


Qualitative Metabolite Profiling of Orchis purpurea Huds. by GC and UHPLC/MS Approaches
 , ,
, ,  ,
,  ,
,  ,
,  , and
, and 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Essential Oil Characterization
2.2. Characterization of the Non-Volatile Constituents

{kind=link}
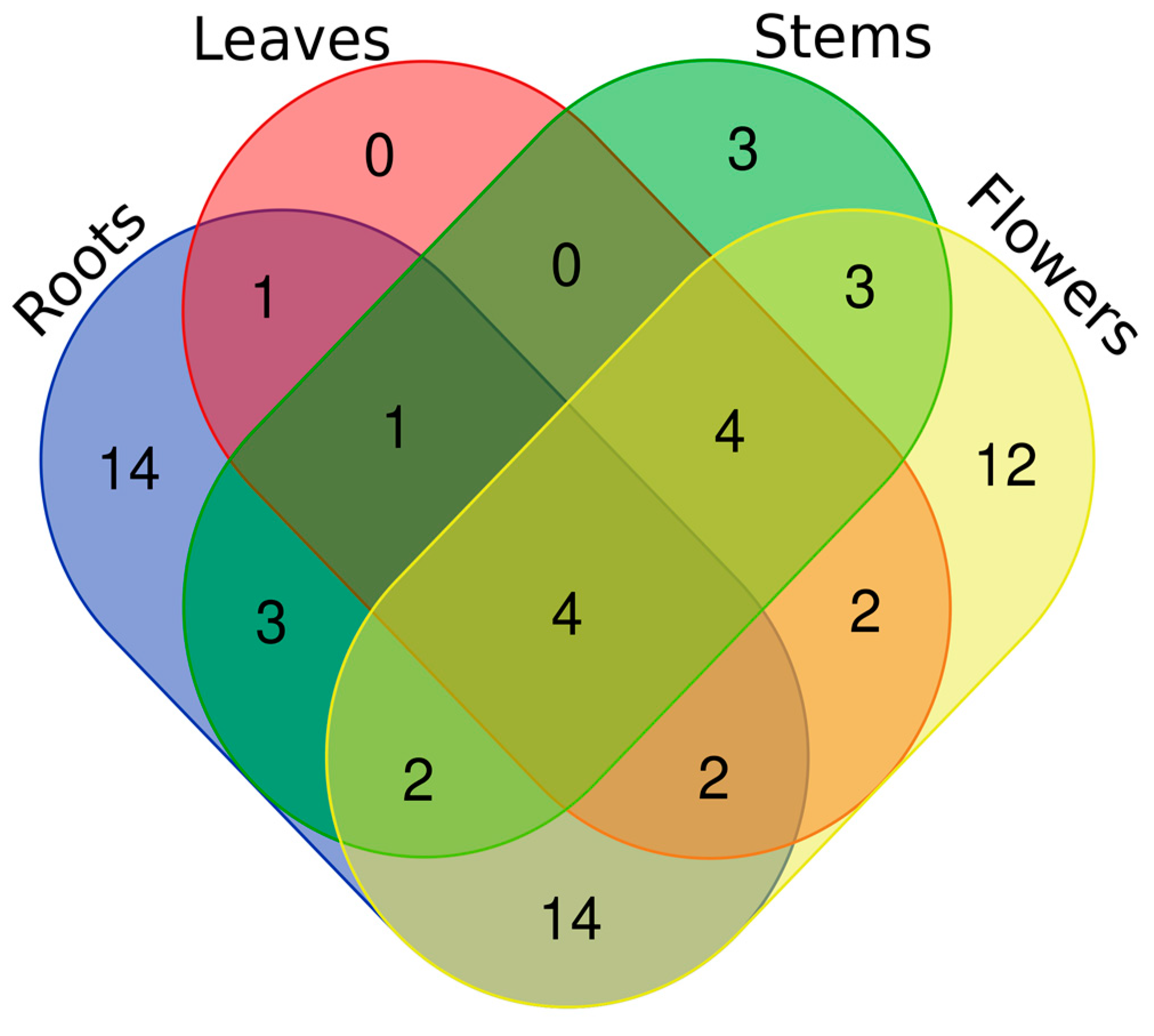{kind=link}
{kind=link}
| N° | Rt (min) | [M − H]− | [M + H]+ | Compounds | Formula | Mass Error (ppm) | Product Ions | Flowers/Leaves | Ref. |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 2.22 | 331.1033 | Koaburaside | C14H20O9 | −0.5 | 123.0445/105.0338 | leaves | [27] | |
| 2 | 3.59 | 205.0855 | Unknown | C12H13O3 | −4.9 | 129.0554/115.0761 | Flowers/leaves | - | |
| 3 | 4.22 | 367.1247 | Coelovirins E | C14H24O11 | 0.3 | 293.1236/143.0710/ 131.0708/99.0811 | Flowers | [28,29] | |
| 4 | 5.42 | 625.1410 | 627.1832 | Quercetin-3-O-gentiobioside | C27H30O17 | −0.1 | 209.0293/191.0190/ 463.0866/301.0346/ 151.0031 | Flowers/leaves | [30] |
| 5 | 5.49 | 325.0932 | p-Coumaroyl-O- hexoside | C15H18O8 | 1.0 | 163.0395/119.0498 | Flowers/ leaves | [31] | |
| 6 | 5.80 | 449.1083 | Cyanidin-3-O- glucoside | C21H20O11 | −0.2 | 287.0549 | Flowers | [32] | |
| 7 | 5.89 | 609.1460 | Luteolin-diglucoside | C27H30O16 | −0.3 | 447.0923/446.0551/ 285.0398/283.0244/ 151.0030 | Flowers/leaves | [30] | |
| 8 | 6.28 | 695.1471 | Kaempferol- malonylhexose- hexose | C30H32O19 | 0.5 | 651.1576/531.1151/ 489.1046/446.0859/ 285.0402 | Flowers/leaves | [33] | |
| 9 | 6.55 | 651.1578 | 2-O-Acetylrutin | C29H32O17 | 1.7 | 489.1041/446.0857/ 285.0401 | Flowers/ leaves | [27] | |
| 10 | 6.56 | 635.2675 | Coelovirins D | C21H47O21 | −1.5 | 349.1143/293.1236/ 277.1286/143.0707 | leaves | [28,29] | |
| 11 | 6.72 | 473.2021 | Coelovirin B | C21H30O12 | −1.5 | 115.0750 | Flowers/ leaves | [28,29] | |
| 12 | 6.91 | 325.0932 | p-Coumaroyl-O- hexoside | C15H18O8 | 1.0 | 163.0395/119.0498 | Flowers/ leaves | [31] | |
| 13 | 7.34 | 787.3220 | Unknown | - | - | 473.1696/285.0981 | Flowers/ leaves | - | |
| 14 | 7.59 | 593.1507 | Luteolin-O-rutinoside | C27H30O19 | −0.4 | 285.0396 | Flowers/ leaves | [27] | |
| 15 | 7.65 | 447.0929 | Kaempferol-hexoside | C21H20O11 | −0.3 | 284.0323/255.0295/ 227.0346 | Flowers | [34] | |
| 16 | 7.68 | 457.2059 | Unknown | C21H30O11 | - | 153.0550/127.0758/ 99.0809 | Flowers/ leaves | - | |
| 17 | 7.88 | 887.3233 | Dactylorhin A | C40H55O22 | 1.1 | 619.2239/439.1606/ 179.0558/153.0553 | Flowers/ leaves | [35] | |
| 18 | 8.15 | 385.1436 | Unknown | - | - | 177.0551/145.0289/ 117.0332 | leaves | - | |
| 19 | 8.37 | 771.2741 | Militarine | C34H46O17 | −1.0 | 457.1220/285.0979/ 153.0555 | Flowers/ leaves | [35] | |
