Improved Production of Industrially Important Essential Oils Through Elicitation in the Adventitious Roots of Artemisia amygdalina


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material, Sterilization and Explant Preparation
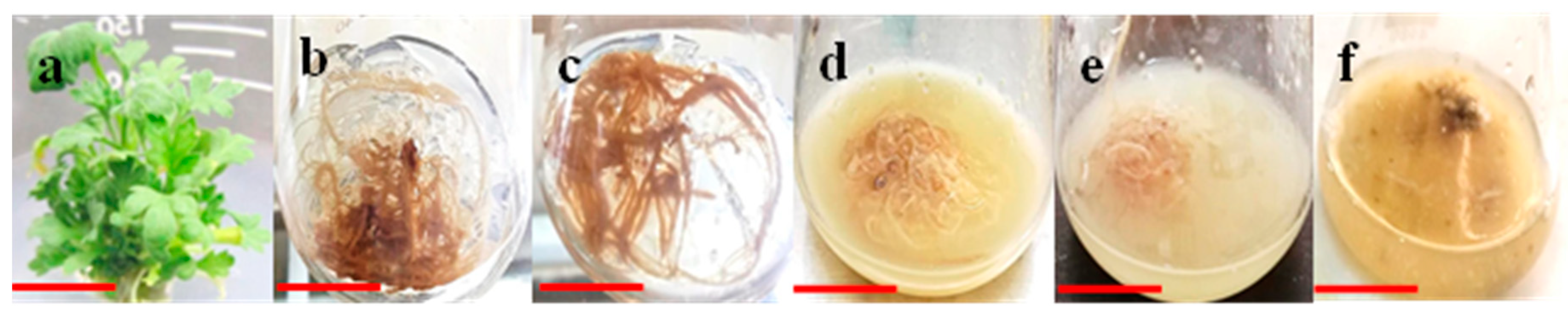
2.2. Establishment of Adventitious Roots (AR) Cultures on Solid Media
2.3. Transient Elicitation of AR and Cultivation in Shake Flask Liquid Media
2.4. Phytochemical Analysis for the Assessment of Effects of Elicitation on the Production of Secondary Metabolites in the In Vitro Raised AR
2.4.1. Determination of Total Phenolic Content (TPC)
2.4.2. Determination of Total Flavonoid Content (TFC)
2.4.3. Phenylalanine Ammonia Lyase (PAL) Activity
2.4.4. DPPH Free Radical Scavenging Assay
2.5. Quantitative Analysis for the Assessment of Effects of Elicitation on the Production of Essential Oils in the In Vitro Raised AR Using Gas Chromatography-Mass Spectrometry (GC–MS)
2.6. Statistical Analysis
3. Results and Discussion
3.1. Effects of Auxins on Induction and Growth of Adventitious Rooting in Explants
3.2. Effects of Varying Levels of Sucrose on Induction and Growth of Adventitious Rooting in Explants
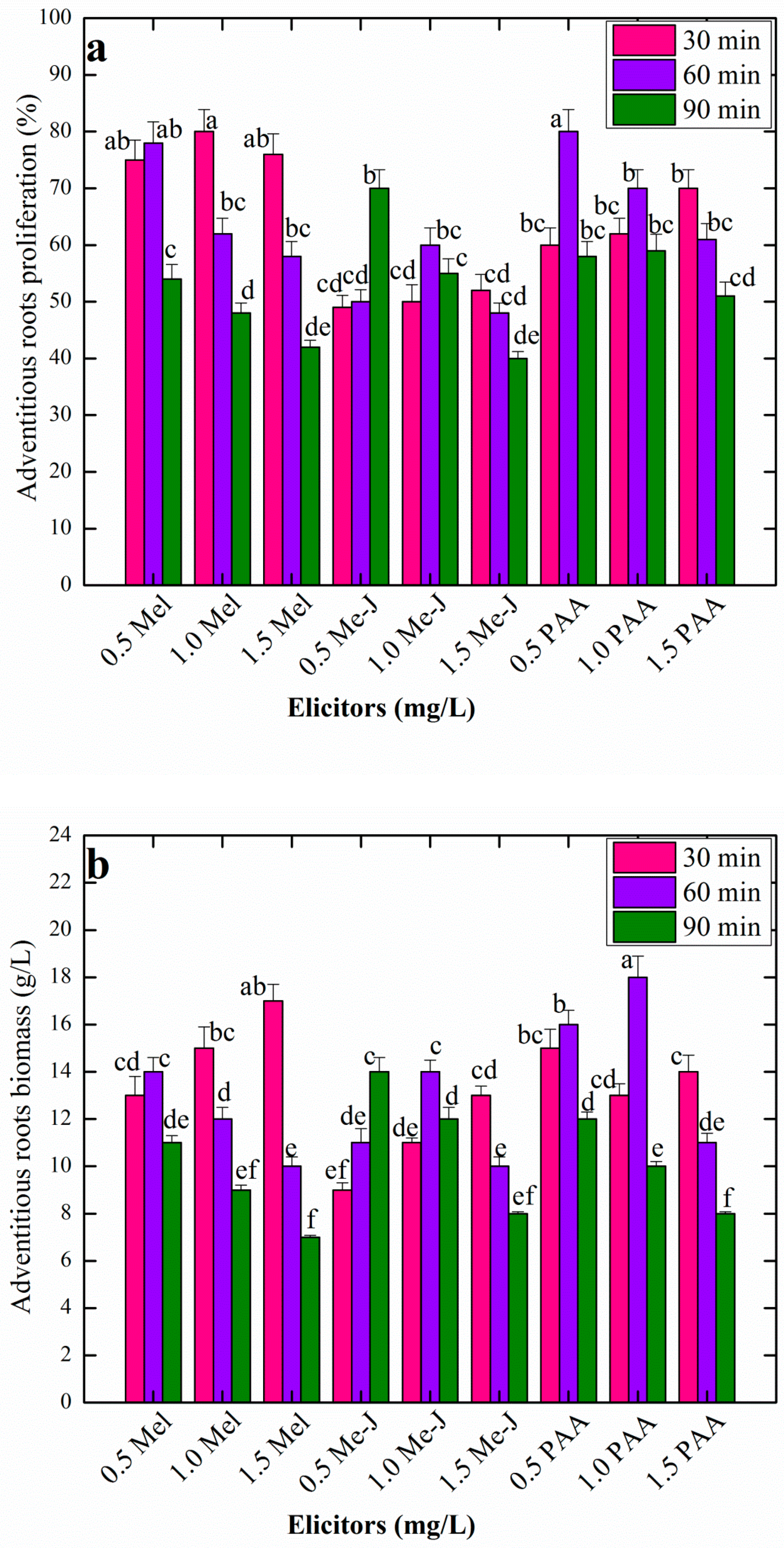
3.3. Effects of AR Pretreatment With Varying Levels of Elicitors at Different Time Periods on AR Growth in Liquid Culture Media
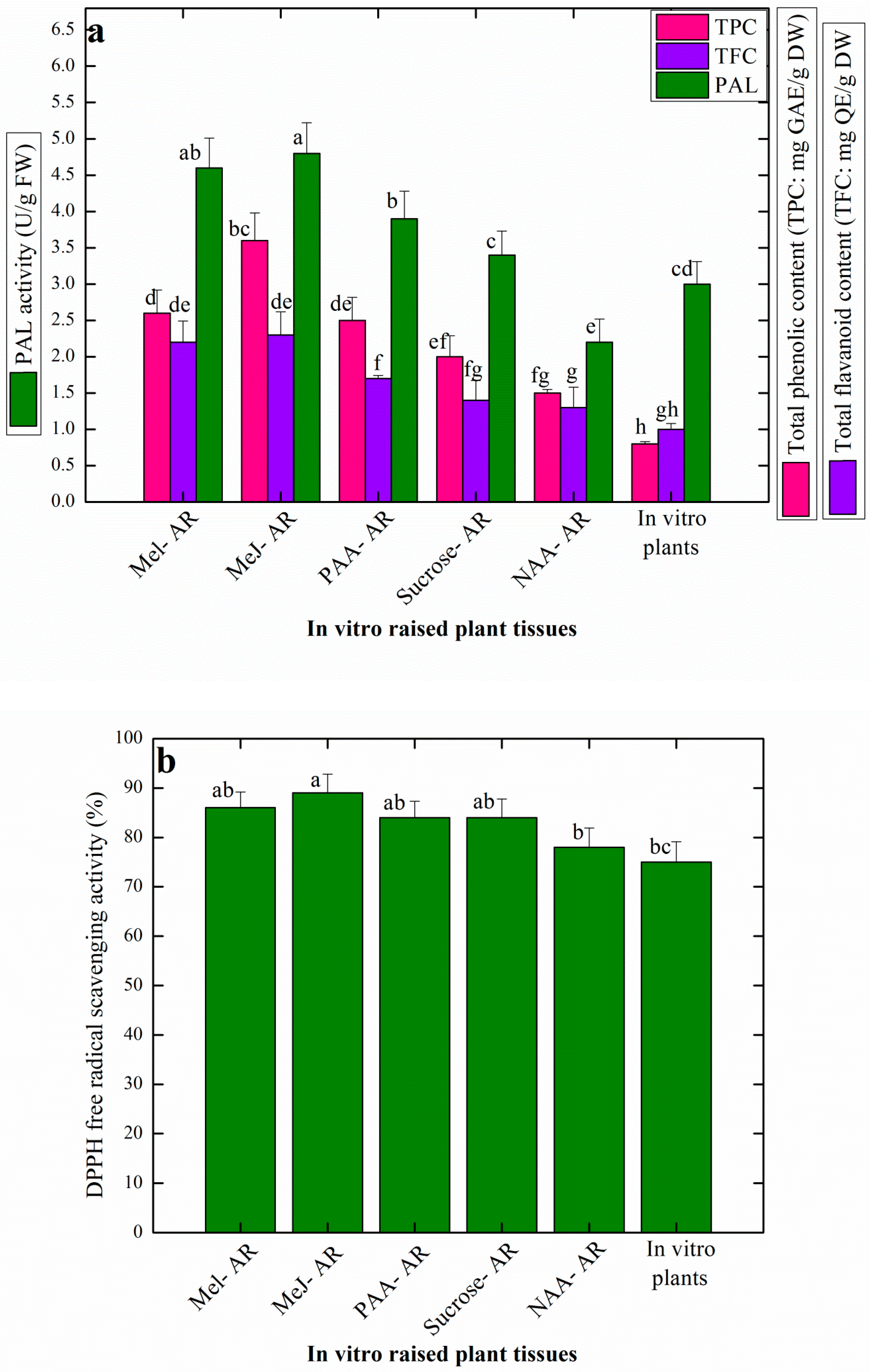
3.4. Effects of Elicitation on Antioxidant Profile of AR Raised in Vitro
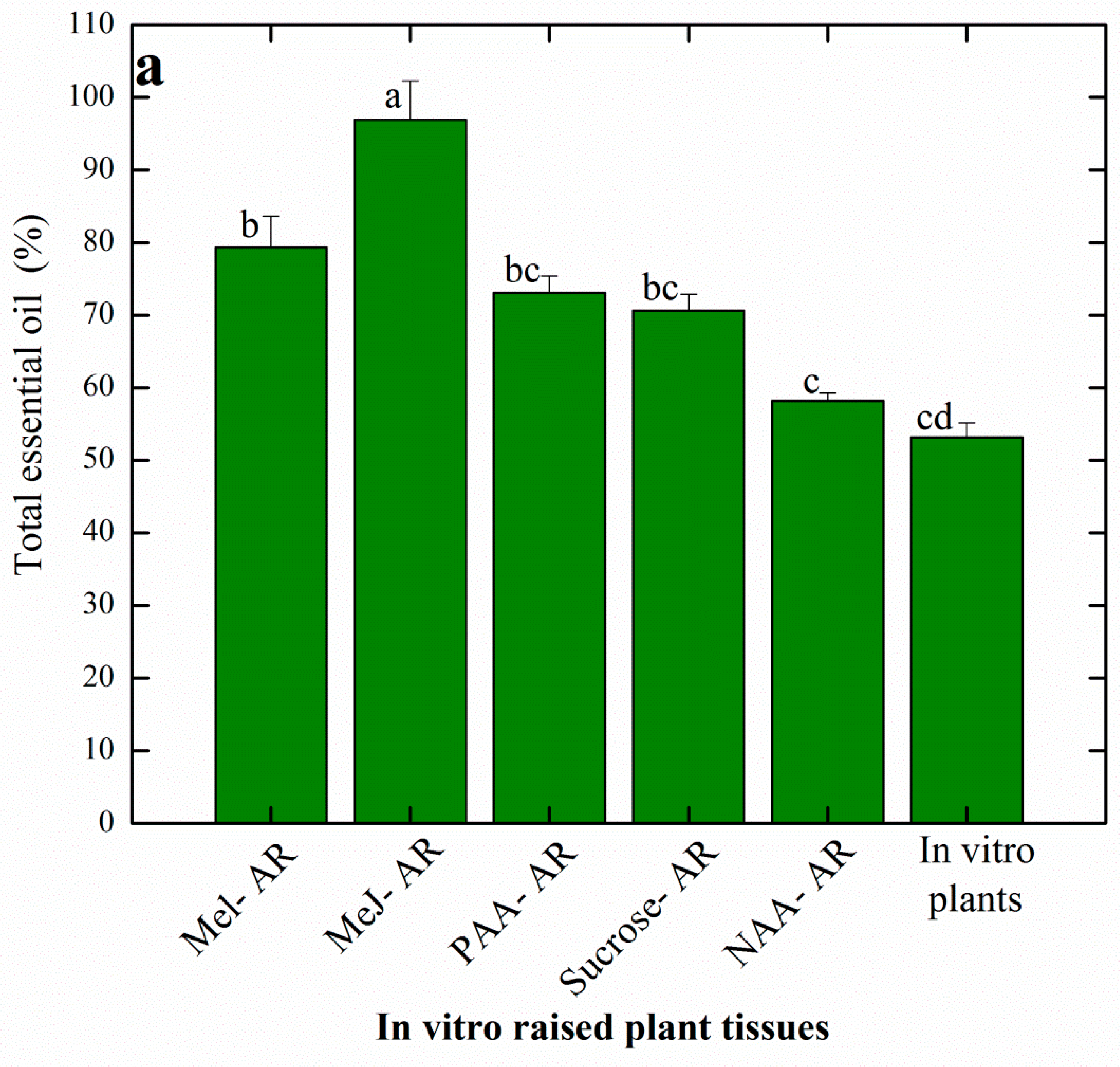
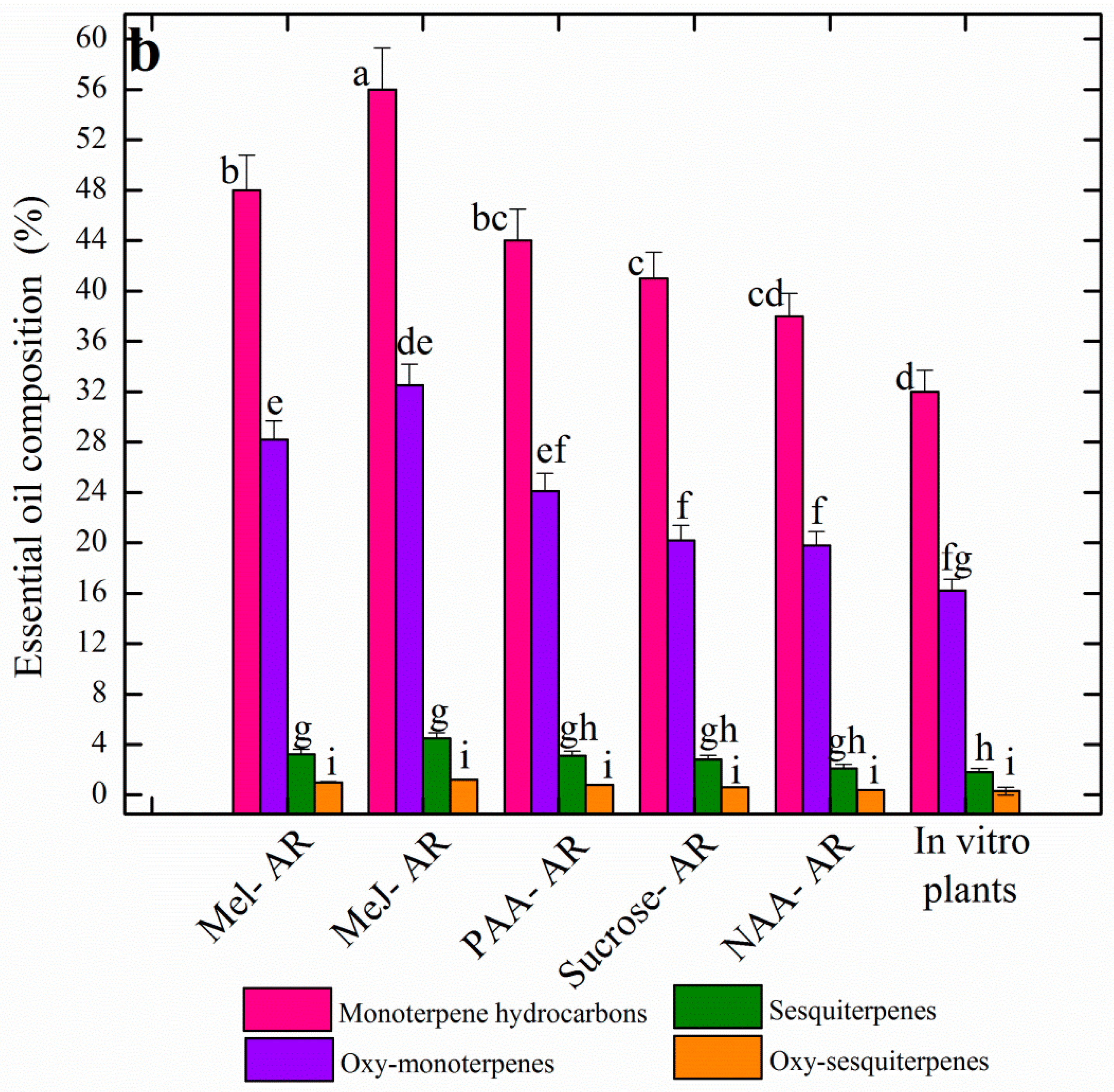
3.5. Effects of Elicitation on Essential Oil Profiles of AR Raised in Vitro
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Edwards, S.; Da-Costa-Rocha, I.; Lawrence, M.J.; Cable, C.; Heinrich, M. Use and efficacy of herbal medicines: Part 1—Historical and traditional use. Pharm. J. 2012, 289, 161–162. [Google Scholar]
- Kazmi, A.; Khan, M.A.; Ali, H. Biotechnological approaches for production of bioactive secondary metabolites in Nigella sativa: An up-to-date review. Int. J. Second. Metab. 2019, 6, 172–195. [Google Scholar] [CrossRef]
- Inoue, M.; Hayashi, S.; Craker, L.E. Role of Medicinal and Aromatic Plants: Past, Present, and Future. In Pharmacognosy-Medicinal Plants; IntechOpen: London, UK, 2019. [Google Scholar] [Green Version]
- Bora, K.S.; Sharma, A. Neuroprotective effect of Artemisia absinthium L. on focal ischemia and reperfusion-induced cerebral injury. J. Ethnopharmacol. 2010, 129, 403–409. [Google Scholar] [CrossRef] [PubMed]
- Dar, A.; Dar, G.H.; Reshi, Z. Conservation of Artemisia amygdalina—A critically endangered endemic plant species of Kashmir Himalaya. Endanger. Species Update 2006, 23, 34–40. [Google Scholar]
