


Phytochemical Screening, Antioxidant Effect and Sperm Quality of the Bomba ceiba Stamen Extracts on Charolais Cattle Sperm Induced by Ferrous Sulfate


Abstract
:1. Introduction
2. Results
2.1. Phytochemical Contents
2.2. Phytochemical Screening Using Liquid Chromatography Quadrupole Time-of-Flight Mass Spectrometry (LC-QTOF-MS)
2.3. Antioxidant Properties in Cell-Free System
2.4. Cytotoxicity Analysis
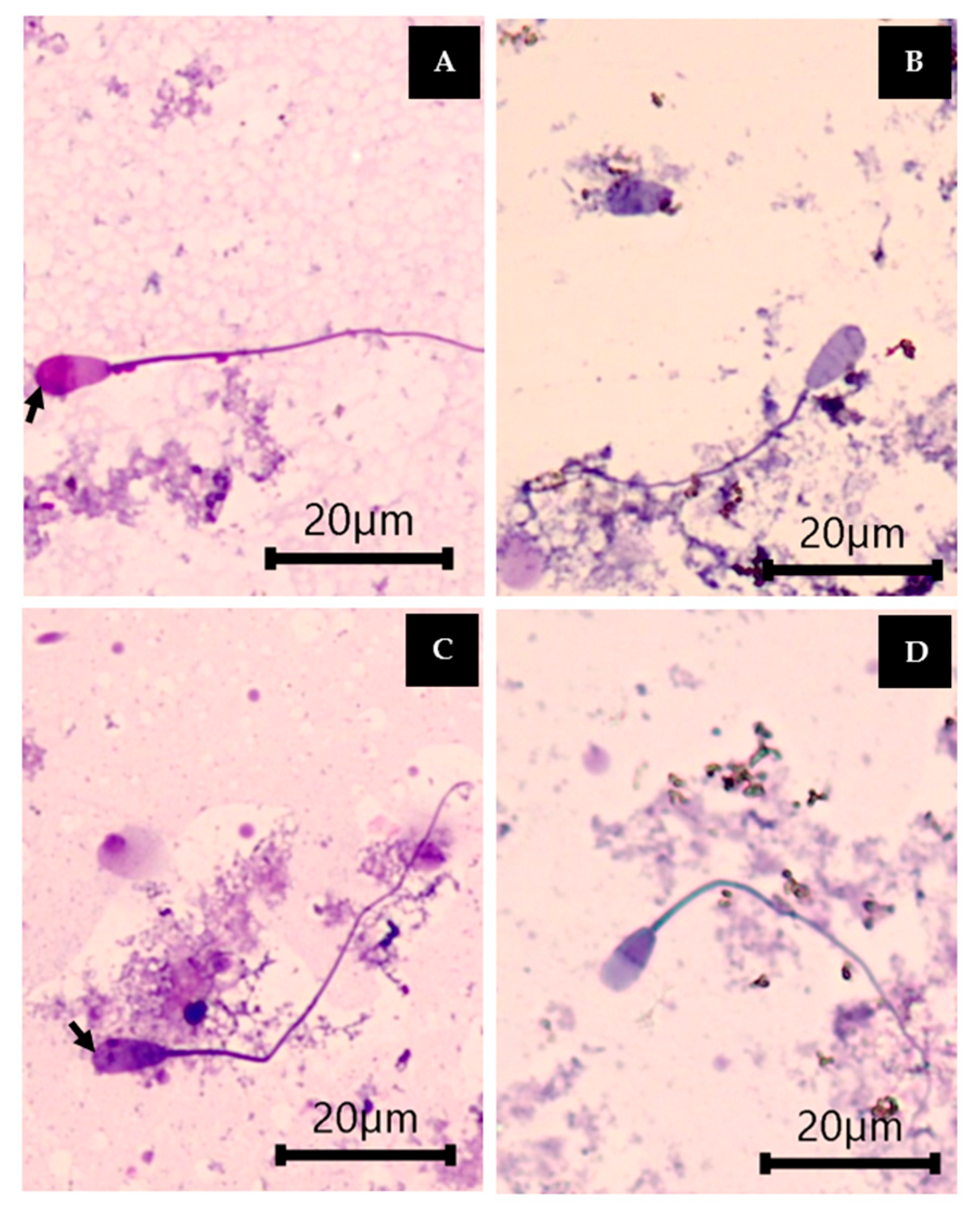
2.5. Sperm Quality Analysis
2.5.1. Sperm Motility
2.5.2. Sperm Viability and Acrosome Integrity
2.5.3. Sperm Morphology
2.6. Antioxidant Properties of Sperm
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Plant Collection
4.3. Plant Extraction
- The decoction method is a traditional extraction method used in recipes and traditional medicine. The B. ceiba stamen hot aqueous extract (BAE) was prepared by soaking dried samples in hot distilled water at 75–80 °C for 3–5 min [13];
- Ultrasonic extraction is a modern method that uses ultrasound energy and solvents to extract target compounds from natural products. The B. ceiba stamen ultrasonic extract (BUE) was prepared by soaking dried samples in distilled water and subjecting them to ultrasonication in a water bath (Elma, Singen, Germany) at 50/60 Hz and 40 °C for 30 min [58];
- Microwave-assisted extraction is a modern method that uses electromagnetic energy to break the cell membrane of plant cells. The B. ceiba stamen microwave-assisted extract (BMAE) was prepared by soaking dried samples in distilled water and pre-heating them using a magnetic stirrer at room temperature for 90 min. Then, suspensions were irradiated with microwaves (Milestone ETHOS UP, Sorisole, Italy) two times as follows: 1 min power on (heating to the desired temperature of about 70–80 °C) and 6 min power off [59].
