Arbuscular Mycorrhizal Fungi Can Compensate for the Loss of Indigenous Microbial Communities to Support the Growth of Liquorice (Glycyrrhiza uralensis Fisch.)


 and
and 
Abstract
:1. Introduction
2. Results
2.1. Mycorrhizal Colonization
2.2. Plant Growth Parameters
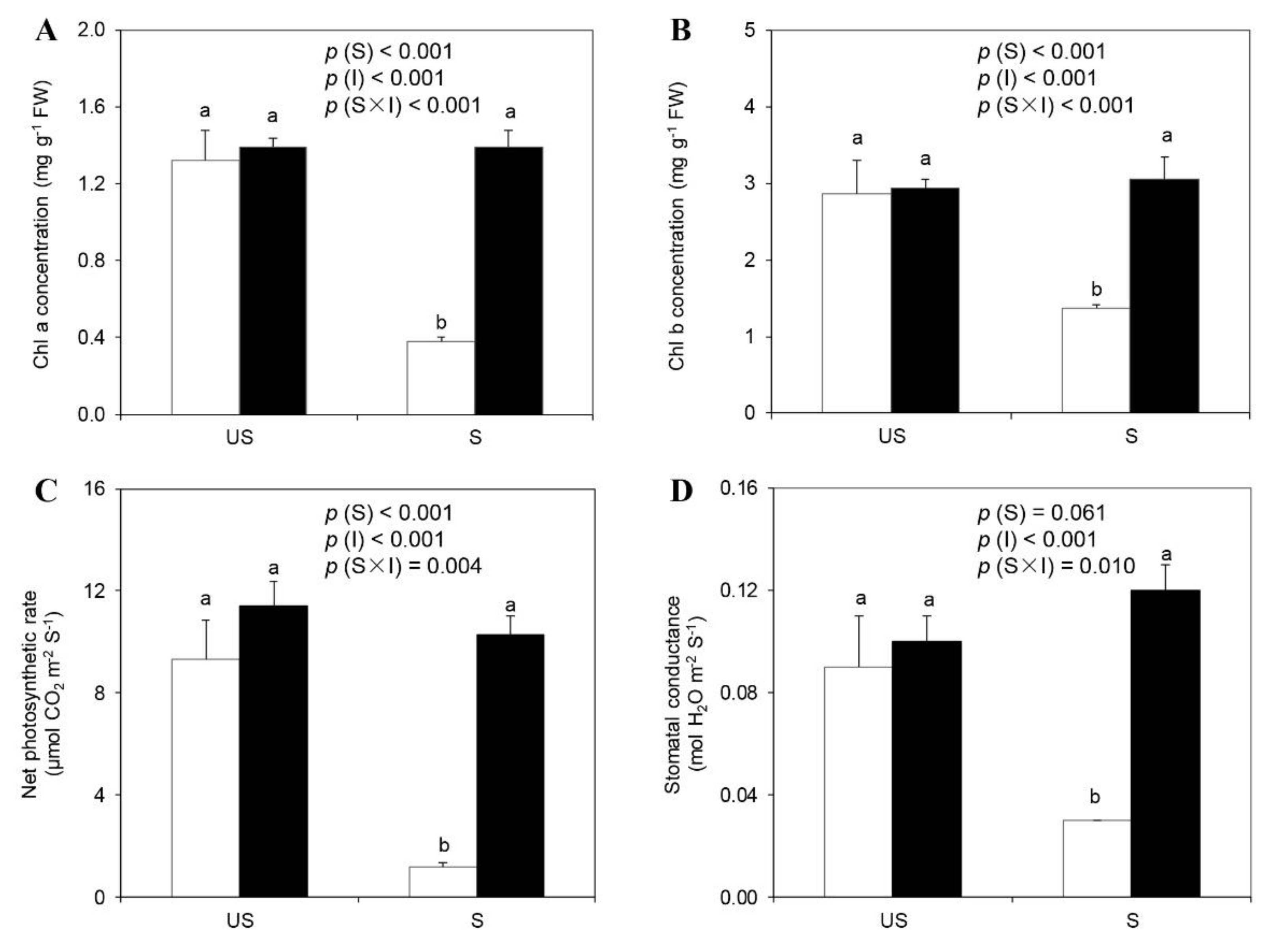
2.3. Chlorophyll Concentration, Net Photosynthetic Rate, and Stomatal Conductance
2.4. Plant Nutrient Concentrations
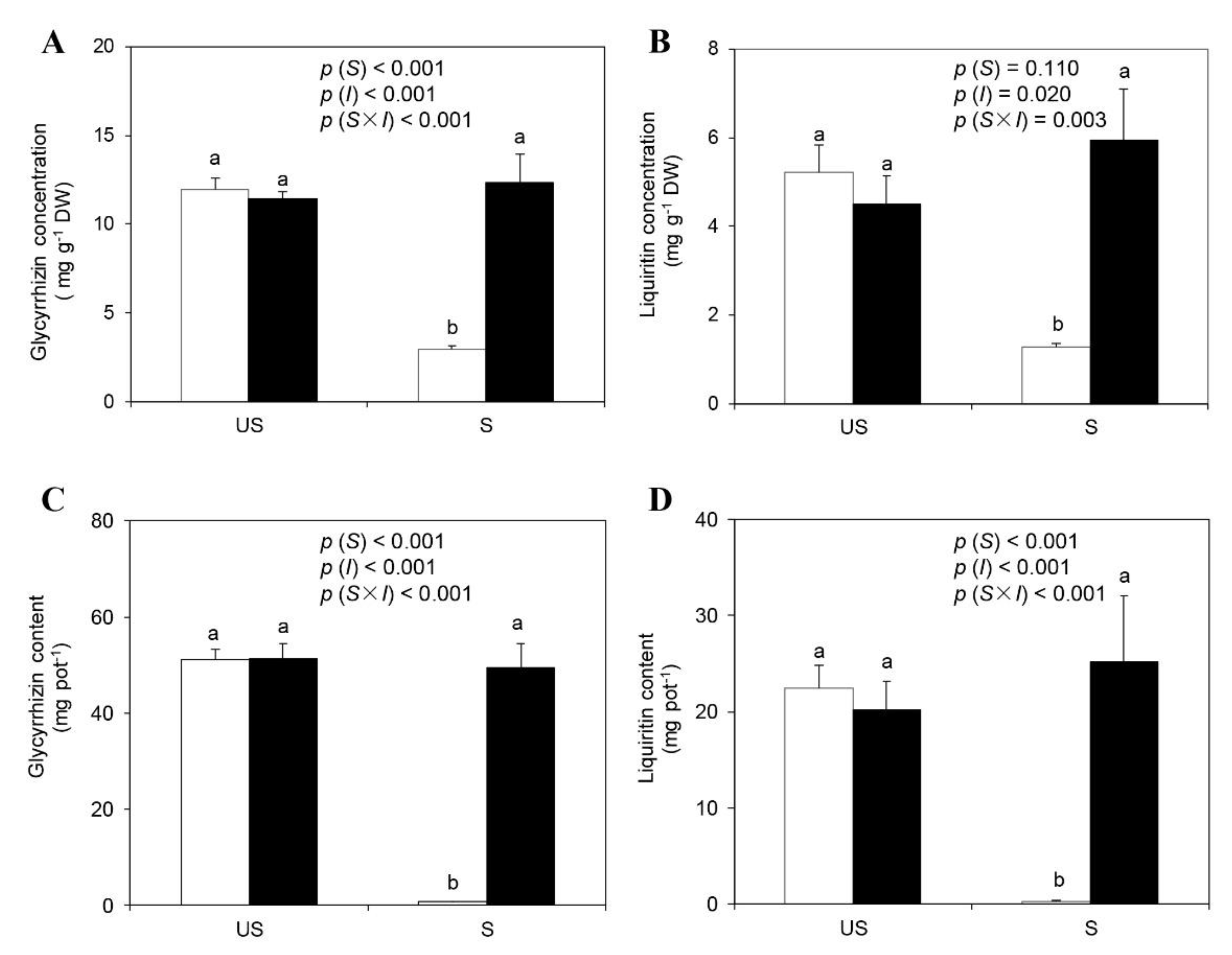
2.5. Root Glycyrrhizin, Liquiritin Concentrations, and Yields
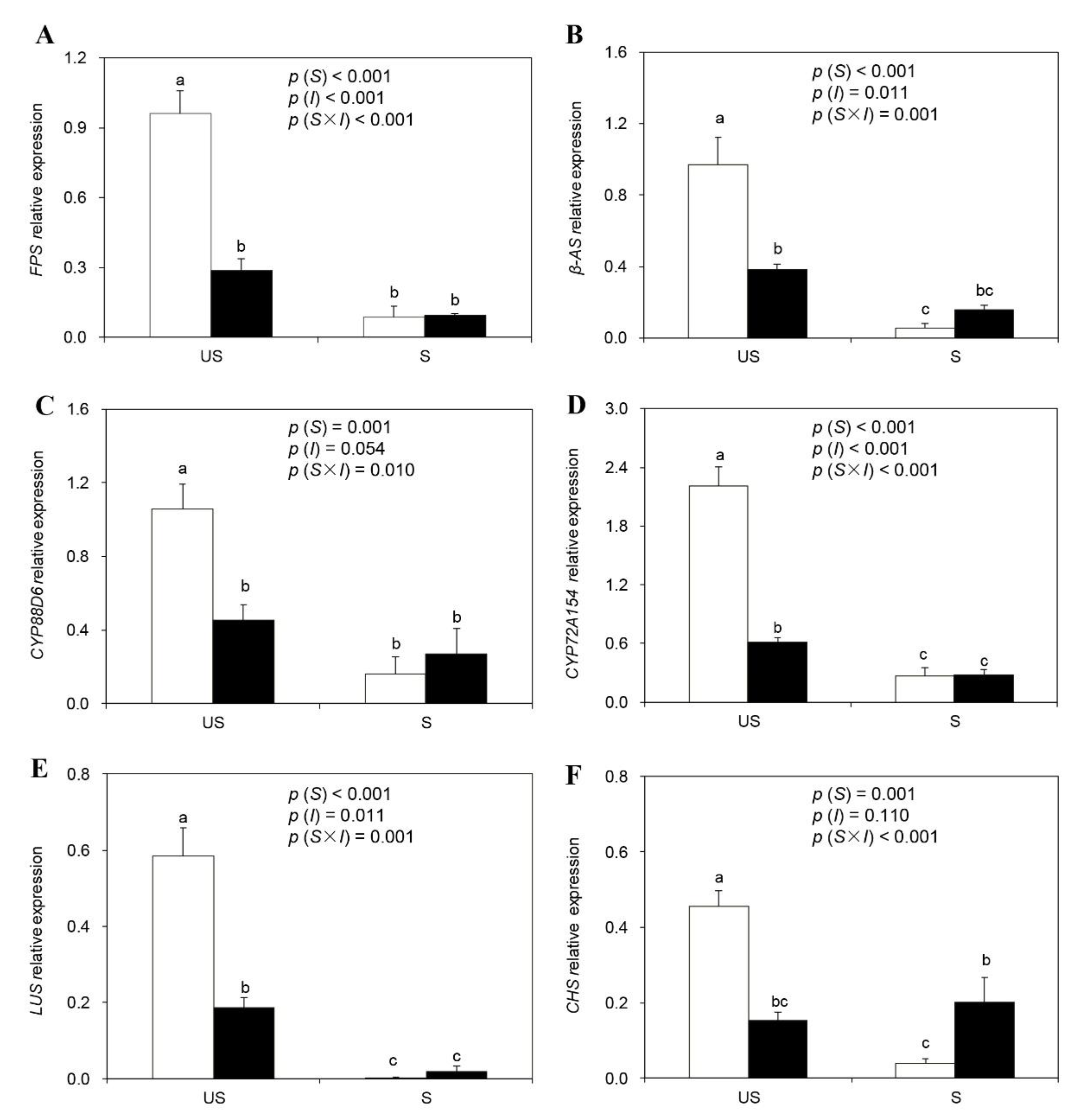
2.6. Expression of Genes for the Biosynthesis of Glycyrrhizin and Liquiritin
3. Discussion
4. Materials and Methods
4.1. Experimental Materials
4.2. Experimental Procedure
4.3. Net Photosynthetic Rate (Pn) and Stomatal Conductance (Cond) of Plants
4.4. Plant Growth
4.5. AM Fungal Colonization
4.6. Chlorophyll Analysis
4.7. Elemental Analysis
4.8. Glycyrrhizin and Liquiritin Analyses
4.9. RNA Isolation and Quantitative Real-Time PCR (qRT-PCR) Analysis
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Van Nuland, M.E.; Wooliver, R.C.; Pfennigwerth, A.A.; Read, Q.D.; Ware, I.M.; Mueller, L.; Fordyce, J.A.; Schweitzer, J.A.; Bailey, J.K. Plant-soil feedbacks: Connecting ecosystem ecology and evolution. Funct. Ecol. 2016, 30, 1032–1042. [Google Scholar] [CrossRef]
- Walder, F.; van der Heijden, M.G.A. Regulation of resource exchange in the arbuscular mycorrhizal symbiosis. Nat. Plants 2015, 1, 15159. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.E.; Smith, F.A. Roles of arbuscular mycorrhizas in plant nutrition and growth: New paradigms from cellular to ecosystem scales. Annu. Rev. Plant Biol. 2011, 62, 227–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, Z.; Fayolle, L.; van Tuinen, D.; Chatagnier, O.; Li, X.; Gianinazzi, S.; Gianinazzi-Pearson, V. Local and systemic mycorrhiza-induced protection against the ectoparasitic nematode Xiphinema index involves priming of defence gene responses in grapevine. J. Exp. Bot. 2012, 63, 3657–3672. [Google Scholar] [CrossRef] [PubMed]
- Hao, Z.; Xie, W.; Chen, B. Arbuscular mycorrhizal symbiosis affects plant immunity to viral infection and accumulation. Viruses 2019, 11, 534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gianinazzi, S.; Gollotte, A.; Binet, M.N.; van Tuinen, D.; Redecker, D.; Wipf, D. Agroecology: The key role of arbuscular mycorrhizas in ecosystem services. Mycorrhiza 2010, 20, 519–530. [Google Scholar] [CrossRef]
- Li, T.; Hu, Y.J.; Hao, Z.P.; Li, H.; Wang, Y.S.; Chen, B.D. First cloning and characterization of two functional aquaporin genes from an arbuscular mycorrhizal fungus Glomus intraradices. N. Phytol. 2013, 197, 617–630. [Google Scholar] [CrossRef]
