Systematic Description of Epidendroideae Pollinium/Tetrads/Pollen
The findings for the investigated Epidendroideae species are presented according to their subdivision into lower and higher epidendroids and arranged alphabetically by genera. Each species is described in terms of the morphology and size of its pollinarium/pollinium, tetrad types, and the size of its single pollen grains, initially based on LM studies. Subsequently, the ornamentation and the ultrastructure of the pollen wall, and its cell content, are delineated based on SEM and TEM. The pollen terminology used follows Halbritter et al. [
10].
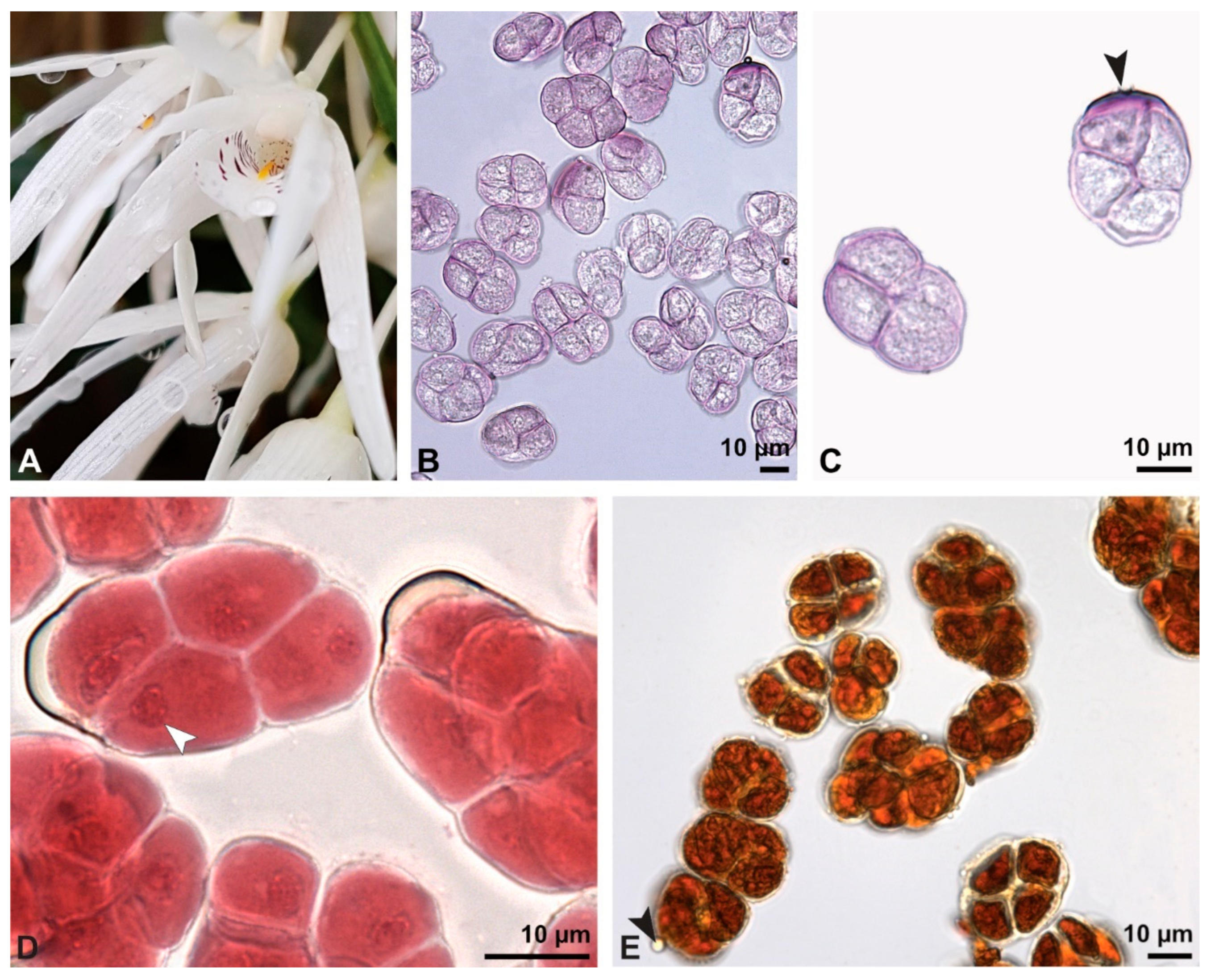
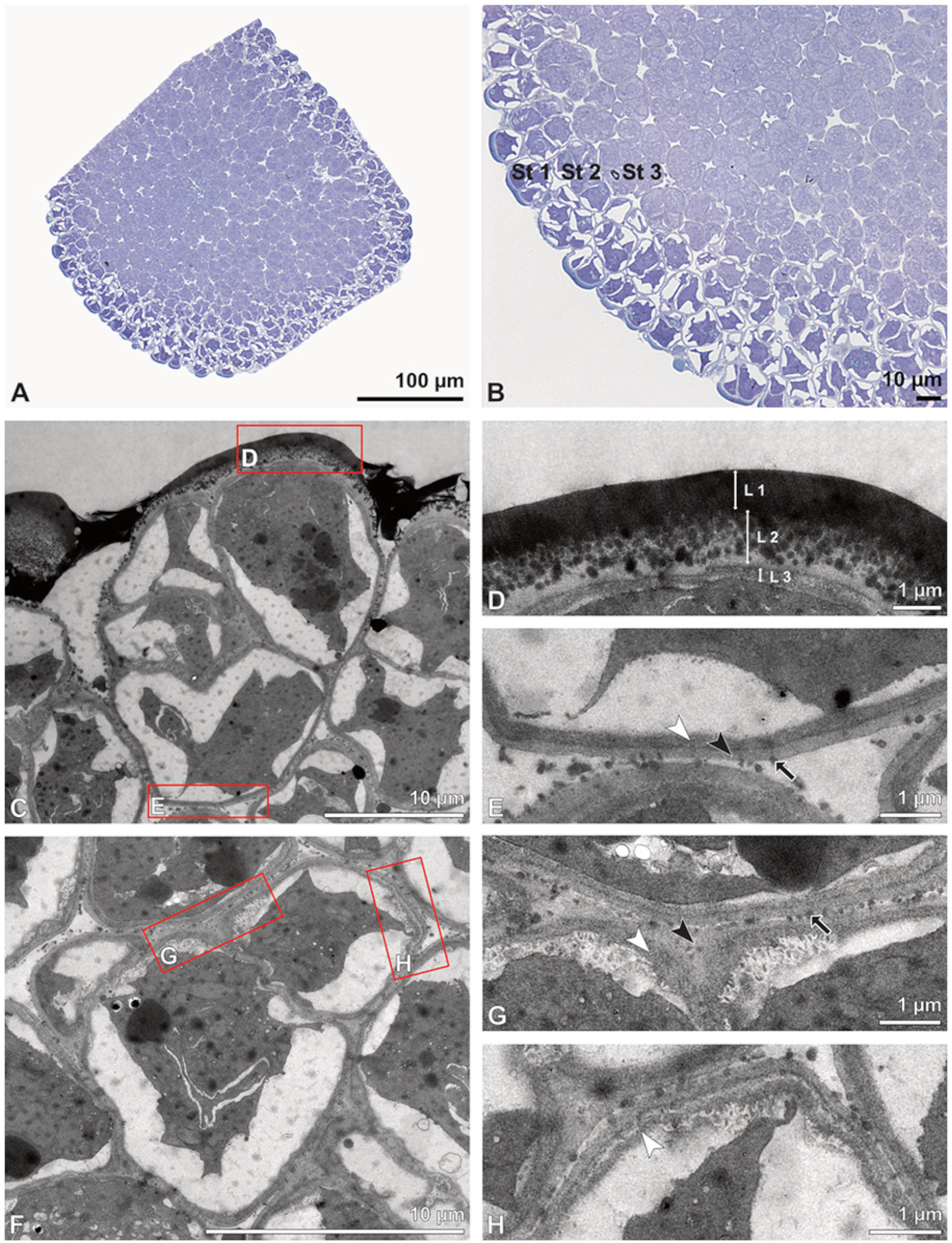
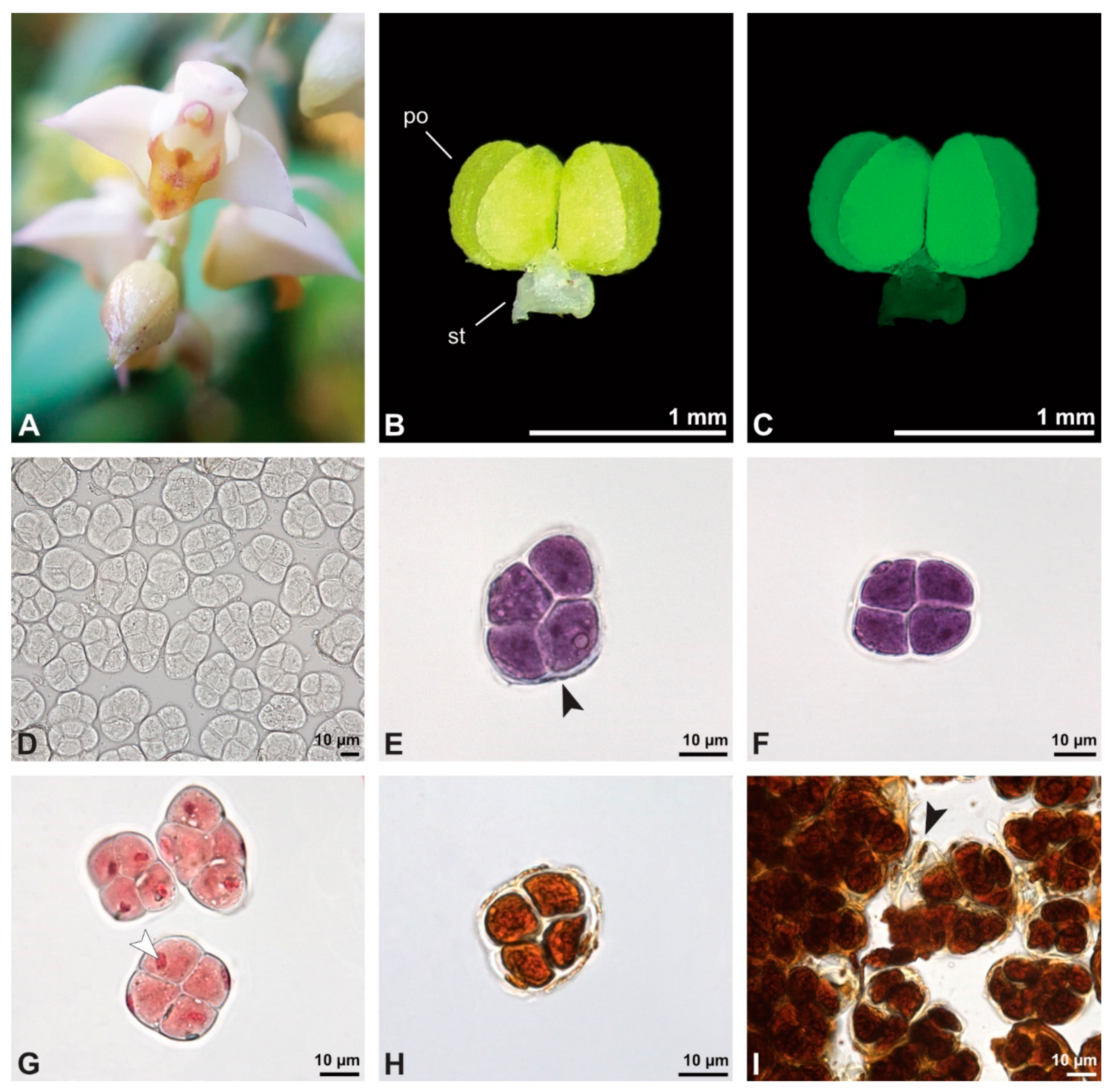
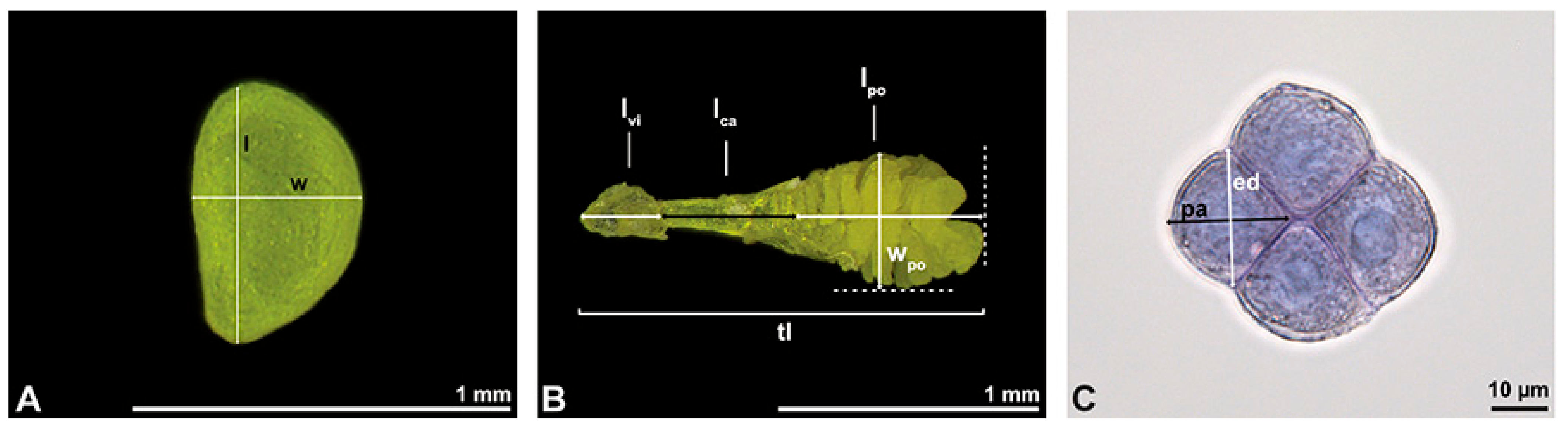
The pollen dispersal unit of
Bulbophyllum retusiusculum comprises a compact bean-shaped pollinium (
Figure 2B and
Figure 3A) measuring 0.69 mm in length and 0.46 mm in width (LM), showing autofluorescence (
Figure 2C,
Table 1). The pollinium is composed of tetrads (
Figure 2D–I and
Figure 3B) glued together by pollen coatings (
Figure 3C,D). Various types of tetrads have been identified, including tetrahedral, planar-tetragonal, planar-rhomboid, and decussate (
Figure 2D–G). In LM, the hydrated pollen grains within tetrads range in size from 10.8 µm to 14.3 µm (
Table 1). Mature pollen grains are inaperturate and two-celled at anthesis (
Figure 2H and
Figure 7A). Toluidine blue staining of the tetrads revealed differences in the pollen wall structure (
Figure 2E).
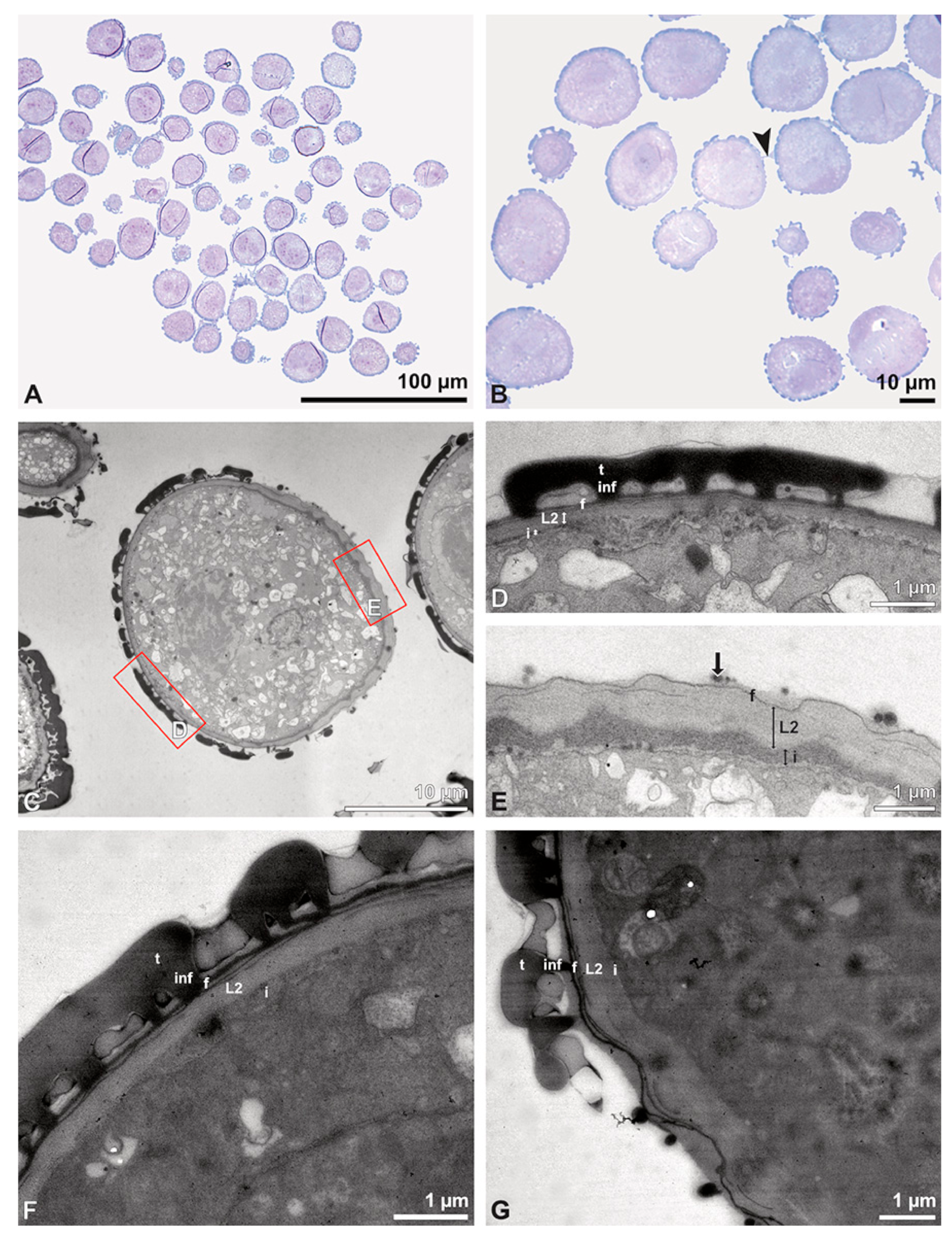
Figure 3.
SEM micrographs of Bulbophyllum retusiusculum pollinium/pollen. Note: (A) Overview of hydrated (DMP + CPD) pollinia. (B) Tetrad arrangement. (C) Tetrads with psilate, perforate to fossulate ornamentation, and pollen coatings in between (white arrowhead). (D) Exine of outer tetrads; perforate, fossulate, granulate (black arrow), with fine striate ornamentation and pollen coatings (white arrowhead) connecting the tetrads.
Figure 3.
