


Last-Century Forest Dynamics in a Highland Pyrenean National Park and Implications for Conservation

Abstract
:1. Introduction
2. Study Area
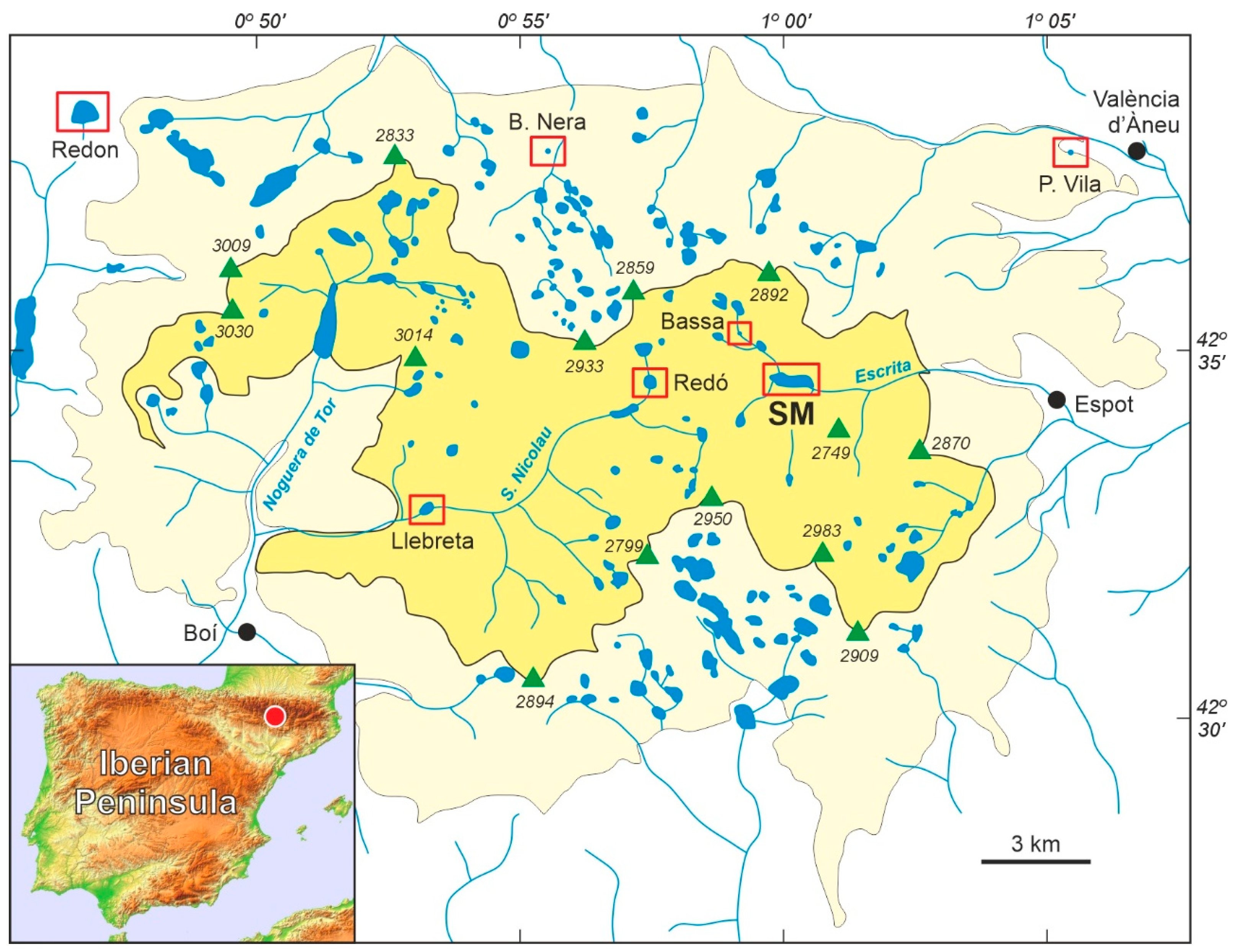
2.1. General Description
2.2. Previous Studies
3. Methods
3.1. Coring and Dating
3.2. Pollen Analysis
4. Results and Interpretation
4.1. Chronology and Sedimentation
4.2. Vegetation Dynamics
5. Discussion and Conclusions
5.1. Summary of Main Trends
5.2. Comparisons with Other Localities and Proxies
5.3. Conservation Insights
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Willis, K.J.; Birks, H.J.B. What is natural? The need for a long-term perspective in biodiversity conservation. Science 2006, 314, 1261–1265. [Google Scholar] [CrossRef] [PubMed]
- Willis, K.J.; Araújo, M.B.; Bennett, K.D.; Figueroa-Rangel, B.; Froyd, C.A.; Myers, N. How can a knowledge of the past help to conserve the future? Biodiversity conservation and the relevance of long-term ecological studies. Philos. Trans. R. Soc. B 2007, 362, 175–187. [Google Scholar] [CrossRef] [PubMed]
- Willis, K.J.; Bailey, R.M.; Bhagwat, S.A.; Birks, H.J.B. Biodiversity baselines, thresholds and resilience: Testing predictions and assumptions using paleoecological data. Trends Ecol. Evol. 2010, 25, 583–591. [Google Scholar] [CrossRef] [PubMed]
- Davies, A.L.; Colombo, S.; Nick, H. Improving the application of long-term ecology in conservation and land management. J. Appl. Ecol. 2014, 51, 63–70. [Google Scholar] [CrossRef]
- Seddon, A.W.R.; Mackay, A.W.; Baker, A.G.; Birks, H.J.B.; Breman, E.; Buck, C.; Ellis, E.C.; Froyd, C.A.; Gill, J.F.; Gillson, L.; et al. Looking forward through the past: Identification of 50 priority research questions in paleoecology. J. Ecol. 2014, 102, 256–267. [Google Scholar] [CrossRef]
- Rull, V.; Vegas-Vilarrúbia, T.; Corella, J.P.; Trapote, M.C.; Montoya, E.; Valero-Garcés, B. A unique Pyrenean varved record provides a detailed reconstruction of Mediterranean vegetation and land-use dynamics over the last three millennia. Quat. Sci. Rev. 2021, 268, 107128. [Google Scholar] [CrossRef]
- Gil-Farrero, J. Un segle de canvis en la gestió dels recursos naturals al Parc Nacional d’Aigüestortes i Estany de Sant Maurici. In Proceedings of the XII Jornades Sobre Recerca al Parc Nacional d’Aigüestortes i Estany de Sant Maurici, Espot, Spain, 13–15 October 2021; pp. 207–216. [Google Scholar]
- Gil-Farrero, J. Conservación, divulgación e imagen pública de la naturaleza durante el franquismo; El Parque Nacional de Aigüestortes i Estany de Sant Maurici. Rubrica Contemp. 2022, 11, 249. [Google Scholar] [CrossRef]