- Ashraf, M.; Hayat, M.Q.; Jabeen, S.; Shaheen, N.; Khan, M.A.; Yasmin, G. Artemisia, L. species recognized by the local community of the northern areas of Pakistan as folk therapeutic plants. J. Med. Plants Res. 2010, 4, 112–119. [Google Scholar]
- Mubashir, K.; Ganai, B.A.; Ghazanfar, K.; Akbar, S.; Malik, A.H.; Masood, A. Evaluation of Artemisia amygdalina D. for anti-inflammatory and immunomodulatory potential. ISRN Inflamm. 2013, 2013. [Google Scholar] [CrossRef]
- Hayat, M.Q.; Ashraf, M.; Khan, M.A.; Yasmin, G.; Shaheen, N.; Jabeen, S. Diversity of foliar trichomes and their systematic implications in the genus Artemisia (Asteraceae). Int. J. Agric. Biol. 2009, 11, 542–546. [Google Scholar]
- Rasool, R.; Ganai, B.A.; Kamili, A.N.; Akbar, S. Antioxidant potential in callus culture of Artemisia amygdalina Decne. Nat. Prod. Res. 2012, 26, 2103–2106. [Google Scholar]
- Zeb, S.; Ali, A.; Zaman, W.; Zeb, S.; Ali, S.; Ullah, F.; Shakoor, A. Pharmacology, Taxonomy and Phytochemistry of the genus Artemisia specifically from Pakistan: A comprehensive review. Pharmaceutic. Biomed. Res. 2019, 4. [Google Scholar] [CrossRef]
- Lone, S.H.; Bhat, K.A.; Naseer, S.; Rather, R.A.; Khuroo, M.A.; Tasduq, S.A. Isolation, cytotoxicity evaluation and HPLC-quantification of the chemical constituents from Artemisia amygdalina Decne. J. Chromatogr. B 2013, 940, 135–141. [Google Scholar] [CrossRef]
- Ghazanfar, K.; Ganai, B.A.; Akbar, S.; Mubashir, K.; Dar, S.A.; Dar, M.Y.; Tantry, M.A. Antidiabetic activity of Artemisia amygdalina Decne in streptozotocin induced diabetic rats. BioMed Res. Int. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Rather, M.A.; Ganai, B.A.; Kamili, A.N.; Qayoom, M.; Akbar, S.; Masood, A.; Rasool, R.; Wani, S.H.; Qurishi, M.A. Comparative GC–FID and GC–MS analysis of the mono and sesquiterpene secondary metabolites produced by the field grown and micropropagated plants of Artemisia amygdalina Decne. Acta Physiologiae Plantarum 2012, 34, 885–890. [Google Scholar] [CrossRef]
- Khanam, M.N.; Anis, M.; Ahmad, S. Establishment of adventitious root cultures of Allamanda cathartica L. for the production of iridoid glycosides and its identification using HPTLC MS. Ind. Crops Prod. 2018, 125, 198–206. [Google Scholar] [CrossRef]
- Khan, M.A.; Khan, T.; Riaz, M.S.; Ullah, N.; Ali, H.; Nadhman, A. Plant cell nanomaterials interaction: Growth, physiology and secondary metabolism. Compr. Anal. Chem. 2019, 84, 23–54. [Google Scholar]
