
Genome-Wide Association Analysis Uncovers Genes Associated with Resistance to Head Smut Pathotype 5 in Senegalese Sorghum Accessions


Abstract
:1. Introduction
2. Results
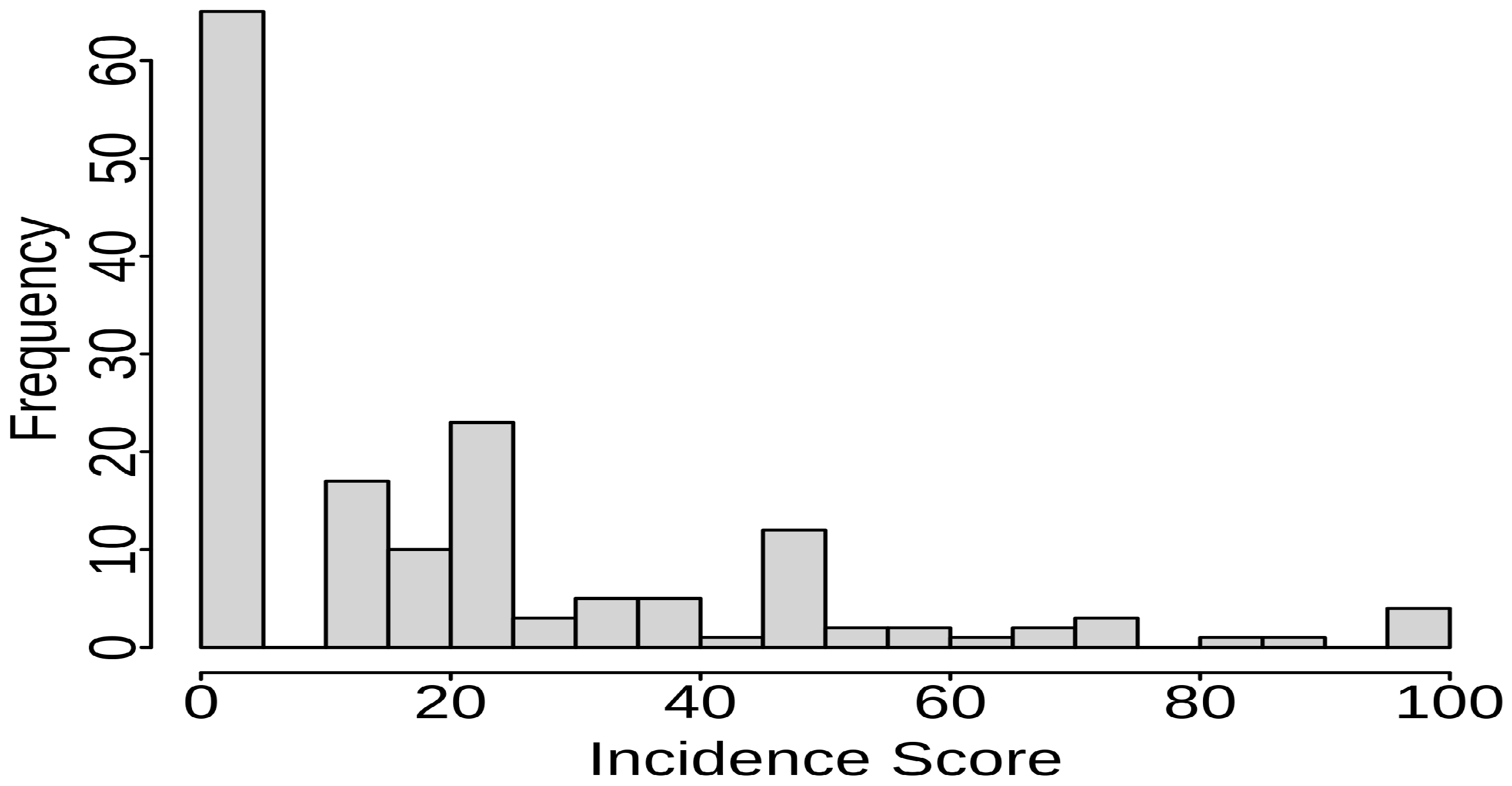
Phenotypic Variation
3. Correlations between Head Smut and Seed Traits in Senegalese Accessions
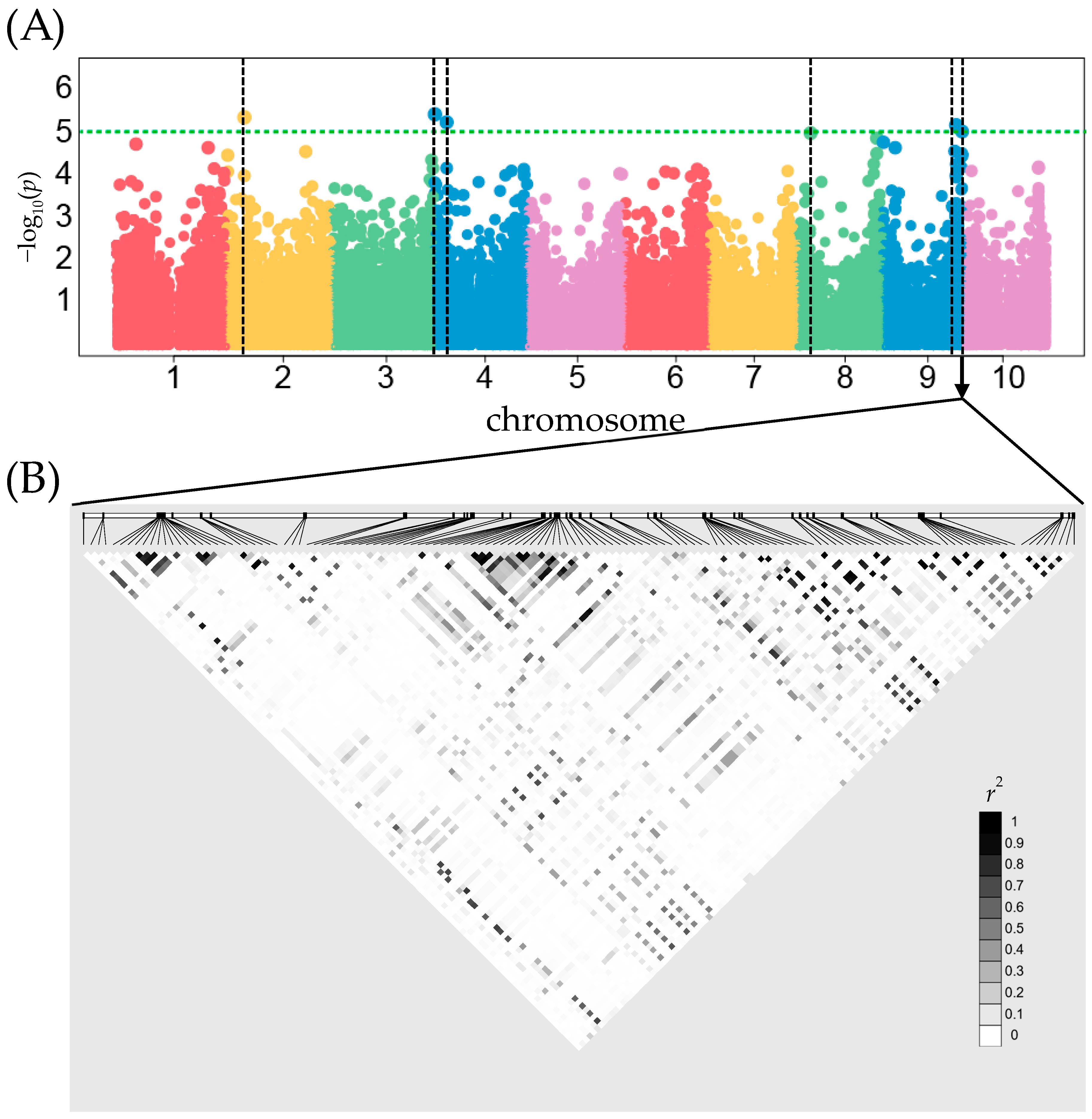
Genome-Wide Association Study
4. Discussion
5. Material and Methods
5.1. Sorghum Lines
5.2. Disease Evaluations
5.3. Statistical Analysis
5.4. Genome-Wide Association Study
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baloch, F.S.; Altaf, M.T.; Liaqat, W.; Bedir, M.; Nadeem, M.A.; Cömertpay, G.; Sun, H.J. Recent advancements in the breeding of sorghum crop: Current status and future strategies for marker-assisted breeding. Front. Genet. 2023, 14, 1150616. [Google Scholar] [CrossRef] [PubMed]
- Hariprasanna, K.; Rakshit, S. Economic Importance of Sorghum. In The Sorghum Genome. Compendium of Plant Genomes; Rakshit, S., Wang, Y.H., Eds.; Springer: Cham, Switzerland, 2016; pp. 1–25. [Google Scholar]
- Widodo, S.; Triastono, J.; Sahara, D.; Pustika, A.B.; Kristamtini; Purwaningsih, H.; Arianti, F.D.; Praptana, R.H.; Romdon, A.S.; Sutardi; et al. Economic Value, Farmers Perception, and Strategic Development of Sorghum in Central Java and Yogyakarta, Indonesia. Agriculture 2023, 13, 516. [Google Scholar] [CrossRef]
- Kumar, A.A. Sorghum: A State of the Art and Future Perspectives; Ignacio, A., Ciampitti, I.A., Vara Prasad, P.V., Eds.; American Society of Agronomy and Crop Science Society of America, Inc.: Madison, WI, USA, 2019; Volume 58, Chapter 5; p. 97. [Google Scholar]
- Prom, L.K.; Perumal, R.; Erattaimuthu, S.R.; Erpelding, J.E.; Montes, N.; Odvody, G.N.; Greenwald, C.; Jin, Z.; Frederiksen, R.; Magill, C. Virulence and molecular genotyping studies of Sporisorium reilianum Isolates in Sorghum. Plant Dis. 2011, 95, 523–529. [Google Scholar] [CrossRef]
- Ahn, E.; Hu, Z.; Perumal, R.; Prom, L.K.; Odvody, G.; Upadhyaya, H.D.; Magill, C.W. Genome wide association analysis of sorghum mini core lines regarding anthracnose, downy mildew, and head smut. PLoS ONE 2019, 14, e0216671. [Google Scholar] [CrossRef]
- Mengistu, G.; Shimelis, H.; Laing, M.; Lule, D. Assessment of Farmers’ Perceptions of Production Constraints, and Their Trait Preferences of Sorghum in Western ETHIOPIA: Implications for Anthracnose Resistance Breeding. Acta Agric. Scand. Sect. B Soil Plant Sci. 2018, 69, 241–249. [Google Scholar] [CrossRef]