- López-Ráez, J.A.; Flors, V.; García, J.M.; Pozo, M.J. AM symbiosis alters phenolic acid content in tomato roots. Plant Signal. Behav. 2010, 5, 1138–1140. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Y.; Guo, L.P.; Chen, B.D.; Hao, Z.P.; Wang, J.Y.; Huang, L.Q.; Yang, G.; Cui, X.M.; Yang, L.; Wu, Z.X.; et al. Arbuscular mycorrhizal symbiosis and active ingredients of medicinal plants: Current research status and prospectives. Mycorrhiza 2013, 23, 253–265. [Google Scholar] [CrossRef]
- Oliver, M.A.; Gregory, P.J. Soil, food security and human health: A review. Eur. J. Soil Sci. 2015, 66, 257–276. [Google Scholar] [CrossRef]
- Avio, L.; Turrini, A.; Giovannetti, M.; Sbrana, C. Designing the ideotype mycorrhizal symbionts for the production of healthy food. Front. Plant Sci. 2018, 9, 1089. [Google Scholar] [CrossRef] [PubMed]
- Toju, H.; Peay, K.G.; Yamamichi, M.; Narisawa, K.; Hiruma, K.; Naito, K.; Fukuda, S.; Ushino, M.; Nakaoka, S.; Onoda, Y.; et al. Core microbiomes for sustainable agroecosystems. Nat. Plants 2018, 4, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Asmelash, F.; Bekele, T.; Birhane, E. The potential role of arbuscular mycorrhizal fungi in the restoration of degraded lands. Front. Microbiol. 2016, 7, 1095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banerjee, S.; Schlaeppi, K.; van der Heijden, M.G.A. Keystone taxa as drivers of microbiome structure and functioning. Nat. Rev. Microbiol. 2018, 16, 567–576. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.; Walder, F.; Buchi, L.; Meyer, M.; Held, A.Y.; Gattinger, A.; Keller, T.; Charles, R.; van der Heijden, M.G.A. Agricultural intensification reduces microbial network complexity and the abundance of keystone taxa in roots. ISME J. 2019, 13, 1722–1736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seki, H.; Sawai, S.; Ohyama, K.; Mizutani, M.; Ohnishi, T.; Sudo, H.; Fukushima, E.O.; Akashi, T.; Aoki, T.; Saito, K.; et al. Triterpene functional genomics in licorice for identification of CYP72A154 involved in the biosynthesis of glycyrrhizin. Plant Cell 2011, 23, 4112–4123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizzato, G.; Scalabrin, E.; Radaelli, M.; Capodaglio, G.; Piccolo, O. A new exploration of licorice metabolome. Food Chem. 2017, 221, 959–968. [Google Scholar] [CrossRef]
- Wei, G.H.; Yang, X.Y.; Zhang, J.W.; Gao, J.M.; Ma, Y.Q.; Fu, Y.Y.; Wang, P. Rhizobialide: A new stearolactone produced by Mesorhizobium sp CCNWGX022, a rhizobial endophyte from Glycyrrhiza uralensis. Chem. Biodivers. 2007, 4, 893–898. [Google Scholar] [CrossRef]
- Wang, Y. Diversity of Arbuscular Mycorrhizal Fungi of Licorice (Glycyrrhiza uralensis Fisch) and the Effect of Inoculation; Inner Mongolia University Press: Hohhot, China, 2008; pp. 10–24. [Google Scholar]