SEM micrographs of Bulbophyllum retusiusculum pollinium/pollen. Note: (A) Overview of hydrated (DMP + CPD) pollinia. (B) Tetrad arrangement. (C) Tetrads with psilate, perforate to fossulate ornamentation, and pollen coatings in between (white arrowhead). (D) Exine of outer tetrads; perforate, fossulate, granulate (black arrow), with fine striate ornamentation and pollen coatings (white arrowhead) connecting the tetrads.
Under LM and SEM, the outermost tetrads constitute a thick pollen wall (
Figure 2F) with psilate to fine striate to fossulate ornamentation and perforations (
Figure 3C,D). Contrary to the outer thick-walled tetrads of the pollinium, the inner tetrads have only thin pollen walls (
Figure 2G).
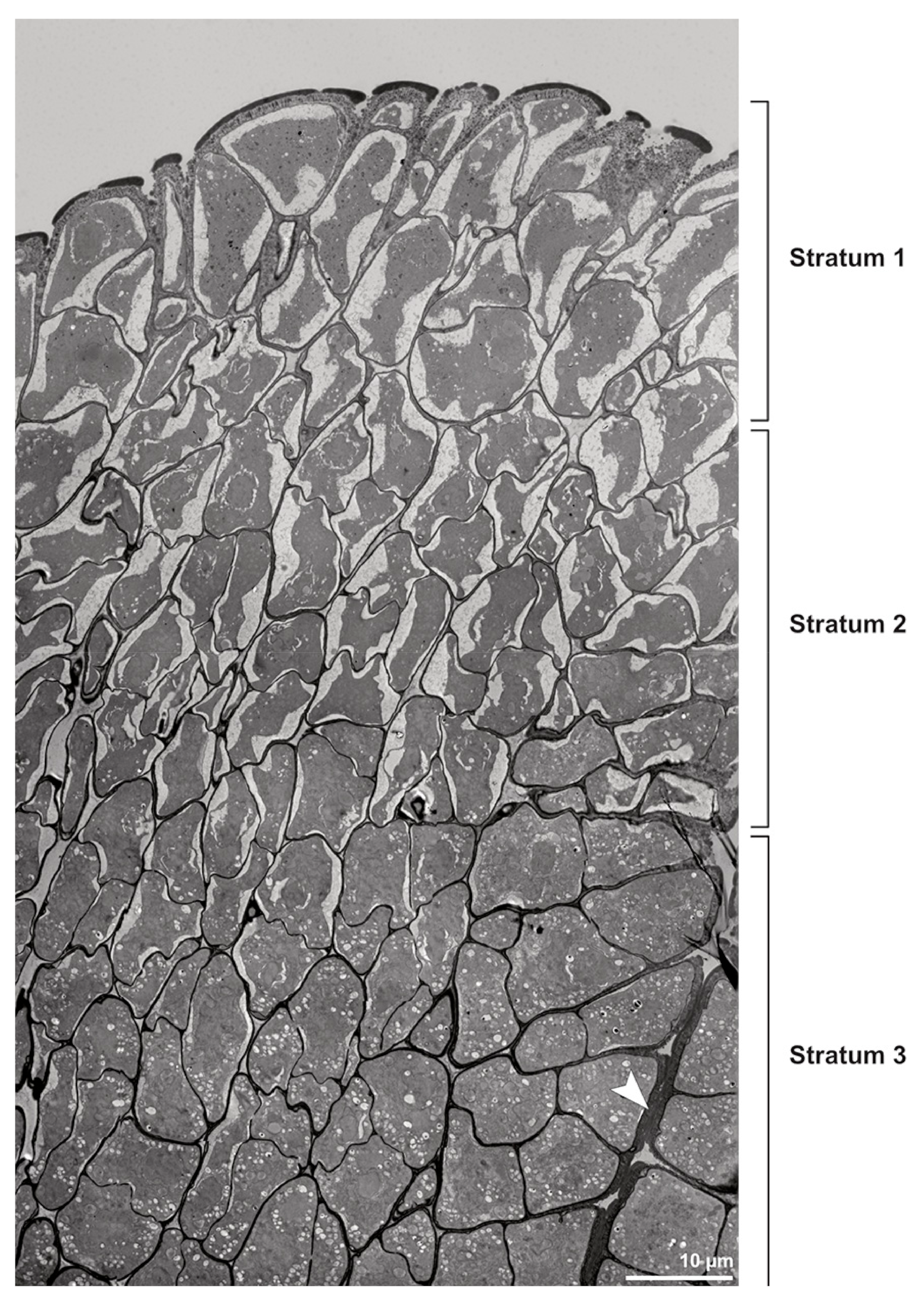
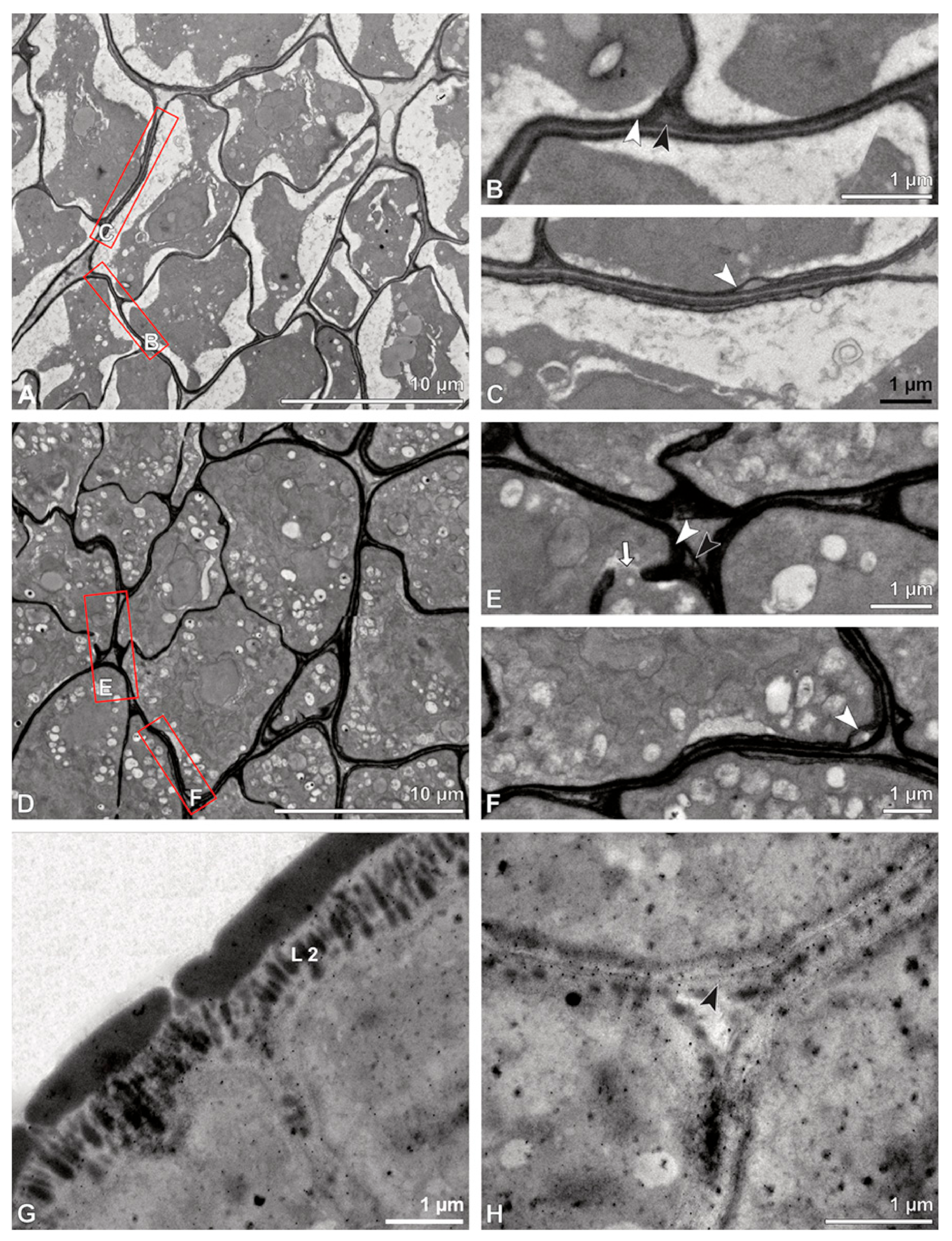
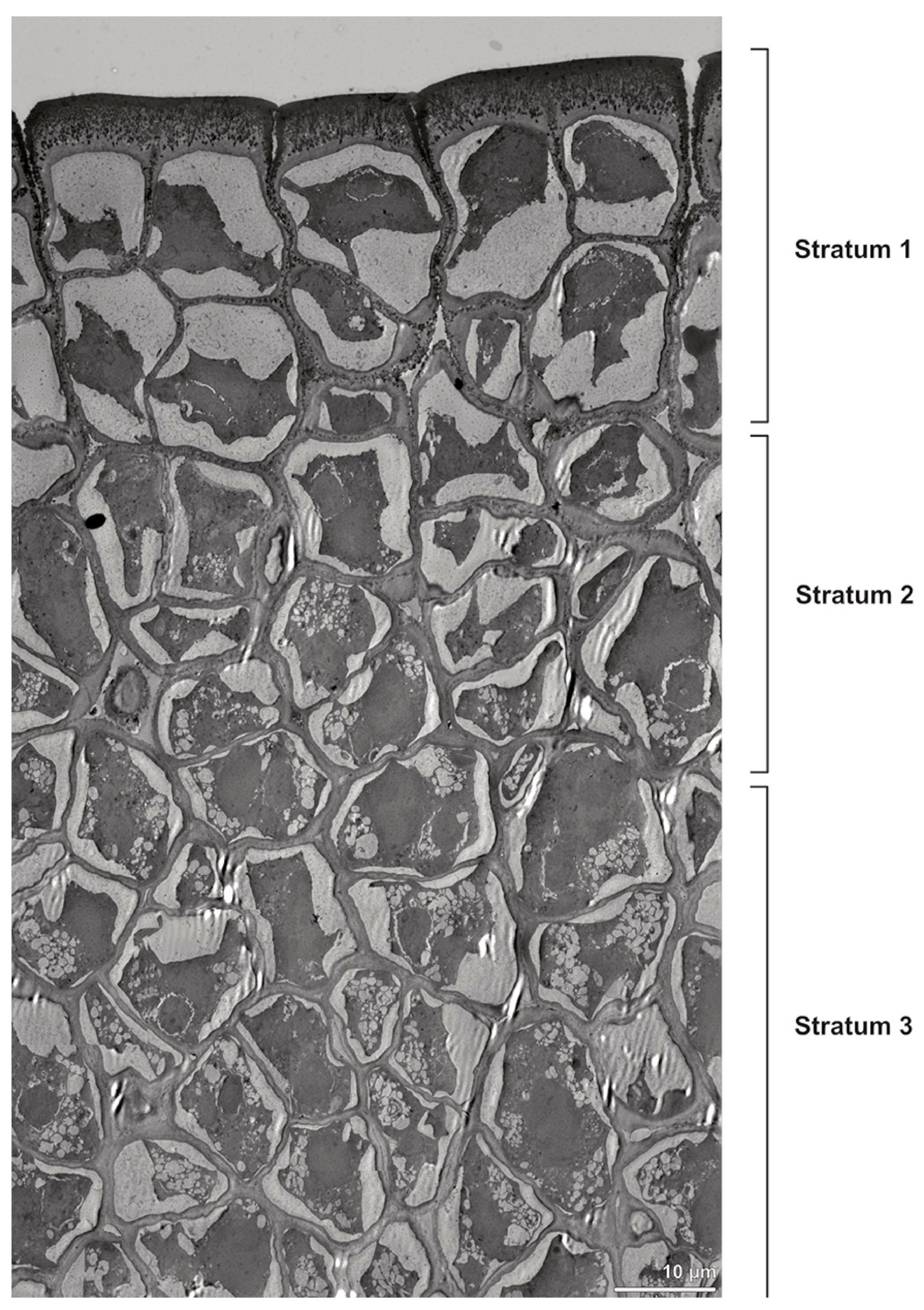
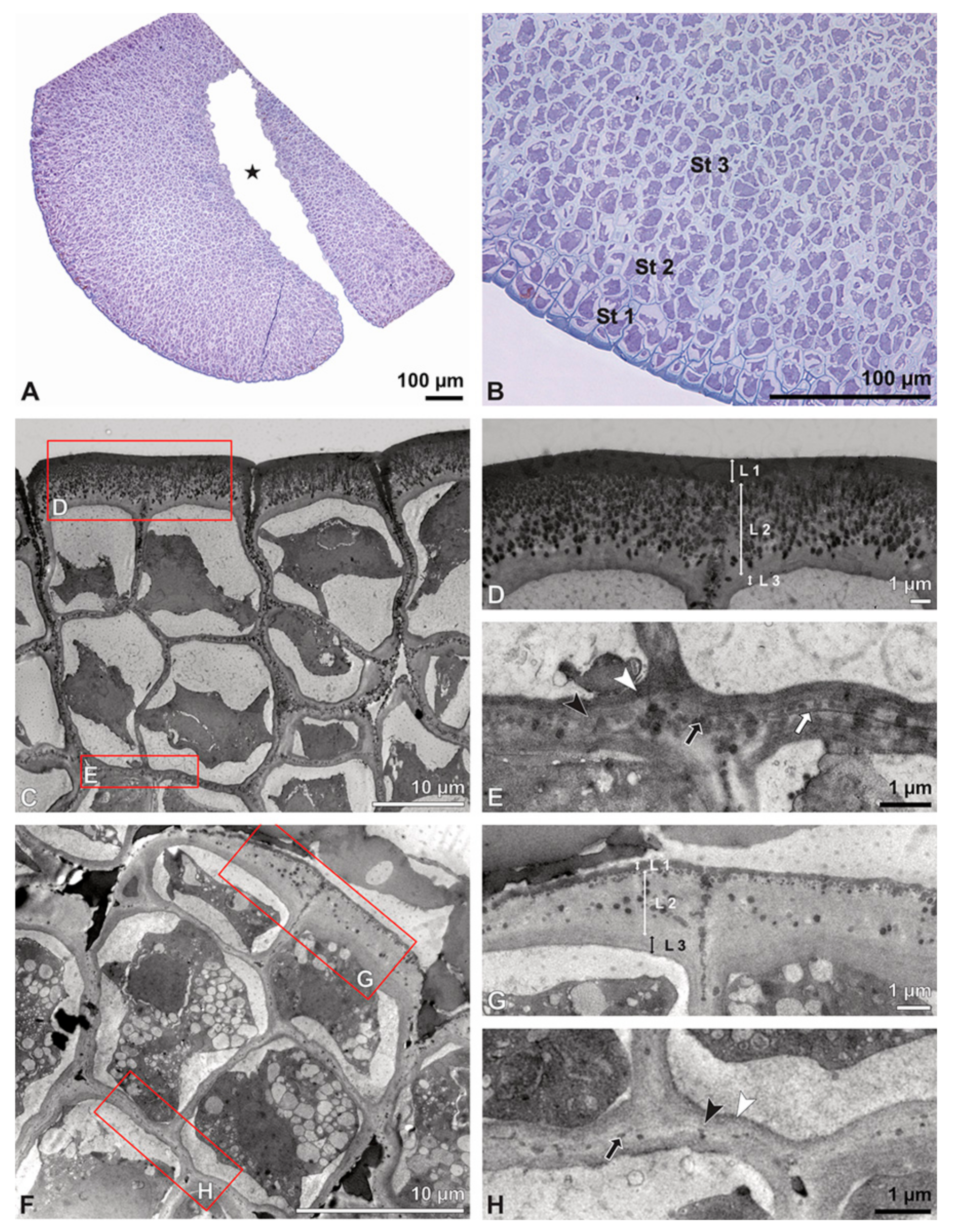
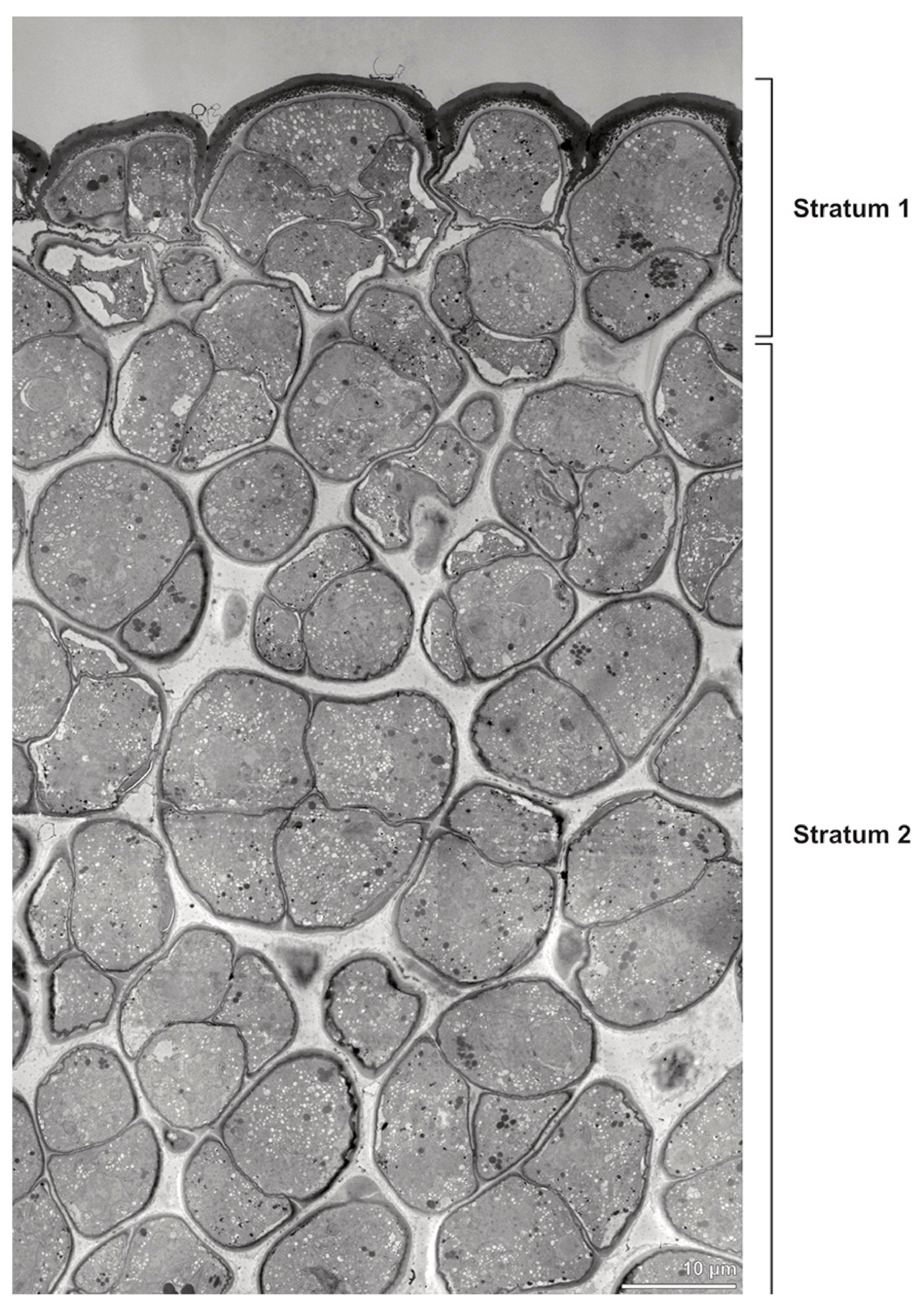
Ultrastructural studies reveal a trilaminate structure to the pollinium, consisting of outer tetrads with thick pollen walls and detached protoplasts (stratum 1), tetrads with detached protoplasts and reduced pollen walls (stratum 2), and inner tetrads with reduced pollen walls (stratum 3) (
Figure 4 and
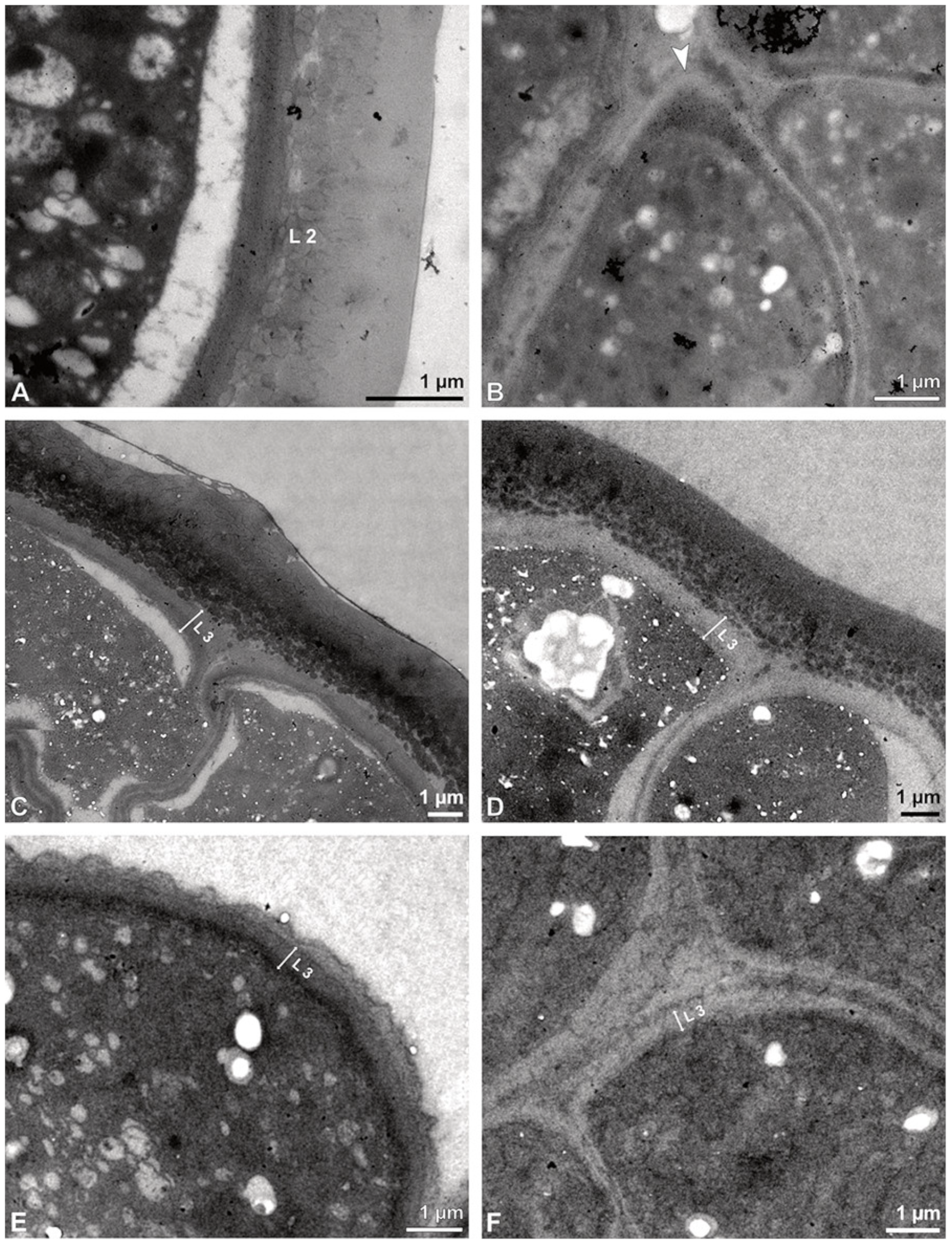
Figure 5A,B). The peripheral part of the pollinium (stratum 1) shows tetrads with pollen walls comprising three layers (
Figure 5C,D). The outermost layer (layer 1), a continuous, compact sporopollenin layer (
Figure 5C,D), constitutes a common pollen wall layer on the distal half of the outer pollen grains only (
Figure 5C). In the contact zones of neighbouring outer tetrads of the pollinium, the compact sporopollenin layer transitions into smaller discontinuous subunits (
Figure 5C). A subjacent electron-translucent pollen wall layer (layer 2) surrounds each pollen grain (monad), forming a common layer around each tetrad (
Figure 5C,F). Layer 2 is much thicker than layer 1 and comprises electron-dense columellate to granulate elements (
Figure 5D). It decreases inwardly in thickness and density (
Figure 5C). The chemical composition of layer 2 appears to be polysaccharidic, as staining with potassium permanganate does not indicate that it has a lipidic nature (
Figure 6G,H). The thin innermost wall (layer 3) is monolayered and most likely an intine. At the distal half of the monads, this layer is undulating due to the detachment of the generative cell (
Figure 5E,H). The pollen walls of the inner pollen grains of the outer tetrads are reduced and consist only of layer 2 and layer 3 (intine) (
Figure 5E). The protoplasts of all outer tetrads of stratum 1 are detached from the intine, have no intact nuclei and are poor in organelles (
Figure 4 and
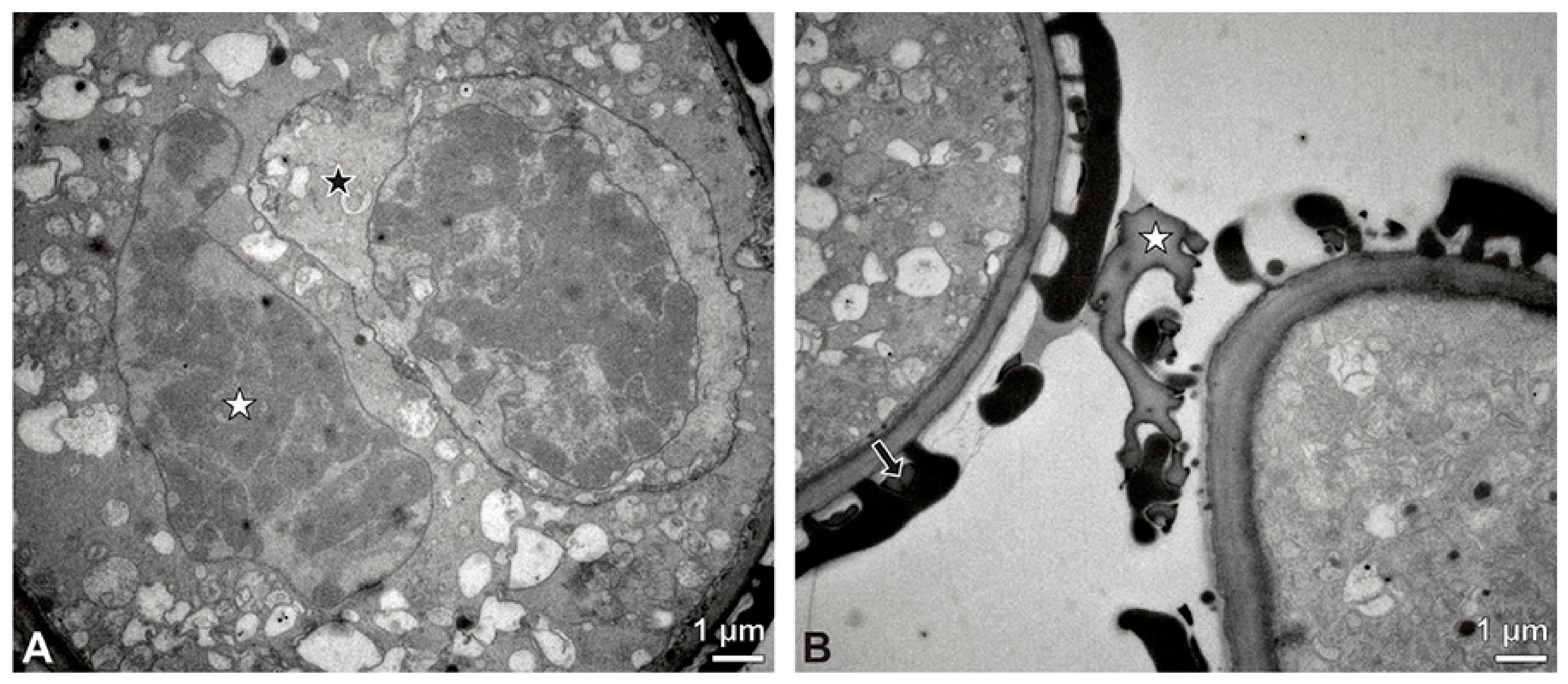
Figure 5C), suggesting infertility. In the contact area of two pollinia (
