehensive Analysis of Flavonoid Compounds
2.3. Principal Component Analysis (PCA) of C. drupifera Samples
2.4. Analysis of Differential Flavonoid Metabolites by OPLS-DA
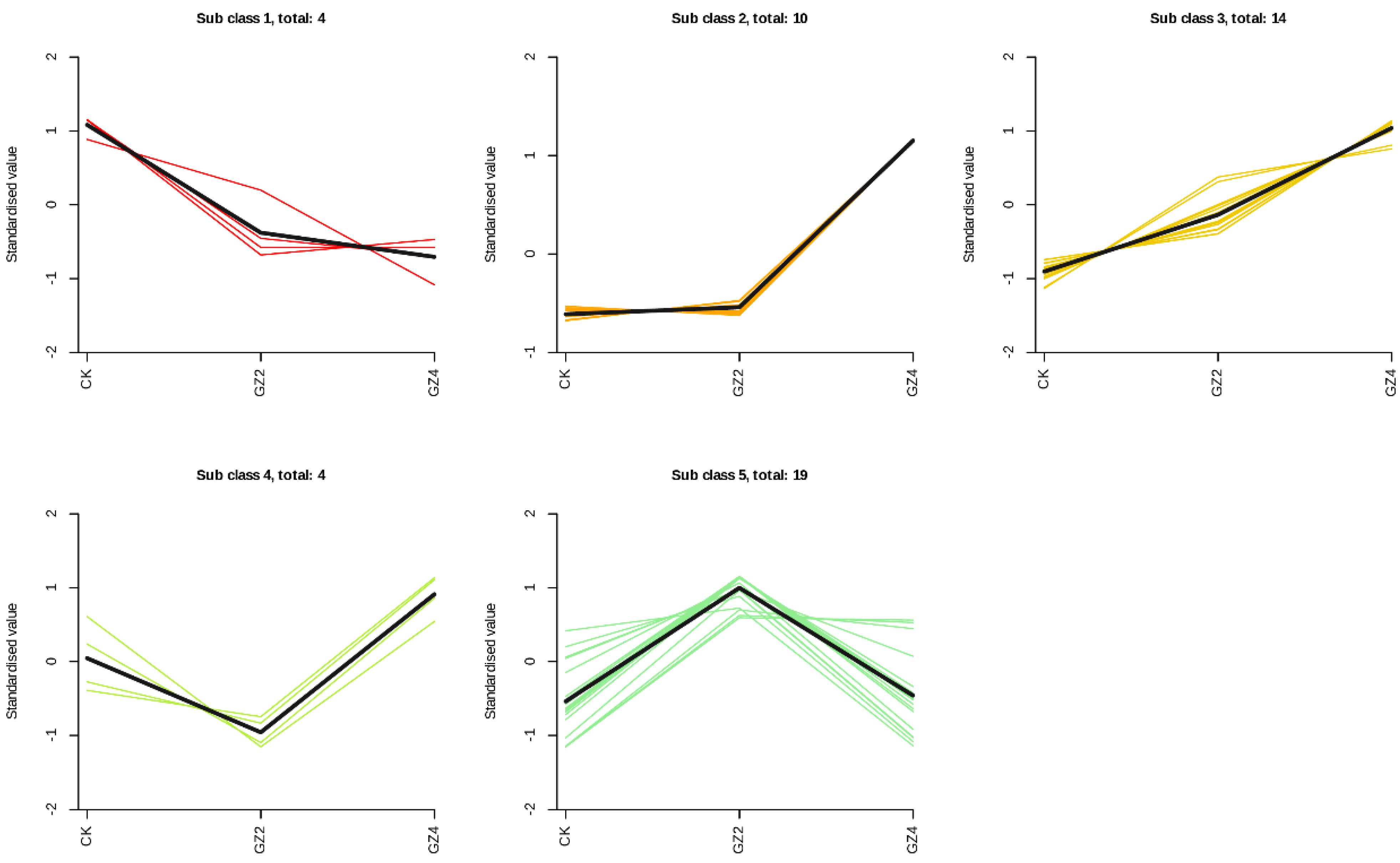
2.5. Screening and Classification of Differential Flavonoid Metabolites
2.6. Trends of Differential Flavonoid Metabolites under Aluminum Stress Treatments at Different Concentrations
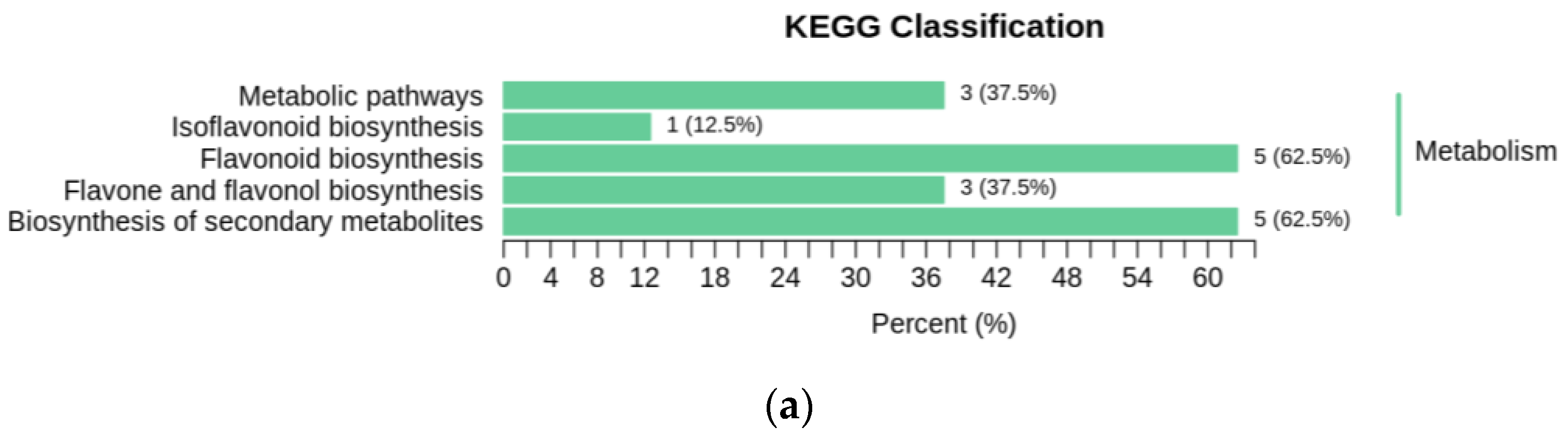
2.7. Enrichment Analysis of Differential Flavonoid Metabolites
2.8. Effect of Flavonoid Pretreatment on Antioxidant Activity
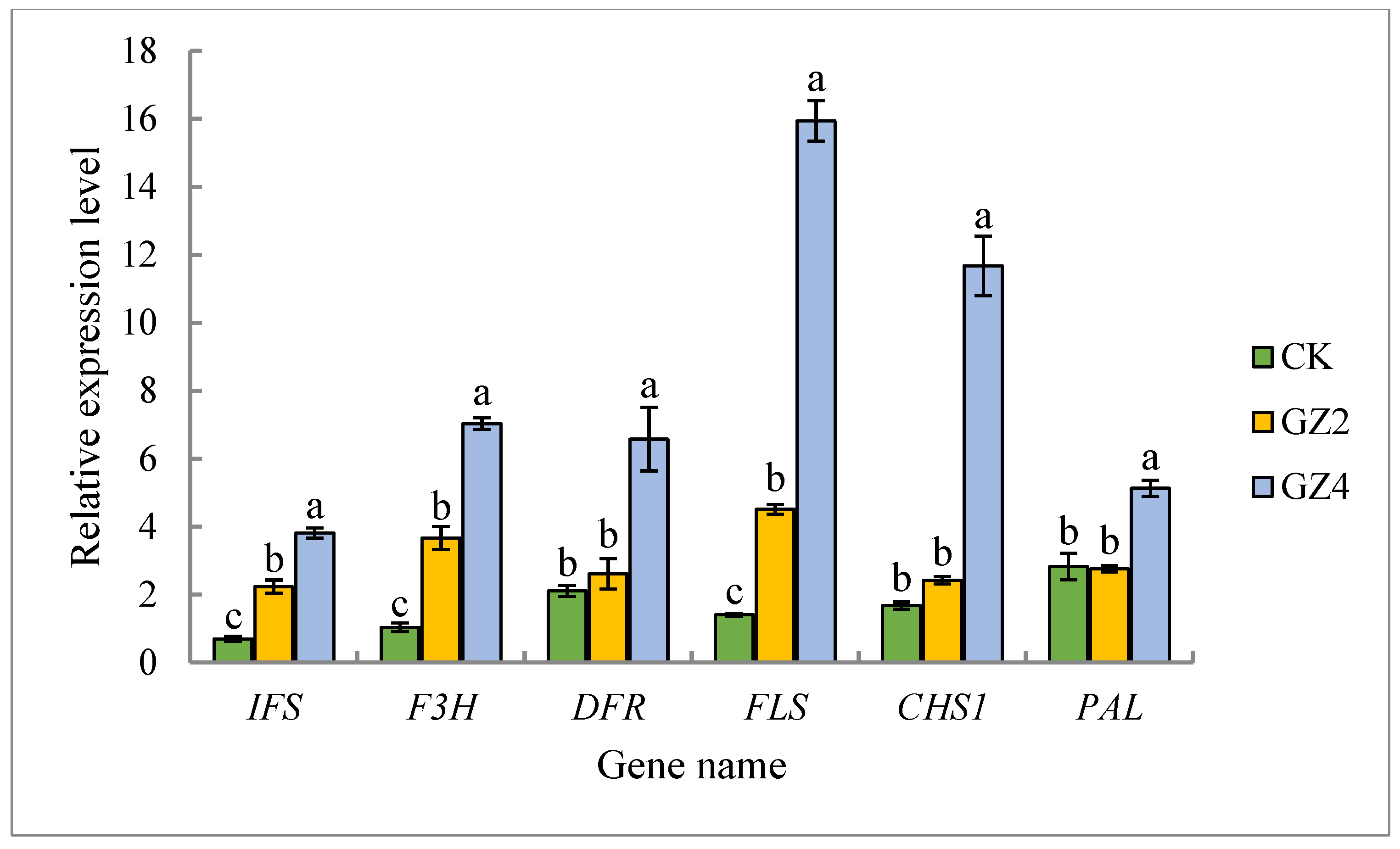
2.9. Validation of Metabolomic Data by qRT-PCR Expression Analysis of Key Flavonoid Synthesis Genes
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Treatments
4.2. Sample Preparation and Extraction
4.3. UPLC Conditions
4.4. ESI-MS/MS Conditions
4.5. Multivariate Statistical Analysis
4.6. Antioxidative Activity
4.7. Quantitative Real-Time PCR (qRT-PCR)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhuang, R.L. Chinese Camellia, 2nd ed.; Forestry Publishing House: Beijing, China, 2008; pp. 11–89. [Google Scholar]
- Li, Y.; Liao, B.; Wang, Y.; Luo, H.; Wang, S.; Li, C.; Song, W.; Zhang, K.; Yang, B.; Lu, S.; et al. Transcriptome and metabolome analyses provide insights into the relevance of pericarp thickness variations in Camellia drupifera and Camellia oleifera. Front. Plant Sci. 2022, 13, 1016475. [Google Scholar] [CrossRef]
- Qin, S.; Rong, J.; Zhang, W.; Chen, J. Cultivation history of Camellia oleifera and genetic resources in the Yangtze River Basin. Biodivers. Sci. 2018, 26, 384–395. [Google Scholar] [CrossRef]
- Yao, X.H. Oil-Tea Camellia Cultivars in China, 1st ed.; Forestry Publishing House: Beijing, China, 2016; Volume 11, pp. 32–33. [Google Scholar]
- Delhaize, E.; Ryan, P.R. Aluminum Toxicity and Tolerance in Plants. Plant Physiol. 1995, 107, 315–321. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.F.; Chen, Z.C.; Shen, R.F. Molecular mechanisms of Al tolerance in gramineous plants. Plant Soil 2014, 381, 1–12. [Google Scholar] [CrossRef]
- Kouki, R.; Ayachi, R.; Ferreira, R.; Sleimi, N. Behavior of Cucumis sativus L. in presence of aluminum stress: Germination, plant growth, and antioxidant enzymes. Food Sci. Nutr. 2021, 9, 3280–3288. [Google Scholar] [CrossRef]
- Ma, L.; Yang, S. Growth and physiological response of Kandelia obovata and Bruguiera sexangula seedlings to aluminum stress. Environ. Sci. Pollut. Res. 2022, 29, 43251–43266. [Google Scholar] [CrossRef]