| 20 | 8.57 | 533.1344 | Unknown | C21H29O17 | 2.2 | 390.0738/333.0760 | Flowers/ leaves | - | |
| 21 | 8.78 | 1033.3541 | Unknown saponin | - | - | 765.2605/436.1606/ 619.2237/325.0923 | Flowers | - | |
| 22 | 9.21 | 753.2615 | Unknown | - | - | 439.1609/153.0554 | leaves | - | |
| 23 | 9.49 | 1063.3664 | Unknown saponin | - | - | 749.2662/569.2034/ 439.1611/153.0554 | Flowers | - |
3. Materials and Methods
3.1. Chemicals
3.2. Plant Material
3.3. Extraction of O. purpurea
3.3.1. Steam Distillation
3.3.2. Soxhlet Extraction
3.4. Gas Chromatographic Analysis
3.5. UPLC-MS Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hinsley, A.; de Boer, H.J.; Fay, M.F.; Gale, S.W.; Gardiner, L.M.; Gunasekara, R.S.; Kumar, P.; Masters, S.; Metusala, D.; Roberts, D.L.; et al. A review of the trade in orchids and its implications for conservation. Bot. J. Linn. Soc. 2018, 186, 435–455. [Google Scholar] [CrossRef]
- Fonge, B.A.; Essomo, S.E.; Bechem, T.E.; Tabot, P.T.; Arrey, B.D.; Afanga, Y.; Assoua, E.M. Market trends and ethnobotany of orchids of Mount Cameroon. J. Ethnobiol. Ethnomed. 2019, 15, 29. [Google Scholar] [CrossRef] [PubMed]
- Dhanda, S.; Bullough, L.-A.; Whitehead, D.; Grey, J.; White, K. A Review of the Edible Orchid Trade; Royal Botanic Gardens: London, UK, 2022; Volume 9. [Google Scholar]
- De Boer, H.J.; Ghorbani, A.; Manzanilla, V.; Raclariu, A.-C.; Kreziou, A.; Ounjai, S.; Osathanunkul, M.; Gravendeel, B. DNA metabarcoding of orchid-derived products reveals widespread illegal orchid trade. Proc. R. Soc. B Biol. Sci. 2017, 284, 20171182. [Google Scholar] [CrossRef] [PubMed]
- Tsaballa, A.; Kelesidis, G.; Krigas, N.; Sarropoulou, V.; Bagatzounis, P.; Grigoriadou, K. Taxonomic Identification and Molecular DNA Barcoding of Collected Wild-Growing Orchids Used Traditionally for Salep Production. Plants 2023, 12, 3038. [Google Scholar] [CrossRef] [PubMed]
- Masters, S.; Anthoons, B.; Madesis, P.; Saroja, S.G.; Schermer, M.; Gerritsen, W.; Karahan, A.; Verdoes, R.; Schwallier, R.; van Andel, T.; et al. Quantifying an online wildlife trade using a web crawler. Biodivers. Conserv. 2022, 31, 855–869. [Google Scholar] [CrossRef]
- Kemsawasd, V.; Jayasena, V.; Karnpanit, W. Incidents and Potential Adverse Health Effects of Serious Food Fraud Cases Originated in Asia. Foods 2023, 12, 3522. [Google Scholar] [CrossRef]
- Chase, M.W.; Cameron, K.M.; Freudenstein, J.V.; Pridgeon, A.M.; Salazar, G.; van den Berg, C.; Schuiteman, A. An updated classification of Orchidaceae. Bot. J. Linn. Soc. 2015, 177, 151–174. [Google Scholar] [CrossRef]