4.4. Phytochemical Contents
4.4.1. Total Phenolic Content
4.4.2. Total Flavonoid Content
4.4.3. Total Tannin Content
4.4.4. Total Monomeric Anthocyanin Content
4.4.5. Lycopene Content
4.5. Phytochemical Screening of Extract Using LC-QTOF-MS
4.6. Antioxidant Properties in Cell-Free System
4.6.1. 2,2-Diphenyl-1-Picrylhydrazyl (DPPH) Radical Scavenging Assay
4.6.2. 2,2′-Azino-di-[3-Ethylbenzthiazoline Sulfonate (ABTS) Radical Scavenging Assay
4.6.3. Lipid Peroxidation (LPO) Assay
4.6.4. Inhibition of Formation of Advance Oxidation Protein Products (AOPPs)
4.6.5. Inhibition of Formation of Advance Glycation End Products (AGEs)
4.6.6. Ferric-Reducing Antioxidant Power (FRAP) Assay
4.7. Cytotoxicity Analysis
4.8. Experimental Design
4.9. Analysis of Sperm Characteristics
4.9.1. Sperm Motility
4.9.2. Sperm Viability and Acrosome Integrity
4.9.3. Sperm Morphology
4.10. Antioxidant Properties in Sperm
4.10.1. Lipid Peroxidation (LPO) Assay
4.10.2. Inhibition of Formation of Advance Oxidation Protein Products (AOPPs)
4.10.3. Inhibition of Formation of Advance Glycation End Products (AGEs)
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arafa, A.; Foda, D.; Mahmoud, A.; Metwally, N.; Farrag, A. Bombax ceiba flowers extract ameliorates hepatosteatosis induced by ethanol and relatively moderate fat diet in rats. Toxicol. Rep. 2019, 6, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Kriintong, N.; Katisart, T. In vitro antioxidant and antidiabetic activities of leaf and flower extracts from Bombax ceiba. Pharmacogn. Res. 2020, 12, 194–198. [Google Scholar]
- Guang-Kai, X.; Xiao-Ying, Q.; Guo-Kai, W.; Guo-Yong, X.; Xu-Sen, L.; Chen-Yu, S.; Bao-Lin, L.; Min-Jian, Q. Antihyperglycemic, antihyperlipidemic and antioxidant effects of standard ethanol extract of Bombax ceiba leaves in high-fat-diet-and streptozotocin-induced Type 2 diabetic rats. Chin. J. Nat. Med. 2017, 15, 168–177. [Google Scholar]
- Thongmee, O.; Rodhetbhai, C.; Siltragool, W. Lanna food: The cultural management strategy for the creative economy development. HASSS 2015, 15, 105–120. [Google Scholar]
- Yasien, S.; Iqbal, M.M.; Javed, M.; Alnuwaiser, M.A.; Iqbal, S.; Mahmood, Q.; Elkaeed, E.B.; Dera, A.A.; Alrbyawi, H.; Pashameah, R.A. Comparative evaluation of various extraction techniques for secondary metabolites from Bombax ceiba L. flowering plants along with in vitro anti-diabetic performance. Bioengineering 2022, 9, 486. [Google Scholar] [CrossRef] [PubMed]
- Bhargava, C.; Thakur, M.; Yadav, S. Effect of Bombax ceiba L. on spermatogenesis, sexual behaviour and erectile function in male rats. Andrologia 2012, 44, 474–478. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.S.; Verma, N.K.; Rathore, B.; Nayak, G.; Singhai, A.K.; Singh, P. Cardioprotective effect of Bombax ceiba flowers against acute adriamycin-induced myocardial infarction in rats. Rev. Bras. Farmacogn. 2011, 21, 704–709. [Google Scholar] [CrossRef]
- Huang, S.K.H.; Hsieh, C.Y.; Fu, P.W.; Lee, C.J.; Domingo, G.C.; Alimboyoguen, A.B.; Lin, P.Y.; Hung, L.C.; Roxas, T.J.R.; De Castro-Cruz, K.A. Phytochemical constituent analysis, antioxidative effect, and anti-inflammatory activity of Bombax ceiba flowers. Biointerface Res. Appl. Chem. 2023, 13, 408. [Google Scholar]
- Davis, T. Stamen number and pollen size in levo-and dextro-rotary flowers of Bombacaceae. Rev. Palaeobot. Palynol. 1967, 3, 133–139. [Google Scholar] [CrossRef]
