


Update on the Basic Understanding of Fusarium graminearum Virulence Factors in Common Wheat Research
 ,
,
Abstract
:1. Introduction
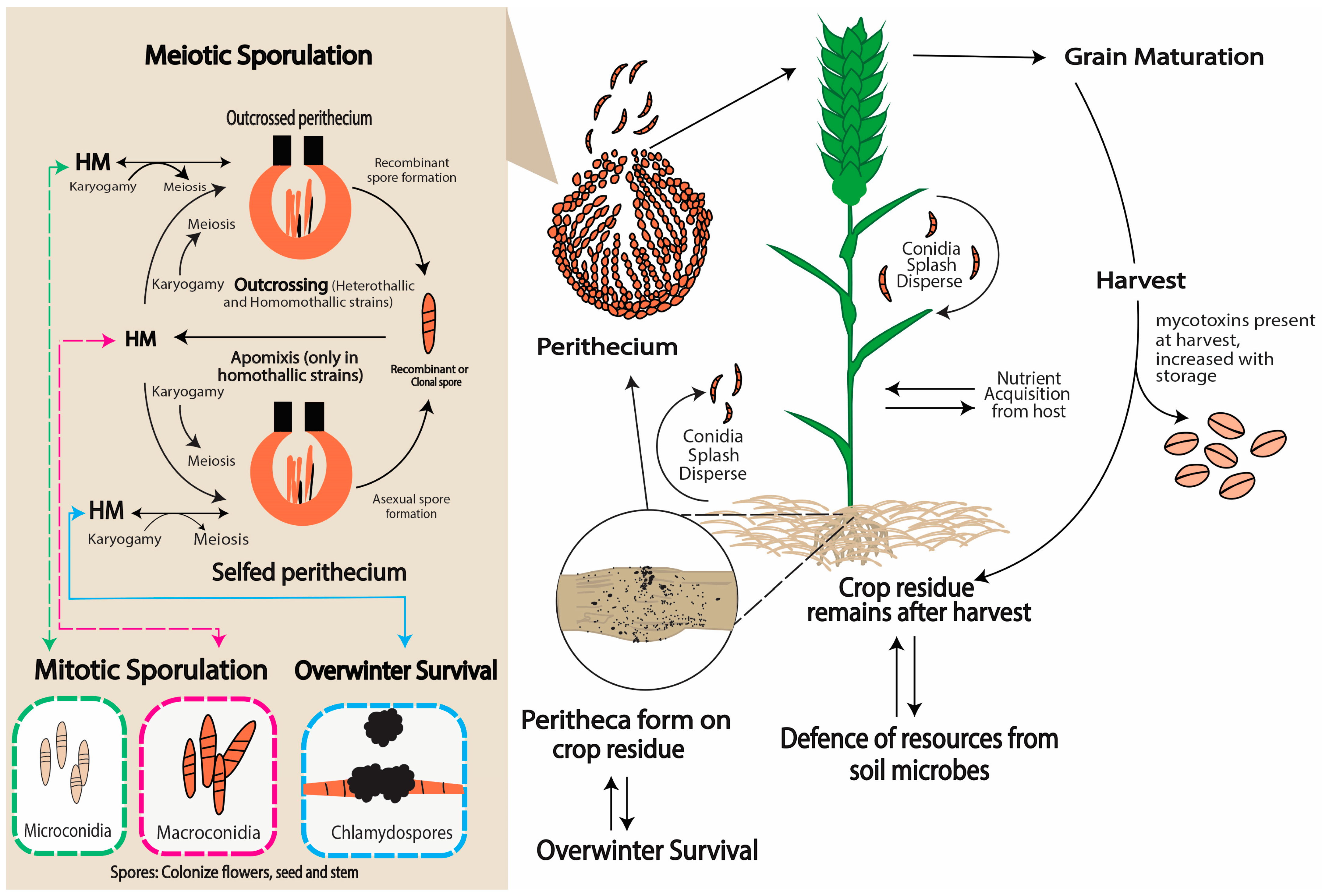
2. Distinguished Features of Fusarium graminearum Life Cycle
3. Fusarium graminearum Germination Leading Up to Infection
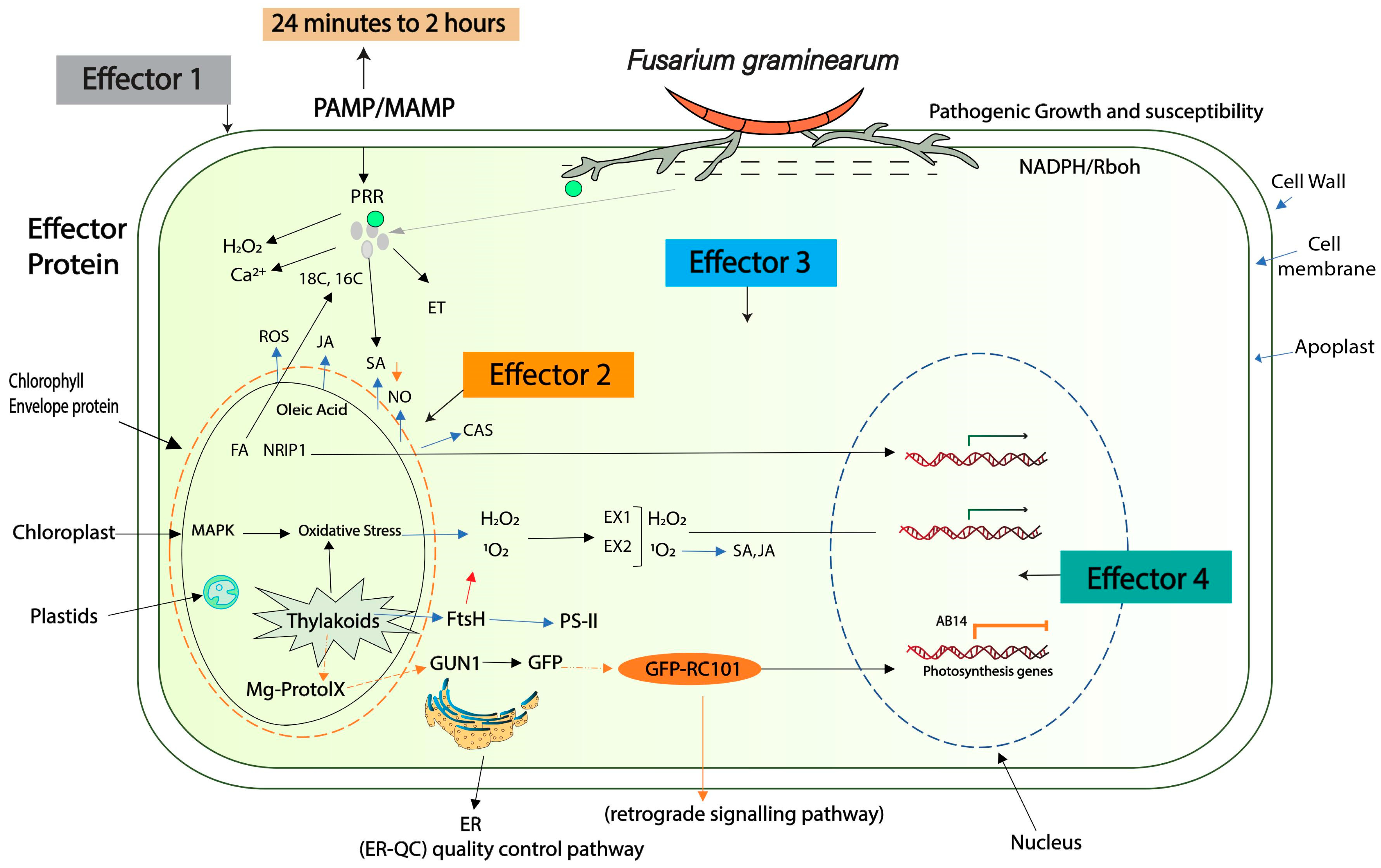
4. Role of Effectors in Colonization of Host Plant
5. Host Plant Resistance Mechanism against Effector Invasion
5.1. Effectors Inhibit R-Genes Expression through Chromatin Remodeling
5.2. Effector Protein Evolution through Repeat-Induced Point Mutation
6. Important Unresolved Questions
6.1. Molecular Pathogenicity
6.2. How Can We Develop Resistant Cultivars More Efficiently?
6.3. Climate Change on FHB Disease and Solution
6.4. Strengthen the Study of Molecular Mechanisms of Pathogenic Bacteria’s Pathogenesis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Hao, Y.; Rasheed, A.; Zhu, Z.; Wulff, B.B.H.; He, Z. Harnessing Wheat Fhb1 for Fusarium Resistance. Trends Plant Sci. 2020, 25, 1–3. [Google Scholar] [PubMed]
- Stack, R.W. History of Fusarium Head Blight with Emphasis on North America. In Fusarium Head Blight of Wheat and Barley; Leonard, K.J., Bushnell, W.R., Eds.; The American Phytopathological Society: St. Paul, MI, USA, 2003. [Google Scholar]
- McMullen, M. Impacts of Fusarium Head Blight on the North American Agriculture Community: The Power of One Disease to Catapult Change. In Fusarium Head Blight of Wheat and Barley; Leonard, K.J., Bushnell, W.R., Eds.; The American Phytopathological Society: St. Paul, MI, USA, 2003. [Google Scholar]
- Burgess, L.W.; Klein, T.A.; Bryden, W.L.; Tobin, N.F. Head Blight of Wheat Caused by Fusarium-Graminearum group 1 in New South Wales Australia. Australas. Plant Pathol. 1983, 16, 72–78. [Google Scholar]
- Wulff, B.B.H.; Jones, J.D.G. Breeding a Fungal Gene into wheat. Science 2020, 368, 822–823. [Google Scholar] [PubMed]
- Parry, D.W.; Jenkinson, P.; McLeod, L. Fusarium Ear Blight (scab) in Small Grain Cereals-a Review. Plant Pathol. 1995, 44, 207–238. [Google Scholar]
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [PubMed]
- Schulze, S.; Yu, L.; Hua, C.; Zhang, L.; Kolb, D.; Weber, H.; Ehinger, A.; Svenja, C.; Stahl, M.; Franz-Wachtel, M.; et al. The Arabidopsis TIR-NBS-LRR Protein CSA1 Guards BAK1-BIR3 Homeostasis and Mediates Convergence of Pattern- and Effector-induced Immune responses. Cell Host Microbe 2022, 30, 1717–1731. [Google Scholar] [CrossRef] [PubMed]
- Zipfel, C. Plant Pattern-recognition Receptors. Trends Immunol. 2014, 35, 345–351. [Google Scholar] [PubMed]
- Balint-Kurti, P. The Plant Hypersensitive Response: Concepts, Control and Consequences. Mol. Plant Pathol. 2019, 20, 1163–1178. [Google Scholar] [PubMed]
- Blümke, A.; Falter, C.; Herrfurth, C.; Sode, B.; Bode, R.; Schäfer, W.; Feussner, I.; Voigt, C.A. Secreted fungal effector lipase releases free fatty acids to inhibit innate immunity-related callose formation during wheat head infection. Plant Physiol. 2014, 165, 346–358. [Google Scholar]