- Prom, L.K.; Perumal, R.; Isakeit, T.; Erattaimuthu, S.; Magill, C. Response of sorghum accessions against newly documented pathotypes 5 and 6 of head smut pathogen, Sporisorium reilianum. Am. J. Plant Sci. 2021, 12, 432–443. [Google Scholar] [CrossRef]
- Frederiksen, R.A. Disease and Disease Management in Sorghum. In Sorghum—Origin, History, Technology and Production; Smith, C., Frederiksen, R.A., Eds.; John Wiley & Sons, Inc.: New York, NY, USA, 2000; pp. 497–533. ISBN 978-0-4712-4237-6. [Google Scholar]
- Frederiksen, R.A. Head smut. In Compendium of Sorghum Diseases; Frederiksen, R.A., Odvody, G.N., Eds.; American Phytopathological Society (APS Press): St. Paul, MN, USA, 2000; ISBN 978-0-89054-240-8. [Google Scholar]
- Indira, S.; Xu, X.; Iamsupasit, N.; Shetty, H.S.; Vasanthi, N.S.; Singh, S.D.; Bandyopadhyay, R. Diseases of sorghum and pearl millet in Asia. In Sorghum and Millet Diseases; Leslie, J.F., Ed.; Blackwell Publishing: Hoboken, NJ, USA, 2002; pp. 393–402. [Google Scholar]
- Boyles, R.E.; Cooper, E.A.; Myers, M.T.; Brenton, Z.; Rauh, B.L.; Morris, G.P.; Kresovich, S. Genome-Wide Association Studies of Grain Yield Components in Diverse Sorghum Germplasm. Plant Genome 2016, 9, 1–17. [Google Scholar] [CrossRef]
- Menkir, A.; Ejeta, G.; Butler, L.; Melakeberhan, A. Physical and chemical kernel properties associated with resistance to grain mold in sorghum. Cereal Chem. 1996, 73, 613–617. [Google Scholar]
- Patil, N.Y.; Klein, R.R.; Williams, C.L.; Collins, S.D.; Knoll, J.E.; Burrell, A.M.; Anderson, W.F.; Rooney, W.L.; Klein, P.E. Quantitative trait loci associated with anthracnose resistance in sorghum. Crop Sci. 2017, 57, 877–890. [Google Scholar] [CrossRef]
- Upadhyaya, H.D.; Wang, Y.; Sharma, R.; Sharma, S. Identification of genetic markers linked to anthracnose resistance in sorghum using association analysis. Theor. Appl. Genet. 2013, 126, 1649–1657. [Google Scholar] [CrossRef]
- Jambunathan, R.; Kherdekar, M.S.; Stenhouse, J.W. Sorghum Grain Hardness and Its Relationship to Mold Susceptibility and Mold Resistance. J. Agric. Food Chem. 1992, 40, 1403–1408. [Google Scholar] [CrossRef]