- Li, L.; Sinkko, H.; Montonen, L.; Wei, G.H.; Lindstrom, K.; Rasanen, L.A. Biogeography of symbiotic and other endophytic bacteria isolated from medicinal Glycyrrhiza species in China. FEMS Microbiol. Ecol. 2012, 79, 46–68. [Google Scholar] [CrossRef] [Green Version]
- Egamberdieva, D.; Li, L.; Lindstrom, K.; Rasanen, L.A. A synergistic interaction between salt-tolerant Pseudomonas and Mesorhizobium strains improves growth and symbiotic performance of liquorice (Glycyrrhiza uralensis Fish.) under salt stress. Appl. Microbiol. Biotechnol. 2016, 100, 2829–2841. [Google Scholar] [CrossRef]
- Zhao, K.; Zhao, C.; Liao, P.; Zhang, Q.; Li, Y.B.; Liu, M.K.; Ao, X.L.; Gu, Y.F.; Liao, D.C.; Xu, K.W.; et al. Isolation and antimicrobial activities of actinobacteria closely associated with liquorice plants Glycyrrhiza glabra L. and Glycyrrhiza inflate BAT. in Xinjiang, China. Microbiology 2016, 162, 1135–1146. [Google Scholar] [CrossRef] [PubMed]
- Lekberg, Y.; Koide, R.T. Is plant performance limited by abundance of arbuscular mycorrhizal fungi? A meta-analysis of studies published between 1988 and 2003. N. Phytol. 2005, 168, 189–204. [Google Scholar] [CrossRef] [PubMed]
- Hosseinzadeh, H.; Nassiri-Asl, M. Pharmacological effects of Glycyrrhiza spp. and its bioactive constituents: Update and review. Phytother. Res. 2015, 29, 1868–1886. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.M.; Shang, E.X.; Zhao, J.L.; Fan, X.S.; Duan, J.A.; Qian, D.W.; Tao, W.W.; Tang, Y.P. Data mining and frequency analysis for licorice as a “Two-Face” herb in Chinese Formulae based on Chinese Formulae Database. Phytomedicine 2014, 21, 1281–1286. [Google Scholar] [CrossRef] [PubMed]
- Mochida, K.; Sakurai, T.; Seki, H.; Yoshida, T.; Takahagi, K.; Sawai, S.; Uchiyama, H.; Muranaka, T.; Saito, K. Draft genome assembly and annotation of Glycyrrhiza uralensis, a medicinal legume. Plant J. 2017, 89, 181–194. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Liu, S.; Wang, J.; Li, J.; Li, J.; Gao, W. Gene expression of glycyrrhizin acid and accumulation of endogenous signaling molecule in Glycyrrhiza uralensis Fisch. adventitious roots after Saccharomyces cerevisiae and Meyerozyma guilliermondii applications. Biotechnol. Appl. Biochem. 2017, 64, 700–711. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Hao, Z.; Yu, M.; Wu, Z.; Zhao, A.; Li, J.; Zhang, X.; Chen, B. Improved phosphorus nutrition by arbuscular mycorrhizal symbiosis as a key factor facilitating glycyrrhizin and liquiritin accumulation in Glycyrrhiza uralensis. Plant Soil 2019, 439, 243–257. [Google Scholar] [CrossRef]
