

Genome-Wide Analysis Elucidates the Roles of GhTIR1/AFB Genes Reveals the Function of Gh_D08G0763 (GhTIR1) in Cold Stress in G. hirsutum

 ,
,
Abstract
:1. Introduction
2. Material and Methods
2.1. Identification of TIR1/AFBs in G. hirsutum and Phylogenetic Tree Construction
2.2. Analysis of Motif and the Exon/Intron Structure of GhTIR1/AFB Genes
2.3. Analysis of Cis-Elements of Upstream Sequences
2.4. Transcriptome Analysis
2.5. Plant Materials and Treatments
2.6. Subcellular Localization Analysis
2.7. Yeast Two-Hybrid Assay
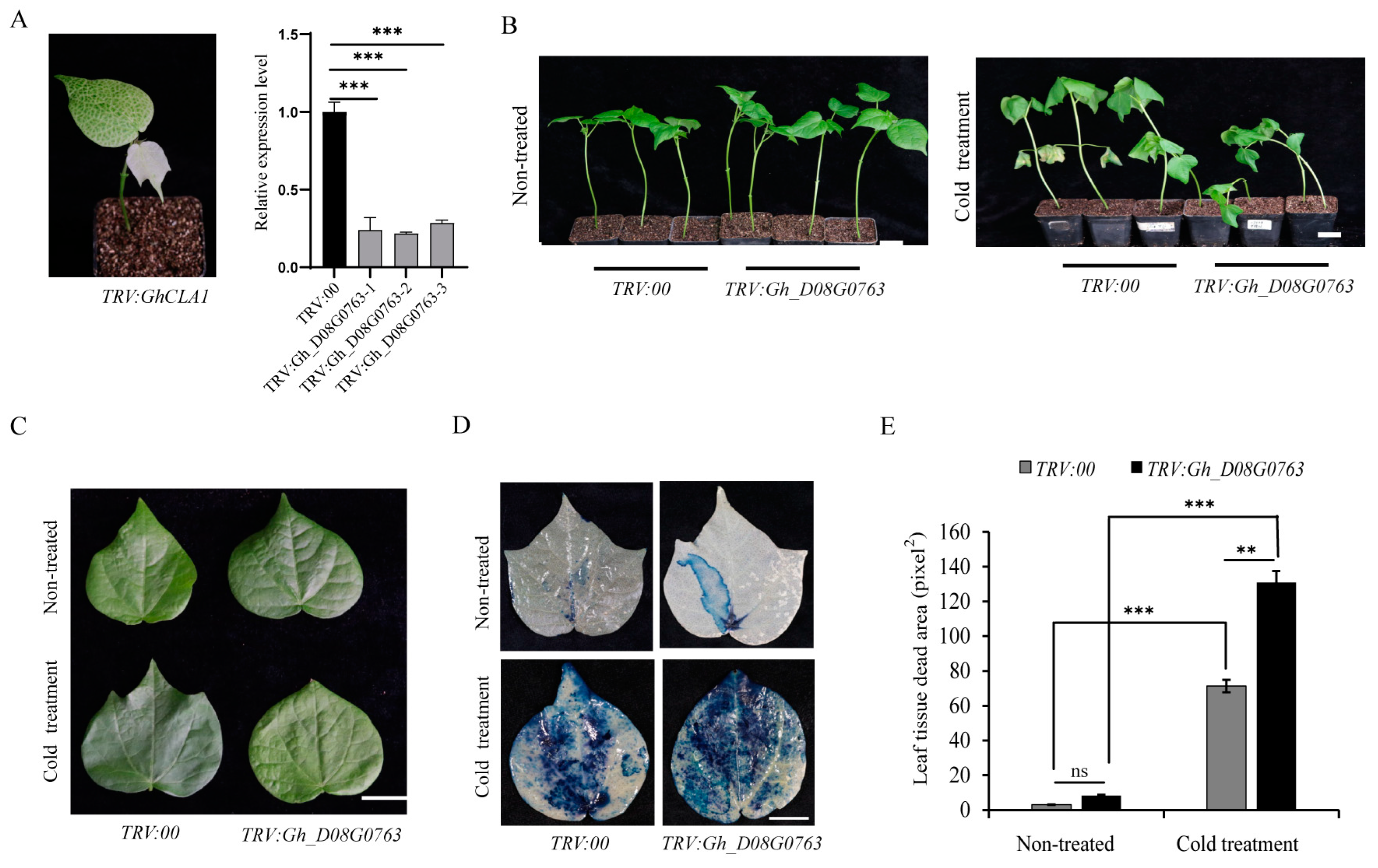
2.8. VIGS for Gene Functional Verification
2.9. Trypan Blue Staining Experiment
2.10. Determination of Enzyme Activity Level of Gene Expression
2.11. RNA Extraction and RT-qPCR Analysis
2.12. Primer Sequence Design
3. Results and Analysis
3.1. Identification and Evolutionary Analysis of TIR1/AFBs in G. hirsutum
3.2. Analysis of Conserved Protein Motifs and Gene Structure of GhTIR1/AFBs
3.3. Analysis of Cis-Acting Elements in the Promoter Regions of GhTIR1/AFBs
3.4. Expression Analysis of GhTIR1/AFBs under Phytohormone and Abiotic Stress
3.5. Subcellular Localization of Gh_D08G076
3.6. Yeast Two-Hybrid Verification of the Interaction between GhTIR1/AFBs and GhSKP1 Protein
3.7. Phenotype Observation of Gh_D08G0763 Knock-Out by VIGS
3.8. Trypan Blue Staining Analysis of VIGS Plant Leaves
3.9. Determination of Enzyme Activity in Plants Silenced by VIGS
4. Discussion
4.1. Bioinformatics Analysis of the GhTIR1/AFB Gene in G. hirsutum
4.2. The GhTIR1/AFB Gene Family Responded Positively to Abiotic Stress
4.3. Possible Regulatory Mechanisms of Gh_D08G0763 in Response to Cold Stress
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Yu, Z.; Zhang, F.; Friml, J.; Ding, Z. Auxin signaling: Research advances over the past 30 years. J. Integr. Plant Biol. 2022, 64, 371–392. [Google Scholar] [CrossRef]
- Xu, Y.; Lou, H.; Zhao, R. Cloning and expression of the catalase gene (KatA) from Pseudomonas aeruginosa and the degradation of AFB1 by recombinant catalase. J. Sci. Food Agric. 2023, 103, 792–798. [Google Scholar] [CrossRef] [PubMed]
- Leyser, O. Auxin Signaling. Plant Physiol. 2018, 176, 465–479. [Google Scholar] [CrossRef]
- Chen, H.; Ma, B.; Zhou, Y.; He, S.J.; Tang, S.Y.; Lu, X.; Xie, Q.; Chen, S.Y.; Zhang, J.S. E3 ubiquitin ligase SOR1 regulates ethylene response in rice root by modulating stability of Aux/IAA protein. Proc. Natl. Acad. Sci. USA 2018, 115, 4513–4518. [Google Scholar] [CrossRef]
