


Crinum bulbispermum, a Medicinal Geophyte with Phytostabilization Properties in Metal-Enriched Mine Tailings

Abstract
:1. Introduction
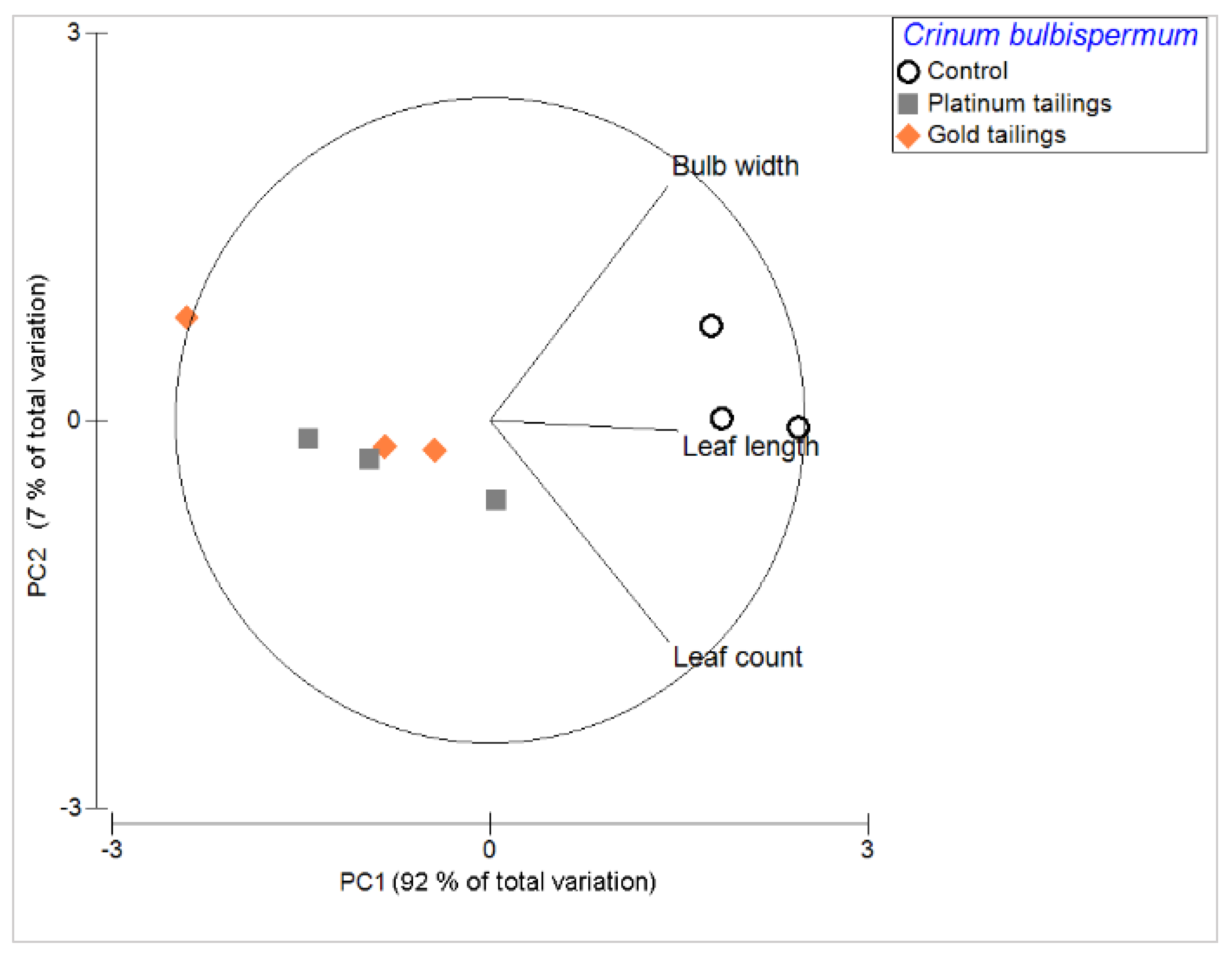
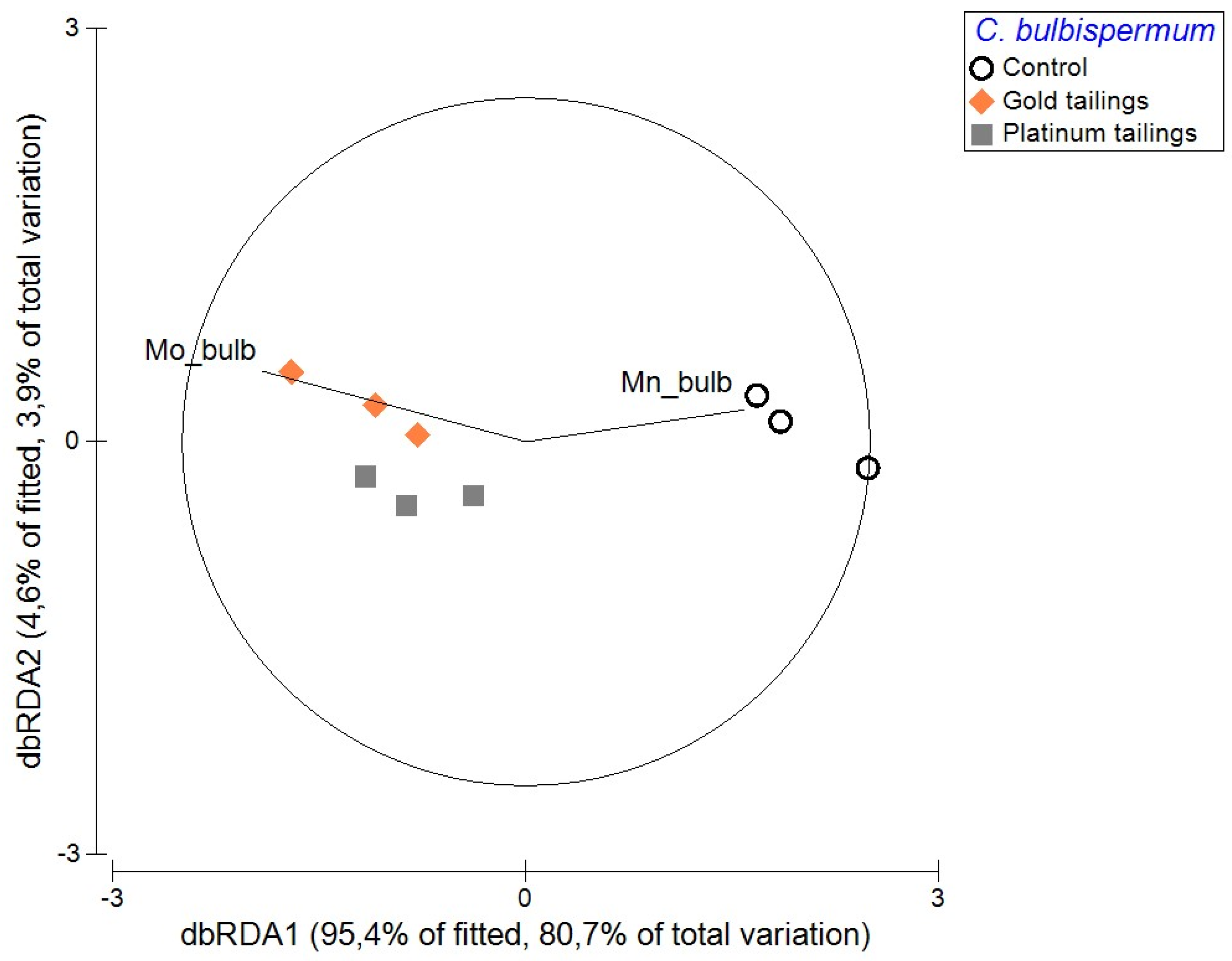
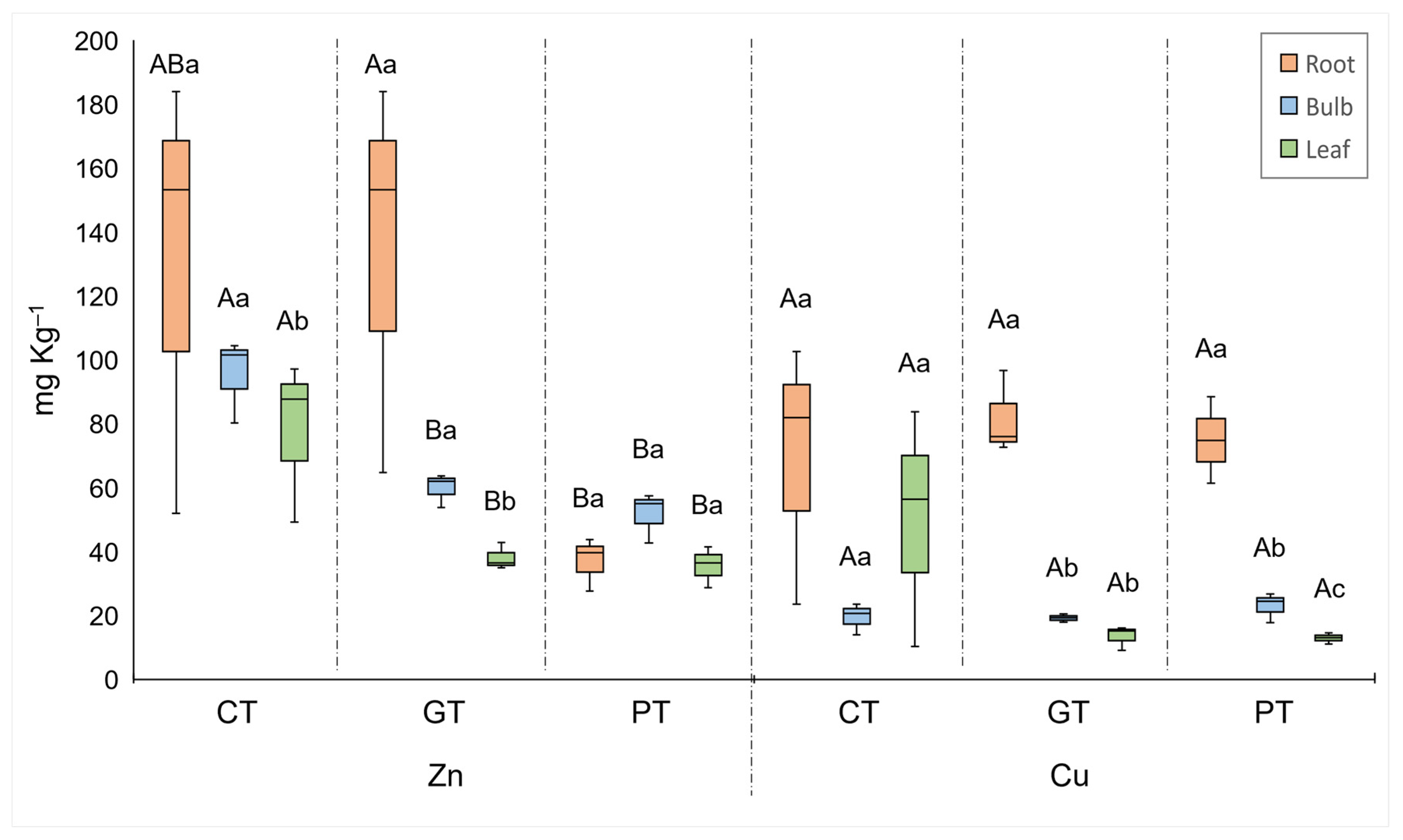
2. Results
3. Discussion
3.1. What Is the Growth Performance of Crinum Plants on Tailings?
3.2. Do Crinum Plants Take Up Metals When Grown on Tailings?
3.3. Which Organs (Roots, Bulbs, or Leaves) Take Up PTMs?
3.4. Key PTMs Associated with the Plant Organs and Each Treatment
3.5. Bioaccumulation
4. Materials and Methods
4.1. Focal Species
4.2. Pot Trials
4.3. Measurements
4.4. Statistical Analyses
4.4.1. Plant Growth
4.4.2. Bioaccumulation
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Egoh, B.N.; Reyers, B.; Rouget, M.; Richardson, D.M. Identifying Priority Areas for Ecosystem Service Management in South African Grasslands. J. Environ. Manag. 2011, 92, 1642–1650. [Google Scholar] [CrossRef] [PubMed]
- Reyers, B.; Nel, J.; Egoh, B.; Jonas, Z.; Rouget, M. Background Information Report No. 1: Grassland Biodiversity Profile and Spatial Biodiversity Priority Assessment; CSIR Report Number: ENV-S-C; SANBI: Pretoria, South Africa, 2005; p. 102. [Google Scholar]
- Sheoran, V.; Sheoran, A.; Poonia, P. Soil Reclamation of Abandoned Mine Land by Revegetation: A Review. Int. J. Soil Sediment Water 2010, 3, 13. Available online: https://scholarworks.umass.edu/intljssw/vol3/iss2/13 (accessed on 13 September 2023).
- Miao, Z.; Bai, Z.; Gao, L. Ecological Rebuilding and Land Reclamation in Surface Mines in Shanxi Province, China. J. Environ. Sci. 2000, 12, 486–497. [Google Scholar]
- Kumar, N.; Kumar, A.; Singh, M. Floristic Diversity Assessment in Ecologically Restored Limestone (Building Stone) Mine near Chechat Village, Kota District, Rajasthan. Ecologia 2014, 4, 16–25. [Google Scholar] [CrossRef]
