
Unlocking the Secret to Higher Crop Yield: The Potential for Histone Modifications




Abstract
:1. Introduction
2. Histone Modifications
2.1. Histone Methylation
2.2. Histone Acetylation
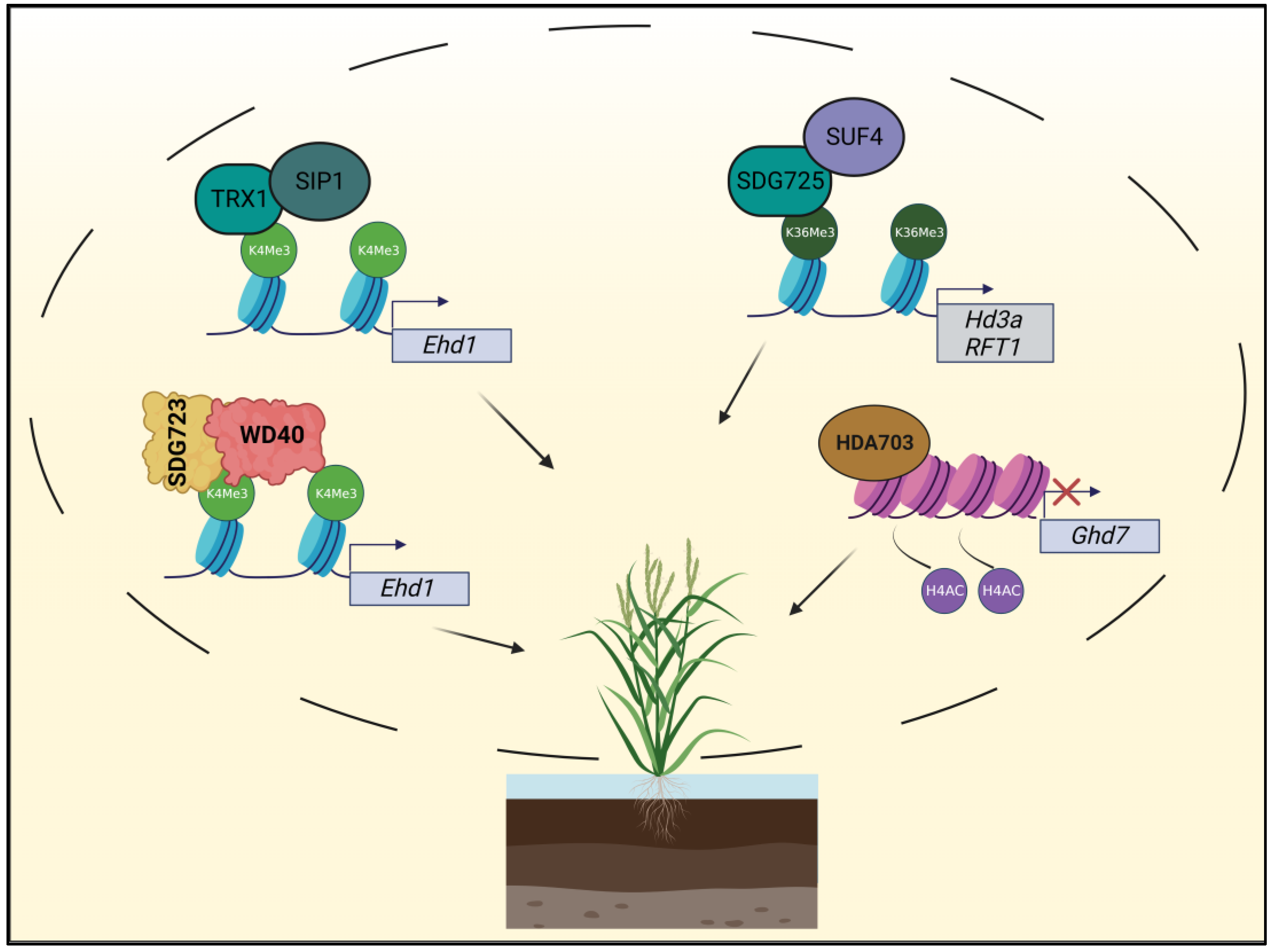
3. Role of Histone Modifications in Improving Yield in Rice (Oryza sativa L.)
3.1. Flowering
3.2. Shoot Development and Grain Formation
3.3. Abiotic Stress
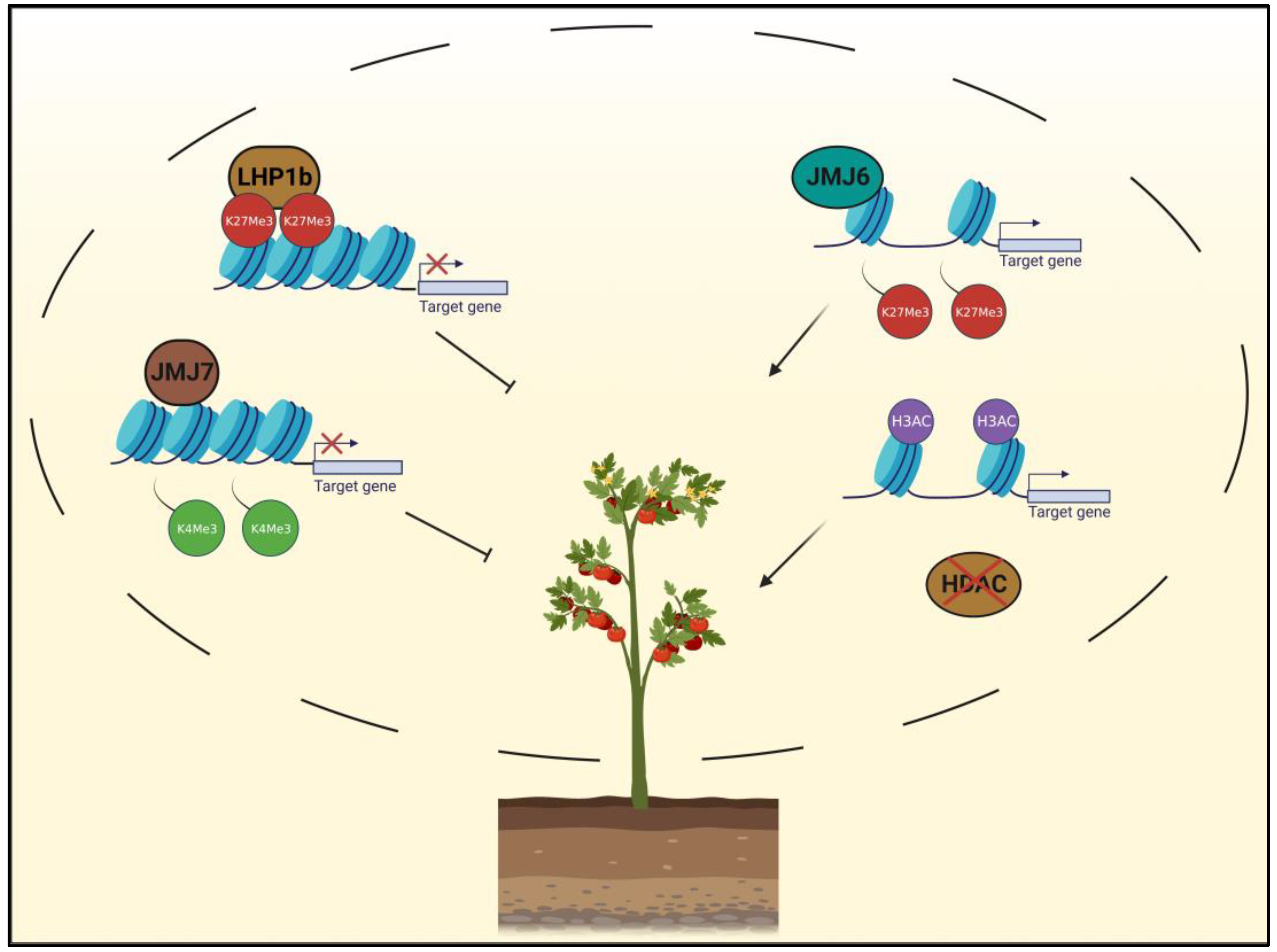
4. Role of Histone Modifications in Improving Fruit Quality in Tomato (Solanum lycopersicum)
4.1. Fruit Ripening
4.2. Abiotic Stress
5. Epigenetic Engineering in Crop Improvement
Epi-Breeding and CRISPR-Cas9 Applications
6. Conclusions and Future Perspectives
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Duan, C.-G.; Zhu, J.-K.; Cao, X. Retrospective and Perspective of Plant Epigenetics in China. J. Genet. Genom. 2018, 45, 621–638. [Google Scholar] [CrossRef]
- Chang, Y.-N.; Zhu, C.; Jiang, J.; Zhang, H.; Zhu, J.-K.; Duan, C.-G. Epigenetic Regulation in Plant Abiotic Stress Responses. J. Integr. Plant Biol. 2020, 62, 563–580. [Google Scholar] [CrossRef]
- Tang, D.; Gallusci, P.; Lang, Z. Fruit Development and Epigenetic Modifications. New Phytol. 2020, 228, 839–844. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-M.; To, T.K.; Nishioka, T.; Seki, M. Chromatin Regulation Functions in Plant Abiotic Stress Responses. Plant Cell Environ. 2010, 33, 604–611. [Google Scholar] [CrossRef]
- Gallo-Franco, J.J.; Sosa, C.C.; Ghneim-Herrera, T.; Quimbaya, M. Epigenetic Control of Plant Response to Heavy Metal Stress: A New View on Aluminum Tolerance. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Pfluger, J.; Wagner, D. Histone Modifications and Dynamic Regulation of Genome Accessibility in Plants. Curr. Opin. Plant Biol. 2007, 10, 645–652. [Google Scholar] [CrossRef]
- Berger, S.L. The Complex Language of Chromatin Regulation during Transcription. Nature 2007, 447, 407–412. [Google Scholar] [CrossRef] [PubMed]
- Karanthamalai, J.; Chodon, A.; Chauhan, S.; Pandi, G. DNA N6-Methyladenine Modification in Plant Genomes—A Glimpse into Emerging Epigenetic Code. Plants 2020, 9, 247. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Song, H.; Wei, Y.; Li, P.; Hu, N.; Liu, J.; Zhang, B.; Peng, R. High Throughput Deep Sequencing Elucidates the Important Role of LncRNAs in Foxtail Millet Response to Herbicides. Genomics 2020, 112, 4463–4473. [Google Scholar] [CrossRef]
- Czajka, K.; Mehes-Smith, M.; Nkongolo, K. DNA Methylation and Histone Modifications Induced by Abiotic Stressors in Plants. Genes Genom. 2022, 44, 279–297. [Google Scholar] [CrossRef]
- Gibney, E.R.; Nolan, C.M. Epigenetics and Gene Expression. Heredity 2010, 105, 4–13. [Google Scholar] [CrossRef]
- Shi, J.; Dong, A.; Shen, W.H. Epigenetic Regulation of Rice Flowering and Reproduction. Front. Plant Sci. 2015, 5, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Gallusci, P.; Hodgman, C.; Teyssier, E.; Seymour, G.B. DNA Methylation and Chromatin Regulation during Fleshy Fruit Development and Ripening. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef]
- Luger, K.; Rechsteiner, T.J.; Flaus, A.J.; Waye, M.M.Y.; Richmond, T.J. Characterization of Nucleosome Core Particles Containing Histone Proteins Made in Bacteria11Edited by A. Klug. J. Mol. Biol. 1997, 272, 301–311. [Google Scholar] [CrossRef] [PubMed]
- Bannister, A.J.; Kouzarides, T. Regulation of Chromatin by Histone Modifications. Cell Res. 2011, 21, 381–395. [Google Scholar] [CrossRef]