- Khan, M.A.; Abbasi, B.H.; Shah, N.A.; Yücesan, B.; Ali, H. Analysis of metabolic variations throughout growth and development of adventitious roots in Silybum marianum L. (Milk thistle), a medicinal plant. Plant Cell Tissue Organ Cult. PCTOC 2015, 123, 501–510. [Google Scholar] [CrossRef]
- Ali, H.; Khan, M.A.; Kayani, W.K.; Dilshad, E.; Rani, R.; Khan, R.S. Production of biomass and medicinal metabolites through adventitious roots in Ajuga bracteosa under different spectral lights. J. Photochem. Photobiol. B Biol. 2019, 193, 109–117. [Google Scholar] [CrossRef]
- Murthy, H.N.; Hahn, E.J.; Paek, K.Y. Adventitious roots and secondary metabolism. Chin. J. Biotechnol. 2008, 24, 711–716. [Google Scholar] [CrossRef]
- Ho, T.-T.; Lee, J.-D.; Jeong, C.-S.; Paek, K.-Y.; Park, S.-Y. Improvement of biosynthesis and accumulation of bioactive compounds by elicitation in adventitious root cultures of Polygonum multiflorum. Appl. Microbiol. Biotechnol. 2018, 102, 199–209. [Google Scholar] [CrossRef]
- Lee, E.J.; Park, S.Y.; Paek, K.Y. Enhancement strategies of bioactive compound production in adventitious root cultures of Eleutherococcus koreanum Nakai subjected to methyl jasmonate and salicylic acid elicitation through airlift bioreactors. Plant Cell Tissue Organ Cult. PCTOC 2015, 120, 1–10. [Google Scholar] [CrossRef]
- Khan, T.; Abbasi, B.H.; Khan, M.A.; Azeem, M. Production of biomass and useful compounds through elicitation in adventitious root cultures of Fagonia indica. Ind. Crops Prod. 2017, 108, 451–457. [Google Scholar] [CrossRef]
- Han, L.; Piao, X.; Jiang, J.; Jiang, X.; Yin, C.; Lian, M. A high production of flavonoids and anthraquinones via adventitious root culture of Oplopanax elatus and evaluating antioxidant activity. Plant Cell Tissue Organ Cult. PCTOC 2019, 137, 173–179. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiologia Plantarum 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Velioglu, Y.; Mazza, G.; Gao, L.; Oomah, B. Antioxidant activity and total phenolics in selected fruits, vegetables, and grain products. J. Agricult. Food Chem. 1998, 46, 44113–44117. [Google Scholar] [CrossRef]
- Chang, C.-C.; Yang, M.-H.; Wen, H.-M.; Chern, J.-C. Estimation of total flavonoid content in propolis by two complementary colorimetric methods. J. Food Drug Anal. 2002, 10, 178–182. [Google Scholar]
- Khan, M.A.; Abbasi, B.H.; Ahmed, N.; Ali, H. Effects of light regimes on in vitro seed germination and silymarin content in Silybum marianum. Ind. Crops Prod. 2013, 46, 105–110. [Google Scholar] [CrossRef]
- Abbasi, B.H.; Khan, M.A.; Mahmood, T.; Ahmad, M.; Chaudhary, M.F.; Khan, M.A. Shoot regeneration and free-radical scavenging activity in Silybum marianum L. Plant Cell Tissue Organ Cult. PCTOC 2010, 101, 371–376. [Google Scholar] [CrossRef]