- Saxena, G.; Purchase, D.; Mulla, S.I.; Saratale, G.D.; Bharagava, R.N. Phytoremediation of Heavy Metal-Contaminated Sites: Eco-Environmental Concerns, Field Studies, Sustainability Issues, and Future Prospects. Rev. Environ. Contam. Toxicol. 2020, 249, 71–131. [Google Scholar] [PubMed]
- Adhikari, S.; Siebert, S.J.; Jordaan, A. Evidence of Chromium Dust Pollution on the Leaves of Food and Medicinal Plants from Mining Areas of Sekhukhuneland, South Africa. S. Afr. J. Bot. 2021, 143, 226–237. [Google Scholar] [CrossRef]
- Shen, Z.-G.; Li, X.-D.; Wang, C.-C.; Chen, H.-M.; Chua, H. Lead Phytoextraction from Contaminated Soil with High-Biomass Plant Species. J. Environ. Qual. 2002, 31, 1893–1900. [Google Scholar] [CrossRef]
- Ali, H.; Khan, E.; Ilahi, I. Environmental Chemistry and Ecotoxicology of Hazardous Heavy Metals: Environmental Persistence, Toxicity, and Bioaccumulation. J. Chem. 2019, 2019, 6730305. [Google Scholar] [CrossRef]
- Van Rensburg, L.; Morgenthal, T. The Effect of Woodchip Waste on Vegetation Establishment during Platinum Tailings Rehabilitation. S. Afr. J. Sci. 2004, 100, 294–300. [Google Scholar]
- Langridge, P. Micronutrient Toxicity and Deficiency. In Wheat Improvement: Food Security in a Changing Climate; Springer: Berlin/Heidelberg, Germany, 2022; pp. 433–449. [Google Scholar]
- Djingova, R.; Kuleff, I. Chapter 5 Instrumental Techniques for Trace Analysis. Trace Met. Environ. 2000, 4, 137–185. [Google Scholar] [CrossRef]
- Van Assche, F.; Clijsters, H. Effects of Metals on Enzyme Activity in Plants. Plant Cell Environ. 1990, 13, 195–206. [Google Scholar] [CrossRef]
- Jadia, C.D.; Fulekar, M.H. Phytoremediation of Heavy Metals: Recent Techniques. Afr. J. Biotechnol. 2009, 8, 921–928. [Google Scholar]
- Robinson, B.H.; Bañuelos, G.; Conesa, H.M.; Evangelou, M.W.H.; Schulin, R. The Phytomanagement of Trace Elements in Soil. CRC Crit. Rev. Plant Sci. 2009, 28, 240–266. [Google Scholar] [CrossRef]