- Robustelli della Cuna, F.S.; Cortis, P.; Esposito, F.; De Agostini, A.; Sottani, C.; Sanna, C. Chemical Composition of Essential Oil from Four Sympatric Orchids in NW-Italy. Plants 2022, 11, 826. [Google Scholar] [CrossRef]
- Duan, Z.; Yan, Q.; Wu, F.; Wang, Y.; Wang, S.; Zong, X.; Zhou, P.; Zhang, J. Genome-Wide Analysis of the UDP-Glycosyltransferase Family Reveals Its Roles in Coumarin Biosynthesis and Abiotic Stress in Melilotus albus. Int. J. Mol. Sci. 2021, 22, 10826. [Google Scholar] [CrossRef]
- Martino, E.; Ramaiola, I.; Urbano, M.; Bracco, F.; Collina, S. Microwave-assisted extraction of coumarin and related compounds from Melilotus officinalis (L.) Pallas as an alternative to Soxhlet and ultrasound-assisted extraction. J. Chromatogr. A 2006, 1125, 147–151. [Google Scholar] [CrossRef]
- Priyadarsini, S.S.; Kumar, P.R. Coumarins in orchidaceae: A review connecting ethnomedicine and therapeutic expectations. Rasayan J. Chem. 2022, 14, 2591–2597. [Google Scholar] [CrossRef]
- Ortiz, A.; Sansinenea, E. Phenylpropanoid Derivatives and Their Role in Plants’ Health and as antimicrobials. Curr. Microbiol. 2023, 80, 380. [Google Scholar] [CrossRef] [PubMed]
- Mabry, T.J.; Ulubelen, A. Chemistry and utilization of phenylpropanoids including flavonoids, coumarins, and lignans. J. Agric. Food Chem. 1980, 28, 188–196. [Google Scholar] [CrossRef] [PubMed]
- Stringlis, I.A.; de Jonge, R.; Pieterse, C.M.J. The Age of Coumarins in Plant–Microbe Interactions. Plant Cell Physiol. 2019, 60, 1405–1419. [Google Scholar] [CrossRef] [PubMed]
- Robustelli della Cuna, F.S.; Boselli, C.; Papetti, A.; Mannucci, B.; Calevo, J.; Tava, A. Composition of Volatile Fraction from Inflorescences and Leaves of Dendrobium moschatum (Orchidaceae). Nat. Prod. Commun. 2018, 13, 1934578X1801300. [Google Scholar] [CrossRef]
- Robustelli della Cuna, F.S.; Calevo, J.; Bazzicalupo, M.; Sottani, C.; Grignani, E.; Preda, S. Chemical Composition of Essential Oil from Flowers of Five Fragrant Dendrobium (Orchidaceae). Plants 2021, 10, 1718. [Google Scholar] [CrossRef]
- Matos, M.J.; Santana, L.; Uriarte, E.; Abreu, O.A.; Molina, E.; Yordi, E.G. Coumarins—An important class of phytochemicals. In Phytochemicals—Isolation, Characterisation and Role in Human Health; InTech: Rijeka, Croatia, 2015. [Google Scholar]
- Tena, C.; Santiago, A.d.R.; Osuna, D.; Sosa, T. Phytotoxic Activity of p-Cresol, 2-Phenylethanol and 3-Phenyl-1-Propanol, Phenolic Compounds Present in Cistus ladanifer L. Plants 2021, 10, 1136. [Google Scholar] [CrossRef]