- Davies, N. Gas chromatographic retention indices of monoterpenes and sesquiterpenes on methyl silicon and Carbowax 20M phases. J. Chromatogr. A 1990, 503, 1–24. [Google Scholar] [CrossRef]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry; Allured Publishing Corporation: Carol Stream, IL, USA, 2007. [Google Scholar]
- Khan, M.A.; Khan, T.; Ali, H. Plant Cell Culture Strategies for the Production of Terpenes as Green Solvents. In Industrial Applications of Green Solvents; Materials Research Forum: Millersville, PA, USA, 2019; Volume I. [Google Scholar]
- Kazmi, A.; Khan, M.A.; Mohammad, S.; Ali, A.; Kamil, A.; Arif, M.; Ali, H. Elicitation directed growth and production of steviol glycosides in the adventitious roots of Stevia rebaudiana Bertoni. Ind. Crops Prod. 2019, 139, 111530. [Google Scholar] [CrossRef]
- Khan, A.; Khan, M.A.; Alam, M.; Akbar, R.; Ali, A.; Mohammad, S.; Naeem, I.; Rauf, M. Analysis of the differential effects of methyl jasmonate on induction of adventitious roots and antioxidant potential in Artimisia scoparia. Int. J. Biosci. 2019. [Google Scholar] [CrossRef]
- Sujatha, G.; Ranjitha Kumari, B. Establishment of fast growing in vitro root culture system in Artemisia vulgaris. J. Agric. Technol. 2012, 8, 1779–1790. [Google Scholar]
- Hussein, S.; Ibrahim, R. Plant growth regulator eEffect on adventitious roots induction of labisia pumila. Malays. J. Fundam. Appl. Sci. 2014, 10, 49–52. [Google Scholar]
- Ling, A.P.K.; Kok, K.M.; Hussein, S.; Ong, S.L. Effects of plant growth regulators on adventitious roots induction from different explants of Orthosiphon stamineus. Am. Eurasian J. Sustain. Agric. 2009, 3, 493–501. [Google Scholar]
- Yan, Y.-H.; Li, J.-L.; Zhang, X.-Q.; Yang, W.-Y.; Wan, Y.; Ma, Y.-M.; Zhu, Y.-Q.; Peng, Y.; Huang, L.-K. Effect of naphthalene acetic acid on adventitious root development and associated physiological changes in stem cutting of Hemarthria compressa. PLoS ONE 2014, 9, e90700. [Google Scholar] [CrossRef] [PubMed]
- Baque, M.A.; Elgirban, A.; Lee, E.-J.; Paek, K.-Y. Sucrose regulated enhanced induction of anthraquinone, phenolics, flavonoids biosynthesis and activities of antioxidant enzymes in adventitious root suspension cultures of Morinda citrifolia (L.). Acta Physiologiae Plantarum 2012, 34, 405–415. [Google Scholar] [CrossRef]
