
Limitations of Plant Stress Tolerance upon Heat and CO2 Exposure in Black Poplar: Assessment of Photosynthetic Traits and Stress Volatile Emissions


{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
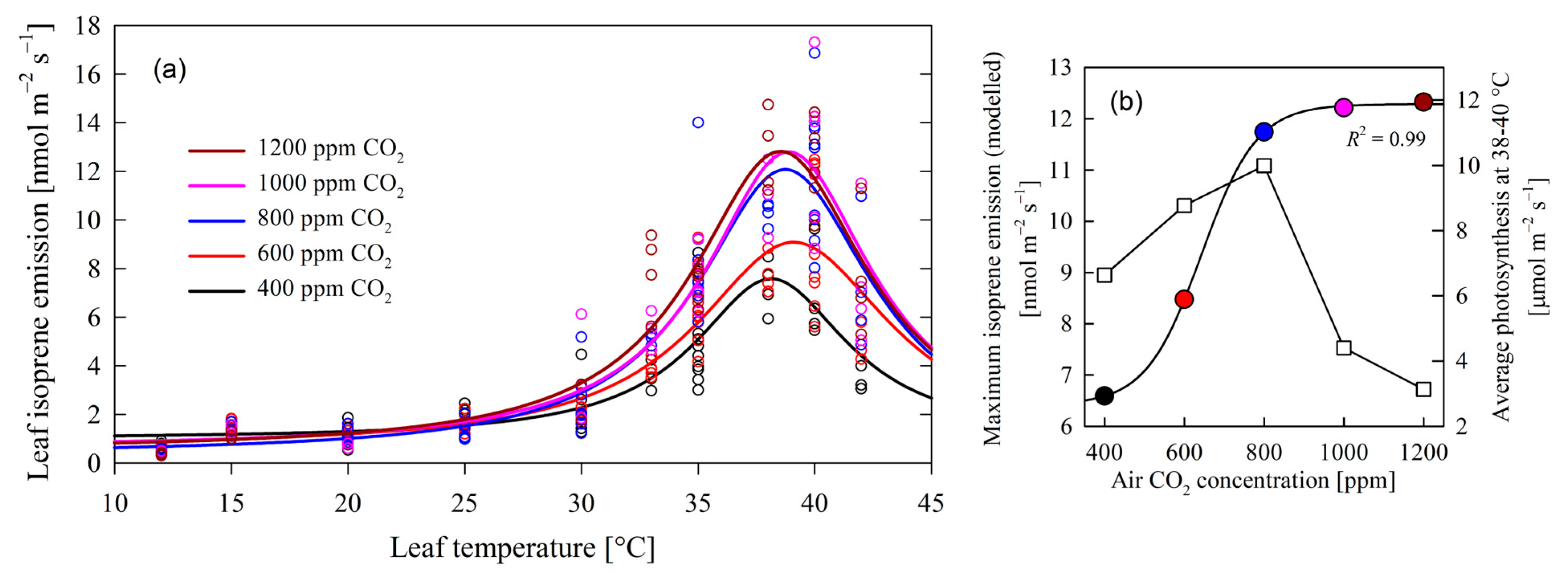
2.1. CO2-Mediated Regulation of Isoprene Emissions and Temperature Optima
- (1)
- As CO2 levels increased from Φisoprene = 6.419 nmol m−2 s−1, there was a corresponding rise in maximum isoprene emission (Figure 1b), progressing from 6.59 nmol m−2 s−1 at 400 ppm CO2 to 11.74 nmol m−2 s−1 at 800 ppm CO2. Based on this regression, the maximum rise in emissions per unit of [CO2] increase was found at 642 ppm CO2, with an emission value of Φisoprene = 9.352 nmol m−2 s−1 and a maximum rate of emissions of 0.05285 nmol m−2 s−1 ppm CO2−1. This positive correlation underlines the influence of CO2 concentration on isoprene biosynthesis, aligning with previous studies highlighting the role of carbon availability in secondary metabolite production [27]. Isoprene is produced in the chloroplast primarily from its immediate precursor dimethylallyl diphosphate (DMADP) via isoprene synthase (IspS), which is synthesized via the methylerythritol 4-phosphate (MEP) pathway [28]. Isoprene production is therefore controlled by the supply of DMADP, and by the activity of isoprene synthase [29,30].