- Chaudhary, P.H.; Tawar, M.G. Pharmacognostic and phytopharmacological overview on Bombax ceiba. Syst. Rev. Pharm. 2019, 10, 20–25. [Google Scholar] [CrossRef]
- Wang, G.K.; Lin, B.B.; Rao, R.; Zhu, K.; Qin, X.Y.; Xie, G.Y.; Qin, M.J. A new lignan with anti-HBV activity from the roots of Bombax ceiba. Nat. Prod. Res. 2013, 27, 1348–1352. [Google Scholar] [CrossRef] [PubMed]
- Joshi, K.R.; Devkota, H.P.; Yahara, S. Chemical analysis of flowers of Bombax ceiba from Nepal. Nat. Prod. Commun. 2013, 8, 583–584. [Google Scholar] [CrossRef]
- Laoung-on, J.; Jaikang, C.; Saenphet, K.; Sudwan, P. Phytochemical screening, antioxidant and sperm viability of Nelumbo nucifera petal extracts. Plants 2021, 10, 1375. [Google Scholar] [CrossRef]
- Laoung-On, J.; Jaikang, C.; Saenphet, K.; Sudwan, P. Effect of Nelumbo nucifera petals extract on antioxidant activity and sperm quality in Charolais cattle sperm induced by mancozeb. Plants 2022, 11, 637. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhang, M.; Haenen, G.R.; Vervoort, L.; Moalin, M. Flavonoids seen through the energy perspective. Int. J. Mol. Sci. 2021, 23, 187. [Google Scholar] [CrossRef] [PubMed]
- Martin, L.J.; Touaibia, M. Improvement of testicular steroidogenesis using flavonoids and isoflavonoids for prevention of late-onset male hypogonadism. Antioxidants 2020, 9, 237. [Google Scholar] [CrossRef] [PubMed]
- Saleem, R.; Ahmad, S.I.; Ahmed, M.; Faizi, Z.; Zikr-ur-Rehman, S.; Ali, M.; Faizi, S. Hypotensive activity and toxicology of constituents from Bombax ceiba stem bark. Biol. Pharm. Bull. 2003, 26, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Bisht, S.; Faiq, M.; Tolahunase, M.; Dada, R. Oxidative stress and male infertility. Nat. Rev. Urol. 2017, 14, 470–485. [Google Scholar] [CrossRef] [PubMed]
- Kaltsas, A. Oxidative stress and male infertility: The protective role of antioxidants. Medicina 2023, 59, 1769. [Google Scholar] [CrossRef] [PubMed]
- Darbandi, M.; Darbandi, S.; Agarwal, A.; Sengupta, P.; Durairajanayagam, D.; Henkel, R.; Sadeghi, M.R. Reactive oxygen species and male reproductive hormones. Reprod. Biol. Endocrinol. 2018, 16, 87. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Virk, G.; Ong, C.; Du Plessis, S.S. Effect of oxidative stress on male reproduction. World J. Mens Health 2014, 32, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Bansal, A.; Bilaspuri, G. Effect of ferrous sulphate and ascorbic acid on motility, viability and lipid peroxidation of crossbred cattle bull spermatozoa. Animal 2008, 2, 100–104. [Google Scholar] [CrossRef] [PubMed]
- Knazicka, Z.; Duranova, H.; Fialkova, V.; Miskeje, M.; Jambor, T.; Makarevich, A.V.; Roychoudhury, S.; Kovacik, A.; Massanyi, P.; Lukac, N. In vitro effect of ferrous sulphate on bovine spermatozoa motility parameters, viability and Annexin V-labeled membrane changes. PLoS ONE 2021, 16, e0257766. [Google Scholar] [CrossRef]
- Sedha, S.; Kumar, S.; Shukla, S. Role of oxidative stress in male reproductive dysfunctions with reference to phthalate compounds. Urol. J. 2015, 12, 2304–2316. [Google Scholar] [PubMed]
- Sudwan, P.; Saenphet, K.; Aritajat, S.; Sitasuwan, N. Effects of Boesenbergia rotunda (L.) Mansf. on sexual behaviour of male rats. Asian J. Androl. 2007, 9, 849–855. [Google Scholar] [CrossRef] [PubMed]