- Berr, A.; Shafiq, S.; Shen, W.-H. Histone Modifications in Transcriptional Activation during Plant Development. Biochim. Biophys. Acta (BBA)-Gene Regul. Mech. 2011, 1809, 567–576. [Google Scholar] [CrossRef]
- Bowler, C.; Benvenuto, G.; Laflamme, P.; Molino, D.; Probst, A.V.; Tariq, M.; Paszkowski, J. Chromatin Techniques for Plant Cells. Plant J. 2004, 39, 776–789. [Google Scholar] [CrossRef] [PubMed]
- Li, Y. Modern Epigenetics Methods in Biological Research. Methods 2021, 187, 104–113. [Google Scholar] [CrossRef]
- Ouyang, W.; Xiong, D.; Li, G.; Li, X. Unraveling the 3D Genome Architecture in Plants: Present and Future. Mol. Plant 2020, 13, 1676–1693. [Google Scholar] [CrossRef]
- Pei, L.; Li, G.; Lindsey, K.; Zhang, X.; Wang, M. Plant 3D Genomics: The Exploration and Application of Chromatin Organization. New Phytol. 2021, 230, 1772–1786. [Google Scholar] [CrossRef]
- Kaiserli, E.; Perrella, G.; Davidson, M.L.H. Light and Temperature Shape Nuclear Architecture and Gene Expression. Curr. Opin. Plant Biol. 2018, 45, 103–111. [Google Scholar] [CrossRef]
- Liu, C.; Cheng, Y.-J.; Wang, J.-W.; Weigel, D. Prominent Topologically Associated Domains Differentiate Global Chromatin Packing in Rice from Arabidopsis. Nat. Plants 2017, 3, 742–748. [Google Scholar] [CrossRef]
- Golicz, A.A.; Bhalla, P.L.; Edwards, D.; Singh, M.B. Rice 3D Chromatin Structure Correlates with Sequence Variation and Meiotic Recombination Rate. Commun. Biol. 2020, 3, 235. [Google Scholar] [CrossRef] [PubMed]
- Mumbach, M.R.; Rubin, A.J.; Flynn, R.A.; Dai, C.; Khavari, P.A.; Greenleaf, W.J.; Chang, H.Y. HiChIP: Efficient and Sensitive Analysis of Protein-Directed Genome Architecture. Nat. Methods 2016, 13, 919–922. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Xie, W. The Role of 3D Genome Organization in Development and Cell Differentiation. Nat. Rev. Mol. Cell Biol. 2019, 20, 535–550. [Google Scholar] [CrossRef] [PubMed]
- Grob, S.; Grossniklaus, U. Chromosome Conformation Capture-Based Studies Reveal Novel Features of Plant Nuclear Architecture. Curr. Opin. Plant Biol. 2017, 36, 149–157. [Google Scholar] [CrossRef]
- Schwope, R.; Magris, G.; Miculan, M.; Paparelli, E.; Celii, M.; Tocci, A.; Marroni, F.; Fornasiero, A.; De Paoli, E.; Morgante, M. Open Chromatin in Grapevine Marks Candidate CREs and with Other Chromatin Features Correlates with Gene Expression. Plant J. 2021, 107, 1631–1647. [Google Scholar] [CrossRef]
- Wang, X.; Chen, C.; He, C.; Chen, D.; Yan, W. Mapping Open Chromatin by ATAC-Seq in Bread Wheat. Front. Plant Sci. 2022, 13, 1074873. [Google Scholar] [CrossRef]
- Liu, C.; Lu, F.; Cui, X.; Cao, X. Histone Methylation in Higher Plants. Annu. Rev. Plant Biol. 2010, 61, 395–420. [Google Scholar] [CrossRef]
- Chachar, S.; Chachar, M.; Riaz, A.; Shaikh, A.A.; Li, X.; Li, X.; Guan, C.; Zhang, P. Epigenetic Modification for Horticultural Plant Improvement Comes of Age. Sci. Hortic. 2022, 292, 110633. [Google Scholar] [CrossRef]
- Shin, H.; Choi, W.L.; Lim, J.Y.; Huh, J.H. Epigenome Editing: Targeted Manipulation of Epigenetic Modifications in Plants. Genes Genom. 2022, 44, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Wei, W.; Zhou, D.-X. Histone Acetylation Enzymes Coordinate Metabolism and Gene Expression. Trends Plant Sci. 2015, 20, 614–621. [Google Scholar] [CrossRef]