- Zhang, H.L. Problem Soils. In Oklahoma Soil Fertility Handbook, 6th ed.; Arnall, B., Hiner, G., Eds.; Oklahoma State University: Stillwater, OK, USA, 2017; pp. 35–56. [Google Scholar]
- Kochian, L.V.; Hoekenga, O.A.; Piñeros, M.A. How do crop plants tolerate acid soils? Mechanisms of aluminum tolerance and phosphorous efficiency. Annu. Rev. Plant Biol. 2004, 55, 459–493. [Google Scholar] [CrossRef]
- Huang, L.; Yuan, J.; Wang, H.; Tan, X.; Niu, G. Aluminum Stress Affects Growth and Physiological Characteristics in Oil Tea. Hortscience 2017, 52, 1601–1607. [Google Scholar] [CrossRef]
- Zhou, J.; Ai, Z.Z.; Wang, H.L.; Niu, G.H. Phosphorus Alleviates Aluminum Toxicity in Camellia oleifera Seedlings. Int. J. Agric. Biol. 2019, 21, 237–243. [Google Scholar]
- Qu, X.; Zhou, J.; Masabni, J.; Yuan, J. Phosphorus relieves aluminum toxicity in oil tea seedlings by regulating the metabolic profiling in the roots. Plant Physiol. Biochem. 2020, 152, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Brunetti, C.; George, R.M.; Tattini, M.; Field, K.; Davey, M.P. Metabolomics in plant environmental physiology. J. Exp. Bot. 2013, 64, 4011–4020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferdinando, M.D.; Brunetti, C.; Fini, A.; Tattini, M. Flavonoids as Antioxidants in Plants under Abiotic Stresses; Springer: New York, NY, USA, 2012; pp. 159–179. [Google Scholar]
- Qu, X.; Hu, S.; Li, T.; Zhang, J.; Wang, B.; Liu, C. Metabolomics Analysis Reveals the Differences Between Bupleurum chinense DC. and Bupleurum scorzonerifolium Willd. Front. Plant Sci. 2022, 13, 933849. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Cheng, J.; Jiang, W.; Chen, S. Metabolomics study of flavonoids in Coreopsis tinctoria of different origins by UPLC-MS/MS. PeerJ 2022, 10, e14580. [Google Scholar] [CrossRef]
- Ferreyra, M.L.F.; Rius, S.P.; Casati, P. Flavonoids: Biosynthesis, biological functions, and biotechnological applications. Front. Plant Sci. 2012, 3, 222. [Google Scholar] [CrossRef] [Green Version]
- Pan, J.; Vicente, A.R.; Martínez, G.A.; Chaves, A.R.; Civello, P.M. Combined use of UV-C irradiation and heat treatment to improve postharvest life of strawberry fruit. J. Sci. Food Agric. 2004, 84, 1831–1838. [Google Scholar] [CrossRef]
