
Variation in Nutritional Components and Antioxidant Capacity of Different Cultivars and Organs of Basella alba

 ,
,  and
and
Abstract
:1. Introduction
2. Results
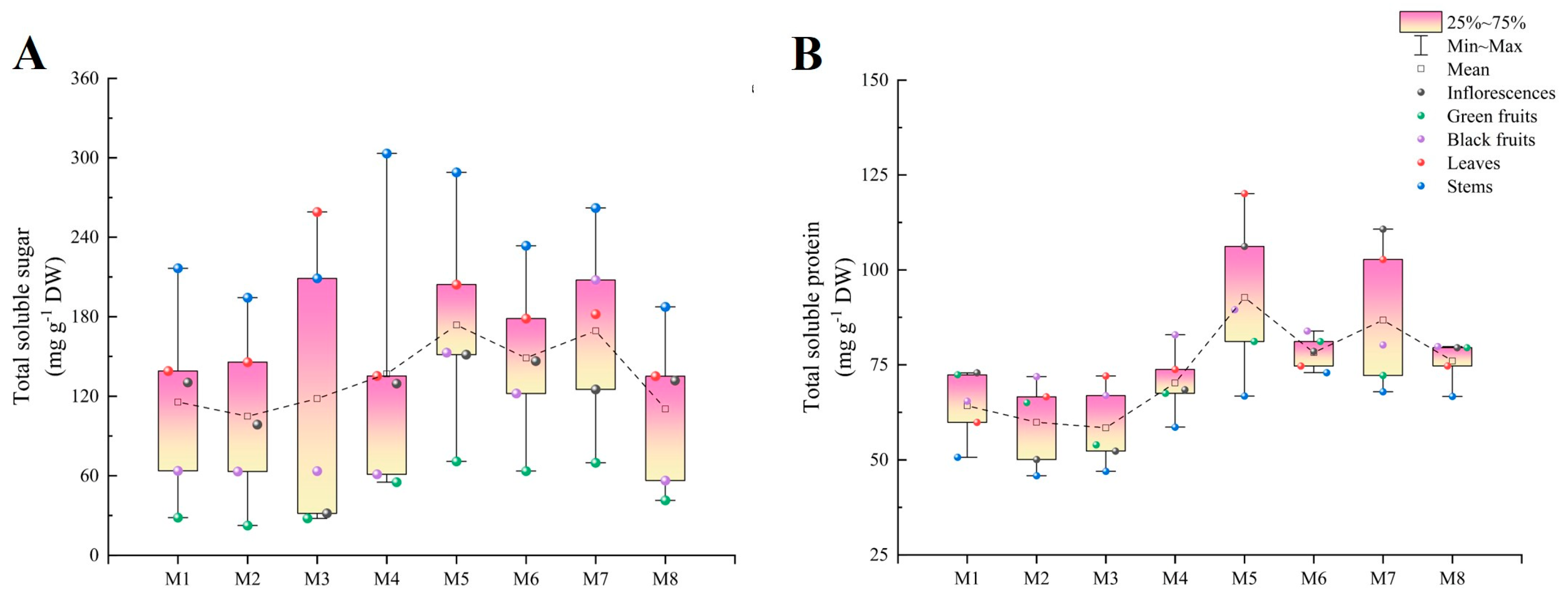
2.1. Total Soluble Sugar
2.2. Total Soluble Protein
2.3. Total Chlorophyll and Total Carotenoids
2.4. Total Ascorbic Acid
2.5. Total Proanthocyanidins, Total Flavonoids, and Total Phenolics
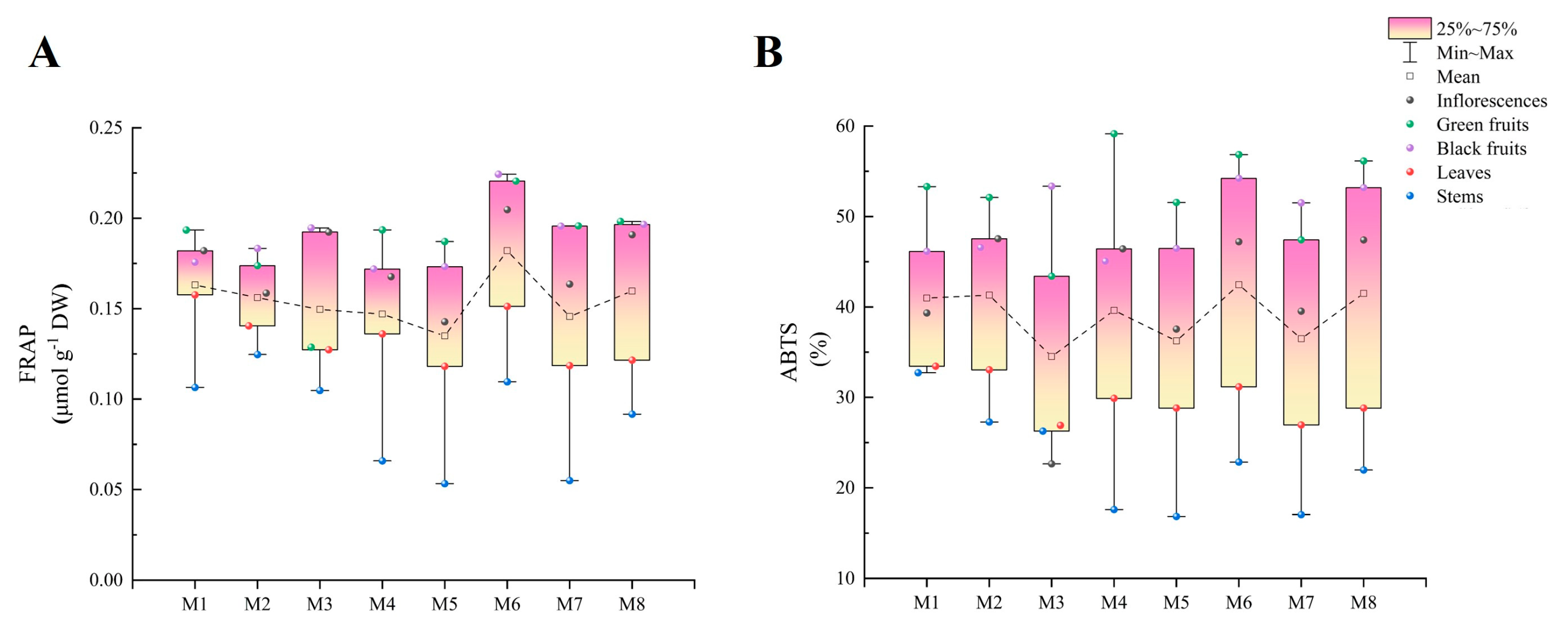
2.6. FRAP Analysis
2.7. ABTS Analysis
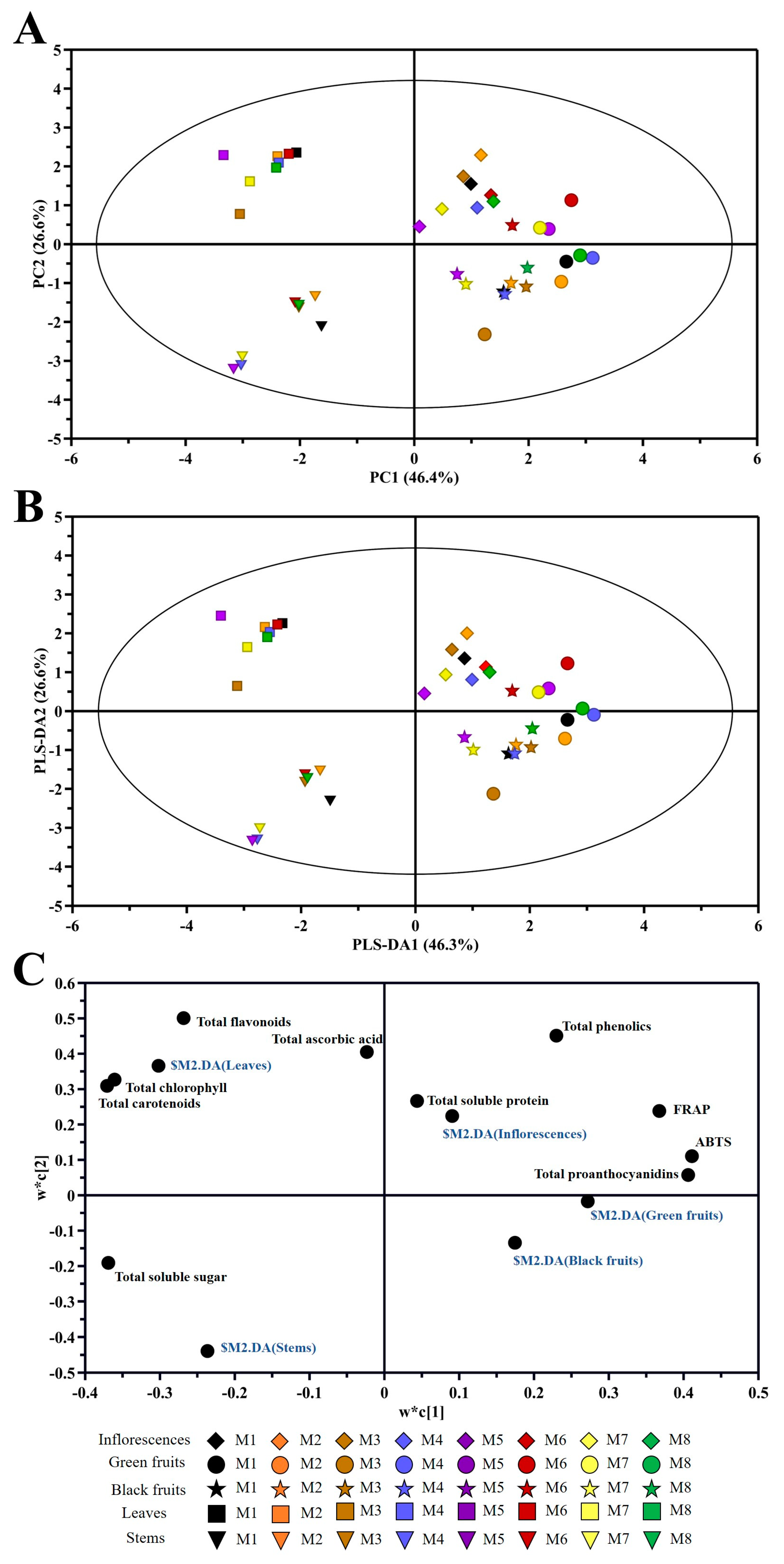
2.8. Principal Component Analysis (PCA)
2.9. Correlation Analysis
2.10. Variance Analysis
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Plant Material
4.3. Total Soluble Sugar Content
4.4. Total Soluble Protein Content
4.5. Total Chlorophyll Content
4.6. Total Carotenoid Content
4.7. Total Ascorbic Acid Content
4.8. Total Proanthocyanidin Content
4.9. Total Flavonoid Content
4.10. Total Phenolic Content
4.11. Ferric Reducing Antioxidant Power (FRAP)
4.12. 2,2-Azinobis (3-ethyl-Benzothiazoline-6-Sulfonic Acid) (ABTS) Assay
4.13. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Fadhila, F.R.; Umar, A.; Chandren, S.; Apriandanu, D.O.B.; Yulizar, Y. Biosynthesis of CoCr2O4/ZnO nanocomposites using Basella alba L. leaves extracts with enhanced photocatalytic degradation of malachite green in aqueous media. Chemosphere 2024, 352, 141215. [Google Scholar] [CrossRef]
- Reddy, M.T.; Begum, H.; Sunil, N.; Rao, P.S.; Kumar, S. Preliminary characterization and evaluation of landraces of indian spinach (Basella spp. L.) for agro-economic and quality traits. Plant Breed. Biotechnol. 2014, 2, 48–63. [Google Scholar] [CrossRef]
- Kumar, S.S.; Manoj, P.; Giridhar, P. Nutrition facts and functional attributes of foliage of Basella spp. LWT 2015, 64, 468–474. [Google Scholar] [CrossRef]
- Lourith, N.; Kanlayavattanakul, M. Ceylon spinach: A promising crop for skin hydrating products. Ind. Crops Prod. 2017, 105, 24–28. [Google Scholar] [CrossRef]
- Kuhnlein, H.V. Finding food sources of vitamin A and provitamin A. Food Nutr. Bull. 2000, 21, 130–134. [Google Scholar] [CrossRef]