- Shu, W.; Liu, Y.; Guo, Y.; Zhou, H.; Zhang, J.; Zhao, S.; Lu, M.A. Populus TIR1 gene family survey reveals differential expression patterns and responses to 1-naphthaleneacetic acid and stress treatments. Front. Plant Sci. 2015, 6, 719. [Google Scholar] [CrossRef]
- Cai, Z.; Zeng, D.E.; Liao, J.; Cheng, C.; Sahito, Z.A.; Xiang, M.; Fu, M.; Chen, Y.; Wang, D. Genome-Wide Analysis of Auxin Receptor Family Genes in Brassica juncea var. tumida. Genes 2019, 10, 165. [Google Scholar] [CrossRef]
- Navarro, L.; Dunoyer, P.; Jay, F.; Arnold, B.; Dharmasiri, N.; Estelle, M.; Voinnet, O.; Jones, J.D. A plant miRNA contributes to antibacterial resistance by repressing auxin signaling. Science 2006, 312, 436–439. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Li, Z.; Xiong, L. A plant microRNA regulates the adaptation of roots to drought stress. FEBS Lett. 2012, 586, 1742–1747. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Li, J.; Cui, L.; Zhang, T.; Wu, Z.; Zhu, P.Y.; Meng, Y.J.; Zhang, K.J.; Yu, X.Q.; Lou, Q.F.; et al. New insights into the roles of cucumber TIR1 homologs and miR393 in regulating fruit/seed set development and leaf morphogenesis. BMC Plant Boil. 2017, 17, 130. [Google Scholar] [CrossRef]
- Morffy, N.; Strader, L.C. Structural Aspects of Auxin Signaling. Cold Spring Harb. Perspect. Biol. 2022, 14, a039883. [Google Scholar] [CrossRef]
- Dharmasiri, N.; Dharmasiri, S.; Jones, A.M.; Estelle, M. Auxin action in a cell-free system. Curr. Biol. 2003, 13, 1418–1422. [Google Scholar] [CrossRef] [PubMed]
- Sevillano, N.; Girón, M.D.; Salido, M.; Vargas, A.M.; Vilches, J.; Salto, R. Internalization of the receptor for advanced glycation end products (RAGE) is required to mediate intracellular responses. J. Biochem. 2009, 145, 21–30. [Google Scholar] [CrossRef]
- Jiang, W.; Jin, Y.M.; Lee, J.; Lee, K.I.; Piao, R.; Han, L.; Shin, J.C.; Jin, R.D.; Cao, T.; Pan, H.Y.; et al. Quantitative trait loci for cold tolerance of rice recombinant inbred lines in low temperature environments. Mol. Cells 2011, 32, 579–587. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Dong, S.; Zhu, H.; Jiang, Q.; Yang, J. Molecular and expression analysis of manganese superoxide dismutase (Mn-SOD) gene under temperature and starvation stress in rotifer Brachionus calyciflorus. Mol. Biol. Rep. 2013, 40, 2927–2937. [Google Scholar] [CrossRef] [PubMed]