- Chutipaijit, S.; Cha-Um, S.; Sompornpailin, K. Differential accumulations of proline and flavonoids in indica rice varieties against salinity. Pak. J. Bot. 2009, 41, 2497–2506. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Fan, R.; Sun, G.; Sun, T.; Fan, Y.; Bai, S.; Guo, S.; Huang, S.; Liu, J.; Zhang, H.; et al. Flavonoids improve drought tolerance of maize seedlings by regulating the homeostasis of reactive oxygen species. Plant Soil 2021, 461, 389–405. [Google Scholar] [CrossRef]
- Rivero, R.M.; Ruiz, J.M.; García, P.C.; López-Lefebre, L.R.; Sánchez, E.; Romero, L. Resistance to cold and heat stress: Accumulation of phenolic compounds in tomato and watermelon plants. Plant Sci. 2001, 160, 315–321. [Google Scholar] [CrossRef]
- He, J.; Yao, L.; Pecoraro, L.; Liu, C.; Wang, J.; Huang, L.; Gao, W. Cold stress regulates accumulation of flavonoids and terpenoids in plants by phytohormone, transcription process, functional enzyme, and epigenetics. Crit. Rev. Biotechnol. 2022, 18, 1–18. [Google Scholar] [CrossRef]
- Kohli, S.K.; Handa, N.; Sharma, A.; Gautam, V.; Arora, S.; Bhardwaj, R.; Wijaya, L.; Alyemeni, M.N.; Ahmad, P. Interaction of 24-epibrassinolide and salicylic acid regulates pigment contents, antioxidative defense responses, and gene expression in Brassica juncea L. seedlings under Pb stress. Environ. Sci. Pollut. Res. 2018, 25, 15159–15173. [Google Scholar] [CrossRef]
- Kaur, R.; Yadav, P.; Sharma, A.; Thukral, A.K.; Kumar, V.; Kohli, S.K.; Bhardwaj, R. Castasterone and citric acid treatment restores photosynthetic attributes in Brassica juncea L. under Cd(II) toxicity. Ecotoxicol. Environ. Saf. 2017, 145, 466–475. [Google Scholar] [CrossRef]
- Izbiańska-Jankowska, K.; Arasimowicz-Jelonek, M.; Deckert, J. Phenylpropanoid pathway metabolites promote tolerance response of lupine roots to lead stress. Ecotoxicol. Environ. Saf. 2014, 110, 61–67. [Google Scholar] [CrossRef]
- Su, L.; Lv, A.; Wen, W.; Fan, N.; Li, J.; Gao, L.; Zhou, P.; An, Y. MsMYB741 is involved in alfalfa resistance to aluminum stress by regulating flavonoid biosynthesis. Plant J. 2022, 112, 756–771. [Google Scholar] [CrossRef]
- Babu, T.S.; Akhtar, T.A.; Lampi, M.A.; Tripuranthakam, S.; Dixon, D.G.; Greenberg, B.M. Similar Stress Responses are Elicited by Copper and Ultraviolet Radiation in the Aquatic Plant Lemna gibba: Implication of Reactive Oxygen Species as Common Signals. Plant Cell Physiol. 2003, 44, 1320–1329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sobkowiak, R.; Deckert, J. Proteins induced by cadmium in soybean cells. J. Plant Physiol. 2006, 163, 1203–1206. [Google Scholar] [CrossRef] [PubMed]