- Pandey, R.; Müller, A.; Napoli, C.A.; Selinger, D.A.; Pikaard, C.S.; Richards, E.J.; Bender, J.; Mount, D.W.; Jorgensen, R.A. Analysis of Histone Acetyltransferase and Histone Deacetylase Families of Arabidopsis Thaliana Suggests Functional Diversification of Chromatin Modification among Multicellular Eukaryotes. Nucleic Acids Res. 2002, 30, 5036–5055. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.-E.; Hu, Z.; Zhu, M.; Li, F.; Zhu, Z.; Lu, Y.; Chen, G. The Tomato Histone Deacetylase SlHDA1 Contributes to the Repression of Fruit Ripening and Carotenoid Accumulation. Sci. Rep. 2017, 7, 7930. [Google Scholar] [CrossRef]
- Kurdistani, S.K.; Grunstein, M. Histone Acetylation and Deacetylation in Yeast. Nat. Rev. Mol. Cell Biol. 2003, 4, 276–284. [Google Scholar] [CrossRef]
- Rossetto, D.; Avvakumov, N.; Côté, J. Histone Phosphorylation: A Chromatin Modification Involved in Diverse Nuclear Events. Epigenetics 2012, 7, 1098–1108. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Yan, Q. Histone Ubiquitination and Deubiquitination in Transcription, DNA Damage Response, and Cancer. Front. Oncol. 2012, 2, 26. [Google Scholar] [CrossRef]
- Han, D.; Chen, C.; Xia, S.; Liu, J.; Shu, J.; Nguyen, V.; Lai, J.; Cui, Y.; Yang, C. Chromatin-Associated SUMOylation Controls the Transcriptional Switch between Plant Development and Heat Stress Responses. Plant Commun. 2021, 2, 100091. [Google Scholar] [CrossRef]
- Hauser, M.-T.; Aufsatz, W.; Jonak, C.; Luschnig, C. Transgenerational Epigenetic Inheritance in Plants. Biochim. Biophys. Acta (BBA)-Gene Regul. Mech. 2011, 1809, 459–468. [Google Scholar] [CrossRef]
- Jiang, P.; Wang, S.; Zheng, H.; Li, H.; Zhang, F.; Su, Y.; Xu, Z.; Lin, H.; Qian, Q.; Ding, Y. SIP1 Participates in Regulation of Flowering Time in Rice by Recruiting OsTrx1 to Ehd1. New Phytol. 2018, 219, 422–435. [Google Scholar] [CrossRef] [PubMed]
- Jiang, P.; Wang, S.; Jiang, H.; Cheng, B.; Wu, K.; Ding, Y. The COMPASS-Like Complex Promotes Flowering and Panicle Branching in Rice. Plant Physiol. 2018, 176, 2761–2771. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Liu, Y.; Wang, B.; Luo, Q.; Shi, J.; Gan, J.; Shen, W.-H.; Yu, Y.; Dong, A. The Transcription Factor OsSUF4 Interacts with SDG725 in Promoting H3K36me3 Establishment. Nat. Commun. 2019, 10, 2999. [Google Scholar] [CrossRef]
- Wang, H.; Jiao, X.; Kong, X.; Liu, Y.; Chen, X.; Fang, R.; Yan, Y. The Histone Deacetylase HDA703 Interacts with OsBZR1 to Regulate Rice Brassinosteroid Signaling, Growth and Heading Date through Repression of Ghd7 Expression. Plant J. 2020, 104, 447–459. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Liu, Y.; Shen, W.-H.; Yu, Y.; Dong, A. Chromatin-Remodeling Factor OsINO80 Is Involved in Regulation of Gibberellin Biosynthesis and Is Crucial for Rice Plant Growth and Development. J. Integr. Plant Biol. 2018, 60, 144–159. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.; Tan, F.; Lu, Y.; Liu, X.; Li, T.; Yuan, W.; Zhao, Y.; Zhou, D.-X. WOX11 Recruits a Histone H3K27me3 Demethylase to Promote Gene Expression during Shoot Development in Rice. Nucleic Acids Res. 2018, 46, 2356–2369. [Google Scholar] [CrossRef]
- Wu, K.; Wang, S.; Song, W.; Zhang, J.; Wang, Y.; Liu, Q.; Yu, J.; Ye, Y.; Li, S.; Chen, J.; et al. Enhanced Sustainable Green Revolution Yield via Nitrogen-Responsive Chromatin Modulation in Rice. Science 2020, 367, eaaz2046. [Google Scholar] [CrossRef]
- Yang, J.; Cho, L.-H.; Yoon, J.; Yoon, H.; Wai, A.H.; Hong, W.-J.; Han, M.; Sakakibara, H.; Liang, W.; Jung, K.-H.; et al. Chromatin Interacting Factor OsVIL2 Increases Biomass and Rice Grain Yield. Plant Biotechnol. J. 2019, 17, 178–187. [Google Scholar] [CrossRef]