Figure 4) the tetrads did not show detached protoplasts, indicating the fertility of the mature two-celled pollen grains (
Figure 5F). At the contact area of the outer pollen grains of the tetrads, the pollen wall layers 2 and 3 are slightly thinner (
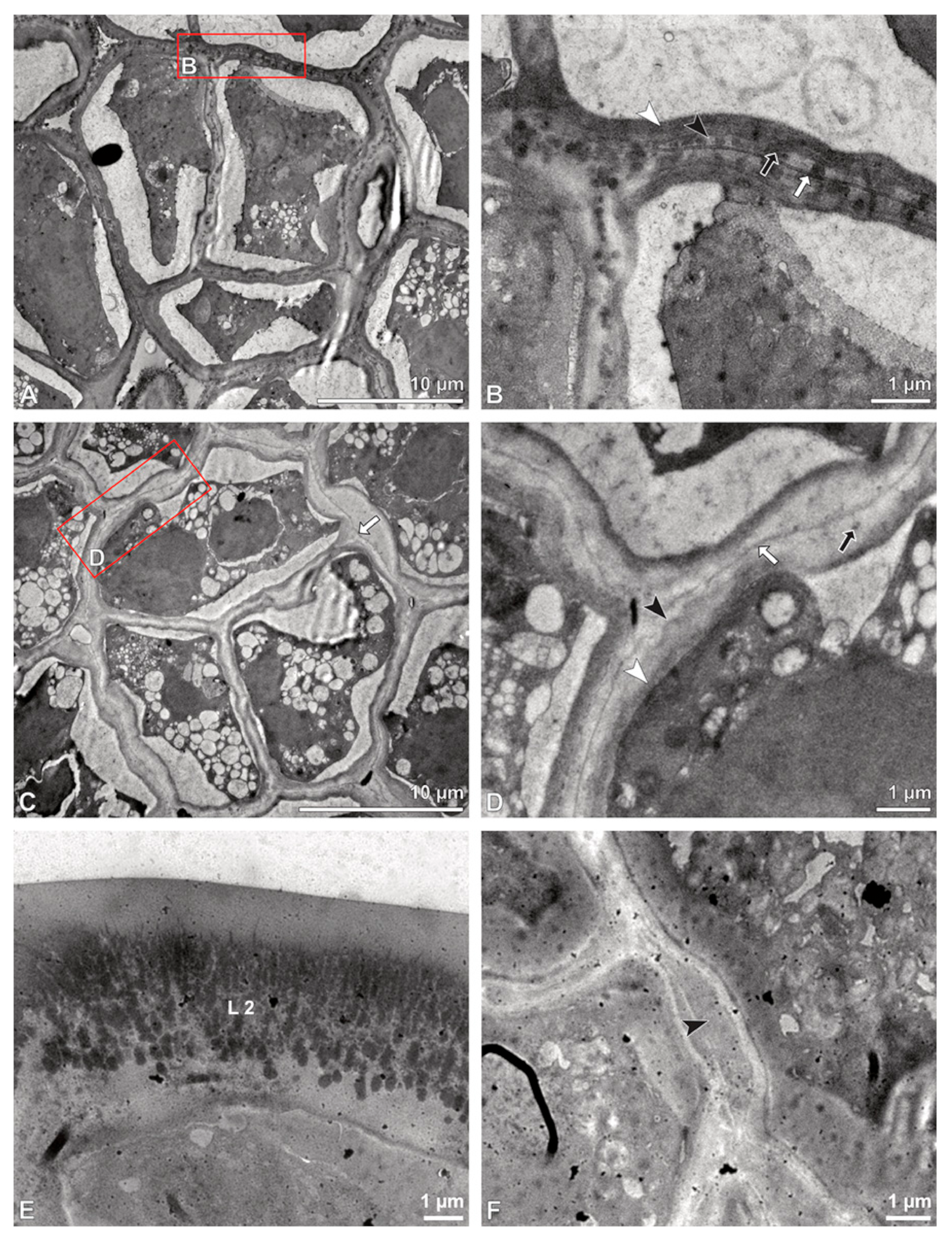
Figure 5G). In stratum 2, the pollinium exhibits tetrads with highly reduced pollen walls (
Figure 6A), consisting of layer 2 and layer 3 only (
Figure 6B). Similar to the tetrads of stratum 1, layer 2 surrounds each tetrad and forms the outer pollen wall layer of each monad (
Figure 6B). The inner layer (layer 3), a monolayered intine, is undulating in the distal half of the monads (
Figure 6C). As observed in the outer tetrads of stratum 1, the protoplasts are detached and poor in organelles (
Figure 4 and
Figure 6A). Tetrads of the inner part of the pollinium (stratum 3) resemble those of stratum 2, with reduced pollen walls consisting of layer 2 and intine (layer 3) (
Figure 6D–F). Contrary to the tetrads of stratum 1 and stratum 2, the tetrads of stratum 3 contain protoplasts richer in organelles and without a detached protoplast (
Figure 4 and
Figure 6D). Pollen reserves like starch are absent in the tetrads of all three strata (
Figure 2I); only some lipid droplets are present (
Figure 5F). Some tetrads of stratum 2 and 3 exhibit plasma bridges between the monads (
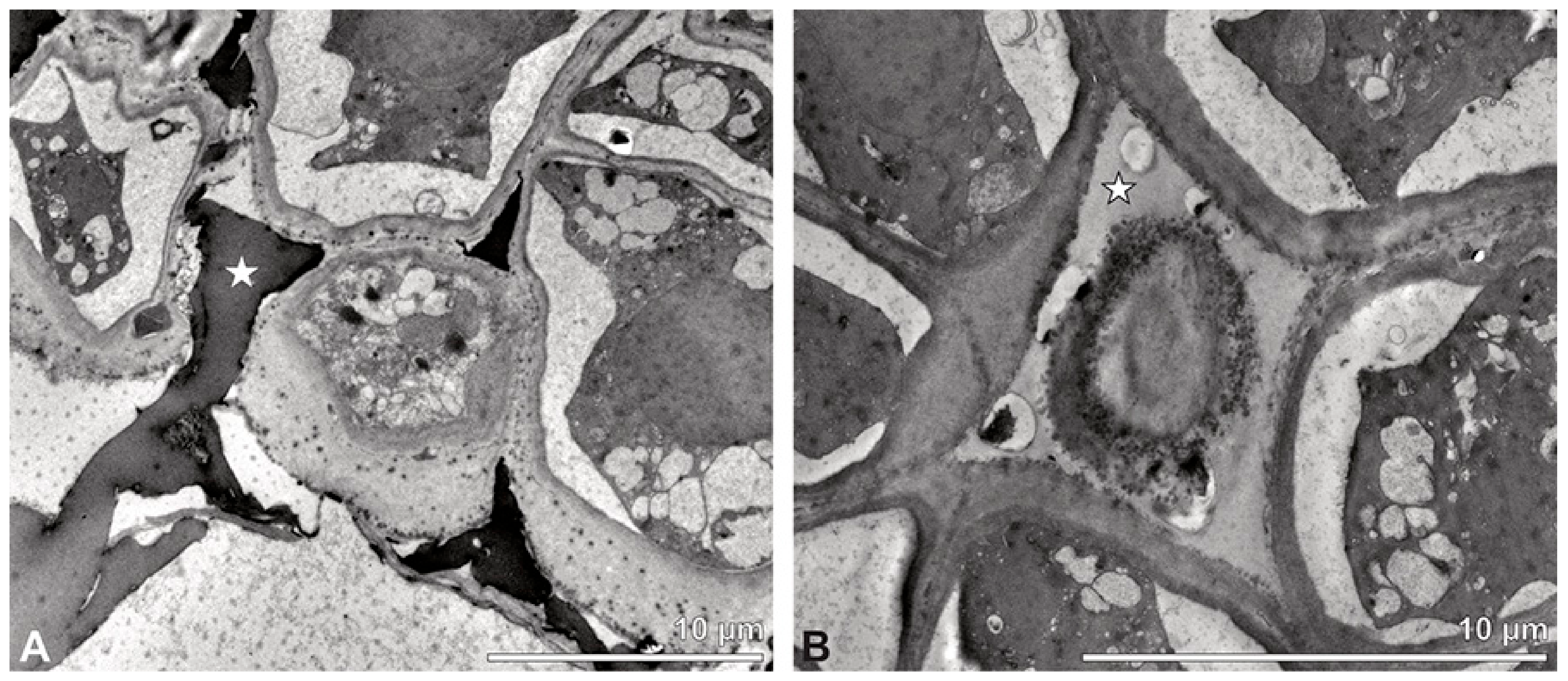
Figure 6E), suggesting that these tetrads have not reached their final mature stage, where plasma bridges are usually entirely closed. The spaces between the tetrads of all strata are filled with an electron-translucent filamentous material (
Figure 7B) gluing the tetrads together.
Remarks: To date, there are only few pollen studies on this genus, although it comprises c. 2168 species [
38]. Schill and Pfeiffer [
24] conducted SEM studies on nine species of
Bulbophyllum, describing a predominantly psilate ornamentation, which is in accordance with our results for
B. retusiusculum. Zavada [
39] has provided only TEM work to date, which has highlighted the structural differences in the pollen wall of
B. imbricatum Lindl. between the peripheral and inner tetrads. Our study on
B. retusiusculum corroborates these findings, and additionally shows differences in the protoplasts’ conditions within tetrads, depending on their position within the pollinium. The peripheral tetrads of stratum 1 exhibit sporopollenin wall elements and detached protoplasts, while tetrads from the centre (stratum 3) display reduced pollen walls without sporopollenin elements and intact protoplasts. Notably, the pollen wall structure deviates from the classical pollen wall model (see [
10]), particularly in the presence of the unique layer 2, which is not comparable to any known wall layer. Additionally, layer 3 deviates from a classical intine, appearing more like a primary cell wall.