- Palanuvej, C.; Hokputsa, S.; Tunsaringkarn, T.; Ruangrungsi, N. In vitro glucose entrapment and alpha-glucosidase inhibition of mucilaginous substances from selected Thai medicinal plants. Sci. Pharm. 2009, 77, 837–849. [Google Scholar] [CrossRef]
- Siriwatanametanon, N.; Fiebich, B.L.; Efferth, T.; Prieto, J.M.; Heinrich, M. Traditionally used Thai medicinal plants: In vitro anti-inflammatory, anticancer and antioxidant activities. J. Ethnopharmacol. 2010, 130, 196–207. [Google Scholar] [CrossRef]
- Saroj, V.; Rao, P.; Rao, S.; Krunal, S.; Shah, C. Pharmacognostical study of Basella alba stem. Int. J. Res. Pharm. Biomed. Sci. 2012, 3, 1093–1094. [Google Scholar]
- Lin, S.M.; Lin, B.H.; Hsieh, W.M.; Ko, H.J.; Liu, C.D.; Chen, L.G.; Chiou, R.Y.Y. Structural identification and bioactivities of red-violet pigments present in Basella alba fruits. J. Agric. Food Chem. 2010, 58, 10364–10372. [Google Scholar] [CrossRef] [PubMed]
- Ashaduzzaman, N.M.; Khan, M.; Satter, M.A.; Rahman, M.M.; Jashim uddin, M.; Amin, M.Z. Assessment of physicochemical properties, nutrient contents and colorant stability of the two varieties of Malabar spinach (Basella alba L.) fruits. Biocatal. Agric. Biotechnol. 2023, 51, 102746. [Google Scholar] [CrossRef]
- Munteanu, I.G.; Apetrei, C. Analytical methods used in determining antioxidant activity: A review. Int. J. Mol. Sci. 2021, 22, 3380. [Google Scholar] [CrossRef]
- Kozioł, L.; Knap, M.; Sutor-Świeży, K.; Górska, R.; Dziedzic, E.; Bieniasz, M.; Mielczarek, P.; Popenda, Ł.; Tyszka-Czochara, M.; Wybraniec, S. Identification and reactivity of pigments in prominent vegetable leaves of Basella alba L. var. ’Rubra’ (Malabar spinach). Food chem. 2024, 445, 138714. [Google Scholar] [CrossRef] [PubMed]
- Jagarwal, P.; Vijayvergia, R.; Kumar, D. Isolation, identification and characterization of flavonoids from Basella alba L. Eur. J. Med. Plants. 2021, 32, 28–38. [Google Scholar] [CrossRef]
- Munteanu; Candoken, E.; Akev, N. Implications for degenerative disorders: Antioxidative activity, total phenols, flavonoids, ascorbic acid, beta-carotene and beta-tocopherol in Aloe vera. Oxid. Med. Cell. Longev. 2014, 2, 99. [Google Scholar] [CrossRef]