- Pawlak-Sprada, S.; Arasimowicz-Jelonek, M.; Podgórska, M.; Deckert, J. Activation of phenylopropanoid pathway in legume plants exposed to heavy metals. Part I. Effects of cadmium and lead on phenylalanine ammonia-lyase gene expression, enzyme activity and lignin content. Acta Biochim. Pol. 2011, 58, 211–216. [Google Scholar] [CrossRef] [Green Version]
- Pawlak-Sprada, S.; Stobiecki, M.; Deckert, J. Activation of phenylpropanoid pathway in legume plants exposed to heavy metals. Part II. Profiling of isoflavonoids and their glycoconjugates induced in roots of lupine (Lupinus luteus) seedlings treated with cadmium and lead. Acta Biochim. Pol. 2011, 58, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Wang, Y.; Ding, Z.; Fan, K.; Ma, D.; Zhang, Y.; Yin, Q. Aluminum induced physiological and proteomic responses in tea (Camellia sinensis) roots and leaves. Plant Physiol. Biochem. 2017, 115, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Li, B.Z.; Fan, R.N.; Fan, Y.T.; Liu, R.N.; Zhang, H.; Chen, T.T.; Liu, J.; Li, H.; Zhao, X.; Song, C.P. The flavonoid biosynthesis regulator PFG3 confers drought stress tolerance in plants by promoting flavonoid accumulation. Environ. Exp. Bot. 2022, 196, 104792. [Google Scholar] [CrossRef]
- Tahara, K.; Hashida, K.; Otsuka, Y.; Ohara, S.; Kojima, K.; Shinohara, K. Identification of a Hydrolyzable Tannin, Oenothein B, as an Aluminum-Detoxifying Ligand in a Highly Aluminum-Resistant Tree, Eucalyptus camaldulensis. Plant Physiol. 2013, 164, 683–693. [Google Scholar] [CrossRef] [Green Version]
- Osawa, H.; Endo, I.; Hara, Y.; Matsushima, Y.; Tange, T. Transient Proliferation of Proanthocyanidin-Accumulating Cells on the Epidermal Apex Contributes to Highly Aluminum-Resistant Root Elongation in Camphor Tree. Plant Physiol. 2010, 155, 433–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Hossain, S.; Ma, H.; Yang, Q.; Gong, X.; Yang, P.; Feng, B. Comparative Metabolomics Reveals Differences in Flavonoid Metabolites among Different Coloured Buckwheat Flowers. J. Food Compos. Anal. 2019, 85, 103335. [Google Scholar] [CrossRef]
- Fu, Z.; Jiang, X.; Li, W.-W.; Shi, Y.; Lai, S.; Zhuang, J.; Yao, S.; Liu, Y.; Hu, J.; Gao, L.; et al. Proanthocyanidin-Aluminum Complexes Improve Aluminum Resistance and Detoxification of Camellia sinensis. J. Agric. Food Chem. 2020, 68, 7861–7869. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Wang, Y.; Ding, Z.; Song, L.; Li, Y.; Ma, D.; Wang, Y.; Shen, J.; Jia, S.; Sun, H.; et al. Aluminum Induced Metabolic Responses in Two Tea Cultivars. Plant Physiol. Biochem. 2016, 101, 162–172. [Google Scholar] [CrossRef]