Description of species
Cephalanthera longifolia (L.) Fritsch (
Figure 8,
Figure 9,
Figure 10,
Figure 11 and
Figure 12;
Table 1,
Table 2 and
Table 3), lower epidendroids: The pollen dispersal unit constitutes a soft banana-shaped pollinium (
Figure 9A) with an average length of 3.2 mm and an average width of 0.3 mm that shows autofluorescence in LM (
Figure 8B,C,
Table 1). It consists of tetrads (
Figure 8D and
Figure 9B) agglutinated by pollen coatings (
Figure 8H). These tetrads easily disintegrate into heteropolar monads, displaying, in LM, a size ranging in their hydrated condition between 26.4 µm and 33.1 µm (
Figure 8I,
Table 1). Monads are ulcerate, spheroidal, isodiametric to oblate, and heteropolar (
Figure 8E,F) and have a suboblate P/E ratio in SEM (
Figure 9D). Within a tetrad, the ulcus is positioned at the proximal pole and is not visible in the intact pollinium (
Figure 9B).
In SEM, the pollen wall ornamentation in the interapertural area is foveolate to reticulate-heterobrochate, with thick muri (
Figure 9C). The size of the lumina ranges from 1.14 µm to 3.31 µm (mean 2.16 µm). The aperture membrane is ornamented (nano- to micro-verrucate, granulate) (
Figure 9D).
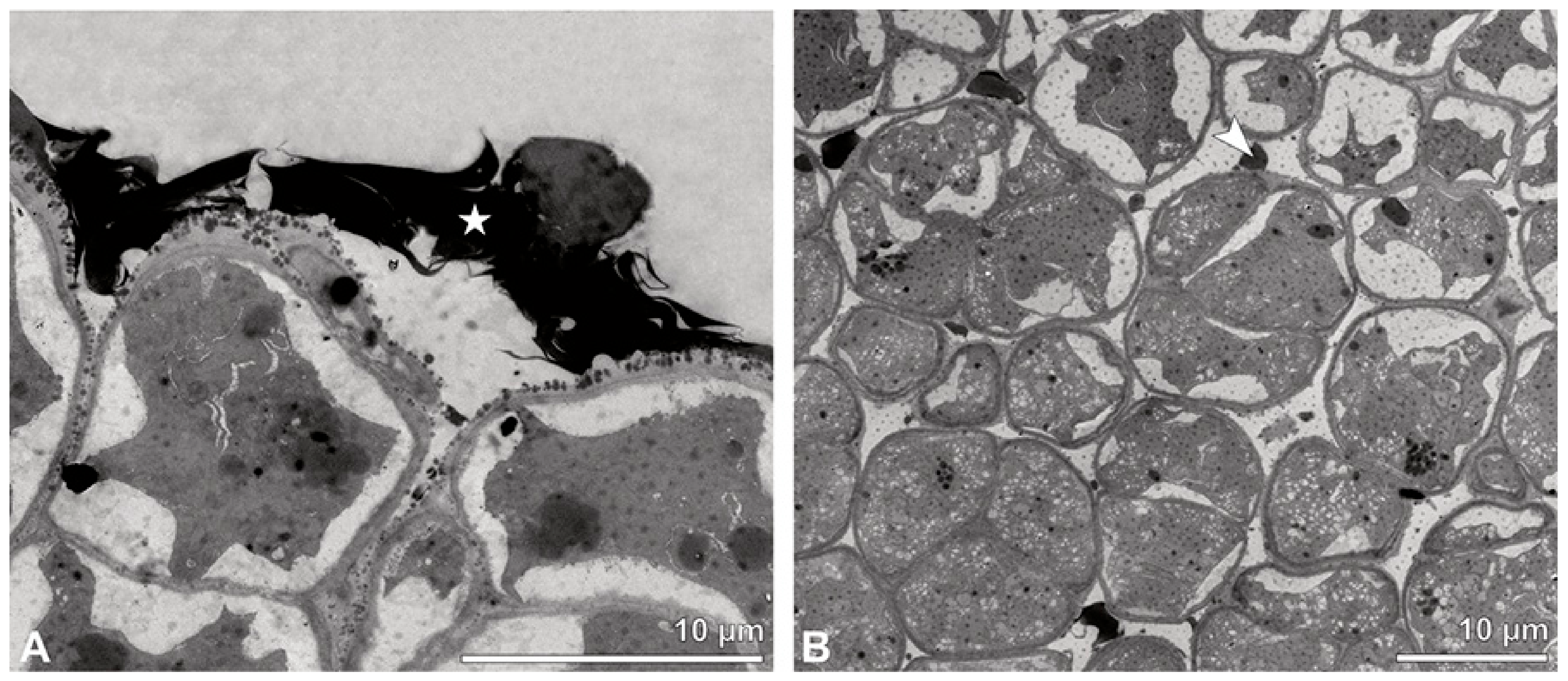
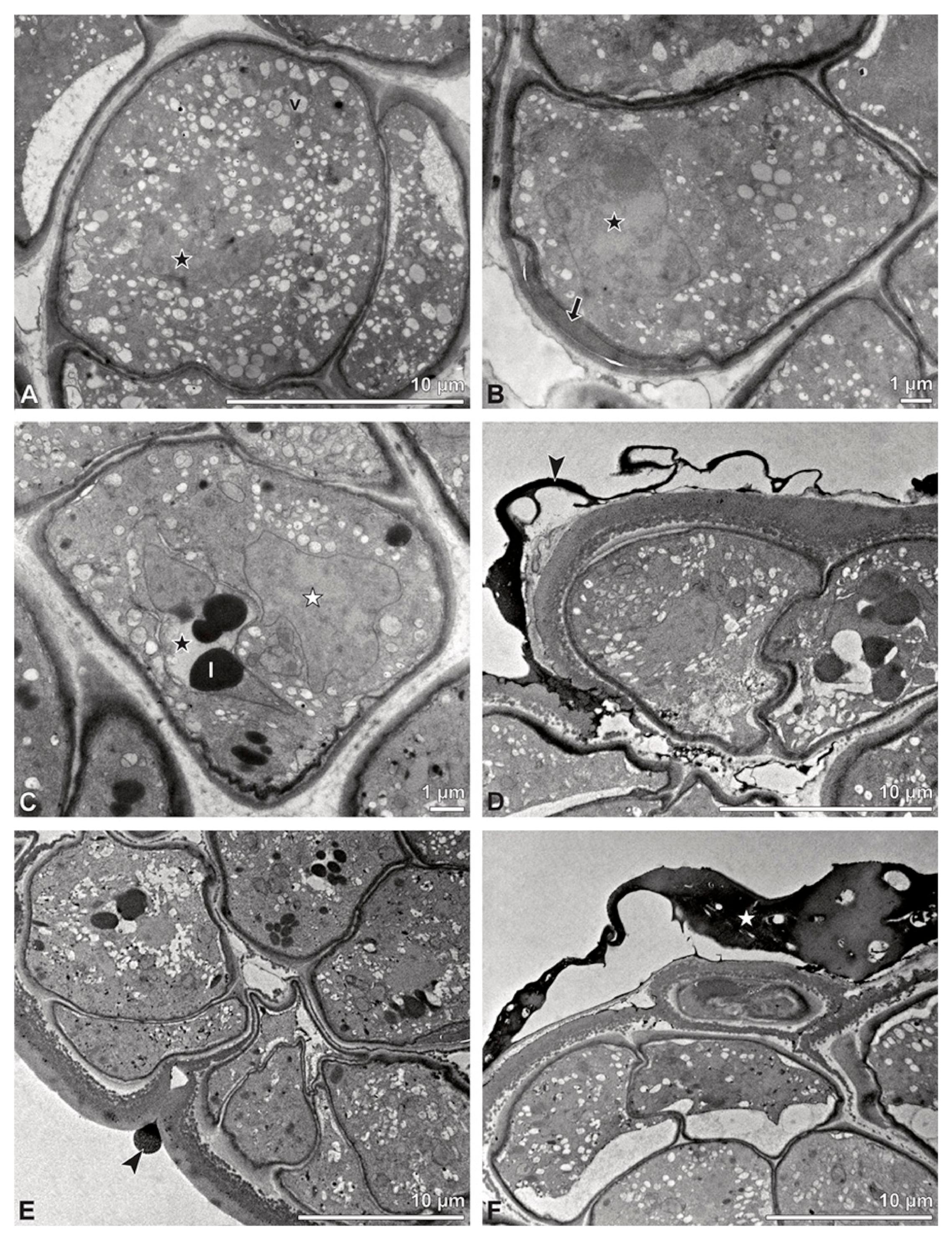
Figure 4.
TEM panorama micrograph showing a cross-section of two adjacent Bulbophyllum retusiusculum pollinia. Note: The pollinia can be divided into three strata based on variations in the tetrad/pollen wall structure and pollen protoplast ultrastructure. This stratification can be traced from the peripheral margin (stratum 1) inwards (stratum 2 and 3). The white arrowhead indicates contact zone of the two pollinia.
Figure 4.
TEM panorama micrograph showing a cross-section of two adjacent Bulbophyllum retusiusculum pollinia. Note: The pollinia can be divided into three strata based on variations in the tetrad/pollen wall structure and pollen protoplast ultrastructure. This stratification can be traced from the peripheral margin (stratum 1) inwards (stratum 2 and 3). The white arrowhead indicates contact zone of the two pollinia.
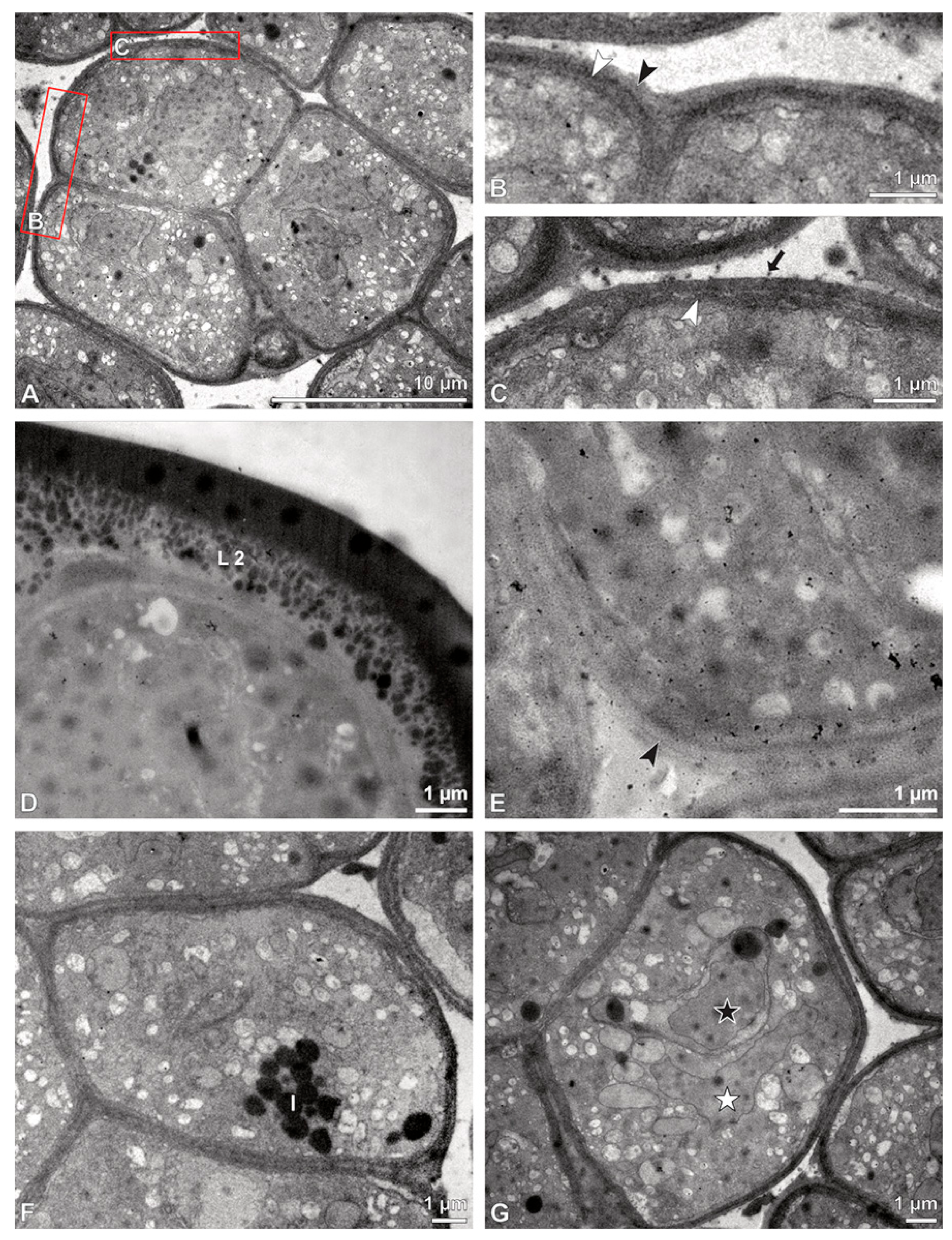
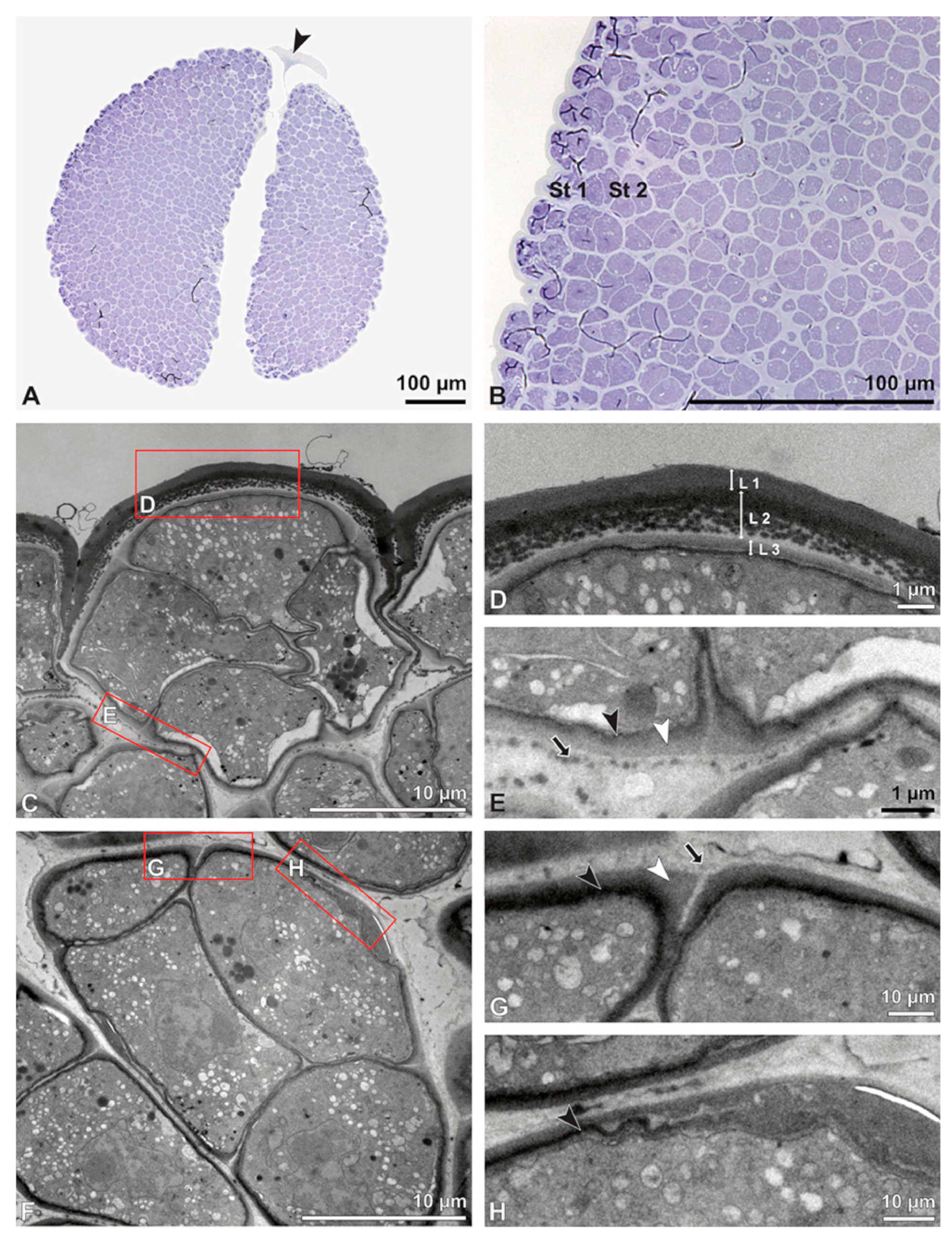
Figure 5.
LM (A,B) and TEM (C–H) micrographs showing cross-sections of Bulbophyllum retusiusculum pollinia/tetrads. Note: (A,B) Semi-thin section of two adjacent pollinia showing different strata (St 1, St 2, and St 3), TBO. (C) Tetrad of outer stratum 1 with detached protoplasts and a three- (D) to two-layered (E) pollen wall; the white arrow points to discontinuous sporopollenin subunits. (D) Detail of pollen wall, showing three layers (L1, L2, and L3). (E) Detail of pollen wall consisting of L2 (black arrowhead) and L3 (intine), with detached undulating areas (white arrowhead). (F) Tetrad of outer stratum 1 close to an adjacent pollinium. (G) Detail of pollen wall, showing three different layers (L1, L2, and L3). (H) Detail of pollen wall, showing L2 (white arrowhead) and L3 (black arrowhead).
Figure 5.
LM (A,B) and TEM (C–H) micrographs showing cross-sections of Bulbophyllum retusiusculum pollinia/tetrads. Note: (A,B) Semi-thin section of two adjacent pollinia showing different strata (St 1, St 2, and St 3), TBO. (C) Tetrad of outer stratum 1 with detached protoplasts and a three- (D) to two-layered (E) pollen wall; the white arrow points to discontinuous sporopollenin subunits. (D) Detail of pollen wall, showing three layers (L1, L2, and L3). (E) Detail of pollen wall consisting of L2 (black arrowhead) and L3 (intine), with detached undulating areas (white arrowhead). (F) Tetrad of outer stratum 1 close to an adjacent pollinium. (G) Detail of pollen wall, showing three different layers (L1, L2, and L3). (H) Detail of pollen wall, showing L2 (white arrowhead) and L3 (black arrowhead).
Figure 6.
TEM micrographs showing cross-sections of Bulbophyllum retusiusculum pollinia/tetrads. Note: (A) Tetrad of stratum 2 with detached pollen protoplasts and two-layered pollen wall. (B) Detail of a pollen wall with two layers: L2 (black arrowhead) and L3 (intine, white arrowhead). (C) Detail of a pollen wall with a distally undulating intine (L3, white arrowhead). (D) Tetrad of the innermost stratum 3. (E). Detail of pollen wall consisting of L2 (black arrowhead) and L3 (white arrowhead) and plasmodesmos (white arrow). (F) Detail of a pollen wall showing the distally undulating intine (L3, white arrowhead). (G) Pollen wall of outer tetrad of stratum 1 with an electron-translucent L2, KMnO4. (H) Contact zone of three tetrads from stratum 3; the black arrowhead points to L2, KMnO4.