- De Souza, A.S.N.; de Oliveira Schmidt, H.; Pagno, C.; Rodrigues, E.; da Silva, M.A.S.; Flores, S.H.; de Oliveira Rios, A. Influence of cultivar and season on carotenoids and phenolic compounds from red lettuce influence of cultivar and season on lettuce. Food Res. Int. 2022, 155, 111110. [Google Scholar] [CrossRef] [PubMed]
- López, A.; Javier, G.A.; Fenoll, J.; Hellı’n, P.; Flores, P. Chemical composition and antioxidant capacity of lettuce: Comparative study of regular-sized (Romaine) and baby-sized (Little Gem and Mini Romaine) types. J. Food Compost. Anal. 2014, 33, 39–48. [Google Scholar] [CrossRef]
- Al-Karaki, G.N.; Othman, Y. Effect of foliar application of amino acid biostimulants on growth, macronutrient, total phenol contents and antioxidant activity of soilless grown lettuce cultivars. S. Afr. J. Bot. 2023, 154, 225–231. [Google Scholar] [CrossRef]
- Martinetti, L.; Ferrante, A.; Bassoli, A.; Borgonovo, G.; Tosca, A.; Spoleto, P. Characterization of some qualitative traits in different perilla cultivars. Acta Hort. 2012, 939, 301–308. [Google Scholar] [CrossRef]
- Gu, H.H.; Wang, J.S.; Zhao, Z.Q.; Sheng, X.G.; Yu, H.F.; Huang, W.B. Characterization of the appearance, health-promoting compounds, and antioxidant capacity of the florets of the loose-curd cauliflower. Int. J. Food Prop. 2014, 18, 392–402. [Google Scholar] [CrossRef]
- Sun, B.; Tian, Y.X.; Jiang, M.; Yuan, Q.; Chen, Q.; Zhang, Y.; Luo, Y.; Zhang, F.; Tang, H.R. Variation in the main health-promoting compounds and antioxidant activity of whole and individual edible parts of baby mustard Brassica juncea var. gemmifera. RSC Adv. 2018, 8, 33845–33854. [Google Scholar] [CrossRef]
- Arnáiz, E.; Bernal, J.; Martín, M.T.; Nozal, M.J.; Bernal, J.L.; Toribio, L. Supercritical fluid extraction of free amino acids from broccoli leaves. J. Chromatogr. A 2012, 1250, 49–53. [Google Scholar] [CrossRef] [PubMed]
- Campas-Baypoli, O.N.; Sánchez-Machado, D.I.; Bueno-Solano, C.; Núñez-Gastélum, J.A.; Reyes-Moreno, C.; López-Cervantes, J. Biochemical composition and physicochemical properties of broccoli flours. Int. J. Food Sci. Nutr. 2009, 60, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, A.; Sharma, U.; Pal Vig, A.; Singh, B.; Arora, S. Free radical scavenging, antiproliferative activities and profiling of variations in the level of phytochemicals in