- Yang, C.C.; Wang, Z.Y.; Cheng, C.M. Insights into Superinfection Immunity Regulation of Xanthomonas Axonopodis Filamentous Bacteriophage cf. Curr. Microbiol. 2023, 81, 42. [Google Scholar]
- Petit-Houdenot, Y.; Fudal, I. Complex Interactions between Fungal Avirulence Genes and Their Corresponding Plant Resistance Genes and Consequences for Disease Resistance Management. Front. Plant Sci. 2017, 8, 1072. [Google Scholar] [PubMed]
- Ausubel, F.M. Are Innate Immune Signaling Pathways in Plants and Animals Conserved? Nat. Immunol. 2005, 6, 973–979. [Google Scholar] [PubMed]
- Flor, H.H. Current Status of the Gene-for-gene Concept. Annu. Rev. Phytopathol. 1971, 9, 275–296. [Google Scholar] [CrossRef]
- Flor, H.H. Inheritance of Pathogenicity in Melampsora lini. Phytopathology 1942, 32, 653–669. [Google Scholar]
- Trail, F. For Blighted Waves of Grain: Fusarium graminearum in the Postgenomics Era. Plant Physiol. 2009, 149, 103–110. [Google Scholar] [PubMed]
- Van, W.S.; Wingfield, B.D.; de Vos, L.; van der Merwe, N.A.; Santana, Q.C.; Steenkamp, E.T. Repeat-Induced Point Mutations Drive Divergence between Fusarium Circinatum and its Close Relatives. Pathogens 2019, 8, 298. [Google Scholar] [CrossRef]
- Klix, V.; Nowrousian, M.; Ringelberg, C.; Loros, J.J.; Dunlap, J.C.; Pöggeler, S. Functional Characterization of MAT1-1-specific Mating-type Genes in the Homothallic Ascomycete Sordaria macrospora Provides New Insights into Essential and Nonessential Sexual Regulators. Eukaryot Cell 2010, 9, 894–905. [Google Scholar]
- Guo, Y.; Yao, S.; Yuan, T.; Wang, Y.; Zhang, D.; Tang, W. The Spatiotemporal Control of KatG2 Catalase-peroxidase Contributes to the Invasiveness of Fusarium graminearum in Host Plants. Mol. Plant Pathol. 2019, 20, 685–700. [Google Scholar]
- Yun, S.H.; Arie, T.; Kaneko, S.; Yoder, O.C.; Turgeon, B.G. Molecular Organization of Mating Type Loci in Heterothallic, Homothallic and Asexual Gibberella/Fusarium species. Fungal Genet. Biol. 2000, 31, 7–20. [Google Scholar] [PubMed]
- Mentges, M.; Glasenapp, A.; Boenisch, M.; Malz, S.; Henrissat, B.; Frandsen, R.J.N.; Güldener, U.; Münsterkötter, M.; Bormann, J.; Lebrun, M.H.; et al. Infection Cushions of Fusarium graminearum are Fungal Arsenals for Wheat Infection. Mol. Plant Pathol. 2020, 21, 1070–1087. [Google Scholar]
- Ofek, P.; Yeini, E.; Arad, G.; Danilevsky, A.; Pozzi, S.; Luna, C.B.; Dangoor, S.I.; Grossman, R.; Ram, Z.; Shomron, N.; et al. Deoxyhypusine hydroxylase: A Novel Therapeutic Target Differentially Expressed in Short-term vs Long-term Survivors of Glioblastoma. Int. J. Cancer 2023, 153, 654–668. [Google Scholar]
- Zhao, F.; Yuan, Z.; Wen, W.; Huang, Z.; Mao, X.; Zhou, M.; Hou, Y. FgMet3 and FgMet14 Related to Cysteine and Methionine Biosynthesis Regulate Vegetative Growth, Sexual Reproduction, Rathogenicity, and Sensitivity to Fungicides in Fusarium graminearum. Front. Plant Sci. 2022, 13, 011709. [Google Scholar]
- Villafana, R.T.; Ramdass, A.C.; Rampersad, S.N. Selection of Fusarium Trichothecene Toxin Genes for Molecular Detection Depends on TRI Gene Cluster Organization and Gene Function. Toxins 2019, 11, 36. [Google Scholar] [CrossRef]