- Mohammad, S.; Khan, M.A.; Ali, A.; Khan, L.; Khan, M.S. Feasible production of biomass and natural antioxidants through callus cultures in response to varying light intensities in olive (Olea europaea. L.) cult. Arbosana. J. Photochem. Photobiol. B Biol. 2019, 193, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Yousaf, R.; Khan, M.A.; Ullah, N.; Khan, I.; Hayat, O.; Shehzad, M.A.; Khan, I.; Taj, F.; Ud Din, N.; Khan, A. Biosynthesis of anti-leishmanial natural products in callus cultures of Artemisia scoparia. Artif. Cells Nanomed. Biotechnol. 2019, 47, 1122–1131. [Google Scholar] [CrossRef]
- Taylor, J.; Van Staden, J. The effect of nitrogen and sucrose concentrations on the growth of Eucomis autumnalis (Mill.) Chitt. plantlets in vitro, and on subsequent anti-inflammatory activity in extracts prepared from the plantlets. Plant Growth Regul. 2001, 34, 49–56. [Google Scholar] [CrossRef]
- Saiman, M.Z.; Mustafa, N.R.; Schulte, A.E.; Verpoorte, R.; Choi, Y.H. Induction, characterization, and NMR-based metabolic profiling of adventitious root cultures from leaf explants of Gynura procumbens. Plant Cell Tissue Organ Cult. PCTOC 2012, 109, 465–475. [Google Scholar] [CrossRef]
- Cui, X.-H.; Murthy, H.N.; Wu, C.-H.; Paek, K.-Y. Sucrose-induced osmotic stress affects biomass, metabolite, and antioxidant levels in root suspension cultures of Hypericum perforatum L. Plant Cell Tissue Organ Cult. PCTOC 2010, 103, 7–14. [Google Scholar] [CrossRef]
- Kim, J.H.; Chang, E.J.; Oh, H.-I. Saponin production in submerged adventitious root culture of Panax ginseng as affected by culture conditions and elicitors. Asia Pacific J. Mol. Biol. Biotechnol. 2005, 13, 87–91. [Google Scholar]
- Saeed, S.; Ali, H.; Khan, T.; Kayani, W.; Khan, M.A. Impacts of methyl jasmonate and phenyl acetic acid on biomass accumulation and antioxidant potential in adventitious roots of Ajuga bracteosa Wall ex Benth., a high valued endangered medicinal plant. Physiol. Mol. Biol. Plants 2017, 23, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Hanaka, A.; Maksymiec, W.; Bednarek, W. The effect of methyl jasmonate on selected physiological parameters of copper-treated Phaseolus coccineus plants. Plant Growth Regul. 2015, 77, 167–177. [Google Scholar] [CrossRef]
- Mbiyu, M.; Muthoni, J.; Kabira, J.; Muchira, C.; Pwaipwai, P.; Ngaruiya, J.; Onditi, J.; Otieno, S. Comparing liquid and solid media on the growth of plantlets from three Kenyan potato cultivars. Am. J. Exp. Agric. 2012, 2, 81. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernández-Ruiz, J. Melatonin promotes adventitious-and lateral root regeneration in etiolated hypocotyls of Lupinus albus L. J. Pineal Res. 2007, 42, 147–152. [Google Scholar] [CrossRef]
- Murch, S.J.; Saxena, P.K. Melatonin: A potential regulator of plant growth and development? In Vitro Cell. Dev. Biol. Plant 2002, 38, 531–536. [Google Scholar] [CrossRef]