- Craig, J.; Frederiksen, R.A. Comparison of sorghum seedling reactions to Sporisorium reilianum in relation to sorghum head smut resistance classes. Plant Dis. 1992, 76, 314–318. [Google Scholar] [CrossRef]
- Ahn, E.; Prom, L.K.; Fall, C.; Magill, C. Response of Senegalese Sorghum Seedlings to Pathotype 5 of Sporisorium reilianum. Crops 2022, 2, 142–153. [Google Scholar] [CrossRef]
- Ahn, E.; Fall, C.; Prom, L.K.; Magill, C. A Genome-Wide Association Study of Senegalese Sorghum Seedlings Responding to Pathotype 5 of Sporisorium reilianum. Plants 2022, 11, 2999. [Google Scholar] [CrossRef]
- Tam, V.; Patel, N.; Turcotte, M.; Bossé, Y.; Paré, G.; Meyre, D. Benefits and limitations of genome-wide association studies. Nat. Rev. Genet. 2019, 20, 467–484. [Google Scholar] [CrossRef]
- Girma, G.; Nida, H.; Seyoum, A.; Mekonen, M.; Nega, A.; Lule, D.; Dessalegn, K.; Bekele, A.; Gebreyohannes, A.; Adeyanju, A.; et al. A Large-scale Genome-wide Association Analyses of Ethiopian Sorghum Landrace Collection Reveal Loci Associated with Important Traits. Front. Plant Sci. 2019, 10, 691–705. [Google Scholar] [CrossRef]
- Cuevas, H.E.; Prom, L.K.; Rosa-Valentin, G. Population structure of the NPGS Senegalese sorghum collection and its evaluation to identify new disease resistant genes. PLoS ONE 2018, 13, e0191877. [Google Scholar] [CrossRef]
- Ahn, E.; Botkin, J.; Ellur, V.; Lee, Y.; Poudel, K.; Prom, L.K.; Magill, C. Genome-Wide Association Study of Seed Morphology Traits in Senegalese Sorghum Cultivars. Plants 2023, 12, 2344. [Google Scholar] [CrossRef]
- Zhang, F.; Ping, J.; Du, Z.; Cheng, Q.; Huang, Y. Identification of a new race of Sporisorium reilianum and characterization of the reaction of sorghum lines to four races of the head smut pathogen. J. Phytopathol. 2010, 159, 342–346. [Google Scholar] [CrossRef]
- Ahn, E.; Park, S.; Hu, Z.; Ellur, V.; Cha, M.; Lee, Y.; Prom, L.K.; Magill, C.W. Genome-Wide Association Study of Seed Morphology-Related TRAITS in Sorghum Mini Core and Senegalese Lines. Preprints 2024, 2024021209. [Google Scholar] [CrossRef]
- van den Burg, H.A.; Tsitsigiannis, D.I.; Rowland, O.; Lo, J.; Rallapalli, G.; Maclean, D.; Takken, F.L.W.; Jones, J.D.G. The F-box protein ACRE189/ACIF1 regulates cell death and defense responses activated during pathogen recognition in tobacco and tomato. Plant Cell 2008, 20, 697–719. [Google Scholar] [CrossRef] [PubMed]