- Novillo, F.; Medina, J.; Rodríguez-Franco, M.; Neuhaus, G.; Salinas, J. Genetic analysis reveals a complex regulatory network modulating CBF gene expression and Arabidopsis response to abiotic stress. J. Exp. Bot. 2012, 63, 293–304. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Zhang, Z.; Xie, S.; Si, T.; Li, Y.; Zhu, J.K. Mutational Evidence for the Critical Role of CBF Transcription Factors in Cold Acclimation in Arabidopsis. Plant Physiol. 2016, 171, 2744–2759. [Google Scholar] [CrossRef] [PubMed]
- Birkeland, S.; Slotte, T.; Krag Brysting, A.; Gustafsson, A.L.S.; Rhoden Hvidsten, T.; Brochmann, C.; Nowak, M.D. What can cold-induced transcriptomes of Arctic Brassicaceae tell us about the evolution of cold tolerance? Mol. Ecol. 2022, 31, 4271–4285. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.B.; Li, Y.; Wang, B.; Chee, P.W. Recent advances in cotton genomics. Int. J. Plant Genom. 2008, 2008, 742304. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; He, P.; Ma, X.; Yang, Z.; Pang, C.; Yu, J.; Wang, G.; Friml, J.; Xiao, G. Auxin-mediated statolith production for root gravitropism. New Phytol. 2019, 224, 761–774. [Google Scholar] [CrossRef]
- Perez, L.M.; Mauleon, R.; Arick, M.A.; Magbanua, Z.V.; Peterson, D.G.; Dean, J.F.D.; Tseng, T.M. Transcriptome analysis of the 2,4-dichlorophenoxyacetic acid (2,4-D)-tolerant cotton chromosome substitution line CS-B15sh and its susceptible parental lines G. hirsutum L. cv. Texas Marker-1 and G. barbadense L. cv. Pima 379. Front. Plant Sci. 2022, 13, 910369. [Google Scholar] [CrossRef]
- Li, X.; Liu, G.; Geng, Y.; Wu, M.; Pei, W.; Zhai, H.; Zang, X.; Li, X.; Zhang, J.; Yu, S.; et al. A genome-wide analysis of the small auxin-up RNA (SAUR) gene family in cotton. BMC Genom. 2017, 18, 815. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef] [PubMed]
- Quevillon, E.; Silventoinen, V.; Pillai, S.; Harte, N.; Mulder, N.; Apweiler, R.; Lopez, R. InterProScan: Protein domains identifier. Nucleic Acids Res. 2005, 33, W116–W120. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Jin, J.; Guo, A.Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [PubMed]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Wu, Y.; Li, J.; Wang, X.; Zeng, Z.; Xu, J.; Liu, Y.; Feng, J.; Chen, H.; He, Y.; et al. TBtools-II: A “one for all, all for one” bioinformatics platform for biological big-data mining. Mol. Plant 2023, 16, 1733–1742. [Google Scholar] [CrossRef] [PubMed]