Figure 6.
TEM micrographs showing cross-sections of Bulbophyllum retusiusculum pollinia/tetrads. Note: (A) Tetrad of stratum 2 with detached pollen protoplasts and two-layered pollen wall. (B) Detail of a pollen wall with two layers: L2 (black arrowhead) and L3 (intine, white arrowhead). (C) Detail of a pollen wall with a distally undulating intine (L3, white arrowhead). (D) Tetrad of the innermost stratum 3. (E). Detail of pollen wall consisting of L2 (black arrowhead) and L3 (white arrowhead) and plasmodesmos (white arrow). (F) Detail of a pollen wall showing the distally undulating intine (L3, white arrowhead). (G) Pollen wall of outer tetrad of stratum 1 with an electron-translucent L2, KMnO4. (H) Contact zone of three tetrads from stratum 3; the black arrowhead points to L2, KMnO4.
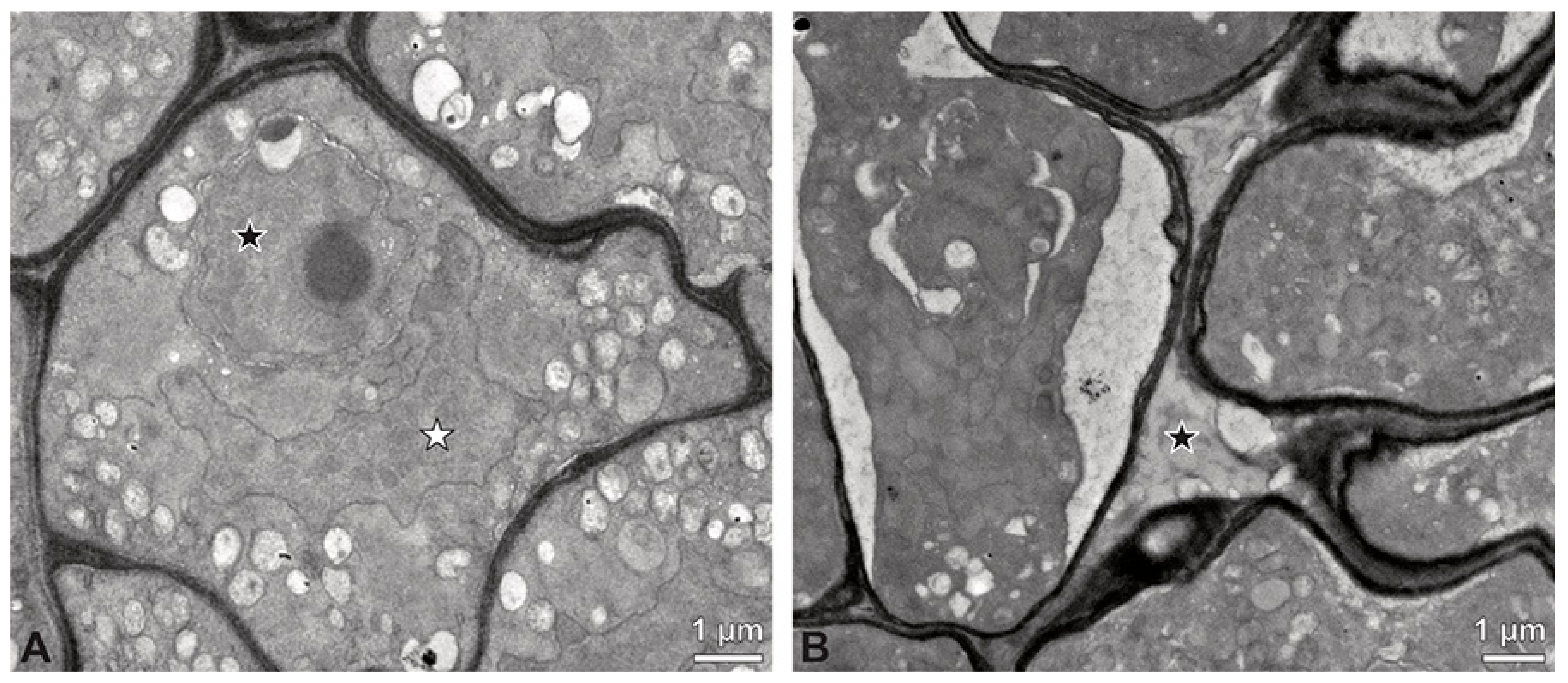
Figure 7.
TEM micrographs showing cross-sections of Bulbophyllum retusiusculum pollinia/tetrads. Note: (A) Mature two-celled pollen grain with generative cell (black star) and vegetative nucleus (white star). (B) Filamentous material between tetrads (black star).
Figure 7.
TEM micrographs showing cross-sections of Bulbophyllum retusiusculum pollinia/tetrads. Note: (A) Mature two-celled pollen grain with generative cell (black star) and vegetative nucleus (white star). (B) Filamentous material between tetrads (black star).
A combined study of the pollinium using LM and TEM shows a loose agglutination of tetrads/monads by pollen coatings (
Figure 10 and
Figure 11A,B). The pollen wall structure of each monad is composed of an ektexine, layer 2, and an intine. The ektexine is subdivided into a discontinuous tectum, columellate infratectum, and thin, continuous foot layer (
Figure 11C,D). At the transition from the interapertural area to the aperture, the foot layer becomes undulating and imbedded in layer 2. An endexine is not present (
Figure 11F,G). The subjacent electron-translucent wall layer 2 and the thin electron-dense intine surround the monads and increase in thickness towards the aperture region (
Figure 11C–E). Layer 2 increases in thickness towards the aperture and shows lamellated electron-dense interstratifications adjacent to the foot layer (
Figure 11C–E). At anthesis, the monads are two-celled (
Figure 8G and
Figure 12A). Reserves in the cytoplasm, like lipids or starch, are absent (
Figure 8H and
Figure 12A). Two different types of pollen coatings are present: electron-dense pollenkitt in the infratectum (
Figure 11D and
Figure 12B) and an additional electron-dense material between the monads (
Figure 12B).
Remarks: The pollinia of
Cephalanthera longifolia and two other species of this genus (
C. damasonium (Mill.) Druce,
C. rubra (L.) Rich.) have been investigated using TEM by Barone Lumaga et al. [
32]. These authors concluded that the ulcerate aperture of
C. longofolia has a distal position and that there is an endexine layer comprising part of the pollen wall. This contradicts our results, which clearly show the aperture to be in a proximal position and that the proclaimed endexine is most likely an intine. Contrary to other species examined in this study,
C. longifolia has a soft pollinium composed of tetrads that disintegrate easily into monads. Also, the structure of the pollen wall is the same regardless of the internal position of the pollen grains within the pollinium. Furthermore, the pollen wall in
C. longifolia is more comparable to the “typical” tectate-collumelate pollen wall of other angiosperms, as portrayed in Halbritter et al. [
10].
Cephalanthera longifolia is the only species of those examined herein that has a reticulate ornamentation and an aperture.
Description of species
Dendrobium × delicatum (F.M. Bailey) F.M. Bailey (
Figure 13,
Figure 14,
Figure 15,
Figure 16,
Figure 17 and
Figure 18;
Table 1,
Table 2 and
Table 3), lower epidendroids: The dispersal unit is an elliptical-shaped pollinium 1.5 mm in length and 0.5 mm in width (SEM) (
Figure 14A,
Table 1). It consists of tetrads (
Figure 14B) adhered together by pollen coatings (
Figure 14D). Various tetrad types were observed within the pollinium: tetrahedral, planar-tetragonal, planar-linear, and decussate (
Figure 13B). Under LM, the size of the hydrated pollen grains within the tetrads ranges from 12.6 µm to 15.9 µm (
Table 1). The monads are inaperturate and two-celled at anthesis (
Figure 13D and
Figure 17G). Staining with basic fuchsin in LM revealed differences in the pollen wall structure of the tetrads, depending on their position within the pollinium (
Figure 13C).
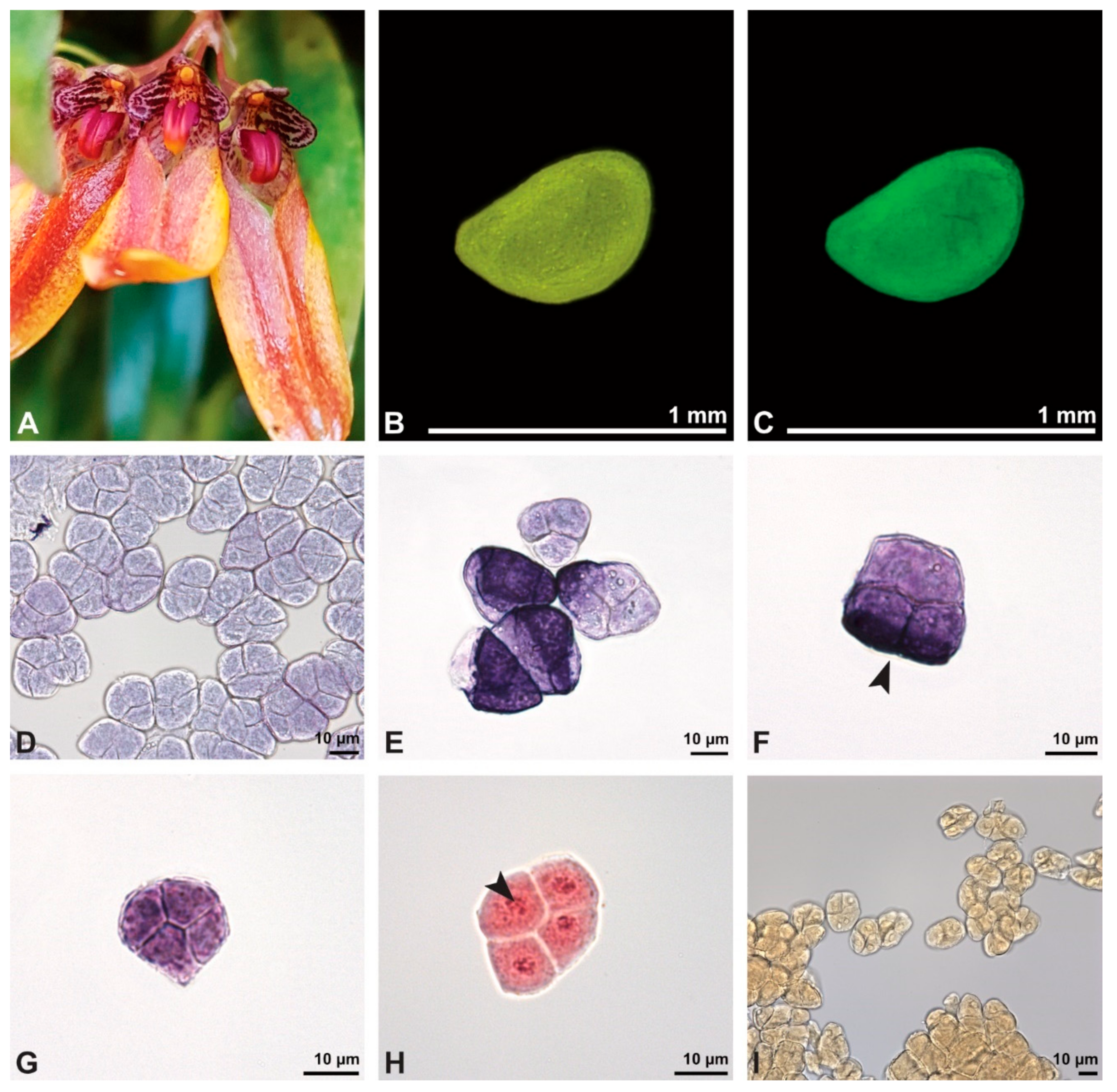
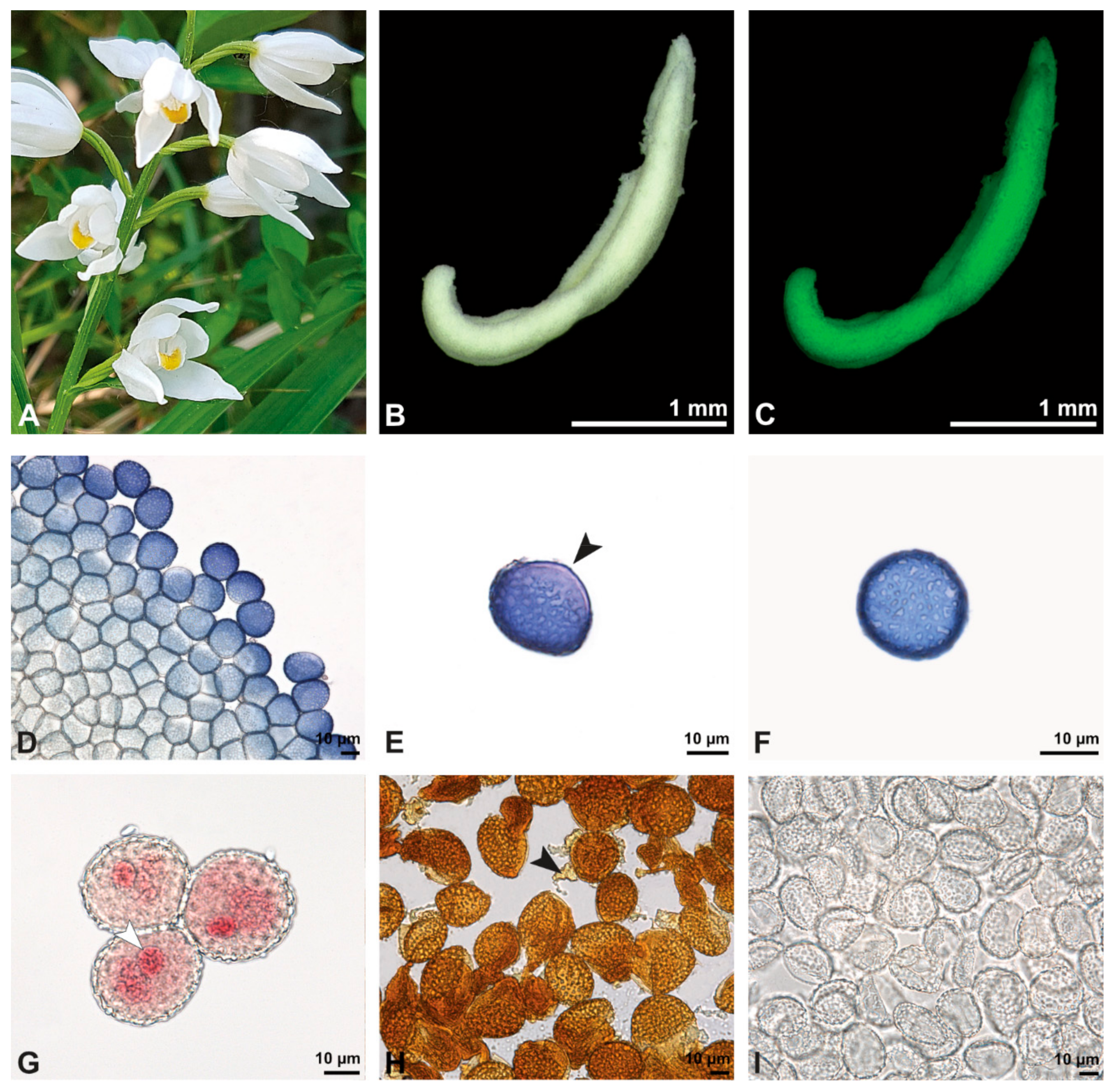
Figure 8.
Macroscopic image of inflorescence/flowers (A) and LM micrographs (B–I) of Cephalanthera longifolia pollen. (B) Pollinium. (C) Pollinium fluorescing under UV light. (D) Arrangement of monads within pollinium, TBO. (E) Ulcerate (black arrowhead) pollen grain from an equatorial view, reticulate ornamentation in interapertural area, TBO. (F) Pollen grain from a polar view, TBO. (G) Binucleate pollen and its generative cell (white arrowhead), Carmine. (H) No starch detected; pollen coatings connect tetrads (black arrowhead), Lugol. (I) Hydrated reticulate pollen.
Figure 8.
Macroscopic image of inflorescence/flowers (A) and LM micrographs (B–I) of Cephalanthera longifolia pollen. (B) Pollinium. (C) Pollinium fluorescing under UV light. (D) Arrangement of monads within pollinium, TBO. (E) Ulcerate (black arrowhead) pollen grain from an equatorial view, reticulate ornamentation in interapertural area, TBO. (F) Pollen grain from a polar view, TBO. (G) Binucleate pollen and its generative cell (white arrowhead), Carmine. (H) No starch detected; pollen coatings connect tetrads (black arrowhead), Lugol. (I) Hydrated reticulate pollen.
In SEM, the outermost tetrads exhibit thick wall elements with an outer layer displaying either a psilate, perforate to fossulate (
Figure 14D), or granulate ornamentation (
Figure 14C). Compared to the outer tetrads, the inner tetrads consist of a thin pollen wall only (
Figure 13C).