- Berti, W.R.; Cunningham, S.D. Phytostabilization of Metals: Using Metals to Clean Up the Environment; Wiley: New York, NY, USA, 2000. [Google Scholar]
- Park, J.H.; Lamb, D.; Paneerselvam, P.; Choppala, G.; Bolan, N.; Chung, J.W. Role of Organic Amendments on Enhanced Bioremediation of Heavy Metal(Loid) Contaminated Soils. J. Hazard. Mater. 2011, 185, 549–574. [Google Scholar] [CrossRef] [PubMed]
- Reeves, R.D. Tropical Hyperaccumulators of Metals and Their Potential for Phytoextraction. Plant Soil 2003, 249, 57–65. [Google Scholar] [CrossRef]
- van der Ent, A.; Baker, A.J.M.; Reeves, R.D.; Pollard, A.J.; Schat, H. Hyperaccumulators of Metal and Metalloid Trace Elements: Facts and Fiction. Plant Soil 2013, 362, 319–334. [Google Scholar] [CrossRef]
- Baker, A.J.; Walker, P.L. Ecophysiology of Metal Uptake by Tolerant Plants. In Heavy Metal Tolerance in Plants: Evolutionary Aspects; Shaw, A.J., Ed.; CRC Press: Boca Raton, FL, USA, 1990; Volume 2, pp. 155–165. [Google Scholar]
- Buisson, E.; Le Stradic, S.; Silveira, F.A.O.; Durigan, G.; Overbeck, G.E.; Fidelis, A.; Fernandes, G.W.; Bond, W.J.; Hermann, J.M.; Mahy, G.; et al. Resilience and Restoration of Tropical and Subtropical Grasslands, Savannas, and Grassy Woodlands. Biol. Rev. 2019, 94, 590–609. [Google Scholar] [CrossRef] [PubMed]
- Morris, C.D.; Scott-Shaw, R. Potential Grazing Indicator Forbs for Two Mesic Grasslands in South Africa. Ecol. Indic. 2019, 107, 105611. [Google Scholar] [CrossRef]
- Moteetee, A.; Van Wyk, B.E. The Medical Ethnobotany of Lesotho: A Review. Bothalia 2011, 41, 209–228. [Google Scholar] [CrossRef]
- Siebert, F.; Dreber, N. Forb Ecology Research in Dry African Savannas: Knowledge, Gaps, and Future Perspectives. Ecol. Evol. 2019, 9, 7875–7891. [Google Scholar] [CrossRef]
- Slooten, E.; Jordaan, E.; White, J.D.M.; Archibald, S.; Siebert, F. South African Grasslands and Ploughing: Outlook for Agricultural Expansion in Africa. S. Afr. J. Sci. 2023, 119, 15540. [Google Scholar] [CrossRef] [PubMed]