- Huang, Y.; Bai, X.; Cheng, N.; Xiao, J.; Li, X.; Xing, Y. Wide Grain 7 Increases Grain Width by Enhancing H3K4me3 Enrichment in the OsMADS1 Promoter in Rice (Oryza Sativa L.). Plant J. 2020, 102, 517–528. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Zhang, N.; Wang, W.-Q.; Shen, S.-Y.; Bai, C.; Song, X.-J. The Ubiquitin-Interacting Motif-Type Ubiquitin Receptor HDR3 Interacts with and Stabilizes the Histone Acetyltransferase GW6a to Control the Grain Size in Rice. Plant Cell 2021, 33, 3331–3347. [Google Scholar] [CrossRef]
- Ullah, F.; Xu, Q.; Zhao, Y.; Zhou, D.-X. Histone Deacetylase HDA710 Controls Salt Tolerance by Regulating ABA Signaling in Rice. J. Integr. Plant Biol. 2021, 63, 451–467. [Google Scholar] [CrossRef]
- Cheng, X.; Zhang, S.; Tao, W.; Zhang, X.; Liu, J.; Sun, J.; Zhang, H.; Pu, L.; Huang, R.; Chen, T. INDETERMINATE SPIKELET1 Recruits Histone Deacetylase and a Transcriptional Repression Complex to Regulate Rice Salt Tolerance. Plant Physiol. 2018, 178, 824–837. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Du, K.; Shi, Y.; Yin, L.; Shen, W.-H.; Yu, Y.; Liu, B.; Dong, A. H3K36 Methyltransferase SDG708 Enhances Drought Tolerance by Promoting Abscisic Acid Biosynthesis in Rice. New Phytol. 2021, 230, 1967–1984. [Google Scholar] [CrossRef] [PubMed]
- Song, T.; Zhang, Q.; Wang, H.; Han, J.; Xu, Z.; Yan, S.; Zhu, Z. OsJMJ703, a Rice Histone Demethylase Gene, Plays Key Roles in Plant Development and Responds to Drought Stress. Plant Physiol. Biochem. 2018, 132, 183–188. [Google Scholar] [CrossRef]
- Li, Z.; Jiang, G.; Liu, X.; Ding, X.; Zhang, D.; Wang, X.; Zhou, Y.; Yan, H.; Li, T.; Wu, K.; et al. Histone Demethylase SlJMJ6 Promotes Fruit Ripening by Removing H3K27 Methylation of Ripening-Related Genes in Tomato. New Phytol. 2020, 227, 1138–1156. [Google Scholar] [CrossRef]
- Liang, Q.; Deng, H.; Li, Y.; Liu, Z.; Shu, P.; Fu, R.; Zhang, Y.; Pirrello, J.; Zhang, Y.; Grierson, D.; et al. Like Heterochromatin Protein 1b Represses Fruit Ripening via Regulating the H3K27me3 Levels in Ripening-Related Genes in Tomato. New Phytol. 2020, 227, 485–497. [Google Scholar] [CrossRef]
- Wang, J.; Li, G.; Li, C.; Zhang, C.; Cui, L.; Ai, G.; Wang, X.; Zheng, F.; Zhang, D.; Larkin, R.M.; et al. NF-Y Plays Essential Roles in Flavonoid Biosynthesis by Modulating Histone Modifications in Tomato. New Phytol. 2021, 229, 3237–3252. [Google Scholar] [CrossRef]
- Ding, X.; Liu, X.; Jiang, G.; Li, Z.; Song, Y.; Zhang, D.; Jiang, Y.; Duan, X. SlJMJ7 Orchestrates Tomato Fruit Ripening via Crosstalk between H3K4me3 and DML2-Mediated DNA Demethylation. New Phytol. 2022, 233, 1202–1219. [Google Scholar] [CrossRef]
- Guo, J.-E.; Hu, Z.; Yu, X.; Li, A.; Li, F.; Wang, Y.; Tian, S.; Chen, G. A Histone Deacetylase Gene, SlHDA3, Acts as a Negative Regulator of Fruit Ripening and Carotenoid Accumulation. Plant Cell Rep. 2018, 37, 125–135. [Google Scholar] [CrossRef]
- Guo, J.-E. Histone Deacetylase Gene SlHDT1 Regulates Tomato Fruit Ripening by Affecting Carotenoid Accumulation and Ethylene Biosynthesis. Plant Sci. 2022, 318, 111235. [Google Scholar] [CrossRef] [PubMed]
- Deng, H.; Chen, Y.; Liu, Z.; Liu, Z.; Shu, P.; Wang, R.; Hao, Y.; Su, D.; Pirrello, J.; Liu, Y.; et al. SlERF.F12 Modulates the Transition to Ripening in Tomato Fruit by Recruiting the Co-Repressor TOPLESS and Histone Deacetylases to Repress Key Ripening Genes. Plant Cell 2022, 34, 1250–1272. [Google Scholar] [CrossRef]
- Cao, X.; Wei, C.; Duan, W.; Gao, Y.; Kuang, J.; Liu, M.; Chen, K.; Klee, H.; Zhang, B. Transcriptional and Epigenetic Analysis Reveals That NAC Transcription Factors Regulate Fruit Flavor Ester Biosynthesis. Plant J. 2021, 106, 785–800. [Google Scholar] [CrossRef]