- Ryan, P.R.; Tyerman, S.; Sasaki, T.; Furuichi, T.; Yamamoto, Y.; Zhang, W.H.; Delhaize, E. The Identification of Aluminium-Resistance Genes Provides Opportunities for Enhancing Crop Production on Acid Soils. J. Exp. Bot. 2010, 62, 9–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, L.M.; Chan, L.I.U.; Cui, B.M.; Ning, W.A.N.G.; Zhuo, Z.H.A.O.; Zhou, L.N.; Huang, K.F.; Ding, J.Z.; Du, H.M.; Jiang, W.; et al. Transcriptomic Responses to Aluminum (Al) Atress in Maize. J. Integr. Agric. 2018, 17, 1946–1958. [Google Scholar] [CrossRef] [Green Version]
- Pourcel, L.; Routaboul, J.M.; Cheynier, V.; Lepiniec, L.; Debeaujon, I. Flavonoid Oxidation in Plants: From Biochemical Properties to Physiological Functions. Trends Plant Sci. 2007, 12, 29–36. [Google Scholar] [CrossRef]
- Brunetti, C.; Fini, A.; Sebastiani, F.; Gori, A.; Tattini, M. Modulation of Phytohormone Signaling: A Primary Function of Flavonoids in Plant-Environment Interactions. Front. Plant Sci. 2018, 9, 1042. [Google Scholar] [CrossRef] [Green Version]
- Ofei-Manu, P.; Wagatsuma, T.; Ishikawa, S.; Tawaraya, K. The Plasma Membrane Strength of the Root-tip Cells and Root Phenolic Compounds are Correiated with Al Tolerance in Several Common Woody Plants. Soil Sci. Plant Nutr. 2001, 47, 359–375. [Google Scholar] [CrossRef]
- Nagata, T.; Hayatsu, M.; Kosuge, N. Identification of Aluminium Forms in Tea Leaves by 27Al NMR. Phytochemistry 1992, 31, 1215–1218. [Google Scholar] [CrossRef]
- Kidd, P.S.; Llugany, M.; Poschenrieder, C.; Gunsé, B.; Barceló, J. The Role of Root Exudates in Aluminium Resistance and Siliconinduced Amelioration of Aluminium Toxicity in Three Varieties of Maize (Zea mays L.). J. Exp. Bot. 2001, 52, 1339–1352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mutha, R.E.; Tatiya, A.U.; Surana, S.J. Flavonoids as Natural Phenolic Compounds and their Role in Therapeutics: An Overview. Future J. Pharm. Sci. 2021, 7, 25. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Ming, R.; Yao, S.; Li, L.; Huang, R.; Tan, Y. Identification of Anthocyanins in the Fruits of Kadsura coccinea using UPLC-MS/MS-based Metabolomics. Biochem. Syst. Ecol. 2021, 98, 104324. [Google Scholar] [CrossRef]