- Sparkes, I.A.; Runions, J.; Kearns, A.; Hawes, C. Rapid, transient expression of fluorescent fusion proteins in tobacco plants and generation of stably transformed plants. Nat. Protoc. 2006, 1, 2019–2025. [Google Scholar] [CrossRef]
- Li, X.; Sun, Y.; Liu, N.; Wang, P.; Pei, Y.; Liu, D.; Ma, X.; Ge, X.; Li, F.; Hou, Y. Enhanced resistance to Verticillium dahliae mediated by an F-box protein GhACIF1 from Gossypium hirsutum. Plant Sci. 2019, 284, 127–134. [Google Scholar] [CrossRef]
- Gao, X.; Britt, R.C., Jr.; Shan, L.; He, P. Agrobacterium-mediated virus-induced gene silencing assay in cotton. J. Vis. Exp. 2011, 54, 2938. [Google Scholar]
- Dong, Y.; Burch-Smith, T.M.; Liu, Y.; Mamillapalli, P.; Dinesh-Kumar, S.P. A ligation-independent cloning tobacco rattle virus vector for high-throughput virus-induced gene silencing identifies roles for NbMADS4-1 and -2 in floral development. Plant Physiol. 2007, 145, 1161–1170. [Google Scholar] [CrossRef]
- Wu, C.; Xiao, S.; Zuo, D.; Cheng, H.; Zhang, Y.; Wang, Q.; Lv, L.; Song, G. Genome-wide analysis elucidates the roles of GhHMA genes in different abiotic stresses and fiber development in upland cotton. Plant Physiol. Biochem. 2023, 194, 281–301. [Google Scholar] [CrossRef] [PubMed]
- Qi, L.; Kwiatkowski, M.; Chen, H.; Hoermayer, L.; Sinclair, S.; Zou, M.; Del Genio, C.I.; Kubeš, M.F.; Napier, R.; Jaworski, K.; et al. Adenylate cyclase activity of TIR1/AFB auxin receptors in plants. Nature 2022, 611, 133–138. [Google Scholar] [CrossRef]
- Gao, X.; Li, F.; Li, M.; Kianinejad, A.S.; Dever, J.K.; Wheeler, T.A.; Li, Z.; He, P.; Shan, L. Cotton GhBAK1 mediates Verticillium wilt resistance and cell death. J. Integr. Plant Biol. 2013, 55, 586–596. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Schiff, M.; Marathe, R.; Dinesh-Kumar, S.P. Tobacco Rar1, EDS1 and NPR1/NIM1 like genes are required for N-mediated resistance to tobacco mosaic virus. Plant J. 2002, 30, 415–429. [Google Scholar] [CrossRef]
- Xu, J.; Duan, X.; Yang, J.; Beeching, J.R.; Zhang, P. Coupled expression of Cu/Zn-superoxide dismutase and catalase in cassava improves tolerance against cold and drought stresses. Plant Signal. Behav. 2013, 8, e24525. [Google Scholar] [CrossRef] [PubMed]
- Ruegger, M.; Dewey, E.; Gray, W.M.; Hobbie, L.; Turner, J.; Estelle, M. The TIR1 protein of Arabidopsis functions in auxin response and is related to human SKP2 and yeast grr1p. Genes 1998, 12, 198–207. [Google Scholar] [CrossRef]