- Siebert, F.; Terblanche, R. Surprise green feasts. Veld Flora 2020, 106, 20–23. [Google Scholar]
- Buisson, E.; Fidelis, A.; Overbeck, G.E.; Schmidt, I.B.; Durigan, G.; Young, T.P.; Alvarado, S.T.; Arruda, A.J.; Boisson, S.; Bond, W.; et al. A Research Agenda for the Restoration of Tropical and Subtropical Grasslands and Savannas. Restor. Ecol. 2021, 29, e13292. [Google Scholar] [CrossRef]
- Pedrini, S.; Lewandrowski, W.; Stevens, J.C.; Dixon, K.W. Optimising Seed Processing Techniques to Improve Germination and Sowability of Native Grasses for Ecological Restoration. Plant. Biol. 2019, 21, 415–424. [Google Scholar] [CrossRef]
- Chibuike, G.U.; Obiora, S.C. Heavy Metal Polluted Soils: Effect on Plants and Bioremediation Methods. Appl. Environ. Soil Sci. 2014, 2014, 752708. [Google Scholar] [CrossRef]
- Wang, Y.; Narayanan, M.; Shi, X.; Chen, X.; Li, Z.; Natarajan, D.; Ma, Y. Plant Growth-Promoting Bacteria in Metal-Contaminated Soil: Current Perspectives on Remediation Mechanisms. Front. Microbiol. 2022, 13, 966226. [Google Scholar] [CrossRef]
- Alejandro, S.; Höller, S.; Meier, B.; Peiter, E. Manganese in Plants: From Acquisition to Subcellular Allocation. Front. Plant. Sci. 2020, 11, 300. [Google Scholar] [CrossRef]
- Bricker, T.M.; Roose, J.L.; Fagerlund, R.D.; Frankel, L.K.; Eaton-Rye, J.J. The Extrinsic Proteins of Photosystem II. BBA—Bioenerg. 2012, 1817, 121–142. [Google Scholar] [CrossRef]
- Broadley, M.; Brown, P.; Cakmak, I.; Rengel, Z.; Zhao, F. Function of Nutrients: Micronutrients. In Marschner’s Mineral Nutrition of Higher Plants; Marschner, P., Ed.; Academic Press: London, UK, 2012; pp. 191–249. [Google Scholar]
- Doncheva, S.; Georgieva, K.; Vassileva, V.; Stoyanova, Z.; Popov, N.; Ignatov, G. Effects of Succinate on Manganese Toxicity in Pea Plants. J. Plant. Nutr. 2005, 28, 47–62. [Google Scholar] [CrossRef]