- Catalan, J. L’Obra Hidràulica en les Pirineus. In Avaluació, Correcció i Prevenció de l’Impacte Mediambiental; ENHER-Fundació La Caixa-FECSA: Barcelona, Spain, 1997. [Google Scholar]
- Rull, V.; Cañellas-Boltà, N.; Vegas-Vilarrúbia, T. Late Holocene forest resilience in the central Pyrenean highlands as deduced from pollen analysis of Lake Sant Maurici sediments. Holocene 2021, 31, 1797–1803. [Google Scholar] [CrossRef]
- Carrillo, E.; Aniz, M. Guía del Parque Nacional de Aigüestortes i Estany de Sant Maurici; Organismo Autónomo de Parques Nacionales-Generalitat de Catalunya: Madrid, Spain, 2013. [Google Scholar]
- Rodríguez-Fernández, R. Parque Nacional de Aigüestortes i Estany de Sant Maurici; Geológica, G., Ed.; Everest: León, Spain, 2010. [Google Scholar]
- Sigro, J.; Pérez-Luque, A.J.; Pérez-Martínez, C.; Vegas-Vilarrubia, T.; Esteban-Parra, M.J. Trends in Temperature and Precipitation in High Mountain Areas in Spain from the Spanish Hig Mountain Climate Database; EGU General Assembly: Vienna, Austria, 2022; p. EGU22-4257. [Google Scholar]
- Vigo, J. L’alta muntanya catalana. Flora i vegetació; Centre Excursionista de Catalunya-Institut d’Estudis Catalans: Barcelona, Spain, 2008. [Google Scholar]
- Calero, M.A.; Valero-Garcés, B.; Rull, V.; Vegas-Vilarrúbia, T.; Garcés-Pastor, S.; López-Vila, J.; Camarero, J.J. El registro sedimentario del lago Sant Maurici (Pirineos Centrales). Geogaceta 2016, 59, 11–14. [Google Scholar]
- Rull, V. Anticipation, discovery and serendipity in Quaternary paleoecology: Personal experiences from the Iberian Pyrenees. Quaternary 2023, 6, 42. [Google Scholar] [CrossRef]
- Esteban, A. La humanización de las altas cuencas de la Garona y las Nogueras (4500 aC—1955 dC); Ministerio de Medio Ambiente: Madrid, Spain, 2023. [Google Scholar]
- Catalan, J.; Pelachs, A.; Gassiot, E.; Antolín, F.; Ballesteros, A.; Batalla, M.; Burjachs, F.; Buchaca, T.; Camarero, L.; Clemente-Conte, I.; et al. Interaccion entre clima y ocupacion humana en la configuracion del paisaje vegetal del Parque Nacional de Aiguestortes i Estany de Sant Maurici a lo largo de los ultimos 15.000 anos. In Proyectos de Investigación en Parques Nacionales: 2009–2012; Ramirez, L., Asensio, B., Eds.; Organismo Autonomo de Parques Nacionales: Madrid, Spain, 2013; pp. 71–92. [Google Scholar]
- Rodríguez, J.-M.; Pérez-Obiol, R.; Pérez- Haase, A.; Nadal-Tersa, J.; Sánchez-Morales, M.; Cunill-Artigas, R. Recerca paleoambiental a l’Estany de la Bassa. In Proceedings of the XII Jornades de Recerca al Parc Nacional d’Aigüestortes i Estany de Sant Maurici, Espot, Spain, 13–15 October 2021; pp. 153–162. [Google Scholar]