- Bvindi, C.; Lee, S.; Tang, L.; Mickelbart, M.V.; Li, Y.; Mengiste, T. Improved Pathogen and Stress Tolerance in Tomato Mutants of SET Domain Histone 3 Lysine Methyltransferases. New Phytol. 2022, 235, 1957–1976. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.-E.; Wang, H.; Yang, Y.; Li, J.; Zhu, Z. Histone Deacetylase Gene SlHDA3 Is Involved in Drought and Salt Response in Tomato. Plant Growth Regul. 2023, 99, 359–372. [Google Scholar] [CrossRef]
- Yu, X.; Gao, Q.; Chen, G.; Guo, J.-E.; Guo, X.; Tang, B.; Hu, Z. SlHDA5, a Tomato Histone Deacetylase Gene, Is Involved in Responding to Salt, Drought, and ABA. Plant Mol. Biol. Rep. 2018, 36, 36–44. [Google Scholar] [CrossRef]
- Guo, J.-E.; Wang, H. Histone Deacetylase Gene SlHDA1 Regulates Drought and Salt Tolerance in Tomato (Solanum Lycopersicum). Sci. Hortic. 2023, 313, 111899. [Google Scholar] [CrossRef]
- Ding, X.; Zhang, D.; Gu, D.; Li, Z.; Liang, H.; Zhu, H.; Jiang, Y.; Duan, X. The Histone H3K27 Demethylase SlJMJ4 Promotes Dark- and ABA-Induced Leaf Senescence in Tomato. Hortic. Res. 2022, 9, uhab077. [Google Scholar] [CrossRef]
- Tamaki, S.; Matsuo, S.; Wong, H.L.; Yokoi, S.; Shimamoto, K. Hd3a Protein Is a Mobile Flowering Signal in Rice. Science 2007, 316, 1033–1036. [Google Scholar] [CrossRef]
- Komiya, R.; Ikegami, A.; Tamaki, S.; Yokoi, S.; Shimamoto, K. Hd3a and RFT1 Are Essential for Flowering in Rice. Development 2008, 135, 767–774. [Google Scholar] [CrossRef] [PubMed]
- Doi, K.; Izawa, T.; Fuse, T.; Yamanouchi, U.; Kubo, T.; Shimatani, Z.; Yano, M.; Yoshimura, A. Ehd1, a B-Type Response Regulator in Rice, Confers Short-Day Promotion of Flowering and Controls FT-like Gene Expression Independently of Hd1. Genes Dev. 2004, 18, 926–936. [Google Scholar] [CrossRef] [PubMed]
- Xue, W.; Xing, Y.; Weng, X.; Zhao, Y.; Tang, W.; Wang, L.; Zhou, H.; Yu, S.; Xu, C.; Li, X.; et al. Natural Variation in Ghd7 Is an Important Regulator of Heading Date and Yield Potential in Rice. Nat. Genet. 2008, 40, 761–767. [Google Scholar] [CrossRef]
- Noh, Y.-S.; Amasino, R.M. PIE1, an ISWI Family Gene, Is Required for FLC Activation and Floral Repression in Arabidopsis. Plant Cell 2003, 15, 1671–1682. [Google Scholar] [CrossRef]
- Zheng, D.; Wang, L.; Chen, L.; Pan, X.; Lin, K.; Fang, Y.; Wang, X.; Zhang, W. Salt-Responsive Genes Are Differentially Regulated at the Chromatin Levels Between Seedlings and Roots in Rice. Plant Cell Physiol. 2019, 60, 1790–1803. [Google Scholar] [CrossRef]
- Yin, W.; Xiao, Y.; Niu, M.; Meng, W.; Li, L.; Zhang, X.; Liu, D.; Zhang, G.; Qian, Y.; Sun, Z.; et al. ARGONAUTE2 Enhances Grain Length and Salt Tolerance by Activating BIG GRAIN3 to Modulate Cytokinin Distribution in Rice[OPEN]. Plant Cell 2020, 32, 2292–2306. [Google Scholar] [CrossRef] [PubMed]
- Giovannoni, J.; Nguyen, C.; Ampofo, B.; Zhong, S.; Fei, Z. The Epigenome and Transcriptional Dynamics of Fruit Ripening. Annu. Rev. Plant Biol. 2017, 68, 61–84. [Google Scholar] [CrossRef]
- Seymour, G.B.; Chapman, N.H.; Chew, B.L.; Rose, J.K.C. Regulation of Ripening and Opportunities for Control in Tomato and Other Fruits. Plant Biotechnol. J. 2013, 11, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, X.; Zhang, Y.; Zhang, A.; You, C.-X. Regulation of Fleshy Fruit Ripening: From Transcription Factors to Epigenetic Modifications. Hortic. Res. 2022, 9, uhac013. [Google Scholar] [CrossRef]