different parts of broccoli (Brassica oleracea italica). Food Chem. 2014, 148, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Le, T.N.; Sakulsataporn, N.; Chiu, C.H.; Hsieh, A.P. Polyphenolic profile and varied bioactivities of processed taiwanese grown broccoli: A comparative study of edible and non-edible parts. Pharmaceuticals 2020, 13, 82. [Google Scholar] [CrossRef] [PubMed]
- Pajak, P.; Socha, R.; Gałkowska, D.; Roznowski, J.; Fortuna, T. Phenolic profile and antioxidant activity in selected seeds and sprouts. Food Chem. 2014, 143, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.S.; Manoj, P.; Giridhar, P. Fourier transform infrared spectroscopy (FTIR) analysis, chlorophyll content and antioxidant properties of native and defatted foliage of green leafy vegetables. J. Food Sci. Technol. 2015, 52, 8131–8139. [Google Scholar] [CrossRef] [PubMed]
- Hounsome, N.; Hounsome, B.; Tomos, D.; Edwards-jones, G. Plant metabolites and nutritional quality of vegetables. J. Food Sci. 2008, 73, R48–R65. [Google Scholar] [CrossRef]
- Yang, W.; Qian, G.T.; Chen, Y.L.; Liu, T.X.; Wan, H.H.; Wang, S.F.; Meng, X.X.; Chen, W.Q.; Wahab, A.T.; Su, Y.; et al. Profiling of polyphenols for in-depth understanding of Tartary buckwheat sprouts: Correlation between cultivars and active components, dynamic changes and the effects of ultraviolet B stress. Food Chem. 2022, 14, 100295. [Google Scholar] [CrossRef]
- Zou, L.; Tan, W.K.; Du, Y.Y.; Lee, H.W.; Liang, X.; Lei, J.J.; Striegel, L.; Weber, N.; Rychlik, M.; Ong, C.N. Nutritional metabolites in Brassica rapa subsp. chinensis var. parachinensis (choy sum) at three different growth stages: Microgreen, seedling and adult plant. Food Chem. 2021, 357, 129535. [Google Scholar] [CrossRef]
- Ashenaf, E.L.; Nyman, M.C.; Holley, J.M.; Mattson, N.S.; Rangarajan, A. Phenotypic plasticity and nutritional quality of three kale cultivars (Brassica oleracea L. var. acephala) under field, greenhouse, and growth chamber environments. Environ. Exp. Bot. 2022, 199, 104895. [Google Scholar] [CrossRef]