- Kaiser, B.N.; Gridley, K.L.; Brady, J.N.; Phillips, T.; Tyerman, S.D. The Role of Molybdenum in Agricultural Plant Production. Ann. Bot. 2005, 96, 745–754. [Google Scholar] [CrossRef]
- McGrath, S.P.; Micó, C.; Zhao, F.J.; Stroud, J.L.; Zhang, H.; Fozard, S. Predicting Molybdenum Toxicity to Higher Plants: Influence of Soil Properties. Environ. Pollut. 2010, 158, 3095–3102. [Google Scholar] [CrossRef] [PubMed]
- Yan, A.; Wang, Y.; Tan, S.N.; Mohd Yusof, M.L.; Ghosh, S.; Chen, Z. Phytoremediation: A Promising Approach for Revegetation of Heavy Metal-Polluted Land. Front. Plant Sci. 2020, 11, 513099. [Google Scholar] [CrossRef] [PubMed]
- Ulhassan, Z.; Gill, R.A.; Huang, H.; Ali, S.; Mwamba, T.M.; Ali, B.; Huang, Q.; Hamid, Y.; Khan, A.R.; Wang, J.; et al. Selenium Mitigates the Chromium Toxicity in Brassicca napus L. by Ameliorating Nutrients Uptake, Amino Acids Metabolism and Antioxidant Defense System. Plant Physiol. Biochem. 2019, 145, 142–152. [Google Scholar] [CrossRef] [PubMed]
- Filek, M.; Zembala, M.; Hartikainen, H.; Miszalski, Z.; Kornaś, A.; Wietecka-Posłuszny, R.; Walas, P. Changes in Wheat Plastid Membrane Properties Induced by Cadmium and Selenium in Presence/Absence of 2,4-Dichlorophenoxyacetic Acid. Plant. Cell. Tissue Organ. Cult. 2009, 96, 19–28. [Google Scholar] [CrossRef]
- Handa, N.; Kohli, S.K.; Sharma, A.; Thukral, A.K.; Bhardwaj, R.; Abd_Allah, E.F.; Alqarawi, A.A.; Ahmad, P. Selenium Modulates Dynamics of Antioxidative Defence Expression, Photosynthetic Attributes and Secondary Metabolites to Mitigate Chromium Toxicity in Brassica juncea L. Plants. Environ. Exp. Bot. 2019, 161, 180–192. [Google Scholar] [CrossRef]
- Li, Y.; Hu, W.; Zhao, J.; Chen, Q.; Wang, W.; Li, B.; Li, Y.F. Selenium Decreases Methylmercury and Increases Nutritional Elements in Rice Growing in Mercury-Contaminated Farmland. Ecotoxicol. Environ. Saf. 2019, 182, 109447. [Google Scholar] [CrossRef]