- Lü, P.; Yu, S.; Zhu, N.; Chen, Y.-R.; Zhou, B.; Pan, Y.; Tzeng, D.; Fabi, J.P.; Argyris, J.; Garcia-Mas, J.; et al. Genome Encode Analyses Reveal the Basis of Convergent Evolution of Fleshy Fruit Ripening. Nat. Plants 2018, 4, 784–791. [Google Scholar] [CrossRef]
- Nardini, M.; Gnesutta, N.; Donati, G.; Gatta, R.; Forni, C.; Fossati, A.; Vonrhein, C.; Moras, D.; Romier, C.; Bolognesi, M.; et al. Sequence-Specific Transcription Factor NF-Y Displays Histone-like DNA Binding and H2B-like Ubiquitination. Cell 2013, 152, 132–143. [Google Scholar] [CrossRef] [PubMed]
- Hu, G.; Huang, B.; Wang, K.; Frasse, P.; Maza, E.; Djari, A.; Benhamed, M.; Gallusci, P.; Li, Z.; Zouine, M.; et al. Histone Posttranslational Modifications Rather than DNA Methylation Underlie Gene Reprogramming in Pollination-Dependent and Pollination-Independent Fruit Set in Tomato. New Phytol. 2021, 229, 902–919. [Google Scholar] [CrossRef] [PubMed]
- Filichkin, S.A.; Ansariola, M.; Fraser, V.N.; Megraw, M. Identification of Transcription Factors from NF-Y, NAC, and SPL Families Responding to Osmotic Stress in Multiple Tomato Varieties. Plant Sci. 2018, 274, 441–450. [Google Scholar] [CrossRef]
- Li, Q.; Sun, W.; Chen, C.; Dong, D.; Cao, Y.; Dong, Y.; Yu, L.; Yue, Z.; Jin, X. Overexpression of Histone Demethylase Gene SlJMJ524 from Tomato Confers Cd Tolerance by Regulating Metal Transport-Related Protein Genes and Flavonoid Content in Arabidopsis. Plant Sci. 2022, 318, 111205. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Li, Z.; Wu, S.; Yu, C.; Wang, X.; Wang, Y.; Peng, Z.; Gao, Y.; Li, R.; Shen, Y.; et al. Coronatine Enhances Chilling Tolerance of Tomato Plants by Inducing Chilling-Related Epigenetic Adaptations and Transcriptional Reprogramming. Int. J. Mol. Sci. 2022, 23, 10049. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.E.; Neumann, M.; Duro, D.I.; Schmid, M. CRISPR-Based Tools for Targeted Transcriptional and Epigenetic Regulation in Plants. PLoS ONE 2019, 14, e0222778. [Google Scholar] [CrossRef] [PubMed]
- Varotto, S.; Tani, E.; Abraham, E.; Krugman, T.; Kapazoglou, A.; Melzer, R.; Radanoviæ, A.; Miladinoviæ, D. Epigenetics: Possible Applications in Climate-Smart Crop Breeding. J. Exp. Bot. 2020, 71, 5223–5236. [Google Scholar] [CrossRef]
- Kapazoglou, A.; Tondelli, A.; Papaefthimiou, D.; Ampatzidou, H.; Francia, E.; Stanca, M.A.; Bladenopoulos, K.; Tsaftaris, A.S. Epigenetic Chromatin Modifiers in Barley: IV. The Study of Barley Polycomb Group (PcG) Genes during Seed Development and in Response to External ABA. BMC Plant Biol. 2010, 10, 73. [Google Scholar] [CrossRef]
- Yang, X.; Kundariya, H.; Xu, Y.Z.; Sandhu, A.; Yu, J.; Hutton, S.F.; Zhang, M.; Mackenzie, S.A. Muts HOMOLOG1-Derived Epigenetic Breeding Potential in Tomato. Plant Physiol. 2015, 168, 222–232. [Google Scholar] [CrossRef]
- Hilton, I.B.; D’Ippolito, A.M.; Vockley, C.M.; Thakore, P.I.; Crawford, G.E.; Reddy, T.E.; Gersbach, C.A. Epigenome Editing by a CRISPR-Cas9-Based Acetyltransferase Activates Genes from Promoters and Enhancers. Nat. Biotechnol. 2015, 33, 510–517. [Google Scholar] [CrossRef]
- Raju, S.K.K.; Shao, M.R.; Sanchez, R.; Xu, Y.Z.; Sandhu, A.; Graef, G.; Mackenzie, S. An Epigenetic Breeding System in Soybean for Increased Yield and Stability. Plant Biotechnol. J. 2018, 16, 1836–1847. [Google Scholar] [CrossRef]
- Liu, B.; Wendel, J.F. Epigenetic Phenomena and the Evolution of Plant Allopolyploids. Mol. Phylogenetics Evol. 2003, 29, 365–379. [Google Scholar] [CrossRef]