- Drabińska, N.; Jez, M.; Nogueira, M. Variation in the accumulation of phytochemicals and their bioactive properties among the aerial parts of cauliflower. Antioxidants 2021, 10, 1597. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.T.; Di, H.M.; Cheng, W.J.; Ren, G.R.; Luo, S.; Ma, J.; Ma, W.; Lian, H.S.; Li, X.M.; Huang, Z.; et al. Variation in the main health-promoting compounds and antioxidant activity of different edible parts of purple flowering stalks (Brassica campestris var. purpuraria) and green flowering stalks (Brassica campestris var. campestris). Plants 2022, 11, 1664. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, F.; Kato, M.; Hyodo, H.; Ikoma, Y.; Sugiura, M.; Yano, M. Ascorbate metabolisom in harvested broccoli. J. Exp. Bot. 2003, 54, 2439–2448. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Q.; Hu, L.P.; Liu, G.M.; Zhang, D.S.; He, H.J. Evaluation of the nutritional quality of chinese kale (Brassica alboglabra Bailey) using UHPLC-Quadrupole-Orbitrap MS/MS-based metabolomics. Molecules 2017, 22, 1262. [Google Scholar] [CrossRef] [PubMed]
- Brown, P.D.; Tokuhisa, J.G.; Reichelt, M.; Gershenzon, J. Variation of glucosinolate accumulation among different organs and developmental stages of Arabidopsis thaliana. Phytochemistry 2003, 62, 471–481. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Liang, Y.; Li, X.; Kang, H. Antioxidant extract from cauliflower leaves effectively improve the stability of pork patties during refrigerated storage. J. Food Process. Preserv. 2020, 44, e14510. [Google Scholar] [CrossRef]
- Chavan, J.J.; Santa-Catarina, C. Multidirectional insights into nutritional, phytochemical, antioxidant capability and multivariate analysis of underutilized edible berry plant (Salacia macrosperma Wight)—A novel source for food and pharmaceutical industry. Food Chem. Adv. 2023, 2, 100284. [Google Scholar] [CrossRef]
- Chen, C.X. Pigments in Fruits and Vegetables: Genomics and Dietetics; Springer: New York, NY, USA, 2015; pp. 11–12. [Google Scholar]
- Diamanti, J.; Capocasa, F.; Denoyes, B.; Petit, A.; Chartier, P.; Faedi, W.; Maltoni, M.L.; Battino, M.; Mezzetti, B. Standardized method for evaluation of strawberry (Fragaria × ananassa Duch.) germplasm collections as a genetic resource for fruit nutritional compounds. J. Food Compos. Anal. 2012, 28, 170–178. [Google Scholar] [CrossRef]