- Sudawadee, Y.; Vipavadee, C.; Kanokporn, S.; Paiwan, S. Effects of Boesenbergia rotunda juice on sperm qualities in male rats. J. Med. Plant. Res. 2011, 5, 3861–3867. [Google Scholar]
- Sudwan, P.; Saenphet, K.; Saenphet, S.; Wongsawad, C. Sperm density and ultrastructure of Sertoli cells in male rats treated with Kaempferia parviflora Wall. Ex Baker extract. Southeast. Asian J. Trop. Med. Public Health 2007, 38, 249. [Google Scholar]
- Laoung-On, J.; Saenphet, K.; Jaikang, C.; Sudwan, P. Effect of Moringa oleifera Lam. leaf tea on sexual behavior and reproductive function in male rats. Plants 2021, 10, 2019. [Google Scholar] [CrossRef] [PubMed]
- Alara, O.R.; Abdurahman, N.H.; Ukaegbu, C.I. Extraction of phenolic compounds: A review. Curr. Res. Food Sci. 2021, 4, 200–214. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Rangel, J.C.; Benavides, J.; Heredia, J.B.; Cisneros-Zevallos, L.; Jacobo-Velázquez, D.A. The Folin–Ciocalteu assay revisited: Improvement of its specificity for total phenolic content determination. Anal. Methods 2013, 5, 5990–5999. [Google Scholar] [CrossRef]
- Thavamoney, N.; Sivanadian, L.; Tee, L.H.; Khoo, H.E.; Prasad, K.N.; Kong, K.W. Extraction and recovery of phytochemical components and antioxidative properties in fruit parts of Dacryodes rostrata influenced by different solvents. J. Food Sci. Technol. 2018, 55, 2523–2532. [Google Scholar] [CrossRef] [PubMed]
- Naczk, M.; Shahidi, F. Phenolics in cereals, fruits, and vegetables: Occurrence, extraction, and analysis. J. Pharm. Biomed. Anal. 2006, 41, 1523–1542. [Google Scholar] [CrossRef] [PubMed]
- Haslam, E. Natural polyphenols (vegetable tannins) as drugs: Possible modes of action. J. Nat. Prod. 1996, 59, 205–215. [Google Scholar] [CrossRef] [PubMed]
- Oancea, S.; Stoia, M.; Coman, D. Effects of extraction conditions on bioactive anthocyanin content of Vaccinium corymbosum in the perspective of food applications. Proc. Eng. 2012, 42, 489–495. [Google Scholar] [CrossRef]
- Zhang, M.; Zhao, J.; Dai, X.; Li, X. Extraction and analysis of chemical compositions of natural products and plants. Separations 2023, 10, 598. [Google Scholar] [CrossRef]
- Ghenabzia, I.; Hemmami, H.; Ilham, B.; Zeghoud, S.; Seghir, B.B.; Hammoudi, R. Different methods of extraction of bioactive compounds and their effect on biological activity: A review. Int. J. Second. Metab. 2023, 10, 469–494. [Google Scholar] [CrossRef]
- Song, Z.; Xiong, X.; Huang, G. Ultrasound-assisted extraction and characteristics of maize polysaccharides from different sites. Ultrason. Sonochem. 2023, 95, 106416. [Google Scholar] [CrossRef]
- Akhtar, I.; Javad, S.; Yousaf, Z.; Iqbal, S.; Jabeen, K. Microwave assisted extraction of phytochemicals an efficient and modern approach for botanicals and pharmaceuticals. Pak. J. Pharm. Sci. 2019, 32, 223–230. [Google Scholar] [PubMed]
- Yadav, N.; Sharma, S.; Joys, J.S.; Kumar, S. Microwave assisted extraction of bioactive compounds: A brief review. J. Indian Chem. Soc. 2020, 97, 1751–1756. [Google Scholar]
- Yang, Y.-C.; Wei, M.-C. Development and characterization of a green procedure for apigenin extraction from Scutellaria barbata D. Don. Food Chem. 2018, 252, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Antony, A.; Farid, M. Effect of temperatures on polyphenols during extraction. Appl. Sci. 2022, 12, 2107. [Google Scholar] [CrossRef]