- Song, Q.; Zhang, T.; Stelly, D.M.; Chen, Z.J. Epigenomic and Functional Analyses Reveal Roles of Epialleles in the Loss of Photoperiod Sensitivity during Domestication of Allotetraploid Cottons. Genome Biol. 2017, 18, 99. [Google Scholar] [CrossRef]
- Zhang, Y.-Y.; Fischer, M.; Colot, V.; Bossdorf, O. Epigenetic Variation Creates Potential for Evolution of Plant Phenotypic Plasticity. New Phytol. 2013, 197, 314–322. [Google Scholar] [CrossRef]
- Lieberman-Lazarovich, M.; Kaiserli, E.; Bucher, E.; Mladenov, V. Natural and Induced Epigenetic Variation for Crop Improvement. Curr. Opin. Plant Biol. 2022, 70, 102297. [Google Scholar] [CrossRef] [PubMed]
- Mladenov, V.; Fotopoulos, V.; Kaiserli, E.; Karalija, E.; Maury, S.; Baranek, M.; Segal, N.; Testillano, P.S.; Vassileva, V.; Pinto, G.; et al. Deciphering the Epigenetic Alphabet Involved in Transgenerational Stress Memory in Crops. Int. J. Mol. Sci. 2021, 22, 7118. [Google Scholar] [CrossRef]
- Miura, K.; Agetsuma, M.; Kitano, H.; Yoshimura, A.; Matsuoka, M.; Jacobsen, S.E.; Ashikari, M. A Metastable DWARF1 Epigenetic Mutant Affecting Plant Stature in Rice. Proc. Natl. Acad. Sci. USA 2009, 106, 11218–11223. [Google Scholar] [CrossRef]
- Miura, K.; Ikeda, M.; Matsubara, A.; Song, X.J.; Ito, M.; Asano, K.; Matsuoka, M.; Kitano, H.; Ashikari, M. OsSPL14 Promotes Panicle Branching and Higher Grain Productivity in Rice. Nat. Genet. 2010, 42, 545–549. [Google Scholar] [CrossRef] [PubMed]
- Quadrana, L.; Almeida, J.; Asís, R.; Duffy, T.; Dominguez, P.G.; Bermúdez, L.; Conti, G.; Corrêa da Silva, J.V.; Peralta, I.E.; Colot, V.; et al. Natural Occurring Epialleles Determine Vitamin E Accumulation in Tomato Fruits. Nat. Commun. 2014, 5, 4027. [Google Scholar] [CrossRef]
- Manning, K.; Tör, M.; Poole, M.; Hong, Y.; Thompson, A.J.; King, G.J.; Giovannoni, J.J.; Seymour, G.B. A Naturally Occurring Epigenetic Mutation in a Gene Encoding an SBP-Box Transcription Factor Inhibits Tomato Fruit Ripening. Nat. Genet. 2006, 38, 948–952. [Google Scholar] [CrossRef] [PubMed]
- Moradpour, M.; Abdulah, S.N.A. CRISPR/DCas9 Platforms in Plants: Strategies and Applications beyond Genome Editing. Plant Biotechnol. J. 2020, 18, 32–44. [Google Scholar] [CrossRef]
- Mutum, R.D.; Balyan, S.C.; Kansal, S.; Agarwal, P.; Kumar, S.; Kumar, M.; Raghuvanshi, S. Evolution of Variety-Specific Regulatory Schema for Expression of Osa-MiR408 in Indica Rice Varieties under Drought Stress. FEBS J. 2013, 280, 1717–1730. [Google Scholar] [CrossRef]
- Wang, W.; Lu, Y.; Li, J.; Zhang, X.; Hu, F.; Zhao, Y.; Zhou, D.-X. SnRK1 Stimulates the Histone H3K27me3 Demethylase JMJ705 to Regulate a Transcriptional Switch to Control Energy Homeostasis. Plant Cell 2021, 33, 3721–3742. [Google Scholar] [CrossRef]
- Xie, Y.; Zhang, Y.; Han, J.; Luo, J.; Li, G.; Huang, J.; Wu, H.; Tian, Q.; Zhu, Q.; Chen, Y.; et al. The Intronic Cis Element SE1 Recruits Trans-Acting Repressor Complexes to Repress the Expression of ELONGATED UPPERMOST INTERNODE1 in Rice. Mol. Plant 2018, 11, 720–735. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Zhang, X.; Yang, Y.; Zhang, H.; Zhu, W.; Nie, W.F. The Histone Variant Sl_H2A.Z Regulates Carotenoid Biosynthesis and Gene Expression during Tomato Fruit Ripening. Hortic. Res. 2021, 8, 85. [Google Scholar] [CrossRef]
- Metje-Sprink, J.; Menz, J.; Modrzejewski, D.; Sprink, T. DNA-Free Genome Editing: Past, Present and Future. Front. Plant Sci. 2019, 9, 1957. [Google Scholar] [CrossRef] [PubMed]
- Agapito-Tenfen, S.Z.; Okoli, A.S.; Bernstein, M.J.; Wikmark, O.-G.; Myhr, A.I. Revisiting Risk Governance of GM Plants: The Need to Consider New and Emerging Gene-Editing Techniques. Front. Plant Sci. 2018, 9, 1874. [Google Scholar] [CrossRef] [PubMed]