- Zhang, Y.Z.; Wei, Z.Z.; Liu, C.H.; Chen, Q.; Xu, B.J.; Guo, Z.R.; Cao, Y.L.; Wang, Y.; Han, Y.N.; Chen, C.; et al. Linoleic Acid Isomerase Gene FgLAI12 Affects Sensitivity to Salicylic Acid, Mycelial Growth and Virulence of Fusarium graminearum. Sci. Rep. 2017, 7, 46129. [Google Scholar] [CrossRef]
- Martinez-Rocha, A.L.; Woriedh, M.; Chemnitz, J.; Willingmann, P.; Kröger, C.; Hadeler, B.; Hauber, J.; Schäfer, W. Posttranslational Hypusination of the Eukaryotic Translation Initiation Factor-5A Regulates Fusarium graminearum Virulence. Sci. Rep. 2016, 6, 24698. [Google Scholar]
- Ellinger, D.; Sode, B.; Falter, C.; Voigt, C.A. Resistance of Callose Synthase Activity to Free Fatty acid Inhibition as an Indicator of Fusarium Head Blight Resistance in Wheat. Plant Signal Behav. 2014, 9, e28982. [Google Scholar]
- Jia, L.J.; Tang, H.Y.; Wang, W.Q.; Yuan, T.L.; Wei, W.Q.; Pang, B.; Gong, X.M.; Wang, S.F.; Li, Y.J.; Zhang, D.; et al. A Linear Nonribosomal Octapeptide from Fusarium graminearum Facilitates Cell-to-cell Invasion of Wheat. Nat. Commun. 2019, 10, 922. [Google Scholar]
- Kamoun, S.A. Catalogue of the Effector Secretome of Plant Pathogenic Oomycetes. Annu. Rev. Phytopathol. 2006, 44, 41–60. [Google Scholar]
- Kamoun, S. Groovy times: Filamentous Pathogen Effectors Revealed. Curr. Opin. Plant Biol. 2007, 10, 358–365. [Google Scholar]
- Zhang, X.W.; Jia, L.J.; Zhang, Y.; Jiang, G.; Li, X.; Zhang, D.; Tang, W.H. In Planta Stage-specific Fungal Gene Profiling Elucidates the Molecular Strategies of Fusarium graminearum Growing inside Wheat Coleoptiles. Plant Cell 2012, 24, 5159–5176. [Google Scholar]
- Kugler, K.G.; Siegwart, G.; Nussbaumer, T.; Ametz, C.; Spannagl, M.; Steiner, B.; Lemmens, M.; Mayer, K.F.; Buerstmayr, H.; Schweiger, W. Quantitative Trait Loci-dependent Analysis of a Gene Co-expression Network Associated with Fusarium Head Blight Resistance in Bread Wheat (Triticum aestivum L.). BMC Genom. 2013, 14, 728. [Google Scholar]
- Jiang, C.; Hei, R.; Yang, Y.; Zhang, S.; Wang, Q.; Wang, W.; Zhang, Q.; Yan, M.; Zhu, G.; Huang, P.; et al. An Orphan Protein of Fusarium graminearum Modulates Host Immunity by Mediating Proteasomal Degradation of TaSnRK1α. Nat. Commun. 2020, 11, 4382. [Google Scholar]
- Min, K.; Son, H.; Lee, J.; Choi, G.J.; Kim, J.C.; Lee, Y.W. Peroxisome Function Is Required for Virulence and Survival of Fusarium graminearum. Mol. Plant Microbe Interact. 2012, 25, 1617–1627. [Google Scholar]
- Hao, G.; McCormick, S.; Usgaard, T.; Tiley, H.; Vaughan, M.M. Characterization of Three Fusarium graminearum Effectors and Their Roles During Fusarium Head Blight. Front. Plant Sci. 2020, 11, 579553. [Google Scholar]
- Walton, J.D. Deconstructing the Cell Wall. Plant Physiol. 1994, 104, 1113–1118. [Google Scholar]
- Wang, Q.; Han, C.; Ferreira, A.O.; Yu, X.; Ye, W.; Tripathy, S.; Kale, S.D.; Gu, B.; Sheng, Y.; Sui, Y.; et al. Transcriptional Programming and Functional Interactions within the Phytophthora sojae RXLR Effector Repertoire. Plant Cell 2011, 23, 2064–2086. [Google Scholar]