- Johnson, S.D.; Jürgens, A. Convergent evolution of carrion and faecal scent mimicry in fly-pollinated angiosperm flowers and a stinkhorn fungus. South African J. Bot. 2010, 76, 796–807. [Google Scholar] [CrossRef]
- Kyte, G.C.; Hetterscheid, W.L.A.; Lewis, M.J.; Boyce, P.C.; Ollerton, J.; Cocklin, E.; Diaz, A.; Simmonds, M.S.J. Inflorescence odours & pollinators Arum & Amoprphophallus. In Reproductive Biology; Owens, S.J., Rudall, P.J., Eds.; Royal Botanic Gardens: London, UK, 1998; pp. 295–315. [Google Scholar]
- Heberle, H.; Meirelles, G.V.; da Silva, F.R.; Telles, G.P.; Minghim, R. InteractiVenn: A web-based tool for the analysis of sets through Venn diagrams. BMC Bioinform. 2015, 16, 169. [Google Scholar] [CrossRef]
- Ertürk, Ö.; Ayvaz, M.Ç.; Çil, E.; Bağdatlı, E. Gas Chromatography-Mass Spectrometry Analysis and Antimicrobial and Antioxidant Activities of Some Orchid (Orchidaceae) Species Growing in Turkey. Braz. Arch. Biol. Technol. 2023, 66, e23210265. [Google Scholar] [CrossRef]
- Miranda, M.; Cuellar, A. Farmacognosia y Productos Naturales; La Habana Félix Varela: Havana, Cuba, 2001. [Google Scholar]
- Zeitler, K.; Rose, C.A. An Efficient Carbene-Catalyzed Access to 3,4-Dihydrocoumarins. J. Org. Chem. 2009, 74, 1759–1762. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Zhang, Z.; Wu, G.; Sun, Y.; Jiang, Y.; Zhang, H.; Song, X.; Wang, W.; Li, Y. Phenolic derivatives with cytotoxic activities from the roots of Fallopia multiflora var. ciliinervis. Phytochem. Lett. 2022, 52, 72–75. [Google Scholar] [CrossRef]
- ChemSpider. Available online: https://www.chemspider.com (accessed on 12 February 2024).
- Wang, X.; Zhong, X.-J.; Zhou, N.; Cai, N.; Xu, J.-H.; Wang, Q.-B.; Li, J.-J.; Liu, Q.; Lin, P.-C.; Shang, X.-Y. Rapid Characterizaiton of Chemical Constituents of the Tubers of Gymnadenia conopsea by UPLC–Orbitrap–MS/MS Analysis. Molecules 2020, 25, 898. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.-Y.; Li, G.-Q.; Shi, J.-G.; Mo, S.-Y.; Wang, S.-J.; Yang, Y.-C. Chemical constituents of the rhizomes of Coeloglossum viride var. bracteatum. J. Asian Nat. Prod. Res. 2004, 6, 49–61. [Google Scholar] [CrossRef] [PubMed]
- Schütz, K.; Kammerer, D.R.; Carle, R.; Schieber, A. Characterization of phenolic acids and flavonoids in dandelion (Taraxacum officinale WEB. ex WIGG) root and herb by high-performance liquid chromatography/electrospray ionization mass spectrometry. Rapid Commun. Mass Spectrom. 