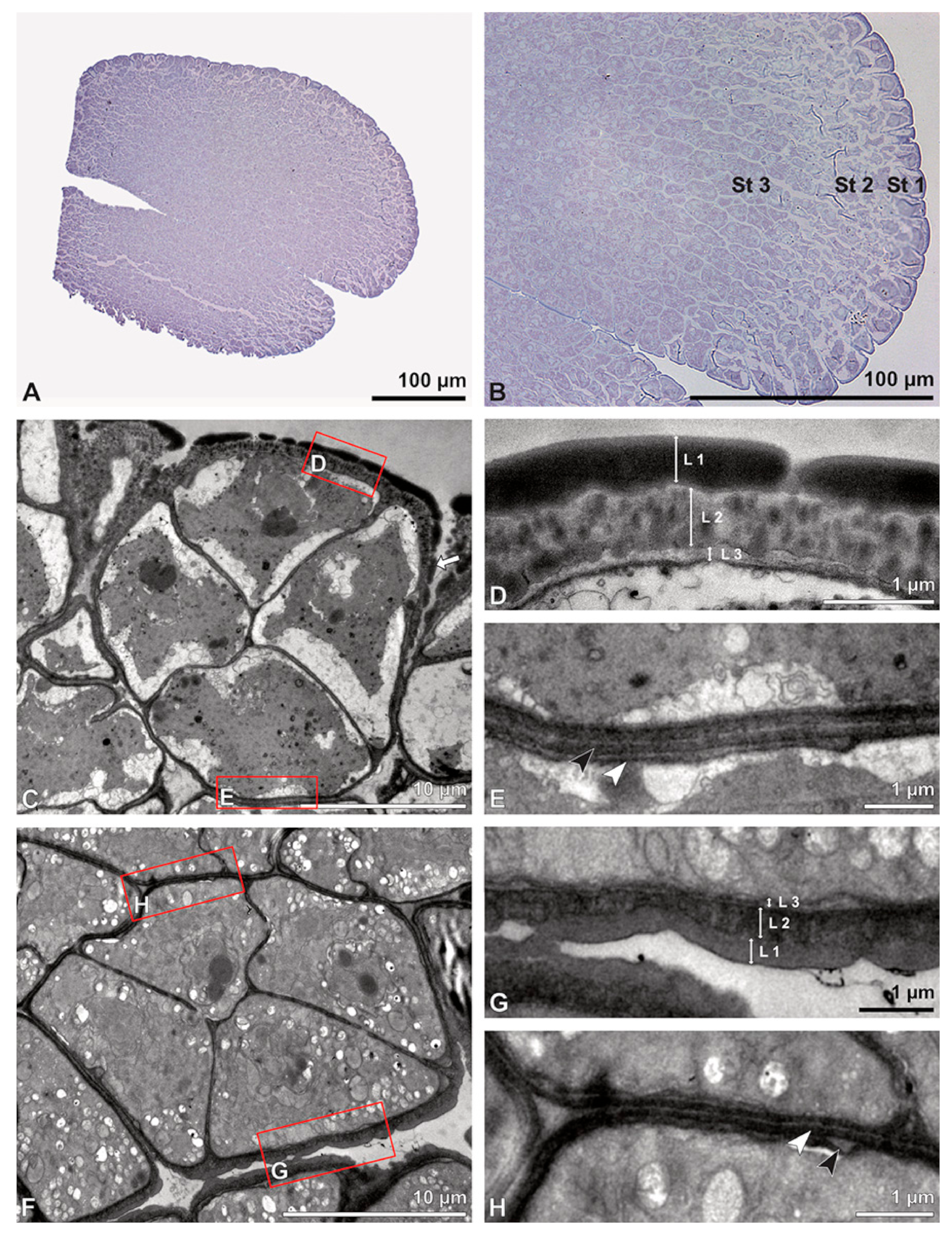
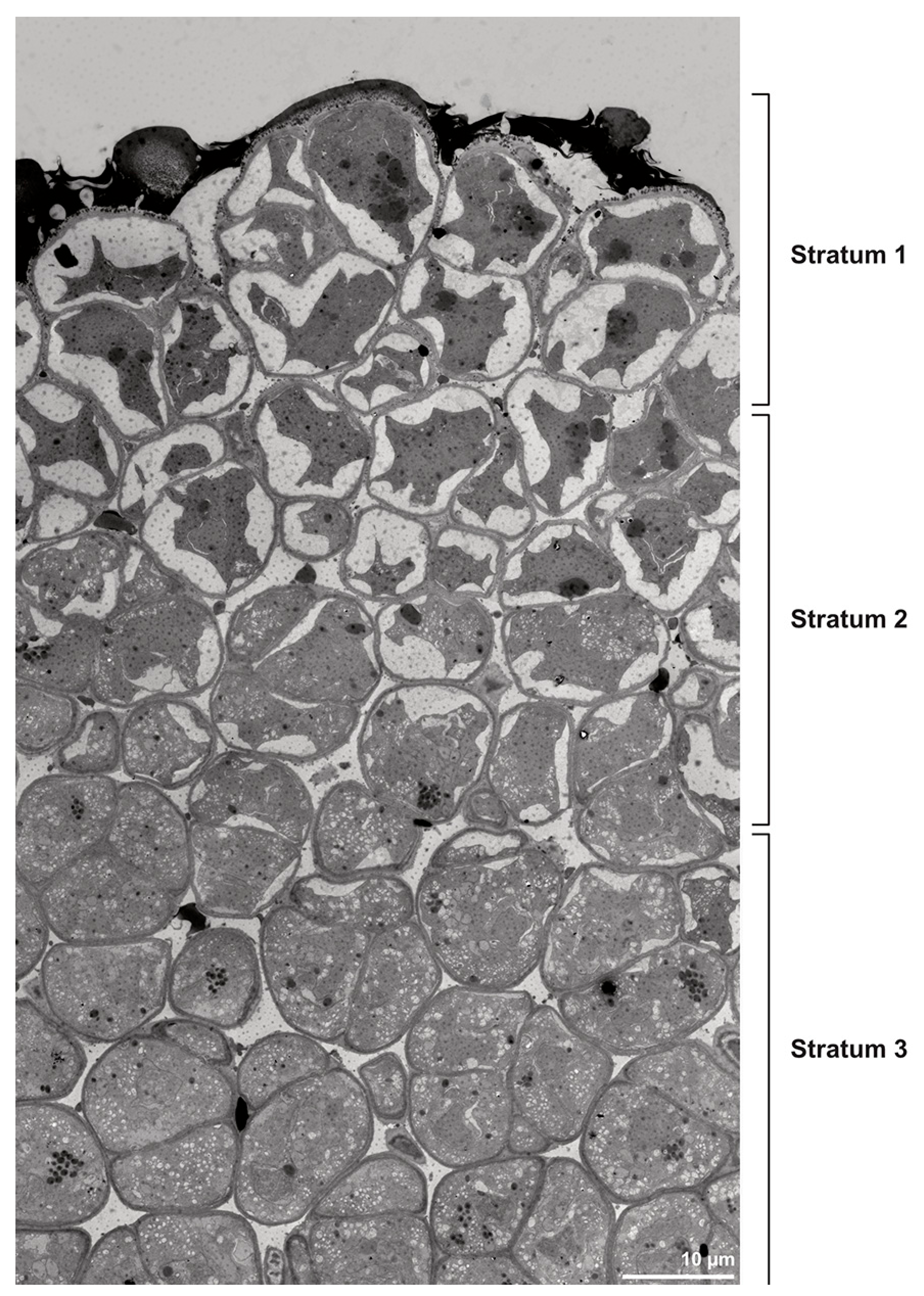
These variations are also clearly visible in TEM, which revealed differences in the cell content as well. Based on this combined study, the pollinium can be divided into three distinct strata, from the peripheral margin to the inside (
Figure 15). Outermost thick-walled tetrads with detached protoplasts characterize the outer stratum 1. In the adjacent stratum 2, the tetrads are thin-walled and the protoplasts are also detached, whereas the inner tetrads of stratum 3 are also thin-walled but with intact protoplasts (
Figure 15 and
Figure 16A,B). The tetrads of the outermost stratum 1 constitute pollen walls with a three-layered structure (
Figure 16C,D). The outer compact sporopollenin wall layer (layer 1) forms a continuous compact wall layer, but only on the distal half of the outer pollen grains of the tetrad (
Figure 16C). This layer decreases in thickness towards the contact zones of adjacent tetrads and transitions into sporopollenin granules that extend inwardly into an electron-translucent pollen wall layer (layer 2) subjacent to layer 1 (
Figure 16C,D). Layer 2 surrounds each pollen grain (monad) and forms a common layer around each tetrad. As staining with potassium permanganate did not indicate a lipidic nature of layer 2, it is most likely polysaccharidic (
Figure 17D,E).The inner layer (layer 3) is most likely a monolayered intine.
Figure 9.
SEM micrographs of hydrated Cephalanthera longifolia pollinium/pollen (EtOH + CPD). Note: (A) Overview of pollinium. (B) Arrangement of monads in the pollinium; remnant of a tetrad (white arrowhead). (C) Distal pole with reticulate, heterobrochate ornamentation. (D) Oblique polar view of a monad with ornamented an aperture membrane (white star) and a reticulate interapertural area showing a columellate infratectum (white arrowhead).
Figure 9.
SEM micrographs of hydrated Cephalanthera longifolia pollinium/pollen (EtOH + CPD). Note: (A) Overview of pollinium. (B) Arrangement of monads in the pollinium; remnant of a tetrad (white arrowhead). (C) Distal pole with reticulate, heterobrochate ornamentation. (D) Oblique polar view of a monad with ornamented an aperture membrane (white star) and a reticulate interapertural area showing a columellate infratectum (white arrowhead).
Figure 10.
TEM panorama micrograph showing the cross-section of a Cephalanthera longifolia pollinium.
Figure 10.
TEM panorama micrograph showing the cross-section of a Cephalanthera longifolia pollinium.
Figure 11.
LM (A,B) and TEM (C–G) micrographs showing the cross-sections of a Cephalanthera longifolia pollinium/monad. Note: (A) Semi-thin section of pollinium showing monads, TBO. (B) Semi-thin section showing the sticky material (black arrowhead) connecting monads, TBO. (C) Overview of a monad. (D) Detail of pollen wall in interapertural area, showing semitectate tectum (t), columellate infratectum (inf), continuous foot layer (f), subjacent layer 2 (L2), and a thin intine (i). (E) Detail of pollen wall in aperture area; ornamented aperture membrane (black arrow) consisting of intine (i), layer 2 (L2) with lamellated interstratifications (black arrowhead), and a thin foot layer (f). (F) Pollen wall in the interapertural area, KMnO4. (G) Pollen wall in the transition zone between the interapertural and aperture areas, KMnO4.
Figure 11.
LM (A,B) and TEM (C–G) micrographs showing the cross-sections of a Cephalanthera longifolia pollinium/monad. Note: (A) Semi-thin section of pollinium showing monads, TBO. (B) Semi-thin section showing the sticky material (black arrowhead) connecting monads, TBO. (C) Overview of a monad. (D) Detail of pollen wall in interapertural area, showing semitectate tectum (t), columellate infratectum (inf), continuous foot layer (f), subjacent layer 2 (L2), and a thin intine (i). (E) Detail of pollen wall in aperture area; ornamented aperture membrane (black arrow) consisting of intine (i), layer 2 (L2) with lamellated interstratifications (black arrowhead), and a thin foot layer (f). (F) Pollen wall in the interapertural area, KMnO4. (G) Pollen wall in the transition zone between the interapertural and aperture areas, KMnO4.
Figure 12.
TEM micrographs showing cross-sections of a Cephalanthera longifolia pollinium/monad. Note: (A) Mature two-celled pollen grain with a vegetative nucleus (white star) and generative cell (black star). (B) Monads with two different pollen coatings, electron-dense in the infratectum (black arrow) and between monads (white star).
Figure 12.
TEM micrographs showing cross-sections of a Cephalanthera longifolia pollinium/monad. Note: (A) Mature two-celled pollen grain with a vegetative nucleus (white star) and generative cell (black star). (B) Monads with two different pollen coatings, electron-dense in the infratectum (black arrow) and between monads (white star).
Figure 13.
Macroscopic image of flowers (A) and LM micrographs of Dendrobium × delicatum pollen (B–E). (B) Various tetrad types, basic fuchsin. (C) Outer tetrad with sporopollenin exine stained dark violet (black arrowhead) and inner thin-walled tetrad, basic fuchsin. (D) Binucleate pollen and its generative cell (white arrowhead), Carmine. (E) Pollen coatings present (black arrowhead); no starch detected, Lugol.
Figure 13.
Macroscopic image of flowers (A) and LM micrographs of Dendrobium × delicatum pollen (B–E). (B) Various tetrad types, basic fuchsin. (C) Outer tetrad with sporopollenin exine stained dark violet (black arrowhead) and inner thin-walled tetrad, basic fuchsin. (D) Binucleate pollen and its generative cell (white arrowhead), Carmine. (E) Pollen coatings present (black arrowhead); no starch detected, Lugol.
Figure 14.
SEM micrographs of Dendrobium × delicatum pollen. Note: (A) Overview of hydrated pollinium (DMP + CPD). (B) Arrangement of tetrads within pollinium. (C) Tetrahedral tetrad with granulate to microverrucate ornamentation (white arrowhead). (D) Outer tetrads with psilate ornamentation and pollen coatings (white arrowhead). (E) Anther cap. (F) Inner surface of anther cap without Ubisch bodies.
Figure 14.
SEM micrographs of Dendrobium × delicatum pollen. Note: (A) Overview of hydrated pollinium (DMP + CPD). (B) Arrangement of tetrads within pollinium. (C) Tetrahedral tetrad with granulate to microverrucate ornamentation (white arrowhead). (D) Outer tetrads with psilate ornamentation and pollen coatings (white arrowhead). (E) Anther cap. (F) Inner surface of anther cap without Ubisch bodies.
Figure 15.
TEM panorama micrograph showing a cross-section of a Dendrobium × delicatum pollinium. The pollinium can be divided into three strata based on the variations in its tetrad/pollen wall structure and pollen protoplast ultrastructure. This stratification can be traced from the peripheral margin (stratum 1) inward (stratum 2 and 3).
Figure 15.
TEM panorama micrograph showing a cross-section of a Dendrobium × delicatum pollinium. The pollinium can be divided into three strata based on the variations in its tetrad/pollen wall structure and pollen protoplast ultrastructure. This stratification can be traced from the peripheral margin (stratum 1) inward (stratum 2 and 3).
Figure 16.
LM (A,B) and TEM (C–H) micrographs showing cross-sections of Dendrobium × delicatum pollinium/tetrads. Note: (A,B). Semi-thin sections of pollinium, showing different strata (St 1, St 2, St 3), TBO. (C). Tetrad of outer stratum 1 with detached protoplasts and three- (D) to two-layered (E) pollen walls. (D) Detail of pollen wall with three layers (L1, L2, L3). (E) Detail of pollen wall with layer 2 (black arrowhead) with loosely scattered granules (black arrow) and layer 3 (intine, white arrowhead). (F) Tetrad of stratum 2 with detached protoplasts and two-layered pollen wall. (G) Detail of pollen wall, layer 2 (L2, black arrowhead) with granules (black arrow) and layer 3 (L3, intine, white arrowhead). (H) Detail of pollen wall with distally undulating area of L3 (intine, white arrowhead).
Figure 16.
LM (A,B) and TEM (C–H) micrographs showing cross-sections of Dendrobium × delicatum pollinium/tetrads. Note: (A,B). Semi-thin sections of pollinium, showing different strata (St 1, St 2, St 3), TBO. (C). Tetrad of outer stratum 1 with detached protoplasts and three- (D) to two-layered (E) pollen walls. (D) Detail of pollen wall with three layers (L1, L2, L3). (E) Detail of pollen wall with layer 2 (black arrowhead) with loosely scattered granules (black arrow) and layer 3 (intine, white arrowhead). (F) Tetrad of stratum 2 with detached protoplasts and two-layered pollen wall. (G) Detail of pollen wall, layer 2 (L2, black arrowhead) with granules (black arrow) and layer 3 (L3, intine, white arrowhead). (H) Detail of pollen wall with distally undulating area of L3 (intine, white arrowhead).
Figure 17.
TEM micrographs showing cross-sections of Dendrobium × delicatum pollinium/tetrads. Note: (A) Tetrad of inner stratum 3 with reduced pollen wall. (B). Detail of pollen wall with layer 2 (L2, black arrowhead) and layer 3 (L3, intine, white arrowhead). (C) Detail of pollen wall with distally undulating layer 3 (intine, white arrowhead) and granules (black arrow) attached to layer 2. (D) Pollen wall of tetrad from outer stratum 1, with granules embedded in layer 2 (L2), KMnO4. (E) Pollen wall of tetrad from inner stratum 3; arrowhead points to L2, KMnO4. (F) Pollen grain with lipid droplets (l). (G) Mature two-celled pollen grain with a generative cell (black star) and vegetative nucleus (white star).
Figure 17.
TEM micrographs showing cross-sections of Dendrobium × delicatum pollinium/tetrads. Note: (A) Tetrad of inner stratum 3 with reduced pollen wall. (B). Detail of pollen wall with layer 2 (L2, black arrowhead) and layer 3 (L3, intine, white arrowhead). (C) Detail of pollen wall with distally undulating layer 3 (intine, white arrowhead) and granules (black arrow) attached to layer 2. (D) Pollen wall of tetrad from outer stratum 1, with granules embedded in layer 2 (L2), KMnO4. (E) Pollen wall of tetrad from inner stratum 3; arrowhead points to L2, KMnO4. (F) Pollen grain with lipid droplets (l). (G) Mature two-celled pollen grain with a generative cell (black star) and vegetative nucleus (white star).
Figure 18.
TEM micrographs showing cross-sections of Dendrobium × delicatum pollinium/tetrads. Note: (A) Pollen coatings (white star) attached to the tetrads of the outer stratum 1. (B) Pollen coatings (white arrowhead) between the tetrads of stratum 2 (with detached protoplasts) and the inner stratum 3.
Figure 18.
TEM micrographs showing cross-sections of Dendrobium × delicatum pollinium/tetrads. Note: (A) Pollen coatings (white star) attached to the tetrads of the outer stratum 1. (B) Pollen coatings (white arrowhead) between the tetrads of stratum 2 (with detached protoplasts) and the inner stratum 3.
The inner pollen grains of the outer tetrads exhibit a reduced pollen wall made of a layer 2 with loosely scattered sporopollenin elements and a monolayered layer 3 (
Figure 16E). The pollen grains in the outer tetrads have detached protoplasts lacking intact nuclei and organelles, which indicates that they have no germination function (
Figure 15). Stratum 2 consists of tetrads with reduced pollen walls (
Figure 16F), comprising only layer 2 and layer 3 (
Figure 16G). Wall layer 2 contains loosely scattered embedded sporopollenin granules, surrounding the monads and additionally forming a common layer around the tetrads (
Figure 16F,G). The subjacent wall layer 3 passes into a undulated area in the distal half of the pollen grains (
Figure 16H). Similar to the outer tetrads of stratum 1, the pollen grains in this region have detached protoplasts and are poor in organelles, indicating that they have no germination function (
Figure 15). The tetrads of stratum 3 (
Figure 17A) share similarities in their pollen wall structure to those of stratum 2, except that wall layer 2 exhibits very few or no sporopollenin granules (
Figure 17B,C). Unlike the tetrads of stratum 1 and 2, the pollen protoplasts are rich in organelles (such as nuclei, vacuoles, lipids) and appear to have a germination function (
Figure 17F). Starch reserves have not been detected at this stage of pollen development (
Figure 13E). Pollen coatings are primarily attached to the ektexine of the outer tetrads (stratum 1) (
Figure 18A), with their quantity decreasing inwardly (
Figure 15). Droplets of similar electron density are also observed in the spaces between the tetrads of all three strata, suggesting the presence of pollen coatings (
Figure 15 and
Figure 18B). Staining with potassium iodine in LM also revealed the presence of pollen coatings (
Figure 13E). Ubisch bodies were not found in the anther cap in SEM (
Figure 14E,F).
Remarks: Pollen studies of
Dendrobium are rare. Schill and Pfeiffer [
24] investigated 28 species including
Dendrobium ×
delicatum using SEM and described their ornamentation as predominantly psilate, which is in accordance with our results. The LM studies of
D. kingianum Bidwill ex. Lindl conducted by Wolter and Schill [
29] and the TEM studies of
D. nobile Lindl. by Zavada [
39] revealed a reduction in the pollen wall structure from the peripheral to the inner tetrads of the pollinia. The present study confirms this and furthermore specifies this reduction (in wall thickness and sporopollenin content) in the wall structure from the peripheral tetrads of stratum 1 to the tetrads of the inner strata 2 and 3. The pollen wall of
D. ×
delicatum deviates from the “typical” angiosperm pollen wall model (see [
10]) by having a differently structured intine, comprising both an endintine and ectintine. Additionally, protoplast detachment extends over strata 1 and 2, similar to
Bulbophyllum retusiusculum.