- Brown, N.A.; Antoniw, J.; Hammond-Kosack, K.E. The Predicted Secretome of the Plant Pathogenic Fungus Fusarium graminearum: A Refined Comparative Analysis. PLoS ONE 2012, 7, e33731. [Google Scholar]
- Köster, P.; DeFalco, T.A.; Zipfel, C. Ca2+ Signals in Plant Immunity. EMBO J. 2022, 41, e110741. [Google Scholar]
- Gebrie, A. Transposable Elements as Essential Elements in the Control of Gene Expression. Mob. DNA 2023, 9, 14. [Google Scholar]
- Lu, S.; Edwards, M.C. Genome-wide Analysis of Small Secreted Cysteine-rich Proteins Identifies Candidate Effector Proteins Potentially Involved in Fusarium graminearum-wheat interactions. Phytopathology 2016, 106, 166–176. [Google Scholar]
- Lapin, G.; Van den Ackerveken, G. Susceptibility to Plant Disease: More than a Failure of Host Immunity. Trends Plant Sci. 2013, 18, 546–554. [Google Scholar]
- Van Schie, C.C.; Takken, F.L. Susceptibility Genes: How to be a Good Host. Annu. Rev. Phytopathol. 2014, 52, 551–581. [Google Scholar]
- Zaidi, S.S.; Mukhtar, M.S.; Mansoor, S. Genome Editing: Targeting Susceptibility Genes for Plant Disease Resistance. Trends Biotechnol. 2018, 36, 898–906. [Google Scholar]
- Guenther, J.C.; Hallen-Adams, H.E.; Bücking, H.; Shachar-Hill, Y.; Trail, F. Triacylglyceride Metabolism by Fusarium graminearum During Colonization and Sexual Development on Wheat. Mole. Plant Microbe Interact. 2009, 12, 1492–1503. [Google Scholar] [CrossRef]
- Hao, G.; Tiley, H.; McCormick, S. Chitin Triggers Tissue-Specific Immunity in Wheat Associated with Fusarium Head Blight. Front. Plant Sci. 2022, 13, 832502. [Google Scholar]
- Zuo, N.; Bai, W.Z.; Wei, W.Q.; Yuan, T.L.; Zhang, D.; Wang, Y.Z.; Tang, W.H. Fungal CFEM Effectors Negatively Regulate a Maize Wall-associated Kinase by Interacting with its Alternatively Spliced Variant to Dampen Resistance. Cell Rep. 2022, 41, 111877. [Google Scholar]
- Houterman, P.M.; Ma, L.; van Ooijen, G.; de Vroomen, M.J.; Cornelissen, B.J.C.; Takken, F.L.W.; Rep, M. The Effector Protein Avr2 of the Xylem-colonizing Fungus Fusarium Oxysporum Activates the Tomato Resistance Protein I-2 Intracellularly. Plant J. 2009, 58, 970–978. [Google Scholar] [CrossRef]
- Catanzariti, A.M.; Lim, G.T.T.; Jones, D.A. The Tomato I-3 Gene: A Novel Gene for Resistance to Fusarium wilt Disease. New Phytol. 2015, 207, 106–118. [Google Scholar] [CrossRef]
- Kong, X.; van Diepeningen, A.D.; van der Lee, T.A.J.; Waalwijk, C.; Xu, J.; Xu, J.; Zhang, H.; Chen, W.; Feng, J. The Fusarium graminearum Histone Acetyltransferases Are Important for Morphogenesis, DON Biosynthesis, and Pathogenicity. Front. Microbiol. 2018, 9, 654. [Google Scholar]
- Luger, K.; Rechsteiner, T.J.; Flaus, A.J.; Waye, M.M.Y.; Richmond, T.J. Characterization of Nucleosome Core Particles Containing Histone Proteins Made in Bacteria. J. Mol. Biol. 1997, 272, 301–311. [Google Scholar] [CrossRef]
- Badeaux, A.I.; Shi, Y. Emerging Roles for Chromatin as a Signal Integration and Storage Platform. Nat. Rev. Mol. Cell Biol. 2013, 14, 211–224. [Google Scholar] [CrossRef]