- Lakehal, A.; Chaabouni, S.; Cavel, E.; Le Hir, R.; Ranjan, A.; Raneshan, Z.; Novák, O.; Păcurar, D.I.; Perrone, I.; Jobert, F.; et al. A Molecular Framework for the Control of Adventitious Rooting by TIR1/AFB2-Aux/IAA-Dependent Auxin Signaling in Arabidopsis. Mol. Plant 2019, 12, 1499–1514. [Google Scholar] [CrossRef]
- Parry, G.; Calderon-Villalobos, L.I.; Prigge, M.; Peret, B.; Dharmasiri, S.; Itoh, H.; Lechner, E.; Gray, W.M.; Bennett, M.; Estelle, M. Complex regulation of the TIR1/AFB family of auxin receptors. Proc. Natl. Acad. Sci. USA 2009, 106, 22540–22545. [Google Scholar] [CrossRef]
- Hu, Z.; Keceli, M.A.; Piisila, M.; Li, J.; Survila, M.; Heino, P.; Brader, G.; Palva, E.T.; Li, J. F-box protein AFB4 plays a crucial role in plant growth, development and innate immunity. Cell Res. 2012, 22, 777–781. [Google Scholar] [CrossRef]
- Du, W.; Lu, Y.; Li, Q.; Luo, S.; Shen, S.; Li, N.; Chen, X. TIR1/AFB proteins: Active players in abiotic and biotic stress signaling. Front. Plant Sci. 2022, 13, 1083409. [Google Scholar] [CrossRef]
- Hey, S.J.; Byrne, E.; Halford, N.G. The interface between metabolic and stress signalling. Ann. Bot. 2010, 105, 197–203. [Google Scholar] [CrossRef]
- Ritonga, F.N.; Zhou, D.; Zhang, Y.; Song, R.; Li, C.; Li, J.; Gao, J. The Roles of Gibberellins in Regulating Leaf Development. Plants 2023, 12, 1243. [Google Scholar] [CrossRef]
- Sybilska, E.; Daszkowska-Golec, A. Alternative splicing in ABA signaling during seed germination. Front. Plant Sci. 2023, 14, 1144990. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Prime Name | Prime Sequence (5′-3′) |
|---|---|
| Gh_D08G0763-GFP-F | TCTAGAAAGCTTCTGCAGATGCATAAGAAAATGGC |
| Gh_D08G0763-GFP-R | CTTGCTCACCATGGTACCGAAAGCCTCAATCCAGA |
| GhSKP1-AD-F | GAGGCCAGTGAATTCATGGCTTCTACGGGTCGGAA |
| GhSKP1-AD-R | GAGCTCGATGGATCCGCTCAAACGCCCATTGATTC |
| Gh_A07G2125-BD-F | ATGGAGGCCGAATTCATGAA CATGAATTATTTCC |
| Gh_A07G2125-BD-R | CAGGTCGACGGATCCCAAAAGCCACACATAC |
| Gh_A10G0207-BD-F | ATGGAGGCCGAATTCATGGAACCATCGGAGCTG |
| Gh_A10G0207-BD-R | CAGGTCGACGGATCCGAGAGTAAGAACAGATG |
| Gh_D08G0763-BD-F | ATGGAGGCCGAATTC ATGCATAAGAAAATGGC |
| Gh_D08G0763-BD-R | CAGGTCGACGGATCCGAAAGCCTCAATCCAGA |
| Gh_D10G0187-BD-F | ATGGAGGCCGAATTCATGGAACCATCGGAGCTG |
| Gh_D10G0187-BD-R | CAGGTCGACGGATCCGAGGGTAAGAACAGATGG |