- Ahn, E.; Prom, L.K.; Hu, Z.; Odvody, G.; Magill, C.W. Genome-wide association analysis for response of Senegalese sorghum accessions to Texas isolates of anthracnose. Plant Genome 2021, 14, e20097. [Google Scholar] [CrossRef] [PubMed]
- Cuevas, H.E.; Prom, L.K.; Cooper, E.A.; Knoll, J.E.; Ni, X. Genome-wide association mapping of anthracnose (Colletotrichum sublineolum) resistance in the US sorghum association panel. Plant Genome 2018, 11, 170099. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.K.; Rai, A.K.; Kanwar, S.S.; Sharma, T.R. Comparative analysis of zinc finger proteins involved in plant disease resistance. PLoS ONE 2012, 7, e42578. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Liu, Y.; Wen, F.; Yao, D.; Wang, L.; Guo, J.; Ni, L.; Zhang, A.; Tan, M.; Jiang, M. A novel rice C2H2-type zinc finger protein, ZFP36, is a key player involved in abscisic acid-induced antioxidant defence and oxidative stress tolerance in rice. J. Exp. Bot. 2014, 65, 5795–5809. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.; Chen, J.; Liu, M.; Zhang, H.; Zhang, S.; Liu, D.; Chen, S. Genome-Wide Analysis of C2H2 Zinc Finger Gene Family and Its Response to Cold and Drought Stress in Sorghum [Sorghum bicolor (L.) Moench]. Int. J. Mol. Sci. 2022, 23, 5571. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhang, L.; Yan, D. Plasmodesmata-Involved Battle Against Pathogens and Potential Strategies for Strengthening Hosts. Front. Plant Sci. 2021, 12, 644870. [Google Scholar] [CrossRef] [PubMed]
- Cheval, C.; Faulkner, C. Plasmodesmal regulation during plant-pathogen interactions. New Phytol. 2018, 217, 62–67. [Google Scholar] [CrossRef]
- Qi, G.; Chen, H.; Wang, D.; Zheng, H.; Tang, X.; Guo, Z.; Cheng, J.; Chen, J.; Wang, Y.; Bai, M.; et al. The BZR1-EDS1 ModuleRegulates Plant Growth-Defense Coordination. Mol. Plant 2021, 14, 2072–2087. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.-H.; Tóth, K.; Kim, D.; Vo, P.H.; Lin, C.-H.; Handakumbura, P.P.; Ubach, A.R.; Evans, S.; Paša-Tolić, L.; Stacey, G. Activation of the Plant Mevalonate Pathway by Extracellular ATP. Nat. Commun. 2022, 13, 450. [Google Scholar] [CrossRef] [PubMed]
- Naets, M.; Van Hemelrijck, W.; Gruyters, W.; Verboven, P.; Nicolaï, B.; Keulemans, W.; De Coninck, B.; Geeraerd, A.H. Time Is of the Essence—Early Activation of the Mevalonate Pathway in Apple Challenged with Gray Mold Correlates with Reduced Susceptibility During Postharvest Storage. Front. Microbiol. 2022, 13, 797234. [Google Scholar] [CrossRef] [PubMed]