- Rull, V.; Vegas-Vilarrúbia, T. Conifer forest dynamics in the Iberian Pyrenees during the Middle Ages. Forests 2021, 12, 1685. [Google Scholar] [CrossRef]
- Sánchez-Cabeza, J.A.; Masqué, P.; Ani-Ragolta, I. 210Pb and 210Po analysis in sediments and soils by microwave acid digestion. J. Radioanal. Nucl Chem. 1998, 227, 19–22. [Google Scholar] [CrossRef]
- Appleby, P.G.; Oldfield, F. The assessment of 210Pb data from sites with varying sediment accumulation rates. Hydrobiologia 1983, 103, 29–35. [Google Scholar] [CrossRef]
- Bennett, K.D.; Willis, K.D. Pollen. In Tracking Environmental Change Using Lake Sediments. Volume 3: Terrestrial, Algal and Siliceous Indicators; Smol, J.P., Birks, H.J.B., Last, W.M., Eds.; Kluwer: New York, NY, USA, 2001; pp. 5–32. [Google Scholar]
- Stockmarr, J. Tablets with spores used in absolute pollen analysis. Pollen Spores 1971, 13, 615–621. [Google Scholar]
- Moore, P.D.; Webb, J.A.; Collinson, M.E. Pollen Analysis; Blackwell: Oxford, UK, 1991. [Google Scholar]
- Reille, M. Pollen et Spores d’Europe et d’Afrique du Nord; URA-CNRS: Marseille, France, 1992. [Google Scholar]
- López-Vila, J.; Montoya, E.; Cañellas-Boltà, N.; Rull, V. Modern non-pollen palynomorphs sedimentation along an elevational transect in the south-central Pyrenees (southwestern Europe) as a tool for Holocene paleoecological reconstruction. Holocene 2014, 24, 327–345. [Google Scholar] [CrossRef]
- Cugny, C.; Mazier, F.; Galop, D. Modern and fossil non-pollen palynomorphs from the Basque mountains (western Pyrenees, France): The use of coprophilous fungi to reconstruct pastoral activity. Veget. Hist. Archaeobot. 2020, 19, 391–408. [Google Scholar] [CrossRef]
- Baker, A.G.; Bhagwat, S.A.; Willis, K.J. Do dung fungal spores make a good proxy for past distribution of large herbivores? Quat. Sci. Rev. 2013, 62, 21–31. [Google Scholar] [CrossRef]
- Gauthier, E.; Jouffroy-Bapicot, I. Detecting human impacts: Non-pollen palynomorphs as proxies for human impact on the environment. In Applications of Non-Pollen Palynomorphs: From Palaeoenvironmental Reconstruction to Biostratigraphy; Marret, F., O’Keefe, J., Osterloff, P., Pound, M., Shumilovkikh, L., Eds.; Geological Society Special Publications 511: London, UK, 2021; pp. 233–244. [Google Scholar]
- Lee, C.M.; van Geel, B.; Gosling, W.D. On the use of spores of coprophilous fungi preserved in sediments to indicate past herbivore presence. Quaternary 2022, 5, 30. [Google Scholar] [CrossRef]