- Lanver, D.; Muller, A.N.; Happel, P.; Schweizer, G.; Haas, F.B.; Franitza, M.; Pellegrin, C.; Reissmann, S.; Altmuller, J.; Rensing, S.A.; et al. The Biotrophic Development of Ustilago Maydis Studied by RNA-Seq Analysis. Plant Cell 2018, 30, 300–323. [Google Scholar] [CrossRef]
- Kong, L.; Qiu, X.; Kang, J.; Wang, Y.; Chen, H.; Huang, J.; Qiu, M.; Zhao, Y.; Kong, G.; Ma, Z.; et al. A Phytophthora Effector Manipulates Host Histone Acetylation and Reprograms Defense Gene Expression to Promote Infection. Curr. Biol. 2017, 27, 981–991. [Google Scholar] [CrossRef]
- Ori, N.; Eshed, Y.; Paran, I.; Presting, G.; Aviv, D.; Tanksley, S.; Zamir, D.; Fluhr, R. The I2C Family from the Wilt Disease Resistance Locus I2 Belongs to the Nucleotide Binding, Leucine-rich Repeat Superfamily of Plant Resistance Genes. Plant Cell 1997, 9, 521–532. [Google Scholar]
- Rep, M.; Meijer, M.; Houterman, P.M.; van der Does, H.C.; Cornelissen, B.J.C. Fusarium oxysporum Evades I-3-Mediated Resistance without Altering the Matching Avirulence Gene. Mol. Plant Microbe Interact. 2005, 18, 15–23. [Google Scholar] [CrossRef]
- Gu, Q.; Wang, Y.; Zhao, X.; Yuan, B.; Zhang, M.; Tan, Z.; Zhang, X.; Chen, Y.; Wu, H.; Luo, Y.; et al. Inhibition of Histone Acetyltransferase GCN5 by a Transcription Factor FgPacC Controls Fungal Adaptation to Host-derived Iron Stress. Nucleic Acid Res. 2022, 50, 6190–6210. [Google Scholar] [CrossRef]
- Zhi, P.; Kong, L.; Liu, J.; Zhang, X.; Wang, X.; Li, H.; Sun, M.; Li, Y.; Chang, C. Histone Deacetylase TaHDT701 Functions in TaHDA6-TaHOS15 Complex to Regulate Wheat Defense Responses to Blumeria graminis f. sp. tritici. Int. J. Mol. Sci. 2020, 21, 2640. [Google Scholar] [CrossRef]
- Sagaram, U.S.; Pandurangi, R.; Kaur, J.; Smith, T.J.; Shah, D.M. Structure-activity Determinants in Antifungal Plant Defensins MsDef1 and MtDef4 with Different Modes of Action against Fusarium graminearum. PLoS ONE 2011, 6, e18550. [Google Scholar] [CrossRef]
- Fouché, S.; Oggenfuss, U.; Chanclud, E.; Croll, D.A. Devil’s Bargain with Transposable Elements in Plant Pathogens. Trends Genet. 2022, 38, 222–230. [Google Scholar] [CrossRef]
- Daboussi, M.J.; Capy, P. Transposable Elements in Filamentous Fungi. Annu. Rev. Microbiol. 2003, 57, 275–299. [Google Scholar] [CrossRef]
- Van Wyk, S.; Wingfield, B.D.; De Vos, L.; van der Merwe, N.A.; Steenkamp, N.A. Genome-wide Analysis of Repeat-Induced Point Mutations in the Ascomycota. Front. Microbiol. 2021, 11, 622368. [Google Scholar] [CrossRef]
- Amselem, J.; Quesneville, H.; Oliver, R.P.; Wincker, P.; Balesdent, M.H.; Howlett, B.J. Effector Diversification within Compartments of the Leptosphaeria Maculans Genome Affected by Repeat-Induced Point mutations. Nat. Commun. 2011, 2, 202. [Google Scholar]
- Komluski, J.; Habig, M.; Stukenbrock, E.H. Repeat-Induced Point Mutation and Gene Conversion Coinciding with Heterochromatin Shape the Genome of a Plant-Pathogenic Fungus. mBio 2023, 14, e0329022. [Google Scholar] [CrossRef]