- Pineli, L.L.O.; Moretti, C.L.; Santos, M.S.; Campos, A.B.; Brasileiro, A.V.; Córdova, A.C.; Chiarello, M.D. Antioxidants and other chemical and physical characteristics of two strawberry cultivars at different ripeness stages. J. Food Compost. Anal. 2011, 125, 348–354. [Google Scholar] [CrossRef]
- Duma, M.; Alsina, I.; Dubova, L.; Erdberga, I. Bioactive compounds in tomatoes at different stages of maturity. Proc. Latv. Acad. Sci. Sect. B 2018, 72, 85–90. [Google Scholar] [CrossRef]
- Taylor, L.; Nunes-Nesi, A.; Parsley, K.; Leiss, A.; Leach, G.; Coates, S.; Wingler, A.; Fernie, A.R.; Hibberd, J.M. Cytosolic pyruvate, orthophosphate dikinase functions in nitrogen remobilization during leaf senescence and limits individual seed growth and nitrogen content. Plant J. 2010, 62, 641–652. [Google Scholar] [CrossRef] [PubMed]
- Vom Dorp, K.; Hölzl, G.; Plohmann, C.; Eisenhut, M.; Abraham, M.; Weber, A.P.M.; Hanson, A.D.; Dörmann, P. Remobilization of phytol from chlorophyll degradation is essential for tocopherol synthesis and growth of Arabidopsis. Plant Cell. 2015, 27, 2846–2859. [Google Scholar] [CrossRef]
- Bordiga, M.; Travaglia, F.; Giuffrida, D.; Mangraviti, D.; Rigano, F.; Mondello, L.; Arlorio, M.; Coïsson, J.D. Characterization of peel and pulp proanthocyanidins and carotenoids during ripening in persimmon “Kaki Tipo” cv, cultivated in Italy. Food Res. Int. 2019, 120, 800–809. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.Q.; Ohlander, M.; Jeppsson, N.; Björk, L.; Trajkovski, V. Changes in antioxidant effects and their relationship to phytonutrients in fruits of sea buckthorn (Hippophae rhamnoides L.) during maturation. J. Agric. Food Chem. 2000, 48, 1485–1490. [Google Scholar] [CrossRef]
- Zhong, R.H.; Wei, J.B.; Liu, B.; Luo, H.H.; Zhang, Z.Q.; Pang, X.Q.; Fang, F. Metabolite and transcriptome profiles of proanthocyanidin biosynthesis in the development of litchi fruit. Int. J. Mol. Sci. 2023, 24, 532. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.R.; Yeap Foo, L. Antioxidant and radical scavenging activities of polyphenols from apple pomace. Food Chem. 2000, 68, 81–85. [Google Scholar] [CrossRef]