- Whitlock, C.; Larsen, C. Charcoal as a fire proxy. In Tracking Environmental Change Using Lake Sediments. Volume 3: Terrestrial, Algal and Siliceous Indicators; Smol, J.P., Birks, H.J.B., Last, W.L., Eds.; Kluwer: New York, NY, USA, 2001; pp. 75–97. [Google Scholar]
- Rull, V. A note on pollen counting in palaeoecology. Pollen Spores 1987, 29, 471–480. [Google Scholar]
- Theuerkauf, M.; Couwenberg, J. ROPES reveals past land cover and PPs from single pollen records. Front. Earth Sci. 2018, 6, 14. [Google Scholar] [CrossRef]
- Dubois, H.; Verkasalo, E.; Claessens, H. Potential of birch (Betula pendula Roth and B. pubescescens Ehrh.) for forestry and forest-based industry sector with the changing climatic and socio-economic context of western Europe. Forests 2020, 11, 336. [Google Scholar] [CrossRef]
- Wu, H.X.; Schoch, C.L.; Boonmmee, S.; Bahkali, A.H.; Chomnuti, P.; Hyde, K.D. A reappraisal of Microthyriaceae. Fungal Divers 2011, 1, 189–248. [Google Scholar] [CrossRef] [PubMed]
- García Álvarez, S.; Morla Juaristi, C.; Solana Gutiérrez, J.; García-Amorena, I. Taxonomic differences between Pinus sylvestris and P. uncinata revealed in the stomata and cutilce characters for use in the study of fossil material. Rev. Palaeobot. Palynol. 2009, 155, 61–68. [Google Scholar] [CrossRef]
- Carrión, J.S.; Munuera, M.; Ochando, J.; López-Sáez, J.A.; Casas-Gallego, M.; González-Sampériz, P.; Badal, E.; Pérez-Díaz, S.; Carrión-Marco, I.; Jiménez-Moreno, G.; et al. Paleofolora y Paleovegetación Ibérica III: Holoceno; Ministerio de Ciencia e Innovación-Fundación Séneca: Madrid, Spain, 2022. [Google Scholar]
- Mercadé, A.; Vigo, J.; Rull, V.; Vegas-Vilarrúbia, T.; Garcés, S.; Lara, A.; Cañellas-Boltà, N. Vegetation and landscape around Lake Montcortès (Catalan pre-Pyrenees) as a tool for palaeoecological studies of lake sediments. Collect. Bot. 2013, 32, 87–101. [Google Scholar] [CrossRef]
- Rull, V.; Vegas-Vilarrúbia, T. A recent Cannabis pollen increase on the Iberian Pyrenees. Sci. Total Environ. 2023, 886, 163947. [Google Scholar] [CrossRef] [PubMed]
- Büntgen, U.; Krusic, P.J.; Verstege, A.; Sangüesa-Barreda, G.; Wagner, S.; Camarero, J.J.; Charpentier, F.; Zorita, E.; Oppenheimer, C.; Koner, O.; et al. New tree-ring evidence from the Pyrenees reveals western Mediterranean climate variability since Medieval times. J. Clim. 2017, 30, 5295–5318. [Google Scholar] [CrossRef]
- Muntán, E.; García, C.; Oller, P.; Martí, G.; García, A.; Gutiérrez, E. Reconstructing snow avalanches in the Southeastern Pyrenees. Nat. Haz. Earth Syst. Sci. 2009, 9, 1599–1612. [Google Scholar] [CrossRef]