- Van Wyk, S.; Harrison, C.H.; Wingfield, B.D.; De Vos, L.; van der Merwe, N.A.; Steenkamp, E.T. The RIPper, a Web-based Tool for Genome-wide Quantification of Repeat-Induced Point (RIP) Mutations. PeerJ 2019, 7, e7447. [Google Scholar] [CrossRef]
- Hartmann, F.E.; Sánchez-Vallet, A.; McDonald, B.A.; Croll, D.A. Fungal Wheat Pathogen Evolved Host Specialization by Extensive Chromosomal Rearrangements. ISME J. 2017, 11, 1189–1204. [Google Scholar] [CrossRef]
- Selker, E.U. Premeiotic Instability of Repeated Sequences in Neurospora crassa. Annu. Rev. Genet. 1990, 24, 579–613. [Google Scholar] [CrossRef]
- Selker, E.U. Repeat-induced Gene Silencing in Fungi. Adv. Genet. 2002, 46, 439–450. [Google Scholar]
- Selker, E.U.; Cambareri, E.B.; Jensen, B.C.; Haack, K. Rearrangement of Duplicated DNA in Specialized Cells of Neurospora. Cell 1987, 51, 741–752. [Google Scholar] [CrossRef]
- Galagan, J.E.; Selker, E.U. RIP: The Evolutionary Cost of Genome Defense. Trends Genet. 2004, 20, 417–423. [Google Scholar] [CrossRef]
- Son, H.; Min, K.; Lee, J.; Raju, N.B.; Lee, Y.W. Meiotic Silencing in the Homothallic Fungus Gibberella zeae. Fungal Biol. 2011, 115, 1290–1302. [Google Scholar] [CrossRef]
- Sridhar, P.S.; Trofimova, D.; Subramaniam, R.; González-Peña Fundora, D.; Foroud, N.A.; Allingham, J.S.; Loewen, M.C. Ste2 receptor-mediated chemotropism of Fusarium graminearum Contributes to its Pathogenicity against Wheat. Sci. Rep. 2020, 10, 10770. [Google Scholar] [CrossRef] [PubMed]
- Urban, M.; King, R.; Hassani-Pak, K.; Hammond-Kosack, K.E. Whole-genome Analysis of Fusarium graminearum Insertional Mutants Identifies Virulence Associated Genes and Unmasks Untagged Chromosomal Deletions. BMC Genom. 2015, 16, 261. [Google Scholar] [CrossRef] [PubMed]
- King, R.; Urban, M.; Hammond-Kosack, M.C.; Hassani-Pak, K.; Hammond-Kosack, K.E. The completed genome sequence of the pathogenic ascomycete fungus Fusarium graminearum. BMC Genom. 2015, 16, 544. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.K.; Jo, S.M.; Kim, G.Y.; Kim, D.W.; Kim, Y.K.; Yun, S.H. A Large-Scale Functional Analysis of Putative Target Genes of Mating-Type Loci Provides Insight into the Regulation of Sexual Development of the Cereal Pathogen Fusarium graminearum. PLoS Genet. 2015, 11, e1005486. [Google Scholar] [CrossRef] [PubMed]
- Irelan, J.T.; Selker, E.U. Cytosine Methylation Associated with Repeat-induced Point Mutation Causes Epigenetic Gene Silencing in Neurospora crassa. Genetics 1997, 146, 509–523. [Google Scholar] [CrossRef]
- Hane, J.K.; Oliver, R.P. In silico Reversal of Repeat-induced Point Mutation (RIP) Identifies the Origins of Repeat Families and Uncovers Obscured Duplicated Genes. BMC Genom. 2010, 11, 655. [Google Scholar] [CrossRef]
- Chen, L.; Wang, H.; Yang, J.; Yang, X.; Zhang, M.; Zhao, Z.; Fan, Y.; Wang, C.; Wang, J. Bioinformatics and Transcriptome Analysis of CFEM Proteins in Fusarium graminearum. J. Fungi 2021, 7, 871. [Google Scholar] [CrossRef]