- Pelagio-Flores, R.; Muñoz-Parra, E.; Ortiz-Castro, R.; López-Bucio, J. Melatonin regulates Arabidopsis root system architecture likely acting independently of auxin signaling. J. Pineal Res. 2012, 53, 279–288. [Google Scholar] [CrossRef]
- Ali, M.; Abbasi, B.H.; Ali, G.S. Elicitation of antioxidant secondary metabolites with jasmonates and gibberellic acid in cell suspension cultures of Artemisia absinthium L. Plant Cell Tissue Organ Cult. PCTOC 2015, 120, 1099–1106. [Google Scholar] [CrossRef]
- Ali, M.; Abbasi, B.H.; Ahmad, N.; Ali, S.S.; Ali, S.; Ali, G.S. Sucrose-enhanced biosynthesis of medicinally important antioxidant secondary metabolites in cell suspension cultures of Artemisia absinthium L. Bioprocess Biosyst. Eng. 2016, 39, 1945–1954. [Google Scholar] [CrossRef]
- Ali, A.; Mohammad, S.; Khan, M.A.; Raja, N.I.; Arif, M.; Kamil, A.; Mashwani, Z.U.R. Silver nanoparticles elicited in vitro callus cultures for accumulation of biomass and secondary metabolites in Caralluma tuberculate. Artif. Cells Nanomed. Biotechnol. 2019, 47, 715–724. [Google Scholar] [CrossRef]
- Mir, M.Y.; Kamili, A.N.; Hassan, Q.P.; Tyub, S. Effect of Light and Dark Conditions on Biomass Accumulation and Secondary Metabolite Production in Suspension Cultures of Artemisia amygdalina Decne. J. Himal. Ecol. Sustain. Dev. 2017, 12, 1–6. [Google Scholar]
- Ali, H.; Khan, M.A.; Ullah, N.; Khan, R.S. Impacts of hormonal elicitors and photoperiod regimes on elicitation of bioactive secondary volatiles in cell cultures of Ajuga bracteosa. J. Photochem. Photobiol. B Biol. 2018, 183, 242–250. [Google Scholar] [CrossRef] [PubMed]
- Açikgöz, M.A.; Kara, Ş.M.; Aygün, A.; Özcan, M.M.; Ay, E.B. Effects of methyl jasmonate and salicylic acid on the production of camphor and phenolic compounds in cell suspension culture of endemic Turkish yarrow (Achillea gypsicola) species. Turk. J. Agric. For. 2019, 43, 351–359. [Google Scholar] [CrossRef]
- Farzi, F.; Fahimi, H.; Sharifi, M.; Mousavi, A. The Effect of Methyl Jasmonate on Monoterpene Composition of the Essential Oil in Shoot Cultures of Sautreja khuzistanica Jamzad. J. Pharm. Res. Int. 2018, 24, 1–7. [Google Scholar] [CrossRef]
- Santos, F.; Rao, V. 1,8-cineol, a food flavoring agent, prevents ethanol-induced gastric injury in rats. Dig. Dis. Sci. 2001, 46, 331–337. [Google Scholar] [CrossRef]
- Kumar, M.; Ando, Y. Single-wall and multi-wall carbon nanotubes from camphor—A botanical hydrocarbon. Diam. Relat. Mater. 2003, 12, 1845–1850. [Google Scholar] [CrossRef]