- Akowuah, G.; Mariam, A.; Chin, J. The effect of extraction temperature on total phenols and antioxidant activity of Gynura procumbens leaf. Pharmacogn. Mag. 2009, 5, 81–85. [Google Scholar]
- Vo, T.P.; Pham, N.D.; Pham, T.V.; Nguyen, H.Y.; Tran, T.N.H.; Tran, T.N.; Nguyen, D.Q. Green extraction of total phenolic and flavonoid contents from mangosteen (Garcinia mangostana L) rind using natural deep eutectic solvents. Heliyon 2023, 9, e14884. [Google Scholar] [CrossRef] [PubMed]
- Aprodu, I.; Milea, Ș.A.; Enachi, E.; Râpeanu, G.; Bahrim, G.E.; Stănciuc, N. Thermal degradation kinetics of anthocyanins extracted from purple maize flour extract and the effect of heating on selected biological functionality. Foods 2020, 9, 1593. [Google Scholar] [CrossRef]
- Chebil, L.; Humeau, C.; Anthoni, J.; Dehez, F.; Engasser, J.-M.; Ghoul, M. Solubility of flavonoids in organic solvents. J. Chem. Eng. Data 2007, 52, 1552–1556. [Google Scholar] [CrossRef]
- Purdy, P.H.; Ericsson, S.A.; Dodson, R.E.; Sternes, K.L.; Garner, D.L. Effects of the flavonoids, silibinin and catechin, on the motility of extended cooled caprine sperm. Small Rumin. Res. 2004, 55, 239–243. [Google Scholar] [CrossRef]
- Gungor, S.; Inanc, M.E.; Ozturk, C.; Korkmaz, F.; Bastan, I.; Cil, B.; Kastelic, J.P. Gallic and carnosic acids improve quality of frozen-thawed ram spermatozoa. Andrologia 2019, 51, e13393. [Google Scholar] [CrossRef]
- Gandhi, G.R.; Vasconcelos, A.B.S.; Wu, D.-T.; Li, H.-B.; Antony, P.J.; Li, H.; Geng, F.; Gurgel, R.Q.; Narain, N.; Gan, R.-Y. Citrus flavonoids as promising phytochemicals targeting diabetes and related complications: A systematic review of in vitro and in vivo studies. Nutrients 2020, 12, 2907. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhao, K.; Jiang, K.; Tao, S.; Li, Y.; Chen, W.; Kou, S.; Gu, C.; Li, Z.; Guo, L. A review of flavonoids from cassia species and their biological activity. Curr. Pharm. Biotechnol. 2016, 17, 1134–1146. [Google Scholar] [CrossRef]
- Moretti, E.; Mazzi, L.; Terzuoli, G.; Bonechi, C.; Iacoponi, F.; Martini, S.; Rossi, C.; Collodel, G. Effect of quercetin, rutin, naringenin and epicatechin on lipid peroxidation induced in human sperm. Reprod. Toxicol. 2012, 34, 651–657. [Google Scholar] [CrossRef]
- Xu, D.; Wu, L.; Yang, L.; Liu, D.; Chen, H.; Geng, G.; Li, Q. Rutin protects boar sperm from cryodamage via enhancing the antioxidative defense. Anim. Sci. J. 2020, 91, e13328. [Google Scholar] [CrossRef] [PubMed]
- Miller, N.J.; Rice-Evans, C.A. Factors influencing the antioxidant activity determined by the ABTS•+ radical cation assay. Free Radic. Res. 1997, 26, 195–199. [Google Scholar] [CrossRef] [PubMed]
- Haminiuk, C.W.I.; Plata-Oviedo, M.S.V.; de Mattos, G.; Carpes, S.T.; Branco, I.G. Extraction and quantification of phenolic acids and flavonols from Eugenia pyriformis using different solvents. J. Food Sci. Technol. 2014, 51, 2862–2866. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, J.; Wang, S.; Ju, H.; Chen, S.; Basioura, A.; Ferreira-Dias, G.; Liu, Z.; Zhu, J. Post-thaw storage temperature Influenced boar sperm quality and lifespan through apoptosis and lipid peroxidation. Animals 2023, 14, 87. [Google Scholar] [CrossRef] [PubMed]
- Marbeuf-Gueye, C.; Stierle, V.; Sudwan, P.; Salerno, M.; Garnier-Suillerot, A. Perturbation of membrane microdomains in GLC4 multidrug-resistant lung cancer cells− modification of ABCC1 (MRP1) localization and functionality. FEBS J. 2007, 274, 1470–1480. [Google Scholar] [CrossRef] [PubMed]