- Wan, Y.; Wang, K.; Liu, Z.; Yu, Y.; Wang, Q.; Li, H. Effect of Selenium on the Subcellular Distribution of Cadmium and Oxidative Stress Induced by Cadmium in Rice (Oryza sativa L.). Environ. Sci. Pollut. Res. 2019, 26, 16220–16228. [Google Scholar] [CrossRef]
- Shahid, M.A.; Balal, R.M.; Khan, N.; Zotarelli, L.; Liu, G.D.; Sarkhosh, A.; Fernández-Zapata, J.C.; Martínez Nicolás, J.J.; Garcia-Sanchez, F. Selenium Impedes Cadmium and Arsenic Toxicity in Potato by Modulating Carbohydrate and Nitrogen Metabolism. Ecotoxicol. Environ. Saf. 2019, 180, 588–599. [Google Scholar] [CrossRef]
- Begum, M.C.; Islam, M.S.; Islam, M.; Amin, R.; Parvez, M.S.; Kabir, A.H. Biochemical and Molecular Responses Underlying Differential Arsenic Tolerance in Rice (Oryza sativa L.). Plant Physiol. Biochem. 2016, 104, 266–277. [Google Scholar] [CrossRef]
- Garg, N.; Singla, P. Arsenic Toxicity in Crop Plants: Physiological Effects and Tolerance Mechanisms. Environ. Chem. Lett. 2011, 9, 303–321. [Google Scholar] [CrossRef]
- Fennell, C.W.; Van Staden, J. Crinum Species in Traditional and Modern Medicine. J. Ethnopharmacol. 2001, 78, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Lux, A.; Martinka, M.; Vaculík, M.; White, P.J. Root Responses to Cadmium in the Rhizosphere: A Review. J. Exp. Bot. 2011, 62, 21–37. [Google Scholar] [CrossRef] [PubMed]
- Ebenebe, P.C.; Shale, K.; Sedibe, M.; Tikilili, P.; Achilonu, M.C. South African Mine Effluents: Heavy Metal Pollution and Impact on the Ecosystem. Int. J. Chem. Sci. 2017, 15, 198. [Google Scholar]
- Rudani, K.; Patel, V.; Prajapati, K. The Importance of Zinc in Plant Growth—A Review. Int. Res. J. Nat. Appl. Sci. 2018, 5, 38–48. [Google Scholar]
- Sturikova, H.; Krystofova, O.; Huska, D.; Adam, V. Zinc, Zinc Nanoparticles and Plants. J. Hazard. Mater. 2018, 349, 101–110. [Google Scholar] [CrossRef]
- Mir, A.R.; Pichtel, J.; Hayat, S. Copper: Uptake, Toxicity and Tolerance in Plants and Management of Cu-Contaminated Soil. BioMetals 2021, 34, 737–759. [Google Scholar] [CrossRef]