- Orhan, I.; Küpeli, E.; Aslan, M.; Kartal, M.; Yesilada, E. Bioassay-guided evaluation of anti-inflammatory and antinociceptive activities of pistachio, Pistacia vera L. J. Ethnopharmacol. 2006, 105, 235–240. [Google Scholar] [CrossRef]
- Türkez, H.; Aydın, E. In vitro assessment of cytogenetic and oxidative effects of α-pinene. Toxicology Ind. Health 2016, 32, 168–176. [Google Scholar] [CrossRef]
- Dai, J.P.; Chen, J.; Bei, Y.F.; Han, B.X.; Wang, S. Influence of borneol on primary mice oral fibroblasts: A penetration enhancer may be used in oral submucous fibrosis. J. Oral Pathol. Med. 2009, 38, 276–281. [Google Scholar] [CrossRef]
- Kalemba, D.; Kunicka, A. Antibacterial and antifungal properties of essential oils. Curr. Med. Chem. 2003, 10, 813–829. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Auxins | Concentration (mg/L) | Percent Root Induction | Number of Days to Root Initiation | Mean Root Number | Mean root Length (cm) | Fresh Biomass (g/L) |
|---|---|---|---|---|---|---|
| Control | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | |
| IBA | 0.5 | 62 ± 2.67 d | 5 | 2.6 ± 0.39 c | 1.15 ± 0.11 cd | 9.5 ± 2.6 b |
| 1.0 | 76 ± 3.78 b | 5 | 4.1 ± 0.45 ab | 2.3 ± 0.12 bc | 12.3 ± 3.1 ab | |
| 1.5 | 67 ± 2.83 c | 5 | 1.7 ± 0.3 cd | 1.12 ± 0.13 c | 9.2 ± 2.7 b | |
| NAA | 0.5 | 78 ± 3.84 ab | 4 | 3.8 ± 0.43 b | 2.06 ± 0.05 b | 12.3 ± 3.1 ab |
| 1.0 | 87 ± 4.25 a | 4 | 6.8 ± 0.48 a | 3.2 ± 0.08 a | 15.7 ± 3.5 a | |
| 1.5 | 71 ± 3.72 bc | 4 | 2.9 ± 0.45 bc | 1.3 ± 0.08 cd | 10.2 ± 2.8 bc | |
| IAA | 0.5 | – | – | – | – | – |
| 1.0 | – | – | – | – | – | |
| 1.5 | – | – | – | – | – |
| Sucrose (w/v) | Percent Root Induction | Number of Days to Root Initiation | Mean Root Number | Mean Root Length (cm) | Fresh Biomass (g/L) |
|---|---|---|---|---|---|
| 0 | 0.0 | 0.0 | 0.0 | 0.0 | |
| 1 | 53 ± 2.67 c | 5 | 3.6 ± 0.39 d | 2.15 ± 0.11 c | 10.5 ± 2.6 c |
| 2 | 65 ± 3.78 bc | 5 | 4.8 ± 0.45 c | 2.80 ± 0.12 bc | 12.3 ± 3.1 bc |
| 3 | 87 ± 4.25 ab | 4 | 6.8 ± 0.48 b | 3.2 ± 0.08 b | 15.7 ± 3.5 ab |
| 4 | 90 ± 4.25 a | 3 | 7.8 ± 0.48 a | 4.20 ± 0.08 a | 17.7 ± 3.5 a |
| 5 | 71 ± 3.72 b | 3 | 4.1 ± 0.43 c | 2.06 ± 0.05 c | 13.3 ± 3.1 b |
| 6 | 59 ± 2.67 c | 4 | 3.9 ± 0.45 cd | 1.80 ± 0.08 d | 11.2 ± 2.8 c |
| S.no | Compound | Class | Molecular Formula | Mel-AR (%) | Me-J-AR (%) | PAA-AR (%) | Sucrose-AR (%) | NAA-AR (%) | In vitro Plants |
|---|---|---|---|---|---|---|---|---|---|
| 1 | α-Thujene | Monoterpene hydrocarbons | C10H16 | 4.71 | 6.8 | 4.2 | 3.2 | 3 | 2.5 |
| 2 | α-Pinene | Monoterpene hydrocarbons | C10H16 | 7.2 | 8.3 | 6.3 | 5.2 | 4.8 | 2.8 |
| 3 | δ-Camphene | Monoterpene hydrocarbons | C10H16 | 4.4 | 5.5 | 4.2 | 4.0 | 3.8 | 3 |