- Opuwari, C.S.; Matshipi, M.N.; Phaahla, M.K.; Setumo, M.A.; Moraswi, R.T.; Zitha, A.A.; Offor, U.; Choma, S.S. Androgenic effect of aqueous leaf extract of Moringa oleifera on Leydig TM3 cells in vitro. Andrologia 2020, 52, e13825. [Google Scholar] [CrossRef] [PubMed]
- Tvrdá, E.; Benko, F.; Slanina, T.; du Plessis, S.S. The role of selected natural biomolecules in sperm production and functionality. Molecules 2021, 26, 5196. [Google Scholar] [CrossRef] [PubMed]
- Das, P.R.; Park, M.J.; Lee, C.M.; Nam, S.H.; Kim, Y.M.; Kim, D.i.; Eun, J.B. Aqueous green tea infusion extracted by ultra-sonication method, but not by conventional method, facilitates GLUT4 membrane translocation in adipocytes which potently ameliorates high-fat diet-induced obesity. J. Food Biochem. 2021, 45, e13561. [Google Scholar] [CrossRef]
- Pan, X.; Niu, G.; Liu, H. Microwave-assisted extraction of tea polyphenols and tea caffeine from green tea leaves. Chem. Eng. Process 2003, 42, 129–133. [Google Scholar] [CrossRef]
- Jeeno, P.; Yadoung, S.; Yana, P.; Hongsibsong, S. Phytochemical profiling and antioxidant capacity of traditional plants, northern Thailand. Plants 2023, 12, 3956. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Extraction | Total Phenolics (µg GAE/g Dried Weight) | Total Tannins (µg TAE/g Dried Weight) | Total Flavonoids (µg QE/g Dried Weight) | Total Monomeric Anthocyanins (µg Cyanidin-3-glucoside E/g Dried Weight) | Lycopene Content (×102 mg/g Dried Weight) | Yield (%) |
|---|---|---|---|---|---|---|---|
| B. ceiba | Decoction | 4.20 ± 0.30 a | 3.55 ± 0.26 a | 0.26 ± 0.02 b | 6.29 ± 0.42 a | 23.84 ± 22.76 | 10.99 ± 4.01 |
| Ultrasonicated | 5.03 ± 0.14 b | 4.29 ± 0.11 b | 0.13 ± 0.04 a | 7.51 ± 0.29 b | 21.44 ± 9.32 | 10.78 ± 4.18 | |
| Microwave-assisted | 4.11 ± 0.10 a | 3.46 ± 0.03 a | 0.10 ±0.05 a | 5.18 ± 0.44 c | 20.73 ± 6.95 | 10.82 ± 3.65 |
| No. | Compound | Formula | RT | Matching Score (%) | m/z | Mass | Mass Diff (db/ppm) |
|---|---|---|---|---|---|---|---|
| 1 | Catechin | C15 H14 O6 | 1.671 | 93.00 | 335.0786 | 290.0805 | 5.11 |
| 2 | Gallic acid | C7 H6 O5 | 2.121 | 92.11 | 169.0139 | 170.0215 | −2.90 |
| 3 | Luteolin 7-β-rutinoside | C27 H30 O15 | 2.388 | 91.81 | 593.1493 | 594.1566 | −3.20 |
| 4 | Kaempferol 3-rutinoside-7-rhamnoside | C33 H40 O19 | 3.488 | 94.04 | 785.2126 | 740.2143 | −2.75 |
| No. | Compound | Formula | RT | Matching Score (%) | m/z | Mass | Mass Diff (db/ppm) |
|---|---|---|---|---|---|---|---|
| 1 | Catechin | C15 H14 O6 | 1.681 | 90.51 | 335.0785 | 290.0802 | 4.16 |
| 2 | Gallic acid | C7 H6 O5 | 2.131 | 83.14 | 169.0139 | 170.0212 | −1.62 |
| 3 | Luteolin 7-β-rutinoside | C27 H30 O15 | 2.397 | 92.07 | 593.1494 | 594.1567 | −2.97 |
| 4 | Kaempferol 3-rutinoside-7-rhamnoside | C33 H40 O19 | 3.481 | 87.96 | 785.2119 | 740.2138 | −3.50 |
| 5 | Rutin | C27 H30 O16 | 3.698 | 85.17 | 609.1440 | 610.1513 | −3.41 |
| No. | Compound | Formula | RT | Matching Score (%) | m/z | Mass | Mass Diff (db/ppm) |
|---|---|---|---|---|---|---|---|
| 1 | Gallic acid | C7 H6 O5 | 2.138 | 98.47 | 169.0138 | 170.0211 | −2.65 |
| 2 | Luteolin 7-β-rutinoside | C27 H30 O15 | 2.455 | 93.31 | 593.1496 | 594.1568 | −2.80 |
| 3 | Kaempferol 3-rutinoside-7-rhamnoside | C33 H40 O19 | 3.556 | 92.77 | 785.2126 | 740.2144 | −2.73 |