- Takeda, S.; Hanano, K.; Kariya, A.; Shimizu, S.; Zhao, L.; Matsui, M.; Tasaka, M.; Aida, M. CUP-SHAPED COTYLEDON1 transcription factor activates the expression of LSH4 and LSH3, two members of the ALOG gene family, in shoot organ boundary cells. Plant J. Cell Mol. Biol. 2011, 66, 1066–1077. [Google Scholar] [CrossRef]
- Schubert, V. SMC proteins and their multiple functions in higher plants. Cytogenet. Genome Res. 2009, 124, 202–214. [Google Scholar] [CrossRef] [PubMed]
- Diaz, M.; Pecinka, A. Scaffolding for Repair: Understanding Molecular Functions of the SMC5/6 Complex. Genes 2018, 9, 36. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Fang, H.; Zhu, C.; Gu, S.; Ding, S.; Yu, J.; Shi, K. Ubiquitylation of PHYTOSULFOKINE RECEPTOR 1 modulates the defense response in tomato. Plant Physiol. 2023, 192, 2507–2522. [Google Scholar] [CrossRef] [PubMed]
- Křeček, P.; Skůpa, P.; Libus, J.; Naramoto, S.; Tejos, R.; Friml, J.; Zažímalová, E. The PIN-FORMED (PIN) protein family of auxin transporters. Genome Biol. 2009, 10, 249. [Google Scholar] [CrossRef] [PubMed]
- Ahn, E.; Fall, C.; Prom, L.K.; Magill, C. Genome-wide association study of Senegalese sorghum seedlings responding to a Texas isolate of Colletotrichum sublineola. Sci. Rep. 2022, 12, 13025. [Google Scholar] [CrossRef] [PubMed]
- Elshire, R.J.; Glaubitz, J.C.; Sun, Q.; Poland, J.A.; Kawamoto, K.; Buckler, E.S.; Mitchell, S.E. A Robust, Simple Genotyping-by-Sequencing (GBS) Approach for High Diversity Species. PLoS ONE 2011, 6, e19379. [Google Scholar] [CrossRef]
- Hu, Z.; Olatoye, M.O.; Marla, S.; Morris, G.P. An integrated genotyping-by-sequencing polymorphism map for over 10,000 sorghum Genotypes. Plant Genome 2019, 12, 180044. [Google Scholar] [CrossRef] [PubMed]
- Upadhyaya, H.D.; Wang, Y.H.; Gowda, C.L.L.; Sharma, S. Association mapping of maturity and plant height using SNP markers with the sorghum mini core collection. Theor. Appl. Genet. 2013, 126, 2003–2015. [Google Scholar] [CrossRef] [PubMed]
- Browning, B.L.; Browning, S.R. Genotype imputation with millions of reference samples. Am. J. Hum. Genet. 2016, 98, 116–126. [Google Scholar] [CrossRef] [PubMed]