- Zhang, Y.; Gao, J.; Qie, Q.; Yang, Y.; Hou, S.; Wang, X.; Li, X.; Han, Y. Comparative Analysis of Flavonoid Metabolites in Foxtail Millet (Setaria italica) with Different Eating Quality. Life 2021, 11, 578. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Teng, J.; Zhu, Y.; Xie, F.; Hou, J.; Ling, Y.; Zhu, H. Metabolomics Study of Flavonoids of Taxilluschinensis on Different Hosts Using UPLC-ESI-MS/MS. Molecules 2021, 26, 7681. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.; Wen, Z.; Tang, T.; Liu, Y.; Dang, F.; Xie, T.; Wu, H. Study on Flavonoid and Bioactivity Features of the Pericarp of Citri Reticulatae ‘chachi’ during Storage. Arab. J. Chem. 2021, 15, 103653. [Google Scholar] [CrossRef]
- Martínez-Villaluenga, C.; Zieliński, H.; Frias, J.; Piskula, M.K.; Kozłowska, H.; Vidal-Valverde, C. Antioxidant Capacity and Polyphenolic Content of High-protein Lupin Products. Food Chem. 2009, 112, 84–88. [Google Scholar] [CrossRef]
- Molynex, P. The use of the stable free radical diphenylpicrylhydrazyl (DPPH) for estimating antioxidant activity. J. Sci. Technol. 2004, 26, 211–219. [Google Scholar]
- Singh, P.; Singh, R.K.; Song, Q.-Q.; Li, H.-B.; Guo, D.-J.; Malviya, M.K.; Verma, K.K.; Song, X.-P.; Lakshmanan, P.; Yang, L.T.; et al. Comparative Analysis of Protein and Differential Responses of Defense-related Gene and Enzyme Activity Reveals the Long-term Molecular Responses of Sugarcane Inoculated with Sporisorium scitamineum. J. Plant Interact. 2021, 16, 12–29. [Google Scholar] [CrossRef]
- Wang, Y.; Jiang, W.; Cheng, J.S.; He, X.G.; Chu, G.H. Differential Analysis of Flavonoid Metabolites and Key Enzyme Gene Ex-pressions in Coreopsis tinctoria Collected from Plateau and Plain Areas. J. Food Saf. Food Qual. 2022, 13, 6956–6963. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | Compounds | Molecular Weight | Formula | VIP | p-Value | Trends | |
|---|---|---|---|---|---|---|---|
| GZ2/CK | GZ4/CK | ||||||
| Flavonoid_22 | Neohesperidin dihydrochalcone | 612.21 | C28H36O15 | 1.20 | 0.0018 | Up | Up |
| Flavonoid_80 | (−)-Gallocatechin gallate | 458.08 | C22H18O11 | 1.17 | 0.0236 | Down | Down |
| Flavonoid_78 | Eriocitrin | 596.17 | C27H32O15 | 1.22 | 0.0005 | Up | Up |
| Flavonoid_101 | Hesperidin | 610.19 | C28H34O15 | 1.21 | 0.0262 | Down | Down |
| Flavonoid_48 | Narirutin | 580.18 | C27H32O14 | 1.22 | 0.0015 | Up | Up |
| Flavonoid_176 | Naringenin-7-glucoside | 434.12 | C21H22O10 | 1.19 | 0.0003 | Up | Up |