Description of species
Oncidium crocidipterum (Rchb. f.) M.W.Chase & N.H.Williams (
Figure 19,
Figure 20,
Figure 21,
Figure 22,
Figure 23 and
Figure 24;
Table 1,
Table 2 and
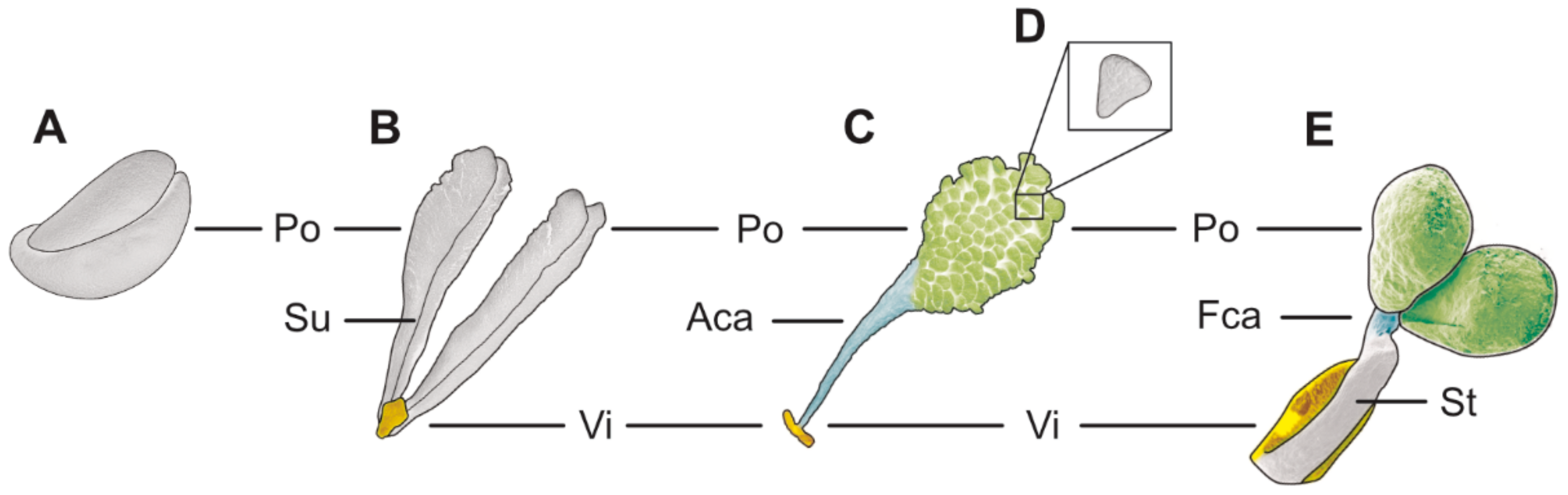
Table 3), higher epidendroids: The pollinarium is irregularly shaped and consists of two pear-shaped pollinia and an appendage, measuring 3.75 mm in length under LM (
Figure 19B and
Figure 20A,
Table 1). Each pollinium shows a posterior suture running from the base to the apex (
Figure 20B). The pollinia, 1.5 mm long and 0.98 mm wide (LM), are attached to a 1.82 mm long (LM) cellular appendage referred to as a stipe (
Figure 19B and
Figure 20A). Both the pollinia and stipe show autofluorescence under LM (
Figure 19C). The frenicular caudicle connects the stipe and pollinium (
Figure 20C). Each pollinium is composed of tetrads agglutinated by pollen coatings (
Figure 24B). Different types of tetrads have been documented: tetrahedral, planar-tetragonal, planar-rhomboid, and decussate (
Figure 19D). In LM, the size range of the hydrated pollen grains within a tetrad is between 12.9 µm and 25.6 µm (
Table 1). Mature pollen grains are inaperturate and two-celled at anthesis, and pollen reserves like lipids or starch have not been detected at this stage (
Figure 19H,I). In LM, germination was observed on pollen hydrated in water and toluidine blue stain (
Figure 19G). Staining with toluidine blue shows the differentiation in the pollen wall structure of tetrads depending on their position within the pollinium (
Figure 19 E,F).
The outermost tetrads exhibit a thick pollen wall in LM (
Figure 19E), with an outer wall layer displaying a psilate, perforate ornamentation in SEM (
Figure 20D). In contrast to the outer tetrads, the inner tetrads have a thin pollen wall (
Figure 19F).
The TEM studies revealed three different strata, based on the differentiation in the pollen’s wall structure: the outermost stratum 1 consists of thick-walled outer tetrads, stratum 2 consists of tetrads with reduced pollen walls, and the innermost stratum 3 comprises tetrads with highly reduced pollen walls (
Figure 21 and
Figure 22B). The tetrads of the outermost stratum 1 exhibit a three-layered pollen wall structure and show slightly different ektexine thicknesses depending on their position (intersutural-intrasutural) within the pollinium (
Figure 22A). The tetrads in the intersutural area (
Figure 22C) constitute a compact, continuous sporopollenin layer (layer 1), located only on the distal half of the outer pollen grains of the tetrad (
Figure 22D). Towards the contact zones of neighbouring outer tetrads of the pollinium, this compact layer 1 transitions into sporopollenin granules, which decrease inwardly in quantity and density (
Figure 22C). The thick, compact electron-translucent layer 2 shows embedded sporopollenin granules with a decreasing density gradient from layer 1 to layer 3 (
Figure 22D). This layer is remarkably thicker distally (
Figure 22C) and surrounds each monad, also forming a common wall layer around each tetrad (
Figure 22C–E). The embedded scattered granules in layer 2 decrease in density inwardly (
Figure 22C). These granules are similar in electron density to layer 1 (
Figure 22D) and, when stained with potassium permanganate, they become electron-dense in TEM, indicating their lipidic nature (
Figure 23E). From a chemical point of view, layer 2 is most likely composed of polysaccharides, since staining with potassium permanganate did not indicate that it has a lipidic nature (
Figure 23E,F). The inner pollen wall layer (layer 3) likely constitutes a monolayered intine (
Figure 22D,E). Contrary to the outer tetrads in the intersutural area, outer tetrads in the intrasutural area (
Figure 22A) display an ektexine composed of a thinner and more electron-translucent distal layer 1 and a subjacent distally thicker layer 2 with only a few embedded sporopollenin granules (
Figure 22A,F). Layer 3 is a monolayered intine (
Figure 22G). The inner pollen grains of the outer tetrads of both areas, intersutural and intrasutural, have a reduced pollen wall made of a thin layer 2 with scattered embedded granules and a thin monolayered layer 3 (intine) (
Figure 22E,H). The pollen grains of stratum 1 show detached protoplasts containing a vegetative nucleus, a generative cell, and many small vacuoles (
Figure 22C,F). Stratum 2 consists of tetrads with a reduced pollen wall structure, composed of layer 2 and layer 3 (
Figure 23A,B). Layer 2, displaying embedded loosely scattered granules, constitutes the outer pollen wall of each monad and forms a common layer around the tetrads (
Figure 23A,B). The inner layer 3 is monolayered (
Figure 23B). As with tetrads of stratum 1, the detached protoplasts and ultrastructure of the cells’ content suggest infertility (
Figure 23A). The tetrads of stratum 3 display highly reduced pollen walls composed of a layer 2 with only a few sparsely embedded granules and a monolayered layer 3 (
Figure 23C,D). Layer 2 surrounds each monad, varies in thickness, and forms a common layer around each tetrad (
Figure 23C). The protoplasts of the pollen grains are detached, containing a vegetative nucleus, a generative cell, and many small vacuoles (
Figure 23C). An additional thin continuous electron-dense layer surrounds all tetrads of all strata (
Figure 22E and
Figure 23B–D). Pollen coatings are present primarily in the intrasutural area attached to the outer tetrads and also between the tetrads of stratum 1 (
Figure 24A). The spaces between tetrads of all strata are filled with an electron-translucent material, probably elastoviscin (
Figure 24B).
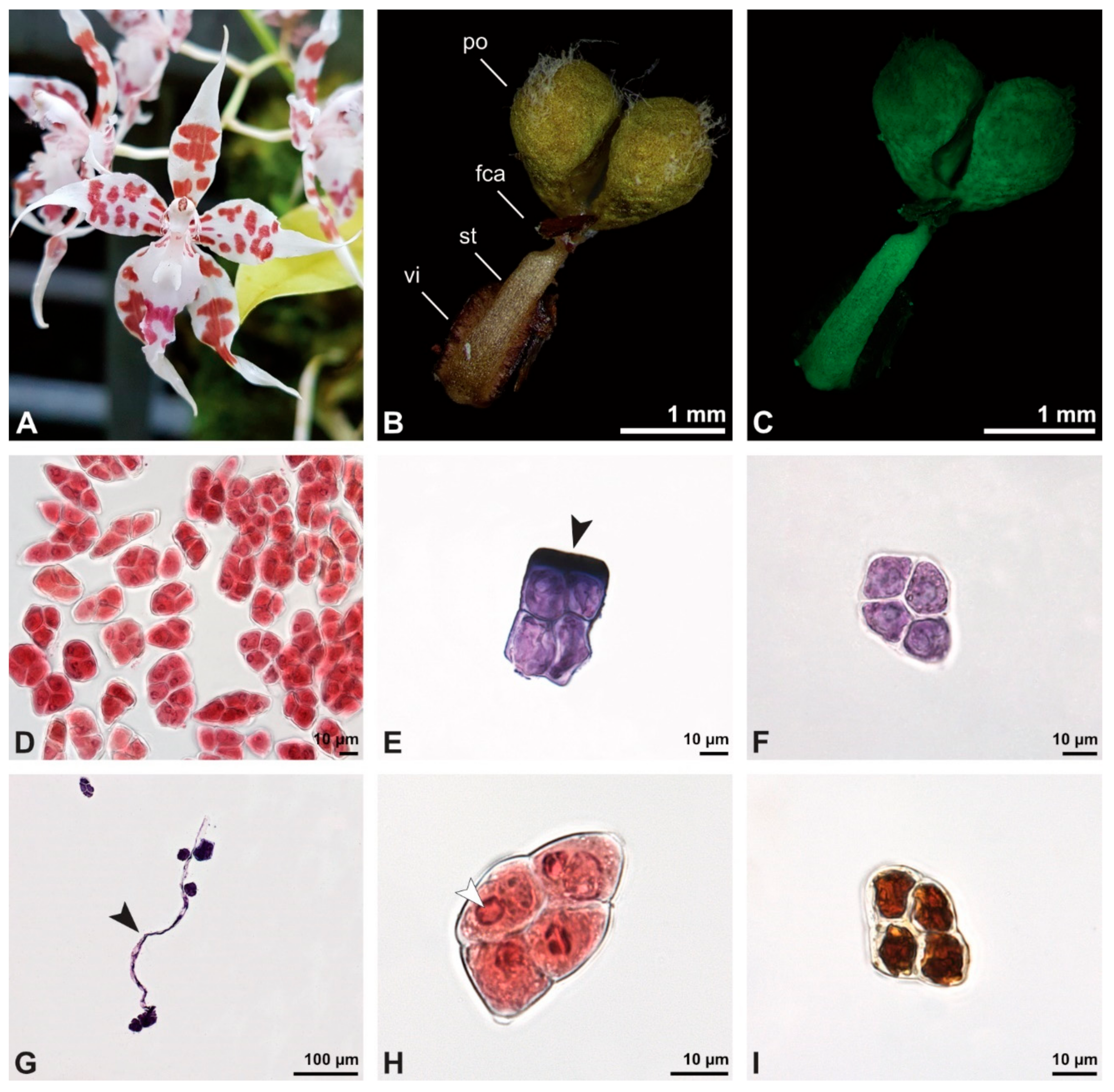
Figure 19.
Macroscopic image of flowers (A) and LM micrographs of Oncidium crocidipterum pollen (B–I). (B) Pollinarium consisting of two pollinia (po), a frenicular caudicle (fca), stipe (st), and viscidium (vi). (C) Pollinarium fluorescing under UV light. (D) Various tetrad types, Carmine. (E) Outer (planar-tetragonal) tetrad with its exine stained dark blue (black arrowhead), TBO. (F) Inner tetrad with thin pollen wall, TBO. (G) Germinating pollen grain with pollen tube (black arrowhead), TBO. (H) Binucleate pollen with a generative cell (white arrowhead), Carmine. (I) No starch detected, Lugol.
Figure 19.
Macroscopic image of flowers (A) and LM micrographs of Oncidium crocidipterum pollen (B–I). (B) Pollinarium consisting of two pollinia (po), a frenicular caudicle (fca), stipe (st), and viscidium (vi). (C) Pollinarium fluorescing under UV light. (D) Various tetrad types, Carmine. (E) Outer (planar-tetragonal) tetrad with its exine stained dark blue (black arrowhead), TBO. (F) Inner tetrad with thin pollen wall, TBO. (G) Germinating pollen grain with pollen tube (black arrowhead), TBO. (H) Binucleate pollen with a generative cell (white arrowhead), Carmine. (I) No starch detected, Lugol.
Remarks: To date, there are no pollen studies on this genus. In contrast to the investigated species above, the PDU type of
Oncidium crocidipterum constitutes a pollinarium (versus pollinia) with two compact pollinia, a frenicular caudicle, a stipe, and a viscidium. The pollen wall’s structure deviates from the “typical” angiosperm pollen wall model (see [
10]) and shows a reduction from the peripheral zones to the centre of the pollinium, as observed in
Bulbophyllum retusiusculum and
Dendrobium ×
delicatum. Similar to
D. ×
delicatum, strata 2 and 3 exhibit only sporopollenin granules that decrease in size and quantity towards the centre of the pollinia. In contrast to
B. retusiusculum and
D. ×
delicatum, there is an additional difference in the wall structure of the outermost tetrads from stratum 1 between the inter- and intrasutural area in
O. crocidipterum. Only in this species does a thin electron-dense layer occur between tetrads, most likely constituting a remnant from the pollen mother cell. Like in
Cephalanthera longifolia, two different types of sticky materials are present within the pollinium.
Figure 20.
SEM micrographs of Oncidium crocidipterum pollinarium/pollen. Note: (A) Overview of hydrated (DMP + CPD) pollinarium consisting of two pollinia (po), a frenicular caudicle (fca), stipe (st), and viscidium (vi). (B) Dorsal view of pollinarium with two pollinia, each showing a suture (white arrowheads). (C) Detail of frenicular caudicle connecting the pollinia and stipe (white arrowhead). (D) Exine of outer tetrads with psilate to granulate (arrowhead) ornamentation.
Figure 20.
SEM micrographs of Oncidium crocidipterum pollinarium/pollen. Note: (A) Overview of hydrated (DMP + CPD) pollinarium consisting of two pollinia (po), a frenicular caudicle (fca), stipe (st), and viscidium (vi). (B) Dorsal view of pollinarium with two pollinia, each showing a suture (white arrowheads). (C) Detail of frenicular caudicle connecting the pollinia and stipe (white arrowhead). (D) Exine of outer tetrads with psilate to granulate (arrowhead) ornamentation.
Figure 21.
TEM panorama micrograph showing a cross-section of an Oncidium crocidipterum pollinium. Note: The pollinium is divided into three strata based on variations in its tetrad/pollen wall structure. Stratum 1 is composed of thick-walled tetrads, stratum 2 of tetrads with reduced pollen walls, and stratum 3 shows tetrads with highly reduced pollen walls.
Figure 21.
TEM panorama micrograph showing a cross-section of an Oncidium crocidipterum pollinium. Note: The pollinium is divided into three strata based on variations in its tetrad/pollen wall structure. Stratum 1 is composed of thick-walled tetrads, stratum 2 of tetrads with reduced pollen walls, and stratum 3 shows tetrads with highly reduced pollen walls.
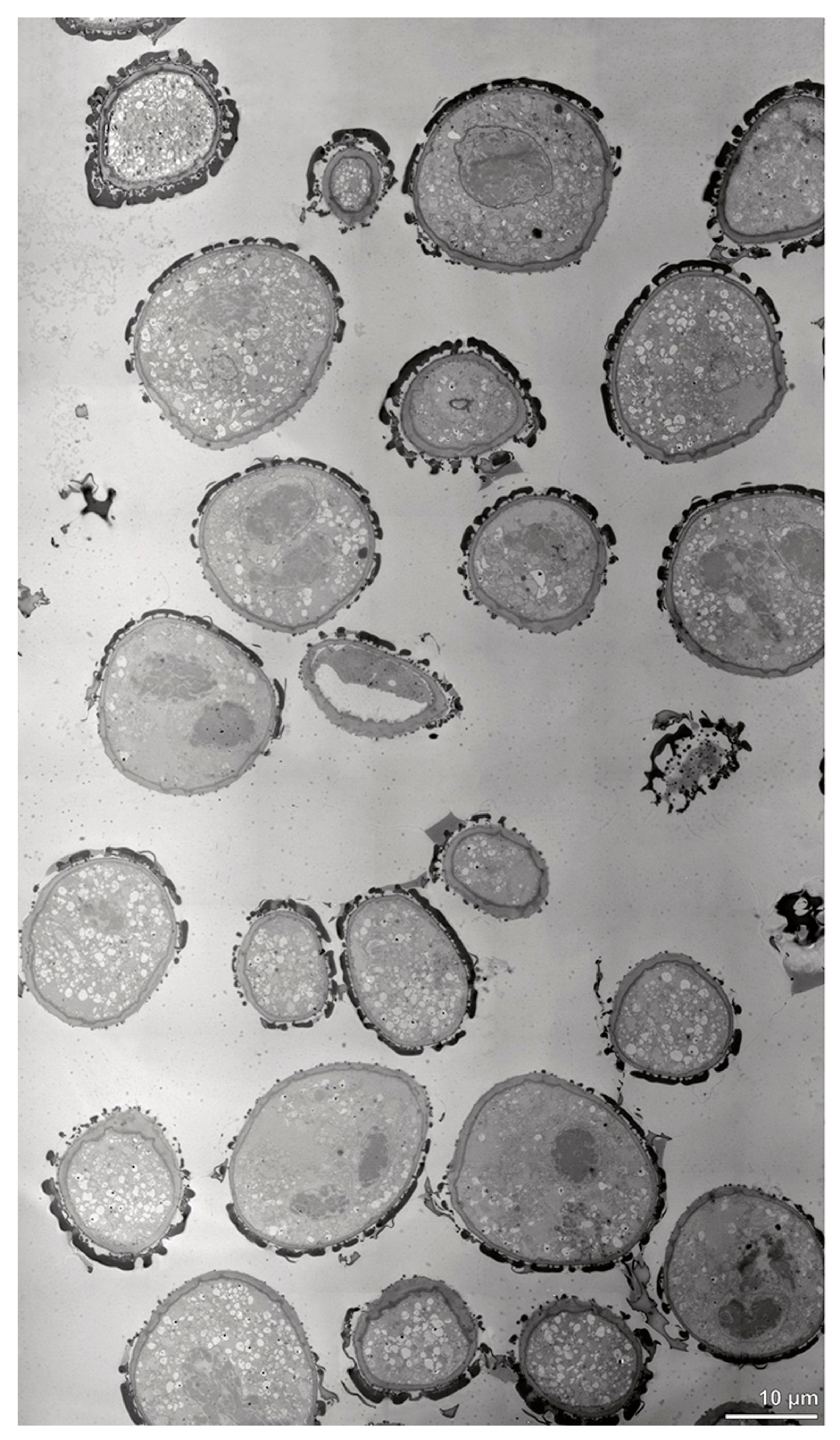
Figure 22.