- Hevia, A.; Sánchez-Salguero, R.; Camarero, J.J.; Buras, A.; Sangüesa-Barreda, G.; Galván, J.D.; Gutiérrez, E. Towards a better understanding of long-term wood-chemistry variations in old-growth forests: A case study on ancient Pinus uncinata trees from the Pyrenees. Sci. Total Environ. 2018, 625, 220–232. [Google Scholar] [CrossRef] [PubMed]
- Galván, J.D.; Camarero, J.J.; Gutiérrez, E. Seeing the trees for the forest: Drivers of individual growth responses to climate in Pinus uncinata mountain forests. J. Ecol. 2014, 102, 1244–1257. [Google Scholar] [CrossRef]
- Sanmiguel-Vallelado, A.; Camarero, J.J.; Gazol, A.; Morán-Tejeda, E.; Sangüesa-Barreda, M.; Alonso-González, E.; Gutiérrez, E.; Alla, A.Q.; Diego, J.; López-Moreno, J.I. Detecting snow-related signals in radial growth of Pinus uncinata mountain forests. Dendrochronologia 2019, 57, 125622. [Google Scholar] [CrossRef]
- Marqués, L.; Ogle, K.; Peltier, D.M.P.; Camarero, J.J. Altered climate memory characterizes tree growth during forest dieback. Agric. For. Meteorol. 2022, 314, 108787. [Google Scholar] [CrossRef]
- Valeriano, C.; Tumajer, J.; Gazol, A.; González, E.; Sánches-Salguero, R.; Colangelo, M.; Linares, J.C.; Valor, T.; Sangüesa-Barreda, G.; Camarero, J.J. Delineating vulnerability to drought using a process-based growth model in Pyrenean silver fir forests. For. Ecol. Manag. 2023, 541, 121069. [Google Scholar] [CrossRef]
- Anderegg, W.R.L.; Kane, J.K.; Anderegg, L.D.L. Consequences of widespread tree mortality triggered by drought and temperature stress. Nat. Clim. Chang. 2013, 3, 30–36. [Google Scholar] [CrossRef]
- Masson-Delmotte, V.; Zhai, P.; Pirani, A.; Connors, S.L.; Péan, C.; Chen, Y.; Goldfarb, L.; Gomis, M.I.; Matthews, J.B.R.; Berger, S.; et al. Climate Change 2021: The Physical Science Basis; Cambridge University Press: Cambridge, UK, 2021. [Google Scholar]
- Fernàndez-Martínez, J.; Alba Fransi, M.; Fleck, I. Ecolophysiological responses of Betula pendula, Pinus uncinata and Rhododendron ferrugineum in the Catalan Pyrenees to low summer rainfall. Tree Physiol. 2016, 36, 1520–1535. [Google Scholar] [CrossRef]
- García-Ruiz, J.M.; Tomás-Faci, G.; Diarte-Blasco, P.; Montes, L.; Domingo, R.; Sebastián, M.; Lasanta, T.; González-Sampériz, P.; López-Moreno, J.I.; Arnáez, J.; et al. Transhumance and long-term deforestation in the subalpine belt of the Central Spanish Pyrenees: An interdisciplinary approach. Catena 2020, 195, 104744. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Depth (cm) | Excess 210Pb (Bq·kg−1) | Date (year) | SAR (cm·y−1) |