- Cai, N.; Liu, R.; Yan, D.; Zhang, N.; Zhu, K.; Zhang, D.; Nong, X.; Tu, X.; Zhang, Z.; Wang, G. Bioinformatics Analysis and Functional Characterization of the CFEM Proteins of Metarhizium anisopliae. J. Fungi 2022, 8, 661. [Google Scholar] [CrossRef]
- Xu, M.; Wang, Q.; Wang, G.; Zhang, X.; Liu, H.; Jiang, C. Combatting Fusarium head blight: Advances in Molecular Interactions between Fusarium graminearum and Wheat. Phytopathol. Res. 2022, 4, 37. [Google Scholar] [CrossRef]
- Rocher, F.; Alouane, T.; Philippe, G.; Martin, M.L.; Label, P.; Langin, T.; Bonhomme, L. Fusarium graminearum Infection Strategy in Wheat Involves a Highly Conserved Genetic Program That Controls the Expression of a Core Effectome. Int. J. Mol. Sci. 2022, 23, 1914. [Google Scholar] [CrossRef]
- Hammond-Kosack, K.E.; Rudd, J.J. Plant Resistance Signalling Hijacked by a Necrotrophic Fungal Pathogen. Plant Signal. Behav. 2008, 3, 993–995. [Google Scholar] [CrossRef]
- Perincherry, L.; Lalak-Kańczugowska, J.; Stępień, Ł. Fusarium-Produced Mycotoxins in Plant-Pathogen Interactions. Toxins 2019, 11, 664. [Google Scholar] [CrossRef]
- Zhao, X.M.; Zhang, X.W.; Tang, W.H.; Chen, L. FPPI: Fusarium graminearum Protein-protein Interaction Database. J. Proteome Res. 2009, 8, 4714–4721. [Google Scholar] [CrossRef]
- Bahrini, I.; Ogawa, T.; Kobayashi, F.; Kawahigashi, H.; Handa, H. Overexpression of the Pathogen-inducible Wheat TaWRKY45 Gene Confers Disease Resistance to Multiple Fungi in Transgenic Wheat Plants. Breed Sci. 2011, 61, 319–326. [Google Scholar] [CrossRef]
- Buerstmayr, M.; Buerstmayr, H. The Effect of the Rht1 Haplotype on Fusarium head blight Resistance in Relation to Type and Level of Background Resistance and in Combination with Fhb1 and Qfhs.ifa-5A. Theor. Appl. Genet. 2022, 135, 1985–1996. [Google Scholar] [CrossRef]
- Mesterhazy, A. What Is Fusarium Head Blight (FHB) Resistance and What Are Its Food Safety Risks in Wheat? Problems and Solutions—A Review. Toxins 2024, 16, 31. [Google Scholar] [CrossRef]
- Berraies, S.; Cuthbert, R.; Knox, R.; Singh, A.; DePauw, R.; Ruan, Y.; Bokore, F.; Henriquez, M.A.; Kumar, S.; Burt, A.; et al. High-density Genetic Mapping of Fusarium head blight Resistance and Agronomic Traits in Spring Wheat. Front. Plant Sci. 2023, 14, 1134132. [Google Scholar] [CrossRef]
- Buerstmayr, M.; Steiner, B.; Buerstmayr, H. Breeding for Fusarium head blight Resistance in Wheat—Progress and Challenges. Plant Breed. 2020, 139, 429–454. [Google Scholar] [CrossRef]
- Wang, H.; Sun, S.; Ge, W.; Zhao, L.; Hou, B.; Wang, K.; Lyu, Z.; Chen, L.; Xu, S.; Guo, J.; et al. Horizontal gene transfer of Fhb7 from fungus underlies Fusarium head blight resistance in wheat. Science 2020, 368, eaba5435. [Google Scholar]
- Su, Z.; Bernardo, A.; Tian, B.; Chen, H.; Wang, S.; Ma, H.; Cai, S.; Liu, D.; Zhang, D.; Li, T.; et al. A deletion mutation in TaHRC confers Fhb1 resistance to Fusarium head blight in wheat. Nat Genet. 2019, 51, 1099–1105. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Name or ID | Gene Family | Sub-Cellular Location | Go Term Annotation |
|---|---|---|---|