- Shabbir, Z.; Sardar, A.; Shabbir, A.; Abbas, G.; Shamshad, S.; Khalid, S.; Tahir, N.; Murtaza, G.; Dumat, C.; Shahid, M. Copper Uptake, Essentiality, Toxicity, Detoxification and Risk Assessment in Soil-Plant Environment. Chemosphere 2020, 259, 127436. [Google Scholar] [CrossRef]
- Soler-Rovira, P.; Madejón, E.; Madejón, P.; Plaza, C. In Situ Remediation of Metal-Contaminated Soils with Organic Amendments: Role of Humic Acids in Copper Bioavailability. Chemosphere 2010, 79, 844–849. [Google Scholar] [CrossRef]
- Lyu, S.; Wei, X.; Chen, J.; Wang, C.; Wang, X.; Pan, D. Titanium as a Beneficial Element for Crop Production. Front. Plant Sci. 2017, 8, 237149. [Google Scholar] [CrossRef]
- Soumare, A.; Diedhiou, A.G.; Thuita, M.; Hafidi, M.; Ouhdouch, Y.; Gopalakrishnan, S.; Kouisni, L. Exploiting Biological Nitrogen Fixation: A Route towards a Sustainable Agriculture. Plants 2020, 9, 1011. [Google Scholar] [CrossRef]
- Juhong, L. Cobalt: Physiological Effects and Uptake Mechanisms in Plants. Ph.D. Thesis, University of Adelaide, Adelaide, Australia, 1998. [Google Scholar]
- Seeda, A.; Abou El-Nour, E.; Mervat, G.; Zaghloul, S. Interaction of Copper, Zinc, and Their Importance in Plant Physiology: Review, Acquisition and Transport. Middle East J. Appl. Sci. 2020, 10, 407–434. [Google Scholar] [CrossRef]
- Tanveer, M.; Shabala, S. Entangling the Interaction between Essential and Nonessential Nutrients: Implications for Global Food Security. In Plant Nutrition and Food Security in the Era of Climate Change; Academic Press: Cambridge, MA, USA, 2022; pp. 1–25. [Google Scholar] [CrossRef]
- Huybrechts, M.; Hendrix, S.; Bertels, J.; Beemster, G.T.S.; Vandamme, D.; Cuypers, A. Spatial Analysis of the Rice Leaf Growth Zone under Controlled and Cadmium-Exposed Conditions. Environ. Exp. Bot. 2020, 177, 104120. [Google Scholar] [CrossRef]
- Verdoorn, I.C. The Genus Crinum in Southern Africa. Bothalia 1973, 11, 27–52. [Google Scholar] [CrossRef]