LM (A,B) and TEM (C–H) micrographs showing cross-sections of Oncidium crocidipterum pollinium/tetrads. Note: (A) Semi-thin section of pollinium; black star marks intrasutural area, TBO. (B) Semi-thin section showing different strata (St 1, St 2, St 3) based on pollen wall differentiation, TBO. (C) Tetrad in intersutural area of outer stratum 1, showing detached protoplasts and a three- (D) to two-layered (E) pollen wall. (D) Detail of pollen wall with three layers (L1, L2, L3). (E) Detail of pollen wall consisting of layer 2 (black arrowhead) with embedded granules (black arrow) and layer 3 (intine, white arrowhead), as well as a thin electron-dense layer (white arrow). (F) Tetrad in intrasutural area of outer stratum 1, showing detached protoplasts and a three- (G) to two-layered (H) pollen wall. (G) Detail of pollen wall with three layers (L1, L2, L3). (H) Detail of pollen wall consisting of layer 2 (black arrowhead) with granules (black arrow) and layer 3 (intine, white arrowhead).
Figure 22.
LM (A,B) and TEM (C–H) micrographs showing cross-sections of Oncidium crocidipterum pollinium/tetrads. Note: (A) Semi-thin section of pollinium; black star marks intrasutural area, TBO. (B) Semi-thin section showing different strata (St 1, St 2, St 3) based on pollen wall differentiation, TBO. (C) Tetrad in intersutural area of outer stratum 1, showing detached protoplasts and a three- (D) to two-layered (E) pollen wall. (D) Detail of pollen wall with three layers (L1, L2, L3). (E) Detail of pollen wall consisting of layer 2 (black arrowhead) with embedded granules (black arrow) and layer 3 (intine, white arrowhead), as well as a thin electron-dense layer (white arrow). (F) Tetrad in intrasutural area of outer stratum 1, showing detached protoplasts and a three- (G) to two-layered (H) pollen wall. (G) Detail of pollen wall with three layers (L1, L2, L3). (H) Detail of pollen wall consisting of layer 2 (black arrowhead) with granules (black arrow) and layer 3 (intine, white arrowhead).
![Plants 13 01114 g022]()
Figure 23.
TEM micrographs showing cross-sections of Oncidium crocidipterum pollinium/tetrads. Note: (A) Tetrad of stratum 2, showing a two-layered pollen wall. (B) Detail of pollen wall, showing layer 2 (black arrowhead) with granules (black arrow) and layer 3 (intine, white arrowhead), along with a thin electron-dense layer (white arrow). (C) Tetrad of stratum 3, showing a two-layered pollen wall (D) and thin electron-dense layer (white arrow). (D) Detail of a pollen wall consisting of layer 2 (black arrowhead) with few granules (black arrow), layer 3 (intine, white arrowhead), and a thin electron-dense layer (white arrow). (E) Pollen wall of a tetrad from the outer stratum 1, showing an electron-translucent layer 2 (L2) with electron-dense granules, KMnO4. (F) Outer pollen wall of a tetrad from stratum 3; black arrowhead points to layer 2, KMnO4.
Figure 23.
TEM micrographs showing cross-sections of Oncidium crocidipterum pollinium/tetrads. Note: (A) Tetrad of stratum 2, showing a two-layered pollen wall. (B) Detail of pollen wall, showing layer 2 (black arrowhead) with granules (black arrow) and layer 3 (intine, white arrowhead), along with a thin electron-dense layer (white arrow). (C) Tetrad of stratum 3, showing a two-layered pollen wall (D) and thin electron-dense layer (white arrow). (D) Detail of a pollen wall consisting of layer 2 (black arrowhead) with few granules (black arrow), layer 3 (intine, white arrowhead), and a thin electron-dense layer (white arrow). (E) Pollen wall of a tetrad from the outer stratum 1, showing an electron-translucent layer 2 (L2) with electron-dense granules, KMnO4. (F) Outer pollen wall of a tetrad from stratum 3; black arrowhead points to layer 2, KMnO4.
Figure 24.
TEM micrographs showing cross-sections of Oncidium crocidipterum pollinium/tetrads. Note: (A) Pollen coatings (white star) in outermost stratum 1 in intrasutural area. (B) Pollen coatings between inner tetrads (white star).
Figure 24.
TEM micrographs showing cross-sections of Oncidium crocidipterum pollinium/tetrads. Note: (A) Pollen coatings (white star) in outermost stratum 1 in intrasutural area. (B) Pollen coatings between inner tetrads (white star).
Description of species
Polystachya cultriformis (Thouars) Lindl. ex Spreng. (
Figure 25,
Figure 26,
Figure 27,
Figure 28,
Figure 29,
Figure 30 and
Figure 31;
Table 1,
Table 2 and
Table 3), higher epidendroids: The pollen mass of the whole anther is arranged in four cup-shaped pollinia, with an average length of 0.4 mm and an average height of 0.62 mm under LM (
Figure 25B and
Figure 26A,
Table 1). The pollinia are connected by a sticky material at the base, known as a frenicular caudicle (
Figure 27B and
Figure 29A), which attaches to a short stipe (
Figure 25B and
Figure 26D,E). The stipe is 0.33 mm long (LM), exhibits a cellular structure (
Figure 26F), and, unlike the pollinium, does not show autofluorescence under LM (
Figure 25C). The combination of the pollinia with these appendages is called the pollinarium, which is irregular-shaped and measures 0.92 mm under LM (
Figure 26A and
Figure 27A). Each pollinium consists of tetrads (
Figure 26B) of various types agglutinated by pollen coatings (elastoviscin) (
Figure 25I and
Figure 26C). The observed tetrad types are planar-tetragonal, planar-rhomboid, tetrahedral, and decussate (
Figure 25D). The size of the hydrated pollen grains within a tetrad in LM ranges between 14.8 µm and 18.2 µm (
Table 1). At anthesis, mature pollen grains are inaperturate and predominantly two-celled (
Figure 25G and
Figure 31C). Pollen reserves, like starch, are absent (
Figure 25H,I); only some lipid droplets have been detected in TEM (
Figure 31C). Toluidine staining of the tetrads in LM revealed differentiation in the pollen wall structure depending on the tetrad’s position within the pollinium, with the outermost tetrads composed of thick wall elements (
Figure 25E).
In SEM, the outer pollen wall layer is psilate (
Figure 26C). Compared to the outer tetrads of the pollinium, the inner tetrads have only thin and unstructured pollen walls (
Figure 25F).
Figure 25.
Macroscopic image of flowers (A) and LM micrographs of Polystachya cultriformis pollen (B–I). (B) Pollinarium consisting of 4 pollinia (po) and a stipe (st). (C) Pollinarium excited with UV light. (D) Various tetrad types. (E) Outer tetrad (planar-rhomboid); exine stained dark blue (black arrowhead), TBO. (F) Inner thin-walled (planar-tetragonal) tetrad, TBO. (G) Binucleate pollen with a generative cell (white arrowhead), Carmine. (H,I) No starch detected; pollen coatings present between tetrads (black arrowhead), Lugol.
Figure 25.
Macroscopic image of flowers (A) and LM micrographs of Polystachya cultriformis pollen (B–I). (B) Pollinarium consisting of 4 pollinia (po) and a stipe (st). (C) Pollinarium excited with UV light. (D) Various tetrad types. (E) Outer tetrad (planar-rhomboid); exine stained dark blue (black arrowhead), TBO. (F) Inner thin-walled (planar-tetragonal) tetrad, TBO. (G) Binucleate pollen with a generative cell (white arrowhead), Carmine. (H,I) No starch detected; pollen coatings present between tetrads (black arrowhead), Lugol.
Figure 26.
SEM micrographs of Polystachya cultriformis pollinarium/pollen. Note: (A) Overview of a hydrated (DMP + CPD) pollinarium comprising 4 pollinia (po) and a stipe (st). (B) Arrangement of tetrads. (C) Tetrads with psilate ornamentation, connected by pollen coatings (white arrowhead). (D) Detail of stipe surface (white star). (E) Detail of the inner part of the stipe. (F) Stipe structure with pollen tetrads attached (white arrowhead).
Figure 26.
SEM micrographs of Polystachya cultriformis pollinarium/pollen. Note: (A) Overview of a hydrated (DMP + CPD) pollinarium comprising 4 pollinia (po) and a stipe (st). (B) Arrangement of tetrads. (C) Tetrads with psilate ornamentation, connected by pollen coatings (white arrowhead). (D) Detail of stipe surface (white star). (E) Detail of the inner part of the stipe. (F) Stipe structure with pollen tetrads attached (white arrowhead).
Figure 27.
SEM micrographs of Polystachya cultriformis pollinarium/pollen. Note: (A) Dorsal view of a hydrated (DMP + CPD) pollinarium. (B) Frenicular caudicle (white stars). (C) Anther cap. (D) No Ubisch bodies detected inside the anther cap.
Figure 27.
SEM micrographs of Polystachya cultriformis pollinarium/pollen. Note: (A) Dorsal view of a hydrated (DMP + CPD) pollinarium. (B) Frenicular caudicle (white stars). (C) Anther cap. (D) No Ubisch bodies detected inside the anther cap.
In TEM, this variation in the pollen wall structure of the tetrads is also clearly visible, allowing the pollinium to be subdivided into two distinct strata: stratum 1, with outermost thick-walled tetrads, and stratum 2, with thin-walled inner tetrads (
Figure 28 and
Figure 29B). The outer tetrads in stratum 1 are characterised by a thick, continuous layer 1 on the distal half of the outer pollen grains (
Figure 29C). This layer decreases in thickness inwardly, finally transitioning into granules surrounding the distal half of the inner pollen grains of the outer tetrads (
Figure 29C,E). The subjacent electron-translucent pollen wall layer (layer 2) contains electron-dense granules of a consistent distribution only on the distal half of the outer pollen grains (
Figure 29D). These granules, exhibiting a contrast similar to that of the ektexine, are thought to consist of sporopollenin. Wall layer 2 surrounds each tetrad, like an ektexine in calymmate tetrads, and also forms the outer wall layer of each pollen grain (monad) (
Figure 29C). Staining with potassium permanganate (
Figure 30A,B) and the Thiéry test (
Figure 30C,D) did not clarify the chemical composition of this layer. Layer 3 constitutes a bilayered intine, composed of an inner electron-dense and an outer electron-translucent polysaccharidic layer (
Figure 30C,D). Layer 3 varies in thickness within the tetrad and is slightly thicker in the distal half (
Figure 29C). In contrast to the outer tetrads of stratum 1, the inner tetrads of stratum 2 have reduced pollen walls composed of pollen wall layer 2 and layer 3 (
Figure 29F). The electron-translucent layer 2 varies in thickness and shows a peripherally diffuse delimitation (
Figure 29G). Like in the outer tetrads of stratum 1, this layer surrounds each tetrad and constitutes the outer wall of the pollen grains (
Figure 29F). The subjacent bilayered intine (layer 3) is thicker on the distal half of the pollen grains (
Figure 29F,G). With the Thiéry test, this layer stains electron-dense, indicating its polysaccharidic nature (
Figure 30E,F). The pollen grains are still in a developing stage (mitosis 1), as seen in TEM, where the formation of the generative cell is still visible in most pollen grains (
Figure 29F,H and
Figure 31A,B). The generative cell/nucleus is formed on the distal half of the pollen grains and is enclosed by an inner, more electron-dense intine layer with an undulating structure (
Figure 29F,H). Even within a single tetrad (of both strata), the pollen grains show different developmental stages, ranging from young microspores with nuclei, lipid droplets, and small vacuoles at the middle stage (
Figure 31A) to microspores at the late stage (mitosis 1) (
Figure 31B), and up to mature two-celled pollen grains with a vegetative nucleus and detached generative cell (
Figure 31C). The pollen grains are poor in organelles, with only a few lipid droplets present (
Figure 31C). In the pollen grains of the outer tetrads of stratum 1, the protoplast is sometimes slightly detached from the pollen wall (
Figure 29C). Pollen coatings are present and attached to the ektexine of the outer tetrads (stratum 1). The coatings differ in their structure, ranging from homogenous to porous (
Figure 31D–F). Ubisch bodies on the inner surface of the anther cap have not been detected in SEM (
Figure 27C,D).
Remarks: Pollen studies on this genus are sparse. To date, only ultrastructural investigations of
Polystachya concreta (Jacq.) Garay & H.R.Sweet, by Wolter and Schill [
29], and
P. pubescens, by Schlag and Hesse [
40], have been conducted. Both studies describe that the sporopollenin elements are limited to the peripheral zones of the pollinia, which also applies to
P. cultriformis. While Schlag and Hesse [
40] depict a bilayered intine for
P. pubescens, Wolter and Schill [
29] delineate a three-layered intine for
P. concreta. For
P. cultriformis we assume a three-layered intine as well. The pollen wall structure of
P. cultriformis deviates from the “typical” angiosperm pollen wall model (see [
10]) and exhibits a reduction in wall structure from the tetrads of stratum 1 (sporopollenin wall elements) to the tetrads of stratum 2 (only granules). Protoplast detachment is infrequent and limited to the tetrads of stratum 1. Furthermore, the sticky materials observed between tetrads and along the peripheral margin of the pollinium differ in structure.
Figure 28.
TEM panorama micrograph showing a cross-section of a Polystachya cultriformis pollinium. Note: The pollinia are divided into three strata based on the variations in their tetrad/pollen wall structure. The peripheral stratum 1 is composed of thick-walled tetrads and the inner stratum 2 of thin-walled tetrads.
Figure 28.
TEM panorama micrograph showing a cross-section of a Polystachya cultriformis pollinium. Note: The pollinia are divided into three strata based on the variations in their tetrad/pollen wall structure. The peripheral stratum 1 is composed of thick-walled tetrads and the inner stratum 2 of thin-walled tetrads.
Figure 29.
LM (A,B) and TEM (C–H) micrographs showing cross-sections of Polystachya cultriformis pollinia/tetrads. Note: (A) Semi-thin section of two pollinia connected by a sticky material (black arrowhead), TBO. (B) Semi-thin section showing different strata (St 1, St 2), TBO. (C) Tetrad of outer stratum 1 with slightly detached protoplasts and a three- to two-layered pollen wall. (D) Detail of pollen wall with three layers (L1, L2, L3). (E) Detail of pollen wall consisting of layer 2 (black arrow), with embedded granules, and layer 3 (bilayered intine, white and black arrowhead). (F) Tetrad of stratum 2. (G) Detail of pollen wall with layer 2 (black arrow) and innermost layer 3 (bilayered intine, white and black arrowheads). (H) Detail of pollen wall layer 3 with inner intine layer (electron-dense) passing into an undulating area on the distal half of the pollen grain (black arrowhead).
Figure 29.
LM (A,B) and TEM (C–H) micrographs showing cross-sections of Polystachya cultriformis pollinia/tetrads. Note: (A) Semi-thin section of two pollinia connected by a sticky material (black arrowhead), TBO. (B) Semi-thin section showing different strata (St 1, St 2), TBO. (C) Tetrad of outer stratum 1 with slightly detached protoplasts and a three- to two-layered pollen wall. (D) Detail of pollen wall with three layers (L1, L2, L3). (E) Detail of pollen wall consisting of layer 2 (black arrow), with embedded granules, and layer 3 (bilayered intine, white and black arrowhead). (F) Tetrad of stratum 2. (G) Detail of pollen wall with layer 2 (black arrow) and innermost layer 3 (bilayered intine, white and black arrowheads). (H) Detail of pollen wall layer 3 with inner intine layer (electron-dense) passing into an undulating area on the distal half of the pollen grain (black arrowhead).
Figure 30.
TEM micrographs showing cross-sections of Polystachya cultriformis pollinium/tetrads. Note: (A) Detail of the pollen wall of a tetrad from the outermost stratum 1, with electron-translucent L2, KMnO4. (B) Detail of the outer pollen wall of a tetrad from the inner stratum 2; white arrowhead points to L2, KMnO4. (C–F) Electron-dense polysaccharides, Thiéry test. (C,D) Pollen wall of a tetrad from the outer stratum 1 with an electron-dense intine (L3) and a Thiéry test control (D) with an electron-translucent intine (L3). (E) Outer pollen wall of a tetrad from stratum 2 with an electron-dense intine (L3) and Thiéry test control (F) with an electron-translucent intine (L3).
Figure 30.
TEM micrographs showing cross-sections of Polystachya cultriformis pollinium/tetrads. Note: (A) Detail of the pollen wall of a tetrad from the outermost stratum 1, with electron-translucent L2, KMnO4. (B) Detail of the outer pollen wall of a tetrad from the inner stratum 2; white arrowhead points to L2, KMnO4. (C–F) Electron-dense polysaccharides, Thiéry test. (C,D) Pollen wall of a tetrad from the outer stratum 1 with an electron-dense intine (L3) and a Thiéry test control (D) with an electron-translucent intine (L3). (E) Outer pollen wall of a tetrad from stratum 2 with an electron-dense intine (L3) and Thiéry test control (F) with an electron-translucent intine (L3).
Figure 31.
TEM micrographs showing cross-sections of Polystachya cultriformis pollinium/tetrads. Note: (A) Microspore stage with a vegetative nucleus (black star) and small vacuoles (v). (B) Young pollen grain with a vegetative nucleus (black star) close to the pollen wall and the formation of a generative cell on the distal half of the pollen grain (black arrow). (C) Mature two-celled pollen grain with a vegetative nucleus (white star), generative cell (black star), and lipid droplets (l). (D) Pollen coatings (black arrowhead) attached to the tetrads of outer stratum 1. (E) Homogenous pollen coatings (black arrowhead) between adjacent tetrads of outer stratum 1. (F) Porous pollen coatings (white star) attached to the ektexine of the tetrads of stratum 1.
Figure 31.
TEM micrographs showing cross-sections of Polystachya cultriformis pollinium/tetrads. Note: (A) Microspore stage with a vegetative nucleus (black star) and small vacuoles (v). (B) Young pollen grain with a vegetative nucleus (black star) close to the pollen wall and the formation of a generative cell on the distal half of the pollen grain (black arrow). (C) Mature two-celled pollen grain with a vegetative nucleus (white star), generative cell (black star), and lipid droplets (l). (D) Pollen coatings (black arrowhead) attached to the tetrads of outer stratum 1. (E) Homogenous pollen coatings (black arrowhead) between adjacent tetrads of outer stratum 1. (F) Porous pollen coatings (white star) attached to the ektexine of the tetrads of stratum 1.



































{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}