2005, 19, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Cadahía, E.; Fernández de Simón, B.; Aranda, I.; Sanz, M.; Sánchez-Gómez, D.; Pinto, E. Non-targeted Metabolomic Profile of Fagus sylvatica L. Leaves using Liquid Chromatography with Mass Spectrometry and Gas Chromatography with Mass Spectrometry. Phytochem. Anal. 2015, 26, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Gecchele, E.; Negri, S.; Cauzzi, A.; Cuccurullo, A.; Commisso, M.; Patrucco, A.; Anceschi, A.; Zaffani, G.; Avesani, L. Optimization of a Sustainable Protocol for the Extraction of Anthocyanins as Textile Dyes from Plant Materials. Molecules 2021, 26, 6775. [Google Scholar] [CrossRef] [PubMed]
- Loizzo, M.R.; Napolitano, A.; Bruno, M.; Geraci, A.; Schicchi, R.; Leporini, M.; Tundis, R.; Piacente, S. LC-ESI-HRMS analysis of glucosinolates, oxylipins and phenols in Italian rocket salad (Diplotaxis erucoides subsp. erucoides (L.) DC.) and evaluation of its healthy potential. J. Sci. Food Agric. 2021, 101, 5872–5879. [Google Scholar] [CrossRef] [PubMed]
- Neilson, A.P.; Hopf, A.S.; Cooper, B.R.; Pereira, M.A.; Bomser, J.A.; Ferruzzi, M.G. Catechin Degradation with Concurrent Formation of Homo- and Heterocatechin Dimers during In Vitro Digestion. J. Agric. Food Chem. 2007, 55, 8941–8949. [Google Scholar] [CrossRef]
- Lin, B.; Guo, S.; Hong, X.; Jiang, X.; Li, H.; Li, J.; Guo, L.; Li, M.; Chen, J.; Huang, B.; et al. Chemical Characterization and Metabolic Profiling of the Compounds in the Chinese Herbal Formula Li Chang Decoction by UPLC-QTOF/MS. Evid.-Based Complement. Altern. Med. 2022, 2022, 1322751. [Google Scholar] [CrossRef]
- De Agostini, A.; Robustelli della Cuna, F.S.; Cortis, P.; Cogoni, A.; Sottani, C.; Soddu, F.; Sanna, C. Volatile Organic Compounds (VOCs) Diversity in the Orchid Himantoglossum robertianum (Loisel.) P. Delforge from Sardinia (Italy). Diversity 2022, 14, 1125. [Google Scholar] [CrossRef]
- Adams, R. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry, 4th ed.; Allured Publishing Cor-poration: Carol Stream, IL, USA, 2007. [Google Scholar]
- Stein, S.E. NIST/EPA/NIH Mass Spectral Database, Version 2.1; Perkin-Elmer Instrument LLC: Waltham, MA, USA, 2000. [Google Scholar]
- Joulain, D.; Konig, W.A. The Atlas of Spectral Data of Sesquiterpene Hydrocarbons; E. B. Verlag: Hamburg, Germany, 1998. [Google Scholar]
- Ribeiro, C.V.C.; Kaplan, M.A.C. Tendências evolutivas de famílias produtoras de cumarinas em angiospermae. Quim. Nova 2002, 25, 533–538. [Google Scholar] [CrossRef]



| Compound | CAS | Roots | Stems | Leaves | Flowers | Identification d | ||
|---|---|---|---|---|---|---|---|---|
| AI a | AI b | % c | % | % | % | |||
| Octane | 111-65-9 | 800 | 800 | 0.07 ± 0.44 | - | - | 0.06 ± 0.04 | STD, MS, RI |
| Furfural | 98-01-1 | 836 | 831 | - | - | 0.09 ± 0.07 | 0.08 ± 0.05 | MS, RI |