- Zhang, H.C.; Liu, J.M.; Lu, H.Y.; Gao, S.L. Enhanced flavonoid production in hairy root cultures of Glycyrrhiza uralensis Fisch by combining the over-expression of chalcone isomerase gene with the elicitation treatment. Plant Cell Rep. 2009, 28, 1205–1213. [Google Scholar] [CrossRef]
- Xie, W.; Hao, Z.P.; Zhou, X.F.; Jiang, X.L.; Xu, L.J.; Wu, S.L.; Zhao, A.H.; Zhang, X.; Chen, B.D. Arbuscular mycorrhiza facilitates the accumulation of glycyrrhizin and liquiritin in Glycyrrhiza uralensis under drought stress. Mycorrhiza 2018, 28, 285–300. [Google Scholar] [CrossRef]
- Köhl, L.; Lukasiewicz, C.E.; van der Heijden, M.G.A. Establishment and effectiveness of inoculated arbuscular mycorrhizal fungi in agricultural soils. Plant Cell Environ. 2016, 39, 136–146. [Google Scholar] [CrossRef]
- Savary, R.; Masclaux, F.G.; Wyss, T.; Droh, G.; Corella, J.C.; Machado, A.P.; Morton, J.B.; Sanders, I.R. A population genomics approach shows widespread geographical distribution of cryptic genomic forms of the symbiotic fungus Rhizophagus irregularis. ISME J. 2018, 12, 17–30. [Google Scholar] [CrossRef] [PubMed]
- Séry, D.J.M.; van Tuinen, D.; Drain, A.; Mounier, A.; Zézé, A. The genus Rhizophagus dominates arbuscular mycorrhizal fungi communities in contrasted cassava field soils in Côte d’Ivoire. Rhizosphere 2018, 7, 8–17. [Google Scholar] [CrossRef]
- Gao, C.; Montoya, L.; Xu, L.; Madera, M.; Hollingsworth, J.; Purdom, E.; Hutmacher, R.B.; Dahlberg, J.A.; Coleman-Derr, D.; Lemaux, P.G.; et al. Strong succession in arbuscular mycorrhizal fungal communities. ISME J. 2019, 13, 214–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaparro, J.M.; Badri, D.V.; Vivanco, J.M. Rhizosphere microbiome assemblage is affected by plant development. ISME J. 2014, 8, 790–803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cobb, A.B.; Wilson, G.W.T.; Goad, C.L.; Bean, S.R.; Tesso, T.T.; Wilson, J.D. Assessing the influence of farm fertility amendments, field management, and sorghum genotypes on soil microbial communities and grain quality. Appl. Soil Ecol. 2017, 119, 367–374. [Google Scholar] [CrossRef]
- Niwa, R.; Koyama, T.; Sato, T.; Adachi, K.; Tawaraya, K.; Sato, S.; Hirakawa, H.; Yoshida, S.; Ezawa, T. Dissection of niche competition between introduced and indigenous arbuscular mycorrhizal fungi with respect to soybean yield responses. Sci. Rep. 2018, 8, 7419. [Google Scholar] [CrossRef]
- Castrillo, G.; Teixeira, P.J.P.L.; Paredes, S.H.; Law, T.F.; de Lorenzo, L.; Feltcher, M.E.; Finkel, O.M.; Breakfield, N.W.; Mieczkowski, P.; Jones, C.D.; et al. Root microbiota drive direct integration of phosphate stress and immunity. Nature 2017, 543, 513–518. [Google Scholar] [CrossRef]
- Panke-Buisse, K.; Poole, A.C.; Goodrich, J.K.; Ley, R.E.; Kao-Kniffin, J. Selection on soil microbiomes reveals reproducible impacts on plant function. ISME J. 2015, 9, 980–989. [Google Scholar] [CrossRef]