- Lipka, A.E.; Tian, F.; Wang, Q.; Peiffer, J.; Li, M.; Bradbury, P.J.; Gore, M.A.; Buckler, E.S.; Zhang, Z. GAPIT: Genome association and prediction integrated tool. Bioinformatics 2012, 28, 2397–2399. [Google Scholar] [CrossRef]
- Liu, X.; Huang, M.; Fan, B.; Buckler, E.S.; Zhang, Z. Iterative usage of fixed and random effect models for powerful and efficient genomewide association studies. PLoS Genet. 2016, 12, e1005767. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.C.; Chow, C.C.; Tellier, L.C.; Vattikuti, S.; Purcell, S.M.; Lee, J.J. Second-generation PLINK: Rising to the challenge of larger and richer datasets. GigaScience 2015, 4, 7. [Google Scholar] [CrossRef] [PubMed]
- Barrett, J.C.; Fry, B.; Maller, J.; Daly, M.J. Haploview: Analysis and visualization of LD and haplotype maps. Bioinformatics 2005, 21, 263–265. [Google Scholar] [CrossRef] [PubMed]
- McCormick, R.F.; Truong, S.K.; Sreedasyam, A.; Jenkins, J.; Shu, S.; Sims, D.; Kennedy, M.; Amirebrahimi, M.; Weers, B.D.; McKinley, B.; et al. The Sorghum bicolor reference genome: Improved assembly, gene annotations, a transcriptome atlas, and signatures of genome organization. Plant J. 2018, 93, 338–354. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Sorghum Accessions Resistant to P5 Head Smut (S. reilianum) | ||
|---|---|---|
| PI 514297 | PI 514376 | PI 514433 |
| PI 514310 | PI 514377 | PI 514434 |
| PI 514317 | PI 514379 | PI 514436 |
| PI 514320 | PI 514381 | PI 514437 |
| PI 514323 | PI 514382 | PI 514438 |
| PI 514338 | PI 514388 | PI 514439 |
| PI 514340 | PI 514392 | PI 514449 |
| PI 514343 | PI 514394 | PI 514452 |
| PI 514345 | PI 514397 | PI 514455 |
| PI 514346 | PI 514398 | PI 514460 |
| PI 514349 | PI 514399 | PI 514461 |
| PI 514351 | PI 514401 | PI 514463 |
| PI 514353 | PI 514403 | PI 514465 |
| PI 514354 | PI 514404 | PI 514466 |
| PI 514355 | PI 514411 | PI 514467 |
| PI 514362 | PI 514412 | PI 514468 |
| PI 514364 | PI 514414 | PI 514471 |
| PI 514366 | PI 514418 | PI 514473 |
| PI 514368 | PI 514419 | PI 514474 |
| PI 514373 | PI 514424 | PI 514475 |
| PI 514375 | PI 514428 | PI 609251 |
| Head Smut Mean Incidence | Seed-Weight | |
|---|---|---|
| Head smut mean incidence | 1.00 *** | 0.49 ** |
| Seed-Area size | 0.16 * | 0.61 *** |
| Seed-Perimeter length | 0.25 ** | 0.67 *** |
| Seed-Length | 0.13 | 0.40 ** |
| Seed-Width | 0.22 ** | 0.74 *** |
| Seed-Length–width ratio (LWR) | −0.23 ** | −0.72 *** |