| Flavonoid_141 | Isosakuranetin | 448.14 | C22H24O10 | 1.11 | 0.0078 | Up | Up |
| Flavonoid_01 | Astilbin | 450.12 | C21H22O11 | 1.20 | 0.0064 | Up | Up |
| Flavonoid_139 | Dihydrokaempferol | 288.06 | C15H12O6 | 1.22 | 0.0021 | Up | Up |
| Flavonoid_56 | Taxifolin | 304.06 | C15H12O7 | 1.21 | 0.0000 | Up | Up |
| Flavonoid_64 | Scutellarein | 286.05 | C15H10O6 | 1.22 | 0.0031 | Up | Up |
| Flavonoid_90 | Cynaroside | 448.10 | C21H20O11 | 1.21 | 0.0140 | Up | Up |
| Flavonoid_67 | Fisetin | 286.05 | C15H10O6 | 1.04 | 0.0205 | Up | Up |
| Flavonoid_194 | Quercimeritrin | 464.10 | C21H20O12 | 1.18 | 0.0057 | Up | Up |
| Flavonoid_02 | Miquelianin | 478.07 | C21H18O13 | 1.21 | 0.0020 | Down | Down |
| Flavonoid_119 | Astragalin | 448.10 | C21H20O11 | 1.21 | 0.0042 | Up | Up |
| Flavonoid_180 | Spiraeoside | 464.10 | C21H20O12 | 1.21 | 0.0031 | Up | Up |
| Flavonoid_152 | Ononin | 430.13 | C22H22O9 | 1.20 | 0.0034 | Up | Up |
| Flavonoid_114 | 6′-O-Acetylglycitin | 488.13 | C24H24O11 | 1.21 | 0.1004 | Up | Up |
| Flavonoid_102 | Theaflavin | 564.13 | C29H24O12 | 1.22 | 0.0013 | Down | - |
| Flavonoid_150 | (-)-Catechin gallate | 442.09 | C22H18O10 | 1.13 | 0.0360 | Up | - |
| Flavonoid_108 | Pinocembrin | 256.07 | C15H12O4 | 1.12 | 0.0866 | Up | - |
| Flavonoid_158 | Isosakuranetin | 286.08 | C16H14O5 | 1.21 | 0.0044 | Up | - |
| Flavonoid_179 | Poncirin | 594.19 | C28H34O14 | 1.21 | 0.0040 | Up | - |
| Flavonoid_204 | Linarin | 592.18 | C28H32O14 | 1.22 | 0.0008 | Up | - |
| Flavonoid_50 | Luteolin | 286.05 | C15H10O6 | 1.19 | 0.0089 | Up | - |
| Flavonoid_118 | Narcissin | 624.17 | C28H32O16 | 1.21 | 0.0014 | Down | - |
| Flavonoid_86 | Avicularin | 434.08 | C20H18O11 | 1.20 | 0.0007 | Up | - |
| Flavonoid_138 | Baimaside | 626.15 | C27H30O17 | 1.21 | 0.0009 | Up | - |
| Flavonoid_137 | Tiliroside | 594.14 | C30H26O13 | 1.21 | 0.0001 | Up | - |
| Flavonoid_65 | Phlorizin | 436.14 | C21H24O10 | 1.05 | 0.0736 | - | Down |
| Flavonoid_201 | Trilobatin | 436.14 | C21H24O10 | 1.15 | 0.0123 | - | Up |
| Flavonoid_177 | Phloretin | 274.08 | C15H14O5 | 1.15 | 0.0027 | - | Down |
| Flavonoid_191 | Naringenin chalcone | 272.07 | C15H12O5 | 1.16 | 0.0000 | - | Up |
| Flavonoid_55 | (−)-Catechin | 290.08 | C15H14O6 | 1.15 | 0.0001 | - | Up |
| Flavonoid_147 | (−)-Epicatechin | 290.08 | C15H14O6 | 1.16 | 0.0016 | - | Up |
| Flavonoid_195 | (−)-Gallocatechin | 306.07 | C15H14O7 | 1.03 | 0.1439 | - | Down |