- Shi, S.; Tian, L.; Ma, L.; Tian, C. Community structure of rhizomicrobiomes in four medicinal herbs and its implication on growth management. Microbiology 2018, 87, 425–436. [Google Scholar] [CrossRef]
- Hao, Z.; Xie, W.; Jiang, X.; Wu, Z.; Zhang, X.; Chen, B. Arbuscular mycorrhizal fungus improves rhizobium-Glycyrrhiza seedling symbiosis under drought stress. Agronomy 2019, 9, 572. [Google Scholar] [CrossRef] [Green Version]
- Wagg, C.; Bender, S.F.; Widmer, F.; van der Heijden, M.G.A. Soil biodiversity and soil community composition determine ecosystem multifunctionality. Proc. Natl. Acad. Sci. USA 2014, 111, 5266–5270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cobb, A.B.; Wilson, G.W.T.; Goad, C.L.; Bean, S.R.; Kaufman, R.C.; Herald, T.J.; Wilson, J.D. The role of arbuscular mycorrhizal fungi in grain production and nutrition of sorhgum genotypes: Enhancing sustainability through plant-microbial partnership. Agric. Ecosyst. Environ. 2016, 233, 432–440. [Google Scholar] [CrossRef]
- Kang, S.M.; Radhakrishnan, R.; Khan, A.L.; Kim, M.J.; Park, J.M.; Kim, B.R.; Shin, D.H.; Lee, I.J. Gibberellin secreting rhizobacterium, Pseudomonas putida H-2-3 modulates the hormonal and stress physiology of soybean to improve the plant growth under saline and drought conditions. Plant Physiol. Bioch. 2014, 84, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Van de Mortel, J.E.; de Vos, R.C.H.; Dekkers, E.; Pineda, A.; Guillod, L.; Bouwmeester, K.; van Loon, J.J.A.; Dicke, M.; Raaijmakers, J.M. Metabolic and transcriptomic changes induced in Arabidopsis by the rhizobacterium Pseudomonas fluorescens SS101. Plant Physiol. 2012, 160, 2173–2188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Welling, M.T.; Liu, L.; Rose, T.J.; Waters, D.L.E.; Benkendorff, K. Arbuscular mycorrhizal fungi: Effects on plant terpenoid accumulation. Plant Biol. 2016, 18, 552–562. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, R.; Anand, G.; Gupta, P.; Mandal, S. Insight into the mechanisms of enhanced production of valuable terpenoids by arbuscular mycorrhiza. Phytochem. Rev. 2017, 16, 677–692. [Google Scholar] [CrossRef]
- Sharma, E.; Anand, G.; Kapoor, R. Terpenoids in plant and arbuscular mycorrhiza-reinforced defence against herbivorous insects. Ann. Bot. 2017, 119, 791–801. [Google Scholar] [CrossRef] [Green Version]
- Miransari, M.; Bahrami, H.A.; Rejali, F.; Malakouti, M.J. Effects of soil compaction and arbuscular mycorrhiza on corn (Zea mays L.) nutrient uptake. Soil Till. Res. 2009, 103, 282–290. [Google Scholar] [CrossRef]