- (2)
- Intriguingly, the observed increase in maximum isoprene emission plateaued beyond 800 ppm CO2 at about 12.285 nmol m−2 s−1 (Figure 1b). This suggests a potential saturation point of isoprene synthesis, possibly attributed to limiting enzymatic activity and substrate availability. This stands in contrast to the presumed stimulatory effect of elevated CO2 concentrations, which is expected to facilitate de novo carbon assimilation and enhance yields for isoprene synthesis and emissions. Such findings prompt further inquiry into the molecular mechanisms of isoprene biosynthesis and the potential existence of regulatory feedback loops, as found in other poplars subjected to increased levels of [CO2] [31].

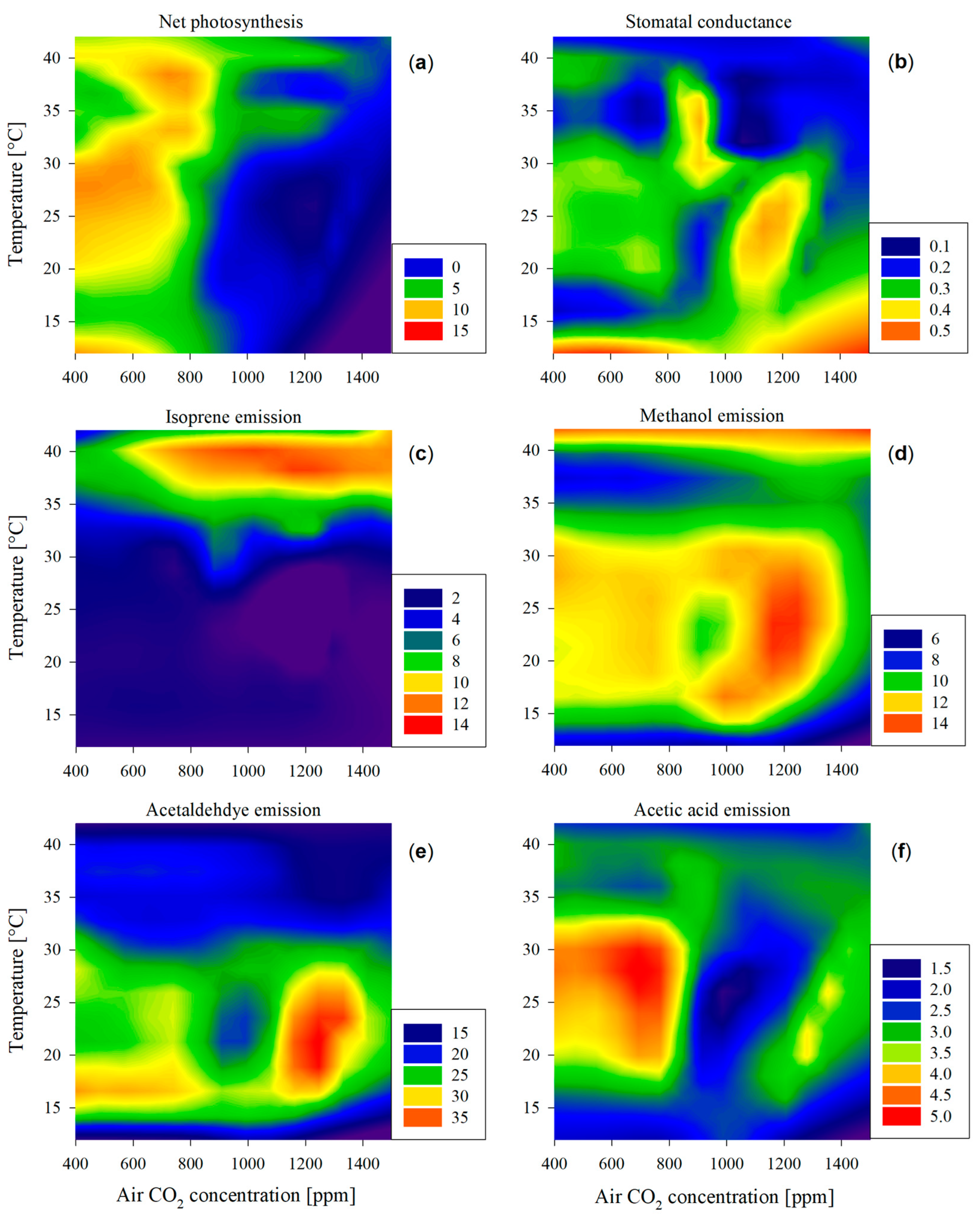
2.2. Controls of CO2 Availability and Temperature on Net Photosynthesis

2.3. Impairment of Carbon Assimilation and Isoprene Emission
2.4. Emission of Oxygenated Compounds under Different Temperatures and CO2 Levels
3. Materials and Methods
3.1. Plant Material
3.2. Analytical Setup
4. Conclusions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kessler, A.; Baldwin, I.T. Defensive Function of Herbivore-Induced Plant Volatile Emissions in Nature. Science 2001, 291, 2141–2144. [Google Scholar] [CrossRef] [PubMed]
- Heil, M.; Silva Bueno, J.C. Within-Plant Signaling by Volatiles Leads to Induction and Priming of an Indirect Plant Defense in Nature. Proc. Natl. Acad. Sci. USA 2007, 104, 5467–5472. [Google Scholar] [CrossRef] [PubMed]
- Hellén, H.; Tykkä, T.; Hakola, H. Importance of Monoterpenes and Isoprene in Urban Air in Northern Europe. Atmos. Environ. 2012, 59, 59–66. [Google Scholar] [CrossRef]
- Graus, M.; Schnitzler, J.P.; Hansel, A.; Cojocariu, C.; Rennenberg, H.; Wisthaler, A.; Kreuzwieser, J. Transient Release of Oxygenated Volatile Organic Compounds During Light-Dark Transitions in Grey Poplar Leaves. Plant Physiol. 2004, 135, 1967–1975. [Google Scholar] [CrossRef]
- García-Plazaola, J.I.; Portillo-Estrada, M.; Fernández-Marín, B.; Kännaste, A.; Niinemets, Ü. Niinemets. Emissions of Carotenoid Cleavage Products Upon Heat Shock and Mechanical Wounding from a Foliose Lichen. Environ. Exp. Bot. 2017, 133, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Harley, P.C.; Niinemets, Ü. Ozone-Induced Foliar Damage and Release of Stress Volatiles Is Highly Dependent on Stomatal Openness and Priming by Low-Level Ozone Exposure in Phaseolus vulgari. Plant Cell Environ. 2017, 40, 1984–2003. [Google Scholar] [CrossRef]
- Jardine, K.J.; Chambers, J.Q.; Holm, J.; Jardine, A.B.; Fontes, C.G.; Zorzanelli, R.F.; Meyers, K.T.; de Souza, V.F.; Garcia, S.; Gimenez, B.O.; et al. Green Leaf Volatile Emissions During High Temperature and Drought Stress in a Central Amazon Rainforest. Plants 2015, 4, 678–690. [Google Scholar] [CrossRef] [PubMed]