- Bordiga, M.; Travaglia, F.; Locatelli, M.; Coïsson, J.D.; Arlorio, M. Characterisation of polymeric skin and seed proanthocyanidins during ripening in six Vitis vinifera L. cv. Food Chem. 2011, 127, 180–187. [Google Scholar] [CrossRef]
- El-Beltagi, H.S.; Shah, S.T.; Mohamed, H.I.; Alam, N.; Sajid, M.; Khan, A.; Basit, A. Physiological response, phytochemicals, antioxidant, and enzymatic activity of date palm (Phoenix dactylifera L.) cultivated under different storage time, harvesting stages, and temperatures. Saudi J. Biol. Sci. 2023, 30, 103818. [Google Scholar] [CrossRef]
- Rastegar, S.; Rahemi, M.; Baghizadeh, A.; Gholami, M. Enzyme activity and biochemical changes of three date palm cultivars with different softening pattern during ripening. Food Chem. 2012, 134, 1279–1286. [Google Scholar] [CrossRef]
- García-Gómez, B.E.; Ruiz, D.; Salazar, J.A.; Rubio, M.; Martínez-García, P.J.; Martinez-Gomez, P. Analysis of metabolites and gene expression changes relative to apricot (Prunus armeniaca L.) fruit quality during development and ripening. Front. Plant Sci. 2020, 11, 1269. [Google Scholar] [CrossRef]
- Fenn, M.A.; Giovannoni, J.J. Phytohormones in fruit development and maturation. Plant J. 2021, 105, 446–458. [Google Scholar] [CrossRef]
- Liyana-Pathirana, C.M.; Shahidi, F.; Alasalvar, C. Antioxidant activity of cherry laurel fruit (Laurocerasus officinalis Roem.) and its concentrated juice. Food Chem. 2006, 99, 121–128. [Google Scholar] [CrossRef]
- Bradford, M. Arapid and sensitive method for the quantization of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Yuan, G.F.; Sun, B.; Yuan, J.; Wang, Q.M. Effect of 1-methylcyclopropene on shelf life, visual quality, antioxidant enzymes and health-promoting compounds in broccoli florets. Food Chem. 2010, 118, 774–781. [Google Scholar] [CrossRef]
- Prior, R.L.; Fan, E.; Ji, H.P.; Howell, A.; Nio, C.; Payne, M.J.; Reed, J. Multi-laboratory validation of a standard method for quantifying proanthocyanidins in cranberry powders. J. Sci. Food Agric. 2010, 90, 1473–1478. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of ‘‘antioxidant power’’: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Subhasree, B.; Baskar, R.; Keerthana, R.L.; Susan, R.L.; Rajasekaran, P. Evaluation of antioxidant potential in selected green leafy vegetables. Food Chem. 2009, 115, 1213–1220. [Google Scholar] [CrossRef]