| Diacetone alcohol | 123-42-2 | 841 | 840 | 1.11 ± 0.24 | 0.08 ± 0.07 | - | - | MS, RI |
| Furfuryl alcohol | 98-00-0 | 855 | 855 | - | - | - | 0.21 ± 0.06 | MS, RI |
| 1-Hexanol | 111-27-3 | 871 | 870 | 0.08 ± 0.06 | - | - | 0.08 ± 0.06 | MS, RI |
| Heptanal | 111-71-7 | 902 | 902 | - | - | - | 0.05 ± 0.03 | MS, RI |
| Unidentified | - | - | 907 | - | - | 0.07 ± 0.06 | 0.22 ± 0.06 | - |
| Benzaldehyde | 100-52-7 | 960 | 958 | 0.07 ± 0.08 | - | 0.07 ± 0.06 | 0.10 ± 0.04 | MS, RI |
| Phenol | 108-95-2 | 985 | 985 | 0.07 ± 0.06 | - | - | - | MS, RI |
| 1-Decene | 872-05-9 | 990 | 991 | 0.11 ± 0.08 | - | - | - | MS, RI |
| Octanal | 124-13-0 | 999 | 1003 | 0.08 ± 0.07 | - | - | - | MS, RI |
| 2,4-Heptadienal | 4313-03-5 | 1010 | 1010 | - | - | - | 0.07 ± 0.05 | MS, RI |
| 2- Ethylhexanol | 104-76-7 | 1031 | 1031 | 0.07 ± 0.06 | - | 0.05 ± 0.04 | 0.11 ± 0.07 | MS, RI |
| Benzyl alcohol | 100-51-6 | 1032 | 1034 | 0.08 ± 0.08 | - | 0.05 ± 0.04 | - | MS, RI |
| Phenylacetaldehyde | 122-78-1 | 1042 | 1042 | 0.14 ± 0.11 | - | - | 0.09 ± 0.05 | MS, RI |
| Acetophenone | 98-86-2 | 1065 | 1065 | 0.14 ± 0.11 | 0.13 ± 0.10 | 0.05 ± 0.03 | - | MS, RI |
| p-Cresol | 106-44-5 | 1076 | 1079 | 1.00 ± 0.20 | 11.58 ± 0.32 | 0.44 ± 0.11 | 12.68 ± 0.20 | MS, RI |
| 2-Phenyl-2-propanol | 617-94-7 | 1089 | 1086 | 0.13 ± 0.07 | 0.11 ± 0.09 | - | - | MS, RI |
| Nonanal | 124-19-6 | 1100 | 1105 | 0.21 ± 0.14 | 0.13 ± 0.11 | 0.05 ± 0.04 | 0.64 ± 0.06 | MS, RI |
| (2E)-2-Nonen-1-al | 2463-53-8 | 1162 | 1161 | - | - | - | 0.09 ± 0.04 | MS, RI |
| 1-nonanol | 143-08-8 | 1169 | 1173 | 0.13 ± 0.10 | 0.07 ± 0.06 | - | - | MS, RI |
| Unidentified | - | - | 1185 | - | 0.12 ± 0.12 | 0.07 ± 0.06 | 0.10 ± 0.06 | - |
| 1-Dodecene | 112-41-4 | 1190 | 1192 | 0.18 ± 0.12 | - | - | - | MS, RI |
| p-Methylguaiacol | 93-51-6 | 1192 | 1193 | - | 0.08 ± 0.07 | 0.04 ± 0.03 | 0.37 ± 0.09 | MS, RI |
| Decanal | 112-31-2 | 1200 | 1206 | 0.17 ± 0.12 | 0.11 ± 0.09 | 0.04 ± 0.04 | 0.09 ± 0.05 | MS, RI |
| p-vinylphenol | 2628-17-3 | 1221 | 1220 | 0.17 ± 0.12 | - | - | 1.20 ± 0.06 | MS, RI |
| 2,3-Dihydro-benzofuran | 496-16-2 | 1221 | 1221 | - | 0.06 ± 0.06 | - | - | MS, RI |
| 3-(1-Methylethyl) phenol | 618-45-1 | 1228 | 1229 | 0.09 ± 0.09 | - | - | - | MS, RI |
| 3,5-Dimethoxy-toluene | 4179-19-5 | 1264 | 1267 | - | - | - | 0.19 ± 0.06 | MS, RI |
| Nonanoic acid | 112-05-0 | 1271 | 1276 | 0.08 ± 0.06 | - | - | 0.67 ± 0.05 | MS, RI |
| Unidentified | - | - | 1308 | - | - | - | 0.41 ± 0.08 | - |
| 2-Methoxy-4-vinylphenol | 7786-61-0 | 1315 | 1315 | 0.16 ± 0.13 | 0.08 ± 0.06 | - | 0.08 ± 0.06 | MS, RI |