- Bond, W.J.; Parr, C.L. Beyond the Forest Edge: Ecology, Diversity and Conservation of the Grassy Biomes. Biol. Conserv. 2010, 143, 2395–2404. [Google Scholar] [CrossRef]
- Bjora, C.; Kwembeya, E.; Nordal, I. Crinum jasonii (Amaryllidaceae): A New Endemic Pan Species of the Luangwa Valley in Zambia with Notes on Different Seed Structures in the Genus. Kew Bull. 2006, 61, 569–577. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Ca | Mg | K | Na | Cd | Cu | Co | Mo | Mn | Zn | pH (H2O) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Control | 2601.6 | 912.7 | 1771.9 | 848.9 | 0.0451 | 16.83 | 32.41 | 2.566 | 442.1 | 55.32 | 5.71 |
| Gold tailings | 1572.5 | 46.3 | 22.2 | 7.9 | 0.1051 | 40.45 | 85.65 | 2.697 | 270.8 | 51.62 | 6.70 |
| Platinum tailings | 261.0 | 35.1 | 13.2 | 6.9 | 0.1237 | 188.5 | 285.4 | 2.579 | 2131 | 170.1 | 7.21 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Clarke, V.C.; Marcelo-Silva, J.; Claassens, S.; Siebert, S.J. Crinum bulbispermum, a Medicinal Geophyte with Phytostabilization Properties in Metal-Enriched Mine Tailings. Plants 2024, 13, 79. https://doi.org/10.3390/plants13010079
Clarke VC, Marcelo-Silva J, Claassens S, Siebert SJ. Crinum bulbispermum, a Medicinal Geophyte with Phytostabilization Properties in Metal-Enriched Mine Tailings. Plants. 2024; 13(1):79. https://doi.org/10.3390/plants13010079
Chicago/Turabian StyleClarke, Vincent C., João Marcelo-Silva, Sarina Claassens, and Stefan J. Siebert. 2024. "Crinum bulbispermum, a Medicinal Geophyte with Phytostabilization Properties in Metal-Enriched Mine Tailings" Plants 13, no. 1: 79. https://doi.org/10.3390/plants13010079
APA StyleClarke, V. C., Marcelo-Silva, J., Claassens, S., & Siebert, S. J. (2024). Crinum bulbispermum, a Medicinal Geophyte with Phytostabilization Properties in Metal-Enriched Mine Tailings. Plants, 13(1), 79. https://doi.org/10.3390/plants13010079