- Jardine, K.J.; Dewhirst, R.A.; Som, S.; Lei, J.; Tucker, E.; Young, R.P.; Portillo-Estrada, M.; Gao, Y.; Su, L.P.; Fares, S.; et al. Cell Wall Ester Modifications and Volatile Emission Signatures of Plant Response to Abiotic Stress. Plant Cell Environ. 2022, 45, 3429–3444. [Google Scholar] [CrossRef]
- Portillo-Estrada, M.; Niinemets, Ü. Massive Release of Volatile Organic Compounds Due to Leaf Midrib Wounding in Populus tremula. Plant Ecol. 2018, 219, 1021–1028. [Google Scholar] [CrossRef]
- Copolovici, L.; Väärtnõu, F.; Portillo Estrada, M.; Niinemets, Ü. Oak Powdery Mildew (Erysiphe alphitoides)-Induced Volatile Emissions Scale with the Degree of Infection in Quercus robur. Tree Physiol. 2014, 34, 1399–1410. [Google Scholar] [CrossRef]
- Brosset, A.; Blande, J.D. Volatile-Mediated Plant-Plant Interactions: Volatile Organic Compounds as Modulators of Receiver Plant Defence, Growth, and Reproduction. J. Exp. Bot. 2022, 73, 511–528. [Google Scholar] [CrossRef] [PubMed]
- Komarova, T.V.; Sheshukova, E.V.; Dorokhov, Y.L. Cell Wall Methanol as a Signal in Plant Immunity. Front. Plant Sci. 2014, 5, 101. [Google Scholar] [CrossRef] [PubMed]
- Midzi, J.; Jeffery, D.W.; Baumann, U.; Rogiers, S.; Tyerman, S.D.; Pagay, V. Stress-Induced Volatile Emissions and Signalling in Inter-Plant Communication. Plants 2022, 11, 2566. [Google Scholar] [CrossRef] [PubMed]
- Zhao, N.; Guan, J.; Ferrer, J.L.; Engle, N.; Chern, M.; Ronald, P.; Tschaplinski, T.J.; Chen, F. Biosynthesis and Emission of Insect-Induced Methyl Salicylate and Methyl Benzoate from Rice. Plant Physiol. Biochem. 2010, 48, 279–287. [Google Scholar] [CrossRef]
- Portillo-Estrada, M.; Ariza-Carricondo, C.; Ceulemans, R. Outburst of Senescence-Related Voc Emissions from a Bioenergy Poplar Plantation. Plant Physiol. Biochem. 2020, 148, 324–332. [Google Scholar] [CrossRef] [PubMed]
- Copolovici, L.; Kännaste, A.; Remmel, T.; Niinemets, Ü. Volatile Organic Compound Emissions from Alnus glutinosa under Interacting Drought and Herbivory Stresses. Environ. Exp. Bot. 2014, 100, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Holopainen, J.K.; Blande, J.D. Molecular Plant Volatile Communication. In Sensing in Nature; López-Larrea, C., Ed.; Springer: New York, NY, USA, 2012; pp. 17–31. [Google Scholar]
- Niinemets, Ü.; Reichstein, M. Controls on the Emission of Plant Volatiles through Stomata: Sensitivity or Insensitivity of the Emission Rates to Stomatal Closure Explained. J. Geophys. Res. Atmos. 2003, 108, 4208. [Google Scholar] [CrossRef]
- Thompson, M.; Gamage, D.; Hirotsu, N.; Martin, A.; Seneweera, S. Effects of Elevated Carbon Dioxide on Photosynthesis and Carbon Partitioning: A Perspective on Root Sugar Sensing and Hormonal Crosstalk. Front. Physiol. 2017, 8, 578. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.; Zhang, Y.X.; Guo, J.P. Effects of Drought Stress on Photosynthetic Physiological Characteristics, Leaf Microstructure, and Related Gene Expression of Yellow Horn. Plant Signal. Behav. 2023, 18, 2215025. [Google Scholar] [CrossRef]