| FGSG_02040T0 | HAT_GNAT >> ELP3 | Nuclear, cytoplasmic | GO:0008080: N-acetyltransferase activity, GO:0046933: hydrogen-transporting ATP synthase activity, rotational mechanism |
| FGSG_00280T0 | HAT_GNAT >> GCN5 | nuclear | GO:0008080: N-acetyltransferase activity |
| FGSG_04254T0 | HAT_MYST >> Esa1 | nuclear | Not found |
| FGSG_06047T0 | HAT_MYST >> Sas2 | nuclear | Not found |
| FGSG_08481T0 | HAT_MYST >> Tip60 | nuclear | GO:0005515: protein binding, GO:0007242: intracellular signaling cascade GO:0006355: regulation of transcription, DNA-dependent GO:0008270: zinc ion binding |
| FGSG_02567T0 | HMT >> DOT1 | nuclear | Not found |
| FGSG_01134T0 | HMT >> PRMT_1 | Mitochondrial, cytoplasmic | GO:0008757: S-adenosylmethionine-dependent methyltransferase activity GO:0005737: cytoplasm GO:0006479: protein amino acid methylation GO:0008276: protein methyltransferase activity |
| FGSG_10718T0 | HMT >> PRMT_1 | cytoplasmic | GO:0008757: S-adenosylmethionine-dependent methyltransferase activity |
| FGSG_07445T0 | HMT >> SET1 | nuclear | GO:0000166: nucleotide binding |
| FGSG_05558T0 | HMT >> SET2 | nuclear | Not found |
| FGSG_01558T0 | HDM >> JHDM3_JMJD2 | Nuclear | GO:0005515: protein binding GO:0006355: regulation of transcription, DNA-dependent GO:0008270: zinc ion binding |
| FGSG_00780T0 | HDAC >> ClassI | Nuclear | GO:0016575: histone deacetylation GO:0004407: histone deacetylase activity GO:0005634: nucleus |
| FGSG_01353T0 | HDAC >> ClassI | Cytoplasmic, nuclear | GO:0016575: histone deacetylation GO:0004407: histone deacetylase activity GO:0005634: nucleus |
| FGSG_04324T0 | HDAC >> ClassIIB | Cytoplasmic, nuclear | Not found |
| FGSG_09218T0 | HDAC >> ClassIII | Nuclear, cytoplasmic | GO:0006342: chromatin silencing GO:0003677: DNA binding GO:0006355: regulation of transcription, DNA-dependent GO:0005677: chromatin silencing complex |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buttar, Z.A.; Cheng, M.; Wei, P.; Zhang, Z.; Lv, C.; Zhu, C.; Ali, N.F.; Kang, G.; Wang, D.; Zhang, K. Update on the Basic Understanding of Fusarium graminearum Virulence Factors in Common Wheat Research. Plants 2024, 13, 1159. https://doi.org/10.3390/plants13081159
Buttar ZA, Cheng M, Wei P, Zhang Z, Lv C, Zhu C, Ali NF, Kang G, Wang D, Zhang K. Update on the Basic Understanding of Fusarium graminearum Virulence Factors in Common Wheat Research. Plants. 2024; 13(8):1159. https://doi.org/10.3390/plants13081159
Chicago/Turabian StyleButtar, Zeeshan Ali, Mengquan Cheng, Panqin Wei, Ziwei Zhang, Chunlei Lv, Chenjia Zhu, Nida Fatima Ali, Guozhang Kang, Daowen Wang, and Kunpu Zhang. 2024. "Update on the Basic Understanding of Fusarium graminearum Virulence Factors in Common Wheat Research" Plants 13, no. 8: 1159. https://doi.org/10.3390/plants13081159
APA StyleButtar, Z. A., Cheng, M., Wei, P., Zhang, Z., Lv, C., Zhu, C., Ali, N. F., Kang, G., Wang, D., & Zhang, K. (2024). Update on the Basic Understanding of Fusarium graminearum Virulence Factors in Common Wheat Research. Plants, 13(8), 1159. https://doi.org/10.3390/plants13081159