| 4 | Rutin | C27 H30 O16 | 3.772 | 88.28 | 609.1441 | 610.1513 | −3.43 |
| Sample | Extraction | IC50 (mg/mL) | FRAB (mEq µmol Gallic Acid/L) | |||
|---|---|---|---|---|---|---|
| DPPH | ABTS | LPO | AGEs | |||
| B. ceiba | Decoction | 0.48 ± 0.02 | 0.94 ± 0.15 a | 2.34 ± 0.12 a | 33.59 ±0.97 | 9.97 ± 0.28 a |
| Ultrasonicated | 0.45 ± 0.05 | 0.54 ± 0.08 b | 1.36 ± 0.70 a | 35.35 ± 1.97 | 12.95 ± 0.18 b | |
| Microwave-assisted | 0.45 ± 0.06 | 0.62 ± 0.06 b | 7.30 ± 1.66 b | 30.35 ± 5.13 | 11.54 ± 0.50 c | |
| Group | Concentration (µg/mL) | Number of Sperm Exhibiting Motility | Number of Sperm Exhibiting Non-Motility | ||
|---|---|---|---|---|---|
| Progressive | Circle | Non-Progressive | |||
| Control | 3.33 ± 0.58 | 3.33 ± 2.08 | 31.67 ± 0.58 | 161.67 ± 1.53 | |
| Ferrous sulfate (Fe) | 20 µg/mL | 0.00 ± 0.00 ## | 0.00 ± 0.00 ## | 0.33 ± 0.58 ## | 199.67 ± 0.58 ## |
| Fe + orange BAE | 6.25 µg/mL | 4.33 ± 3.06 # | 0.00 ± 0.00 * | 30.67 ± 9.61 # | 165.00 ± 12.53 # |
| 12.5 µg/mL | 1.33 ± 0.58 *,# | 0.00 ± 0.00 * | 33.33 ± 7.7 # | 165.33 ± 8.14 # | |
| 25 µg/mL | 4.00 ± 2.00 # | 0.00 ± 0.00 * | 40.67 ± 15.04 # | 155.33 ± 16.86 # | |
| 50 µg/mL | 2.00 ± 0.01 *,# | 0.00 ± 0.00 * | 30.00 ± 3.00 # | 168.00 ± 3.00 *,# | |
| 100 µg/mL | 2.33 ± 1.53 # | 0.00 ± 0.00 * | 30.67 ± 12.22 # | 167.00 ± 13.75 # | |
| Fe + orange BUE | 6.25 µg/mL | 5.67 ± 0.58 *,# | 0.00 ± 0.00 * | 29.67 ± 1.53 # | 164.67 ± 2.08 # |
| 12.5 µg/mL | 3.67 ± 0.58 # | 0.00 ± 0.00 * | 32.00 ± 2.65 # | 164.33 ± 3.06 # | |
| 25 µg/mL | 3.67 ± 0.58 # | 0.00 ± 0.00 * | 41.33 ± 1.53 *,# | 155.00 ± 1.00 *,# | |
| 50 µg/mL | 7.00 ± 3.00 # | 0.00 ± 0.00 * | 68.33 ± 13.50 *,# | 124.67 ± 16.50 *,# | |
| 100 µg/mL | 8.00 ± 1.00 *,# | 0.00 ± 0.00 * | 82.33 ± 22.50 *,# | 109.67 ± 23.50 *,# | |
| Fe + orange BMAE | 6.25 µg/mL | 1.33 ± 0.58 *,# | 0.00 ± 0.00 * | 35.00 ± 4.36 # | 163.67 ± 4.62 # |
| 12.5 µg/mL | 1.00 ± 0.01 *,# | 0.00 ± 0.00 * | 24.00 ± 1.00 *,# | 175.00 ± 1.00 *,# | |
| 25 µg/mL | 0.00 ± 0.00 * | 0.00 ± 0.00 * | 15.33 ± 0.58 *,# | 184.67 ± 0.58 *,# | |
| 50 µg/mL | 1.67 ± 0.58 *,# | 0.00 ± 0.00 * | 29.00 ± 1.00 *,# | 169.33 ± 1.53 *,# | |
| 100 µg/mL | 1.00 ± 0.01 *,# | 0.00 ± 0.00 * | 31.67 ± 3.06 # | 167.33 ± 3.06 *,# | |
| Group | Concentration (µg/mL) | Viable Sperm (%) | Dead Sperm (%) | ||
|---|---|---|---|---|---|
| Intact | Detached | Intact | Detached | ||
| Control | 31.00 ± 6.00 | 28.00 ± 3.00 | 13.67 ± 5.51 | 27.33 ± 8.50 | |
| Ferrous sulfate (Fe) | 20 µg/mL | 4.00 ± 1.00 ## | 23.00 ± 5.00 | 15.00 ± 10.54 | 58.00 ± 6.56 ## |
| Fe + orange BAE | 6.25 µg/mL | 36.00 ± 3.00 # | 40.67 ± 0.58 *,# | 11.00 ± 1.00 | 12.33 ± 3.51 *,# |
| 12.5 µg/mL | 29.33 ± 5.51 # | 39.33 ± 18.50 | 16.33 ± 8.50 | 15.00 ± 4.58 # | |
| 25 µg/mL | 34.67 ± 2.52 # | 36.00 ± 1.00 *,# | 16.33 ± 1.53 | 13.00 ± 2.00 *,# | |
| 50 µg/mL | 19.33 ± 2.52 *,# | 50.00 ± 5.00 *,# | 16.67 ± 1.53 | 14.00 ± 1.00 *,# | |
| 100 µg/mL | 30.67 ± 3.51 # | 29.00 ± 6.00 | 23.33 ± 1.53 * | 17.00 ± 4.00 # | |
| Fe + orange BUE | 6.25 µg/mL | 38.00 ± 18.00 # | 25.00 ± 4.00 | 30.00 ± 18.00 | 7.00 ± 4.00 *,# |
| 12.5 µg/mL | 24.67 ± 8.50 # | 30.00 ± 5.00 | 25.00 ± 2.00 * | 20.33 ± 1.53 # | |
| 25 µg/mL | 19.00 ± 2.65 *,# | 39.33 ± 26.50 | 23.00 ± 15.00 | 18.67 ± 9.07 # | |
| 50 µg/mL | 20.67 ± 17.50 | 40.00 ± 17.00 | 20.33 ± 11.50 | 19.00 ± 12.00 # | |