| Gh_D11G0671-BD-F | ATGGAGGCCGAATTCATGAATTATTTCCCAGATGAAG |
| Gh_D11G0671-BD-R | CAGGTCGACGGATCCCAAAATCAACACATATTCTGG |
| GhUBQ7-F | AGGCATTCCACCTGACCAAC- |
| GhUBQ7-R | CAGCGAGCTTGACCTTCTTC |
| Gh_D08G0763-vigs-F | GTGAGTAAGGTTACCGAATTCTCAAGCTTGTAAGCC |
| Gh_D08G0763-vigs-R | CGTGAGCTCGGTACCGGATCCGCAAGTCCTTGCAAC |
| GhCLA1-vigs-F | CGACGACAAGACCGTGACCATGCACAACATCGATGATTTAG |
| GhCLA1-vigs-F | GAGGAGAAGAGCCGTCATTAGCATGAATGATGAGTAGATTGCAC |
| Gh_A07G2125-qPCR-F | CAGGTTTCTTAGGGAGCTG |
| Gh_A07G2125-qPCR-R | CCACACGGTTTAACCTCAA |
| Gh_A08G0662-qPCR-F | CCAAGGATGGTGATCAGACG |
| Gh_A08G0662-qPCR-R | GAGACCATCAGTTGAAAACC |
| Gh_A08G1014-qPCR-F | TTCGTGGGGAACTGTTACG |
| Gh_A08G1014-qPCR-R | CACTTGCAAATTCGTAAACG |
| Gh_A10G0207-qPCR-F | CATGCACTAAACTTCAGAGGC |
| Gh_A10G0207-qPCR-R | GAACGGGCAGTCTCGTATCTC |
| Gh_A11G1077-qPCR-F | GGAACTGTTACGCTGTTAGC |
| Gh_A11G1077-qPCR-R | TCCGCTTAAGTCGAATCTCC |
| Gh_D08G0763-qPCR-F | AACGCCTGGTGGGTAGATGT |
| Gh_D08G0763-qPCR-R | ACCACATCCCAAAATCCAGA |
| Gh_D08G1288-qPCR-F | GGACTCGAAGTGCTCGCATC |
| Gh_D08G1288-qPCR-R | ACAACCCGAAGAAACAGCGA |
| Gh_D11G1288-qPCR-F | GGACAAATACTTCGACCTGG |
| Gh_D11G1288-qPCR-R | CAACTTGGAAGCAGCATTCG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Wu, C.; Guo, Y.; Ren, X.; Meng, Y.; Gao, Q.; Zhang, F.; Wang, Y.; Guo, J. Genome-Wide Analysis Elucidates the Roles of GhTIR1/AFB Genes Reveals the Function of Gh_D08G0763 (GhTIR1) in Cold Stress in G. hirsutum. Plants 2024, 13, 1152. https://doi.org/10.3390/plants13081152
Zhang X, Wu C, Guo Y, Ren X, Meng Y, Gao Q, Zhang F, Wang Y, Guo J. Genome-Wide Analysis Elucidates the Roles of GhTIR1/AFB Genes Reveals the Function of Gh_D08G0763 (GhTIR1) in Cold Stress in G. hirsutum. Plants. 2024; 13(8):1152. https://doi.org/10.3390/plants13081152
Chicago/Turabian StyleZhang, Xianliang, Cuicui Wu, Yutao Guo, Xiang Ren, Yongming Meng, Qi Gao, Fei Zhang, Yaping Wang, and Jinggong Guo. 2024. "Genome-Wide Analysis Elucidates the Roles of GhTIR1/AFB Genes Reveals the Function of Gh_D08G0763 (GhTIR1) in Cold Stress in G. hirsutum" Plants 13, no. 8: 1152. https://doi.org/10.3390/plants13081152
APA StyleZhang, X., Wu, C., Guo, Y., Ren, X., Meng, Y., Gao, Q., Zhang, F., Wang, Y., & Guo, J. (2024). Genome-Wide Analysis Elucidates the Roles of GhTIR1/AFB Genes Reveals the Function of Gh_D08G0763 (GhTIR1) in Cold Stress in G. hirsutum. Plants, 13(8), 1152. https://doi.org/10.3390/plants13081152