- Troelstra, S.R.; Wagenaar, R.; Smant, W.; Peters, B.A.M. Interpretation of bioassays in the study of interactions between soil organisms and plants: Involvement of nutrient factors. N. Phytol. 2001, 150, 697–706. [Google Scholar] [CrossRef]
- Öztürk, M.; Altay, V.; Hakeem, K.R.; Akçiçek, E. Liquorice-mycorrhiza interactions. In Liquorice: From Botany to Phytochemistry; Springer International Publishing: Cham, Switzerland, 2017; pp. 31–40. [Google Scholar]
- Lugtenberg, B.; Kamilova, F. Plant-growth-promoting rhizobacteria. Annu. Rev. Microbiol. 2009, 63, 541–556. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Aparicio, L.; Dominguez-Begines, J.; Kardol, P.; Avila, J.M.; Ibanez, B.; Garcia, L.V. Plant-soil feedbacks in declining forests: Implications for species coexistence. Ecology 2017, 98, 1908–1921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koerselman, W.; Meuleman, A.F.M. The vegetation N:P ratio: A new tool to detect the nature of nutrient limitation. J. Appl. Ecol. 1996, 33, 1441–1450. [Google Scholar] [CrossRef]
- Bender, S.F.; Schlaeppi, K.; Held, A.; van der Heijden, M.G.A. Establishment success and crop growth effects of an arbuscular mycorrhizal fungus inoculated into Swiss corn fields. Agr. Ecosyst. Environ. 2019, 273, 13–24. [Google Scholar] [CrossRef] [Green Version]
- Hausmann, N.T.; Hawkes, C.V. Order of plant host establishment alters the composition of arbuscular mycorrhizal communities. Ecology 2010, 91, 2333–2343. [Google Scholar] [CrossRef]
- Bandyopadhyay, P.; Bhuyan, S.K.; Yadava, P.K.; Varma, A.; Tuteja, N. Emergence of plant and rhizospheric microbiota as stable interactomes. Protoplasma 2017, 254, 617–626. [Google Scholar] [CrossRef]
- Köhl, L.; van der Heijden, M.G.A. Arbuscular mycorrhizal fungal species differ in their effect on nutrient leaching. Soil Biol. Biochem. 2016, 94, 191–199. [Google Scholar] [CrossRef] [Green Version]
- Verbruggen, E.; van der Heijden, M.G.A.; Rillig, M.C.; Kiers, E.T. Mycorrhizal fungal establishment in agricultural soils: Factors determining inoculation success. N. Phytol. 2013, 197, 1104–1109. [Google Scholar] [CrossRef] [Green Version]
- Shrivastava, G.; Ownley, B.H.; Auge, R.M.; Toler, H.; Dee, M.; Vu, A.; Kollner, T.G.; Chen, F. Colonization by arbuscular mycorrhizal and endophytic fungi enhanced terpene production in tomato plants and their defense against a herbivorous insect. Symbiosis 2015, 65, 65–74. [Google Scholar] [CrossRef]
- Bryant, J.P.; Chapin, F.S.; Klein, D.R. Carbon/nutrient balance of boreal plants in relation to vertebrate herbivory. Oikos 1983, 40, 357–368. [Google Scholar] [CrossRef] [Green Version]
- McNamara, N.P.; Black, H.I.J.; Beresford, N.A.; Parekh, N.R. Effects of acute gamma irradiation on chemical, physical and biological properties of soils. Appl. Soil Ecol. 2003, 24, 117–132. [Google Scholar] [CrossRef]