- Sage, R.F.; Way, D.A.; Kubien, D.S. Rubisco, Rubisco Activase, and Global Climate Change. J. Exp. Bot. 2008, 59, 1581–1595. [Google Scholar] [CrossRef]
- Dusenge, M.E.; Duarte, A.G.; Way, D.A. Plant Carbon Metabolism and Climate Change: Elevated Co2 and Temperature Impacts on Photosynthesis, Photorespiration and Respiration. New Phytol. 2019, 221, 32–49. [Google Scholar] [CrossRef]
- Copolovici, L.; Niinemets, Ü. Environmental Impacts on Plant Volatile Emission. In Deciphering Chemical Language of Plant Communication; Blande, J.D., Glinwood, R., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 35–59. [Google Scholar]
- Pinto, D.M.; Blande, J.D.; Souza, S.R.; Nerg, A.-M.; Holopainen, J.K. Plant Volatile Organic Compounds (Vocs) in Ozone (O3) Polluted Atmospheres: The Ecological Effects. J. Chem. Ecol. 2010, 36, 22–34. [Google Scholar] [CrossRef]
- Cen, Y.P.; Sage, R.F. The Regulation of Rubisco Activity in Response to Variation in Temperature and Atmospheric Co2 Partial Pressure in Sweet Potato. Plant Physiol. 2005, 139, 979–990. [Google Scholar] [CrossRef]
- Niinemets, U. Variation in Leaf Photosynthetic Capacity within Plant Canopies: Optimization, Structural, and Physiological Constraints and Inefficiencies. Photosynth. Res. 2023, 158, 131–149. [Google Scholar] [CrossRef]
- Morfopoulos, C.; Sperlich, D.; Peñuelas, J.; Filella, I.; Llusià, J.; Medlyn, B.E.; Niinemets, Ü.; Possell, M.; Sun, Z.H.; Prentice, I.C. A Model of Plant Isoprene Emission Based on Available Reducing Power Captures Responses to Atmospheric CO2. New Phytol. 2014, 203, 125–139. [Google Scholar] [CrossRef] [PubMed]
- Sharkey, T.D.; Wiberley, A.E.; Donohue, A.R. Isoprene Emission from Plants: Why and How. Ann. Bot. 2008, 101, 5–18. [Google Scholar] [CrossRef] [PubMed]
- Rasulov, B.; Copolovici, L.; Laisk, A.; Niinemets, Ü. Postillumination Isoprene Emission: In Vivo Measurements of Dimethylallyldiphosphate Pool Size and Isoprene Synthase Kinetics in Aspen Leaves. Plant Physiol. 2009, 149, 1609–1618. [Google Scholar] [CrossRef] [PubMed]
- Rasulov, B.; Hüve, K.; Bichele, I.; Laisk, A.; Niinemets, Ü. Temperature Response of Isoprene Emission in Vivo Reflects a Combined Effect of Substrate Limitations and Isoprene Synthase Activity: A Kinetic Analysis. Plant Physiol. 2010, 154, 1558–1570. [Google Scholar] [CrossRef]
- Sahu, A.; Mostofa, M.G.; Weraduwage, S.M.; Sharkey, T.D. Hydroxymethylbutenyl Diphosphate Accumulation Reveals Mep Pathway Regulation for High Co2-Induced Suppression of Isoprene Emission. Proc. Natl. Acad. Sci. USA 2023, 120, e2309536120. [Google Scholar] [CrossRef]
- Zheng, Y.P.; Li, F.; Hao, L.H.; Yu, J.J.; Guo, L.L.; Zhou, H.R.; Ma, C.; Zhang, X.X.; Xu, M. Elevated Co2 Concentration Induces Photosynthetic Down-Regulation with Changes in Leaf Structure, Non-Structural Carbohydrates and Nitrogen Content of Soybean. BMC Plant Biol. 2019, 19, 255. [Google Scholar] [CrossRef]