| (2E,4E)-2,4-Decadienal | 25152-84-5 | 1317 | 1317 | 0.21 ± 0.20 | - | - | 0.08 ± 0.07 | MS, RI |
| p-Hydroxybenzyl alcohol | 623-05-2 | 1357 | 1356 | 0.11 ± 0.13 | - | - | 0.11 ± 0.06 | MS, RI |
| Decanoic acid | 334-48-5 | 1372 | 1372 | - | - | - | 0.05 ± 0.04 | MS, RI |
| Unidentified | - | - | 1379 | 0.17 ± 0.16 | - | - | 0.11 ± 0.06 | - |
| 3,4-dihydro-coumarin | 119-84-6 | 1378 | 1384 | - | 0.12 ± 0.08 | 0.07 ± 0.03 | 0.08 ± 0.07 | MS, RI |
| (E)-damascenone | 23726-93-4 | 1385 | 1386 | - | - | - | 0.12 ± 0.06 | MS, RI |
| 2-Tetradecene | 26952-13-6 | 1389 | 1393 | 0.22 ± 0.17 | - | - | - | MS, RI |
| Tetradecane | 629-59-4 | 1400 | 1400 | - | - | - | 0.05 ± 0.04 | STD, MS, RI |
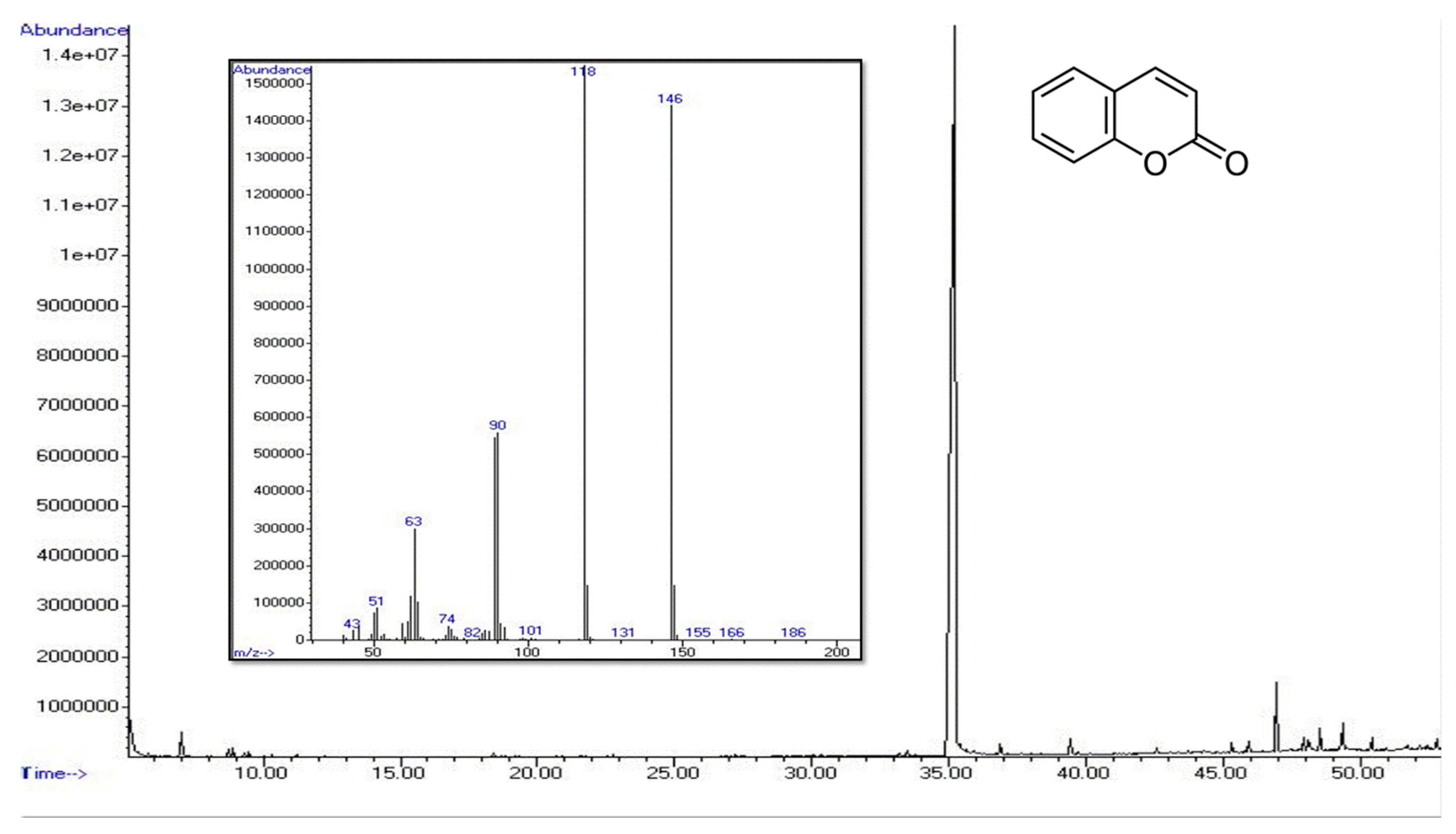
| Coumarin | 91-64-5 | 1445 | 1455 | 32.30 ± 0.32 | 85.98 ± 0.27 | 98.79 ± 0.34 | 69.64 ± 0.22 | MS, RI |
| 2,6-Di-tert-butyl-p-benzoquinone | 719-22-2 | 1469 | 1469 | - | - | - | 0.11 ± 0.06 | MS, RI |
| Unidentified | - | - | 1560 | - | - | - | 0.31 ± 0.12 | - |
| Dodecanoic acid | 143-07-7 | 1565 | 1566 | 0.26 ± 0.17 | - | - | 0.23 ± 0.12 | MS, RI |
| 1-Hexadecene | 629-73-2 | 1590 | 1592 | 0.21 ± 0.15 | - | - | 0.20 ± 0.09 | MS, RI |
| Hexadecane | 544-76-3 | 1600 | 1600 | 0.10 ± 0.08 | - | - | - | STD, MS, RI |
| Methyl dihydro jasmonate | 24851-98-7 | 1656 | 1657 | 0.16 ± 0.15 | - | - | - | MS, RI |
| Tetradecanoic acid | 544-63-8 | 1780 | 1765 | 0.16 ± 0.14 | - | - | 0.24 ± 0.10 | MS, RI |
| 1-Octadecene | 112-88-9 | 1790 | 1794 | 0.19 ± 0.20 | - | - | - | MS, RI |
| Octadecane | 593-45-3 | 1800 | 1800 | 0.15 ± 0.11 | - | - | - | MS, RI |
| 1-Methylethyl tetradecanoate | 110-27-0 | 1828 | 1828 | 0.14 ± 0.10 | - | - | - | MS, RI |
| Cyclohexadecane | 295-65-8 | 1881 | 1881 | - | - | - | 0.46 ± 0.07 | MS, RI |
| 1-Hexadecanol | 36653-82-4 | 1876 | 1887 | 14.03 ± 0.21 | - | - | - | MS, RI |
| Nonadecane | 629-92-5 | 1900 | 1900 | 0.12 ± 0.11 | - | - | 0.10 ± 0.05 | STD, MS, RI |
| 7,9-Di-tert-butyl-1-oxaspiro-(4,5)-deca-6,9-diene | 82304-66-3 | 1929 | 1923 | 0.06 ± 0.06 | - | - | - | MS, RI |
| Hexadecanoic acid | 57-10-3 | 1972 | 1972 | 3.51 ± 0.12 | - | - | 2.54 ± 0.12 | MS, RI |
| Ethyl hexadecanoate | 628-97-7 | 1995 | 1995 | 0.09 ± 0.09 | - | - | 0.15 ± 0.07 | MS, RI |
| Isopropyl palmitate | 142-91-6 | 2026 | 2026 | 0.09 ± 0.06 | - | - | - | MS, RI |
| (E)-15-heptadecenal | 700381-35-7 | 2085 | 2085 | 43.39 ± 0.40 | 0.20 ± 0.11 | - | 0.86 ± 0.08 | MS, RI |
| Heneicosane | 629-94-7 | 2100 | 2100 | - | 0.12 ± 0.06 | - | 0.88 ± 0.08 | STD, MS, RI |
| Unidentified | - | - | - | - | - | - | 0.16 ± 0.04 | - |
| Docosane | 629-97-0 | 2200 | 2200 | - | 0.10 ± 0.06 | - | - | STD, MS, RI |