- Berns, A.E.; Philipp, H.; Narres, H.D.; Burauel, P.; Vereecken, H.; Tappe, W. Effect of gamma-sterilization and autoclaving on soil organic matter structure as studied by solid state NMR, UV and fluorescence spectroscopy. Eur. J. Soil Sci. 2008, 59, 540–550. [Google Scholar] [CrossRef]
- Trouvelot, A.; Kough, J.L.; Gianinazzi-Pearson, V. Mesure du taux de mycorhization VA d’un systeme radiculaire. Recherche de methods d’estimation ayant une signification fonctionnelle. In Physiological and Genetical Aspects of Mycorrhizae; Gianinazzi-Pearson, V., Gianinazzi, S., Eds.; INRA: Paris, France, 1986; pp. 217–221. [Google Scholar]
- Arnon, D.I. Copper enzymes in isolated chloroplasts, polyphenoxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Sterilization (S) | Inoculation (I) | Mycorrhizal Colonization Rate (M%) | Arbuscule Abundance (A%) |
|---|---|---|---|
| US | −M | 31.7 ± 5.2 b | 6.0 ± 1.0 bc |
| +M | 49.9 ± 5.6 a | 16.2 ± 3.2 ab | |
| S | −M | 0.0 ± 0.0 c | 0.0 ± 0.0 c |
| +M | 65.3 ± 4.0 a | 26.6 ± 5.2 a | |
| ANOVA | S | F1,12 = 3.56 ns | F1,12 = 0.61 ns |
| I | F1,12 = 94.07 ** | F1,12 = 36.21 ** | |
| S × I | F1,12 = 29.86 ** | F1,12 = 6.69 * |
| Sterilization (S) | Inoculation (I) | Plant Height (cm) | Shoot Dry Weight (g pot−1) | Root Dry Weight (g pot−1) |
|---|---|---|---|---|
| US | −M | 44.2 ± 2.5 a | 1.92 ± 0.10 a | 4.31 ± 0.10 a |
| +M | 47.1 ± 3.0 a | 2.50 ± 0.20 a | 4.48 ± 0.11 a | |
| S | −M | 10.0 ± 0.9 b | 0.10 ± 0.02 b | 0.23 ± 0.03 b |
| +M | 49.5± 4.4 a | 2.73 ± 0.20 a | 4.11 ± 0.40 a | |
| ANOVA | S | F1,12 = 53.445 ** | F1,12 = 120.56 ** | F1,12 = 389.55 ** |
| I | F1,12 = 80.132 ** | F1,12 = 181.54 ** | F1,12 = 357.90 ** | |
| S × I | F1,12 = 64.295 ** | F1,12 = 135.08 ** | F1,12 = 339.84 ** |
| Sterilization (S) | Inoculation (I) | C (mg g−1) | N (mg g−1) | P (mg g−1) | K (mg g−1) | Ca (mg g−1) | Mg (mg g−1) | C:N | C:P | N:P | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Shoot | US | −M | 435 ± 5 a | 20.24 ± 0.67 b | 1.66 ± 0.09 a | 18.53 ± 0.77 b | 13.93 ± 0.57 b | 2.76 ± 0.16 b | 21.55 ± 0.53 a | 264 ± 14.71 b | 12.2 ± 0.62 b |
| +M | 435 ± 4 a | 19.78 ± 0.49 b | 1.60 ± 0.16 a | 17.81 ± 1.53 b | 11.27 ± 0.35 c | 2.34 ± 0.23 b | 22.03 ± 0.36 a | 280 ± 26.92 b | 12.7 ± 1.16 b | ||
| S | −M | 411 ± 3 b | 24.33 ± 0.58 a | 0.42 ± 0.01 b | 22.85 ± 0.76 a | 25.90 ± 0.75 a | 4.48 ± 0.32 a | 16.93 ± 0.53 b | 962 ± 32.73 a | 57.1 ± 3.07 a | |
| +M | 443 ± 3 a | 20.41 ± 0.88 b | 1.60 ± 0.06 a | 16.83 ± 0.60 b | 11.96 ± 0.95 bc | 2.28 ± 0.21 b | 21.85 ± 1.03 a | 278 ± 12.31 b | 12.7 ± 0.35 b | ||
| ANOVA | S | F1,12 = 4.60 ns | F1,12 = 12.37 ** | F1,12 = 117.23 ** | F1,12 = 2.89 ns | F1,12 = 47.97 ** | F1,12 = 12.18 ** | F1,12 = 13.28 ** | F1,12 = 223.70 ** | F1,12 = 947.92 ** | |