|---|---|---|---|---|
| 1P | 1.0 | 292.5 ± 11.1 | 2007 ± 1 | 0.282 ± 0.013 |
| 2P | 2.0 | 2005 ± 1 | 0.301 ± 0.017 | |
| 3P | 3.1 | 360.4 ± 21.5 | 2002 ± 1 | 0.212 ± 0.015 |
| 4P | 4.1 | 1997 ± 1 | 0.269 ± 0.017 | |
| 5P | 5.1 | 228.4 ± 11.1 | 1993 ± 1 | 0.291 ± 0.017 |
| 6P | 6.1 | 1990 ± 1 | 0.266 ± 0.021 | |
| 7P | 7.1 | 164.9 ± 16.0 | 1986 ± 1 | 0.343 ± 0.037 |
| 8P | 8.1 | 1983 ± 1 | 0.344 ± 0.040 | |
| 9P | 9.2 | 98.3 ± 11.5 | 1980 ± 1 | 0.384 ± 0.049 |
| 10P | 10.2 | 1977 ± 1 | 0.322 ± 0.043 | |
| 11P | 11.2 | 76.4 ± 9.4 | 1974 ± 1 | 0.463 ± 0.062 |
| 12P | 12.2 | 1972 ± 2 | 0.394 ± 0.051 | |
| 13P | 13.2 | 85.8 ± 9.2 | 1969 ± 2 | 0.303 ± 0.037 |
| 14P | 14.3 | 1966 ± 2 | 0.298 ± 0.033 | |
| 15P | 15.3 | 91.1 ± 6.9 | 1963 ± 2 | 0.291 ± 0.029 |
| 16P | 16.3 | 1959 ± 2 | 0.307 ± 0.038 | |
| 17P | 17.3 | 77.3 ± 9.9 | 1956 ± 2 | 0.341 ± 0.052 |
| 18P | 18.3 | 1953 ± 2 | 0.333 ± 0.052 | |
| 19P | 19.9 | 52.5 ± 6.9 | 1950 ± 3 | 0.256 ± 0.043 |
| 20P 1 | 20.4 | 1947 ± 3 | 0.248 ± 0.042 | |
| 21P | 21.4 | 60.5 ± 7.6 | 1944 ± 3 | 0.239 ± 0.041 |
| 22P | 22.4 | 1937 ± 4 | 0.199 ± 0.037 | |
| 23P | 23.4 | 61.8 ± 8.2 | 1932 ± 4 | 0.146 ± 0.030 |
| 24P | 24.4 | 1925 ± 5 | 0.161 ± 0.038 | |
| 25P | 25.5 | 43.5 ± 7.7 | 1919 ± 6 | 0.120 ± 0.035 |
| 1C 2 | 27.0 | 1911 ± 8 | 0.070 ± 0.039 | |
| 2C 2 | 28.0 | 34.8 ± 17.7 | 1889 ± 11 | 0.043 ± 0.037 |
| 3C 2 | 29.0 | 1865 ± 15 | 0.031 ± 0.031 | |
| 4C 2 | 30.0 | 1832 ± 32 | ND | |
| 5C 2 | 31.1 | <0 | ND | ND |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rull, V.; Blasco, A.; Sigro, J.; Vegas-Vilarrúbia, T. Last-Century Forest Dynamics in a Highland Pyrenean National Park and Implications for Conservation. Plants 2024, 13, 1144. https://doi.org/10.3390/plants13081144
Rull V, Blasco A, Sigro J, Vegas-Vilarrúbia T. Last-Century Forest Dynamics in a Highland Pyrenean National Park and Implications for Conservation. Plants. 2024; 13(8):1144. https://doi.org/10.3390/plants13081144
Chicago/Turabian StyleRull, Valentí, Arnau Blasco, Javier Sigro, and Teresa Vegas-Vilarrúbia. 2024. "Last-Century Forest Dynamics in a Highland Pyrenean National Park and Implications for Conservation" Plants 13, no. 8: 1144. https://doi.org/10.3390/plants13081144
APA StyleRull, V., Blasco, A., Sigro, J., & Vegas-Vilarrúbia, T. (2024). Last-Century Forest Dynamics in a Highland Pyrenean National Park and Implications for Conservation. Plants, 13(8), 1144. https://doi.org/10.3390/plants13081144