- Cotrufo, M.F.; Ineson, P.; Scott, A. Elevated Co2 Reduces the Nitrogen Concentration of Plant Tissues. Glob. Chang. Biol. 1998, 4, 43–54. [Google Scholar] [CrossRef]
- Davey, P.A.; Olcer, H.; Zakhleniuk, O.; Bernacchi, C.J.; Calfapietra, C.; Long, S.P.; Raines, C.A. Can Fast-Growing Plantation Trees Escape Biochemical Down-Regulation of Photosynthesis When Grown Throughout Their Complete Production Cycle in the Open Air under Elevated Carbon Dioxide? Plant Cell Environ. 2006, 29, 1235–1244. [Google Scholar] [CrossRef]
- Benomar, L.; Moutaoufik, M.T.; Elferjani, R.; Isabel, N.; DesRochers, A.; El Guellab, A.; Khlifa, R.; Hassania, L.A.I. Thermal Acclimation of Photosynthetic Activity and Rubisco Content in Two Hybrid Poplar Clones. PLoS ONE 2019, 14, e0206021. [Google Scholar] [CrossRef]
- Silim, S.N.; Ryan, N.; Kubien, D.S. Temperature Responses of Photosynthesis and Respiration in Populus balsamifera L.: Acclimation Versus Adaptation. Photosynth. Res. 2010, 104, 19–30. [Google Scholar] [CrossRef] [PubMed]
- de Souza, V.F.; Niinemets, Ü.; Rasulov, B.; Vickers, C.E.; Duvoisin, S.; Araújo, W.L.; Gonçalves, J.F.D. Alternative Carbon Sources for Isoprene Emission. Trends Plant Sci. 2018, 23, 1081–1101. [Google Scholar] [CrossRef] [PubMed]
- Maeda, H.A. Harnessing Evolutionary Diversification of Primary Metabolism for Plant Synthetic Biology. J. Biol. Chem. 2019, 294, 16549–16566. [Google Scholar] [CrossRef] [PubMed]
- Velikova, V.; Sharkey, T.D.; Loreto, F. Stabilization of Thylakoid Membranes in Isoprene-Emitting Plants Reduces Formation of Reactive Oxygen Species. Plant Signal. Behav. 2012, 7, 139–141. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.Y.; Guenther, A.; Potosnak, M.; Geron, C.; Seco, R.; Karl, T.; Kim, S.; Gu, L.H.; Pallardy, S. Isoprene Emission Response to Drought and the Impact on Global Atmospheric Chemistry. Atmos. Environ. 2018, 183, 69–83. [Google Scholar] [CrossRef] [PubMed]
- Niinemets, Ü.; Reichstein, M.; Staudt, M.; Seufert, G.; Tenhunen, J.D. Stomatal Constraints May Affect Emission of Oxygenated Monoterpenoids from the Foliage of Pinus pinea. Plant Physiol. 2002, 130, 1371–1385. [Google Scholar] [CrossRef]
- Jiang, Y.F.; Ye, J.Y.; Rasulov, B.; Niinemets, Ü. Role of Stomatal Conductance in Modifying the Dose Response of Stress-Volatile Emissions in Methyl Jasmonate Treated Leaves of Cucumber (Cucumis sativa). Int. J. Mol. Sci. 2020, 21, 1018. [Google Scholar] [CrossRef]
- Hüve, K.; Christ, M.M.; Kleist, E.; Uerlings, R.; Niinemets, Ü.; Walter, A.; Wildt, J. Simultaneous Growth and Emission Measurements Demonstrate an Interactive Control of Methanol Release by Leaf Expansion and Stomata. J. Exp. Bot. 2007, 58, 1783–1793. [Google Scholar] [CrossRef] [PubMed]
- Karl, T.; Curtis, A.J.; Rosenstiel, T.N.; Monson, R.K.; Fall, R. Transient Releases of Acetaldehyde from Tree Leaves—Products of a Pyruvate Overflow Mechanism? Plant Cell Environ. 2002, 25, 1121–1131. [Google Scholar] [CrossRef]