| 100 µg/mL | 26.67 ± 10.02 # | 25.33 ± 13.50 | 30.00 ± 12.00 | 18.00 ± 8.54 # | |
| Fe + orange BMAE | 6.25 µg/mL | 8.67 ± 3.51 * | 52.00 ± 22.00 # | 22.00 ± 16.52 | 17.33 ± 2.31 # |
| 12.5 µg/mL | 25.67 ± 6.51 *,# | 37.33 ± 7.51 | 18.00 ± 10.00 | 19.00 ± 4.00 # | |
| 25 µg/mL | 14.67 ± 1.53 *,# | 26.00 ± 8.00 | 30.67 ± 10.50 * | 28.67 ± 4.04 # | |
| 50 µg/mL | 19.00 ± 4.00 *,# | 38.33 ± 11.50 | 18.67 ± 2.52 | 24.00 ± 5.00 # | |
| 100 µg/mL | 13.67 ± 11.50 | 52.00 ± 7.00 *,# | 6.00 ± 1.00 * | 28.33 ± 5.51 # | |
| Group | Concentration (µg/mL) | Normal Sperm (%) | Abnormal Sperm (%) | ||
|---|---|---|---|---|---|
| Head Only | Head and Tail | Tail Only | |||
| Control | 45.33 ± 5.51 | 2.67 ± 0.58 | 6.33 ± 1.53 | 45.67 ± 3.51 | |
| Ferrous sulfate (Fe) | 20 µg/mL | 36.00 ± 6.00 | 6.00 ± 4.00 | 17.67 ± 6.51 ## | 40.33 ± 4.51 |
| Fe + orange BAE | 6.25 µg/mL | 64.67 ± 2.52 *,# | 2.00 ± 2.00 | 0.67 ± 0.58 *,# | 32.67 ± 5.03 *,# |
| 12.5 µg/mL | 72.67 ± 9.50 *,# | 0.33 ± 0.58 *,# | 0.67 ± 0.58 *,# | 26.33 ± 8.50 *,# | |
| 25 µg/mL | 77.00 ± 2.00 *,# | 0.00 ± 0.00 *,# | 0.00 ± 0.00 *,# | 23.00 ± 2.00 *,# | |
| 50 µg/mL | 68.33 ± 1.53 *,# | 0.33 ± 0.58 *,# | 1.33 ± 0.58 *,# | 30.00 ± 1.00 *,# | |
| 100 µg/mL | 72.33 ± 0.58 *,# | 0.67 ± 0.58 # | 0.00 ± 0.00 *,# | 27.00 ± 1.00 *,# | |
| Fe + orange BUE | 6.25 µg/mL | 58.33 ± 0.58 *,# | 1.67 ± 0.58 | 2.00 ± 2.00 *,# | 38.00 ± 2.00 * |
| 12.5 µg/mL | 66.33 ± 3.51 *,# | 1.33 ± 0.58 | 3.00 ± 0.01 *,# | 29.33 ± 3.51 *,# | |
| 25 µg/mL | 68.00 ± 3.61 *,# | 1.33 ± 1.15 | 0.00 ± 0.00 *,# | 30.67 ± 2.52 *,# | |
| 50 µg/mL | 69.33 ± 7.51 *,# | 4.00 ± 2.00 | 2.67 ± 1.53 *,# | 24.00 ± 4.00 *,# | |
| 100 µg/mL | 63.67 ± 10.50 *,# | 0.00 ± 0.00 *,# | 0.67 ± 1.15 *,# | 35.67 ± 11.50 | |
| Fe + orange BMAE | 6.25 µg/mL | 55.67 ± 0.58 *,# | 0.00 ± 0.00 *,# | 3.67 ± 1.15 # | 40.67 ± 1.53 |
| 12.5 µg/mL | 49.67 ± 8.50 | 3.00 ± 1.00 | 6.67 ± 1.15 # | 40.67 ± 8.50 | |
| 25 µg/mL | 43.67 ± 6.51 | 1.67 ± 0.58 | 8.67 ± 2.08 | 46.00 ± 4.00 | |
| 50 µg/mL | 62.67 ± 2.52 *,# | 2.67 ± 0.58 | 2.67 ± 2.08 # | 32.00 ± 1.00 *,# | |
| 100 µg/mL | 72.67 ± 0.58 *,# | 1.67 ± 1.53 | 0.67 ± 1.15 *,# | 25.00 ± 0.01 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Laoung-on, J.; Ounjaijean, S.; Sudwan, P.; Boonyapranai, K. Phytochemical Screening, Antioxidant Effect and Sperm Quality of the Bomba ceiba Stamen Extracts on Charolais Cattle Sperm Induced by Ferrous Sulfate. Plants 2024, 13, 960. https://doi.org/10.3390/plants13070960
Laoung-on J, Ounjaijean S, Sudwan P, Boonyapranai K. Phytochemical Screening, Antioxidant Effect and Sperm Quality of the Bomba ceiba Stamen Extracts on Charolais Cattle Sperm Induced by Ferrous Sulfate. Plants. 2024; 13(7):960. https://doi.org/10.3390/plants13070960
Chicago/Turabian StyleLaoung-on, Jiraporn, Sakaewan Ounjaijean, Paiwan Sudwan, and Kongsak Boonyapranai. 2024. "Phytochemical Screening, Antioxidant Effect and Sperm Quality of the Bomba ceiba Stamen Extracts on Charolais Cattle Sperm Induced by Ferrous Sulfate" Plants 13, no. 7: 960. https://doi.org/10.3390/plants13070960
APA StyleLaoung-on, J., Ounjaijean, S., Sudwan, P., & Boonyapranai, K. (2024). Phytochemical Screening, Antioxidant Effect and Sperm Quality of the Bomba ceiba Stamen Extracts on Charolais Cattle Sperm Induced by Ferrous Sulfate. Plants, 13(7), 960. https://doi.org/10.3390/plants13070960