| I | F1,12 = 18.09 ** | F1,12 = 10.65 ** | F1,12 = 105.79 ** | F1,12 = 11.67 ** | F1,12 = 104.19 ** | F1,12 = 30.20 ** | F1,12 = 16.72 ** | F1,12 = 206.51 ** | F1,12 = 884.38 ** | ||
| S × I | F1,12 = 18.01 ** | F1,12 = 6.65 * | F1,12 = 122.85 ** | F1,12 = 7.23 * | F1,12 = 34.44 ** | F1,12 = 13.85 ** | F1,12 = 11.32 ** | F1,12 = 227.12 ** | F1,12 = 955.09 ** | ||
| Root | US | −M | 403 ± 10 b | 16.26 ± 1.75 b | 2.58 ± 0.14 a | 7.48 ± 0.15 a | 3.83 ± 0.20 | 0.69 ± 0.07 b | 25.53 ± 2.23 a | 157 ± 5.20 b | 6.24 ± 0.34 b |
| +M | 413 ± 5 b | 16.72 ± 0.82 b | 2.43 ± 0.11 a | 7.89 ± 0.47 a | 4.15 ± 0.35 | 0.90 ± 0.08 b | 24.85 ± 1.16 a | 171 ± 9.24 b | 6.94 ± 0.57 b | ||
| S | −M | 436 ± 2 a | 29.64 ± 0.41 a | 0.69 ± 0.01 b | 4.28 ± 0.05 b | 4.40 ± 0.37 | 1.55 ± 0.06 a | 14.71 ± 0.13 b | 636 ± 13.23 a | 43.4 ± 0.66 a | |
| +M | 422 ± 1b | 18.89 ± 0.77 b | 2.62 ± 0.15 a | 7.95 ± 0.45 a | 4.19 ± 0.17 | 0.85 ± 0.06 b | 22.45 ± 0.97 a | 153 ± 14.16 b | 6.87 ± 0.76 b | ||
| ANOVA | S | - | F1,12 = 53.87 ** | F1,12 = 167.49 ** | F1,12 = 22.00 ** | F1,12 = 1.53 ns | F1,12 = 40.86 ** | - | F1,12 = 440.14 ** | F1,12 = 47.32 ** | |
| I | - | F1,12 = 23.58 ** | F1,12 = 174.00 ** | F1,12 = 37.24 ** | F1,12 = 0.05 ns | F1,12 = 14.32 ** | - | F1,12 = 455.94 ** | F1,12 = 37.76 ** | ||
| S × I | - | F1,12 = 27.98 ** | F1,12 = 208.10 ** | F1,12 = 23.77 ** | F1,12 = 1.11 ns | F1,12 = 51.36 ** | - | F1,12 = 511.41 ** | F1,12 = 41.40 ** |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, M.; Xie, W.; Zhang, X.; Zhang, S.; Wang, Y.; Hao, Z.; Chen, B. Arbuscular Mycorrhizal Fungi Can Compensate for the Loss of Indigenous Microbial Communities to Support the Growth of Liquorice (Glycyrrhiza uralensis Fisch.). Plants 2020, 9, 7. https://doi.org/10.3390/plants9010007
Yu M, Xie W, Zhang X, Zhang S, Wang Y, Hao Z, Chen B. Arbuscular Mycorrhizal Fungi Can Compensate for the Loss of Indigenous Microbial Communities to Support the Growth of Liquorice (Glycyrrhiza uralensis Fisch.). Plants. 2020; 9(1):7. https://doi.org/10.3390/plants9010007
Chicago/Turabian StyleYu, Meng, Wei Xie, Xin Zhang, Shubin Zhang, Youshan Wang, Zhipeng Hao, and Baodong Chen. 2020. "Arbuscular Mycorrhizal Fungi Can Compensate for the Loss of Indigenous Microbial Communities to Support the Growth of Liquorice (Glycyrrhiza uralensis Fisch.)" Plants 9, no. 1: 7. https://doi.org/10.3390/plants9010007
APA StyleYu, M., Xie, W., Zhang, X., Zhang, S., Wang, Y., Hao, Z., & Chen, B. (2020). Arbuscular Mycorrhizal Fungi Can Compensate for the Loss of Indigenous Microbial Communities to Support the Growth of Liquorice (Glycyrrhiza uralensis Fisch.). Plants, 9(1), 7. https://doi.org/10.3390/plants9010007