- Kreuzwieser, J.; Scheerer, U.; Rennenberg, H. Metabolic Origin of Acetaldehyde Emitted by Poplar (Populus tremula × P. alba) Trees. J. Exp. Bot. 1999, 50, 757–765. [Google Scholar] [CrossRef]
- Portillo-Estrada, M.; Kazantsev, T.; Talts, E.; Tosens, T.; Niinemets, Ü. Emission Timetable and Quantitative Patterns of Wound-Induced Volatiles across Different Leaf Damage Treatments in Aspen (Populus tremula). J. Chem. Ecol. 2015, 41, 1105–1117. [Google Scholar] [CrossRef] [PubMed]
- Turan, S.; Kask, K.; Kanagendran, A.; Li, S.; Anni, R.; Talts, E.; Rasulov, B.; Kännaste, A.; Niinemets, Ü. Lethal Heat Stress-Dependent Volatile Emissions from Tobacco Leaves: What Happens Beyond the Thermal Edge? J. Exp. Bot. 2019, 70, 5017–5030. [Google Scholar] [CrossRef] [PubMed]
- Portillo-Estrada, M.; Zenone, T.; Arriga, N.; Ceulemans, R. Contribution of Volatile Organic Compound Fluxes to the Ecosystem Carbon Budget of a Poplar Short-Rotation Plantation. Glob. Chang. Biol. Bioenergy 2018, 10, 405–414. [Google Scholar] [CrossRef] [PubMed]
- Rasulov, B.; Talts, E.; Niinemets, Ü. Spectacular Oscillations in Plant Isoprene Emission under Transient Conditions Explain the Enigmatic Co2 Response. Plant Physiol. 2016, 172, 2275–2285. [Google Scholar] [CrossRef] [PubMed]
- Portillo-Estrada, M.; Kazantsev, T.; Niinemets, Ü. Fading of Wound-Induced Volatile Release during Populus tremula Leaf Expansion. J. Plant Res. 2017, 130, 157–165. [Google Scholar] [CrossRef]
- Cappellin, L.; Karl, T.; Probst, M.; Ismailova, O.; Winkler, P.M.; Soukoulis, C.; Aprea, E.; Mark, T.D.; Gasperi, F.; Biasioli, F. On Quantitative Determination of Volatile Organic Compound Concentrations Using Proton Transfer Reaction Time-of-Flight Mass Spectrometry. Environ. Sci. Technol. 2012, 46, 2283–2290. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Portillo-Estrada, M. Limitations of Plant Stress Tolerance upon Heat and CO2 Exposure in Black Poplar: Assessment of Photosynthetic Traits and Stress Volatile Emissions. Plants 2024, 13, 1165. https://doi.org/10.3390/plants13081165
Portillo-Estrada M. Limitations of Plant Stress Tolerance upon Heat and CO2 Exposure in Black Poplar: Assessment of Photosynthetic Traits and Stress Volatile Emissions. Plants. 2024; 13(8):1165. https://doi.org/10.3390/plants13081165
Chicago/Turabian StylePortillo-Estrada, Miguel. 2024. "Limitations of Plant Stress Tolerance upon Heat and CO2 Exposure in Black Poplar: Assessment of Photosynthetic Traits and Stress Volatile Emissions" Plants 13, no. 8: 1165. https://doi.org/10.3390/plants13081165
APA StylePortillo-Estrada, M. (2024). Limitations of Plant Stress Tolerance upon Heat and CO2 Exposure in Black Poplar: Assessment of Photosynthetic Traits and Stress Volatile Emissions. Plants, 13(8), 1165. https://doi.org/10.3390/plants13081165