- Sun, B.; Di, H.M.; Zhang, J.Q.; Xia, P.X.; Huang, W.L.; Jian, Y.; Zhang, C.L.; Zhang, F. Effect of light on sensory quality, health-promoting phytochemicals and antioxidant capacity in post-harvest baby mustard. Food Chem. 2021, 339, 128057. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | VC/VP | VO/VP | VCO/VP |
|---|---|---|---|
| Total soluble sugar | 0.108 ** | 0.724 ** | 0.157 ** |
| Total soluble protein | 0.499 ** | 0.206 ** | 0.256 ** |
| Total chlorophyll | 0.006 ** | 0.964 ** | 0.028 ** |
| Total carotenoids | 0.006 ** | 0.961 ** | 0.015 ** |
| Total ascorbic acid | 0.104 ** | 0.601 ** | 0.265 ** |
| Total proanthocyanidins | 0.033 ** | 0.901 ** | 0.062 ** |
| Total flavonoids | 0.011 ** | 0.857 ** | 0.122 ** |
| Total phenolics | 0.097 ** | 0.759 ** | 0.129 ** |
| FRAP | 0.091 ** | 0.766 ** | 0.128 ** |
| ABTS | 0.049 ** | 0.818 ** | 0.121 ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Cheng, W.; Di, H.; Yang, S.; Tian, Y.; Tong, Y.; Huang, H.; Escalona, V.H.; Tang, Y.; Li, H.; et al. Variation in Nutritional Components and Antioxidant Capacity of Different Cultivars and Organs of Basella alba. Plants 2024, 13, 892. https://doi.org/10.3390/plants13060892
Zhang Y, Cheng W, Di H, Yang S, Tian Y, Tong Y, Huang H, Escalona VH, Tang Y, Li H, et al. Variation in Nutritional Components and Antioxidant Capacity of Different Cultivars and Organs of Basella alba. Plants. 2024; 13(6):892. https://doi.org/10.3390/plants13060892
Chicago/Turabian StyleZhang, Yi, Wenjuan Cheng, Hongmei Di, Shihan Yang, Yuxiao Tian, Yuantao Tong, Huanhuan Huang, Victor Hugo Escalona, Yi Tang, Huanxiu Li, and et al. 2024. "Variation in Nutritional Components and Antioxidant Capacity of Different Cultivars and Organs of Basella alba" Plants 13, no. 6: 892. https://doi.org/10.3390/plants13060892
APA StyleZhang, Y., Cheng, W., Di, H., Yang, S., Tian, Y., Tong, Y., Huang, H., Escalona, V. H., Tang, Y., Li, H., Zhang, F., Sun, B., & Huang, Z. (2024). Variation in Nutritional Components and Antioxidant Capacity of Different Cultivars and Organs of Basella alba. Plants, 13(6), 892. https://doi.org/10.3390/plants13060892