| Flavonoid_126 | Eriodictyol | 288.06 | C15H12O6 | 1.16 | 0.0021 | - | Up |
| Flavonoid_124 | Silychristin | 482.12 | C25H22O10 | 1.16 | 0.0000 | - | Up |
| Flavonoid_40 | Dihydromyricetin | 320.05 | C15H12O8 | 1.01 | 0.0187 | - | Up |
| Flavonoid_117 | Engeletin | 434.12 | C21H22O10 | 1.15 | 0.0005 | - | Up |
| Flavonoid_154 | Vitexin | 432.11 | C21H20O10 | 1.13 | 0.0236 | - | Up |
| Flavonoid_115 | Quercitrin | 448.10 | C21H20O11 | 1.16 | 0.0001 | - | Up |
| Flavonoid_106 | Typhaneoside | 770.23 | C34H42O20 | 1.08 | 0.0312 | - | Up |
| Flavonoid_57 | Rutin | 610.15 | C27H30O16 | 1.16 | 0.0000 | - | Up |
| Flavonoid_197 | Isorhamnetin 3-O-glucoside | 478.11 | C22H22O12 | 1.16 | 0.0000 | - | Up |
| Flavonoid_43 | Kaempferol | 286.05 | C15H10O6 | 1.16 | 0.0464 | - | Down |
| Flavonoid_175 | Afzelin | 432.11 | C21H20O10 | 1.16 | 0.0000 | - | Up |
| Flavonoid_73 | Genistin | 432.11 | C21H20O10 | 1.02 | 0.0151 | - | Up |
| Genes | Upstream Primers (5′→3′) | Downstream Primers (5′→3′) | Reference |
|---|---|---|---|
| IFS | ACAACGGCGGAACATACG | ACACTGCTTGCCACTCACC | [54] |
| F3H | AAGAAGTGGAGCAAGGGAAAG | CGGAGACATTGGTGGAGAAA | [54] |
| DFR | ATGTTGCTACACCGTGGTTAC | AAATCGAAAGATCCCTCCTC | [54] |
| FLS | GCACATGATGCAACGCTACT | GGAACCTCTAATTGGGTCCCTC | [27] |
| CHS1 | GCCAAGAGTGGTGTGGATAGG | TGCCCTTTGAGCATTGCGAA | [27] |
| PAL | GGAGGATCAACTCCACATAGC | AGTCACTGCTGGCCTTAACTC | [27] |
| β-Tubulin | ATGTTCAGGCGCAAGGCTT | TCTGCAACCGGGTCATTCAT | [54] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Cheng, J.; Wei, S.; Jiang, W.; Li, Y.; Guo, W.; Dai, W.; Liao, B. Metabolomic Study of Flavonoids in Camellia drupifera under Aluminum Stress by UPLC-MS/MS. Plants 2023, 12, 1432. https://doi.org/10.3390/plants12071432
Wang Y, Cheng J, Wei S, Jiang W, Li Y, Guo W, Dai W, Liao B. Metabolomic Study of Flavonoids in Camellia drupifera under Aluminum Stress by UPLC-MS/MS. Plants. 2023; 12(7):1432. https://doi.org/10.3390/plants12071432
Chicago/Turabian StyleWang, Yi, Junsen Cheng, Shanglin Wei, Wei Jiang, Yongquan Li, Wei Guo, Wenkui Dai, and Boyong Liao. 2023. "Metabolomic Study of Flavonoids in Camellia drupifera under Aluminum Stress by UPLC-MS/MS" Plants 12, no. 7: 1432. https://doi.org/10.3390/plants12071432
APA StyleWang, Y., Cheng, J., Wei, S., Jiang, W., Li, Y., Guo, W., Dai, W., & Liao, B. (2023). Metabolomic Study of Flavonoids in Camellia drupifera under Aluminum Stress by UPLC-MS/MS. Plants, 12(7), 1432. https://doi.org/10.3390/plants12071432