- Davison, J.; Ammann, K. New GMO Regulations for Old: Determining a New Future for EU Crop Biotechnology. GM Crops Food 2017, 8, 13–34. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Species | Physiological Process | Epigenetic Mark | Associated Genes | Effect of Regulation | Reference |
|---|---|---|---|---|---|
| Rice (Oryza sativa) | Flowering | H3K4me3 | OsTRX1, SIP1, Ehd1 | Promotes flowering | [40] |
| OsWDR5a, Ehd1 | Promotes heading date and secondary branch growth | [41] | |||
| H3K36me3 | OsSDG725, OsSUF4, Ehd1 | Promotes flowering | [42] | ||
| De-Acetylation (H4K8, H4K12) | OsHDA703, Ghd7, Ehd1 | Promotes flowering | [43] | ||
| H2A.Z | OsINO80, OsCPS1, OsGA3OX2 | Promotes flowering and reproductive efficiency | [44] | ||
| Grain size and quality | H3K27me3 | OsJMJ705, OsWOX11 | Represses shoot growth | [45] | |
| OsNGR5, OsPRC2 | Promotes tiller number | [46] | |||
| OsVIL2, OsCKX2 | Promotes cytokinin synthesis and plant biomass | [47] | |||
| H3K4me3 | OsMADs | Promotes grain size and yield | [48] | ||
| Acetylation (H3 and H4) | OsHDR3, GW6a | Promotes grain size and yield | [49] | ||
| Abiotic stress | Acetylation (H4K5, H4K16, H3K9) | OsHDA710, OsLEA3, OsABI5 | Increases salt tolerance | [50] | |
| Acetylation (H3) | OsIDS1, OsLEA1, OsSOS1 | Increases salt tolerance | [51] | ||
| H3K36me3 | OsSDG708 | Increases drought resistance | [52] | ||
| H3K4me3 | OsJMJ703 | Increases drought tolerance | [53] | ||
| Tomato (Solanum lycopersicum) | Fruit ripening | H3K27me3 | SlJMJ6 | Represses fruit ripening | [54] |
| SlLHP1b | Represses fruit softening and ethylene accumulation | [55] | |||
| SlNF-YB, SlCHS1 | Represses flavonoid accumulation | [56] | |||
| H3K4me3 | SlJMJ7 | Promotes ethylene biosynthesis | [57] | ||
| Acetylation | SlHDA3, SlACSs, SlE4, SlLOX8 | Promotes ripening and carotenoid accumulation | [58] | ||
| Acetylation (H3) | SlHDT1 | Promotes carotenoid accumulation and ethylene biosynthesis | [59] | ||
| Acetylation (H3K9, H3K27) | SlERF.F12, SlTPL2, SlHDA1/HDA3 | Promotes fruit ripening | [60] | ||
| H3K4me3 | SlNOR, SlAAT1 | Promotes fruit ripening and ester synthesis | [61] | ||
| H3K27me3 | SlNOR, SlAAT1 | Represses fruit ripening and ester synthesis | [61] | ||
| Abiotic stress | H3K4me3 H3K36me3 | SlSDG33/34 | Promotes drought tolerance | [62] | |
| De-Acetylation | SlHDA3 | Promotes drought resistance | [63] | ||
| SlHDA5 | Promotes salt and drought resistance | [64] | |||
| SlHDA1 | Promotes salt and drought resistance | [65] | |||
| H3K27me3 | SlJMJ4 | Represses plant sensitivity to ABA | [66] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fang, W.; Fasano, C.; Perrella, G. Unlocking the Secret to Higher Crop Yield: The Potential for Histone Modifications. Plants 2023, 12, 1712. https://doi.org/10.3390/plants12081712
Fang W, Fasano C, Perrella G. Unlocking the Secret to Higher Crop Yield: The Potential for Histone Modifications. Plants. 2023; 12(8):1712. https://doi.org/10.3390/plants12081712
Chicago/Turabian StyleFang, Weiwei, Carlo Fasano, and Giorgio Perrella. 2023. "Unlocking the Secret to Higher Crop Yield: The Potential for Histone Modifications" Plants 12, no. 8: 1712. https://doi.org/10.3390/plants12081712
APA StyleFang, W., Fasano, C., & Perrella, G. (2023). Unlocking the Secret to Higher Crop Yield: The Potential for Histone Modifications. Plants, 12(8), 1712. https://doi.org/10.3390/plants12081712