| Seed-Circularity | −0.27 ** | −0.30 * |
| Seed-Distance between the intersection of length and width (IS) and center of gravity (CG) | 0.32 *** | 0.53 *** |
| Seed-Brightness | −0.04 | −0.41 ** |
| 4-leaf anthracnose average score | −0.06 | 0.63 |
| 4-leaf anthracnose highest score | 0.01 | 0.61 |
| 8-leaf anthracnose average score | 0.02 | 0.48 |
| Seedling head smut spot appearance rate | 0.22 * | 0.16 |
| Seedling head smut spot appearance time | −0.03 | −0.17 |
| Seed-Weight | 0.49 ** | 1.00 *** |
| Cluster | Members | R2 with Its Own Cluster | R2 with the Next Closest Cluster |
|---|---|---|---|
| 1 | Seed-Width | 0.99 | 0.03 |
| Seed-Area size | 0.94 | 0.02 | |
| Seed-Perimeter length | 0.94 | 0.06 | |
| Seed-Length | 0.72 | 0.01 | |
| Seed-LWR | 0.62 | 0.03 | |
| Seed-Weight | 0.58 | 0.26 | |
| 2 | Seed-Circularity | 0.64 | 0.02 |
| Seedling head smut spot appearance rate | 0.50 | 0.06 | |
| Seed-Distance between IS and CG | 0.55 | 0.21 | |
| Head smut mean incidence | 0.35 | 0.07 | |
| 3 | 4-leaf anthracnose highest score | 0.91 | 0.01 |
| 4-leaf anthracnose average score | 0.92 | 0.04 | |
| 8-leaf anthracnose average score | 0.04 | 0.01 | |
| 4 | Seedling head smut spot appearance time | 0.53 | 0.01 |
| Brightness | 0.53 | 0.03 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahn, E.; Prom, L.K.; Park, S.; Hu, Z.; Magill, C.W. Genome-Wide Association Analysis Uncovers Genes Associated with Resistance to Head Smut Pathotype 5 in Senegalese Sorghum Accessions. Plants 2024, 13, 977. https://doi.org/10.3390/plants13070977
Ahn E, Prom LK, Park S, Hu Z, Magill CW. Genome-Wide Association Analysis Uncovers Genes Associated with Resistance to Head Smut Pathotype 5 in Senegalese Sorghum Accessions. Plants. 2024; 13(7):977. https://doi.org/10.3390/plants13070977
Chicago/Turabian StyleAhn, Ezekiel, Louis K. Prom, Sunchung Park, Zhenbin Hu, and Clint W. Magill. 2024. "Genome-Wide Association Analysis Uncovers Genes Associated with Resistance to Head Smut Pathotype 5 in Senegalese Sorghum Accessions" Plants 13, no. 7: 977. https://doi.org/10.3390/plants13070977
APA StyleAhn, E., Prom, L. K., Park, S., Hu, Z., & Magill, C. W. (2024). Genome-Wide Association Analysis Uncovers Genes Associated with Resistance to Head Smut Pathotype 5 in Senegalese Sorghum Accessions. Plants, 13(7), 977. https://doi.org/10.3390/plants13070977