| 4 | Camphene | Monoterpene hydrocarbons | C10H16 | 5.12 | 6.4 | 4.1 | 3.5 | 3.2 | 2.5 |
| 5 | β-Pinen | Monoterpene hydrocarbons | C10H16 | 4.21 | 6.5 | 3.2 | 2.8 | 2.5 | 12.8 |
| 6 | α–phellandrene | Monoterpene hydrocarbons | C10H16 | 5.15 | 6.1 | 4.2 | 3.8 | 3.6 | 3 |
| 7 | 1,8-Cineole | Oxygenated monoterpenes | C10H18O | 12.4 | 16.2 | 8.14 | 5.36 | 4.8 | 4.2 |
| 8 | γ-Terpinene | Monoterpene hydrocarbons | C10H16 | 5.37 | 6.2 | 4.2 | 3.4 | 3 | 2.4 |
| 9 | Camphor | Oxygenated monoterpenes | C10H16 | 7.1 | 8.4 | 5.4 | 4.5 | 3.5 | 1.0 |
| 10 | L-Borneol | Oxygenated monoterpenes | C10H18O | 6.5 | trace | 9.0 | 6.8 | 6.1 | 9.25 |
| 11 | α-Terpineol | Monoterpene hydrocarbons | C10H18O | 3.84 | 2.9 | 6.7 | 5.4 | trace | 5.23 |
| 12 | β-Elemene | Sesquiterpene hydrocarbons | C15H24 | - | 4.5 | - | 0.85 | 1.1 | 0.5 |
| 13 | Piperitone | Monoterpene hydrocarbons | C10H16O | trace | 2.1 | 1.9 | 10.6 | 8.5 | 2 |
| 14 | Valencene | Oxygenated sesquiterpnes | C15H24 | trace | trace | trace | trace | 0.4 | 0.3 |
| 15 | Verbenol | Oxygenated monoterpenes | C10H16O | 5.1 | 10.2 | 4.2 | 3.5 | trace | 1.2 |
| 16 | Pinocarvone | Oxygenated monoterpenes | C10H14O | 4.2 | 5.6 | 3.1 | 4.54 | 8.9 | 1.55 |
| 17 | β-bisabolene | Sesquiterpene hydrocarbons | C15H24 | 3.0 | - | - | 0.75 | 1.0 | 0.8 |
| 18 | Nerolidol | Sesquiterpene hydrocarbons | C15H26O | trace | 3.1 | 1.2 | - | 0.5 | |
| 19 | α-farnesene | Sesquiterpene hydrocarbons | C15H24 | 1.0 | 1.2 | 1.1 | 0.6 | trace | - |
| 20 | Caryophyllene oxide | Oxygenated sesquiterpnes | C15H24O | trace | - | trace | trace | trace | trace |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Taj, F.; Khan, M.A.; Ali, H.; Khan, R.S. Improved Production of Industrially Important Essential Oils Through Elicitation in the Adventitious Roots of Artemisia amygdalina. Plants 2019, 8, 430. https://doi.org/10.3390/plants8100430
Taj F, Khan MA, Ali H, Khan RS. Improved Production of Industrially Important Essential Oils Through Elicitation in the Adventitious Roots of Artemisia amygdalina. Plants. 2019; 8(10):430. https://doi.org/10.3390/plants8100430
Chicago/Turabian StyleTaj, Faqeer, Mubarak Ali Khan, Huma Ali, and Raham Sher Khan. 2019. "Improved Production of Industrially Important Essential Oils Through Elicitation in the Adventitious Roots of Artemisia amygdalina" Plants 8, no. 10: 430. https://doi.org/10.3390/plants8100430
APA StyleTaj, F., Khan, M. A., Ali, H., & Khan, R. S. (2019). Improved Production of Industrially Important Essential Oils Through Elicitation in the Adventitious Roots of Artemisia amygdalina. Plants, 8(10), 430. https://doi.org/10.3390/plants8100430