| Unidentified | - | - | 2271 | - | 0.12 ± 0.11 | - | - | - |
| Tricosane | 638-67-5 | 2300 | 2300 | - | 0.10 ± 0.07 | - | 0.73 ± 0.11 | STD, MS, RI |
| Tetracosane | 646-31-1 | 2400 | 2400 | - | 0.21 ± 0.11 | - | - | STD, MS, RI |
| 9-pentacosene | 51865-00-0 | 2474 | 2475 | - | - | - | 0.07 ± 0.05 | MS, RI |
| 1-Docosanol | 661-19-8 | 2493 | 2493 | - | - | 0.03 ± 0.02 | 0.27 ± 0.06 | MS, RI |
| Pentacosane | 629-99-2 | 2500 | 2500 | - | 0.18 ± 0.15 | 0.04 ± 0.03 | 2.53 ± 0.05 | STD, MS, RI |
| Hexacosane | 630-01-3 | 2600 | 2600 | - | 0.19 ± 0.15 | - | 0.22 ± 0.07 | STD, MS, RI |
| Heptacosane | 593-49-7 | 2700 | 2700 | - | 0.14 ± 0.11 | 0.04 ± 0.03 | 2.10 ± 0.07 | STD, MS, RI |
| Class | Roots | Stems | Leaves | Flowers |
|---|---|---|---|---|
| Acids | 4.00 | - | - | 3.74 |
| Alcohols | 16.12 | 11.91 | 0.61 | 15.13 |
| Aldeydes | 44.28 | 0.43 | 0.25 | 2.15 |
| Esters (among which coumarin) | 32.77 (32.30) | 86.10 (86.10) | 98.86 (98.86) | 69.87 (69.72) |
| Ketones | 1.11 | 0.08 | 0.05 | 0.52 |
| Saturated hydrocarbons | 0.44 | 1.04 | 0.08 | 7.13 |
| Unsaturated hydrocarbons | 0.90 | - | - | 0.26 |
| Unidentified | 0.31 | 0.37 | 0.14 | 0.91 |
| Miscellanea | 0.06 | - | - | 0.19 |
| Oxygenated monoterpenes | - | - | - | 0.12 |
| Extractive Solvent | |||
|---|---|---|---|
| n-Hexane | Ethyl Acetate | Methanol | |
| Leaves | 0.33% | 0.16% | 3.83% |
| Flowers | 0.95% | 2.05% | 12.77% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cavalloro, V.; Pagliari, S.; Gosetti, F.; Campone, L.; Sottani, C.; Collina, S.; Martino, E.; Robustelli della Cuna, F.S. Qualitative Metabolite Profiling of Orchis purpurea Huds. by GC and UHPLC/MS Approaches. Plants 2024, 13, 1064. https://doi.org/10.3390/plants13081064
Cavalloro V, Pagliari S, Gosetti F, Campone L, Sottani C, Collina S, Martino E, Robustelli della Cuna FS. Qualitative Metabolite Profiling of Orchis purpurea Huds. by GC and UHPLC/MS Approaches. Plants. 2024; 13(8):1064. https://doi.org/10.3390/plants13081064
Chicago/Turabian StyleCavalloro, Valeria, Stefania Pagliari, Fabio Gosetti, Luca Campone, Cristina Sottani, Simona Collina, Emanuela Martino, and Francesco Saverio Robustelli della Cuna. 2024. "Qualitative Metabolite Profiling of Orchis purpurea Huds. by GC and UHPLC/MS Approaches" Plants 13, no. 8: 1064. https://doi.org/10.3390/plants13081064
APA StyleCavalloro, V., Pagliari, S., Gosetti, F., Campone, L., Sottani, C., Collina, S., Martino, E., & Robustelli della Cuna, F. S. (2024). Qualitative Metabolite Profiling of Orchis purpurea Huds. by GC and UHPLC/MS Approaches. Plants, 13(8), 1064. https://doi.org/10.3390/plants13081064