Taxonomic Notes on the ‘Mahat’ (Artocarpus lacucha and A. thailandicus, Moraceae) Species Complex in Thailand

 , , and
, , and
Abstract
:1. Introduction
2. Results
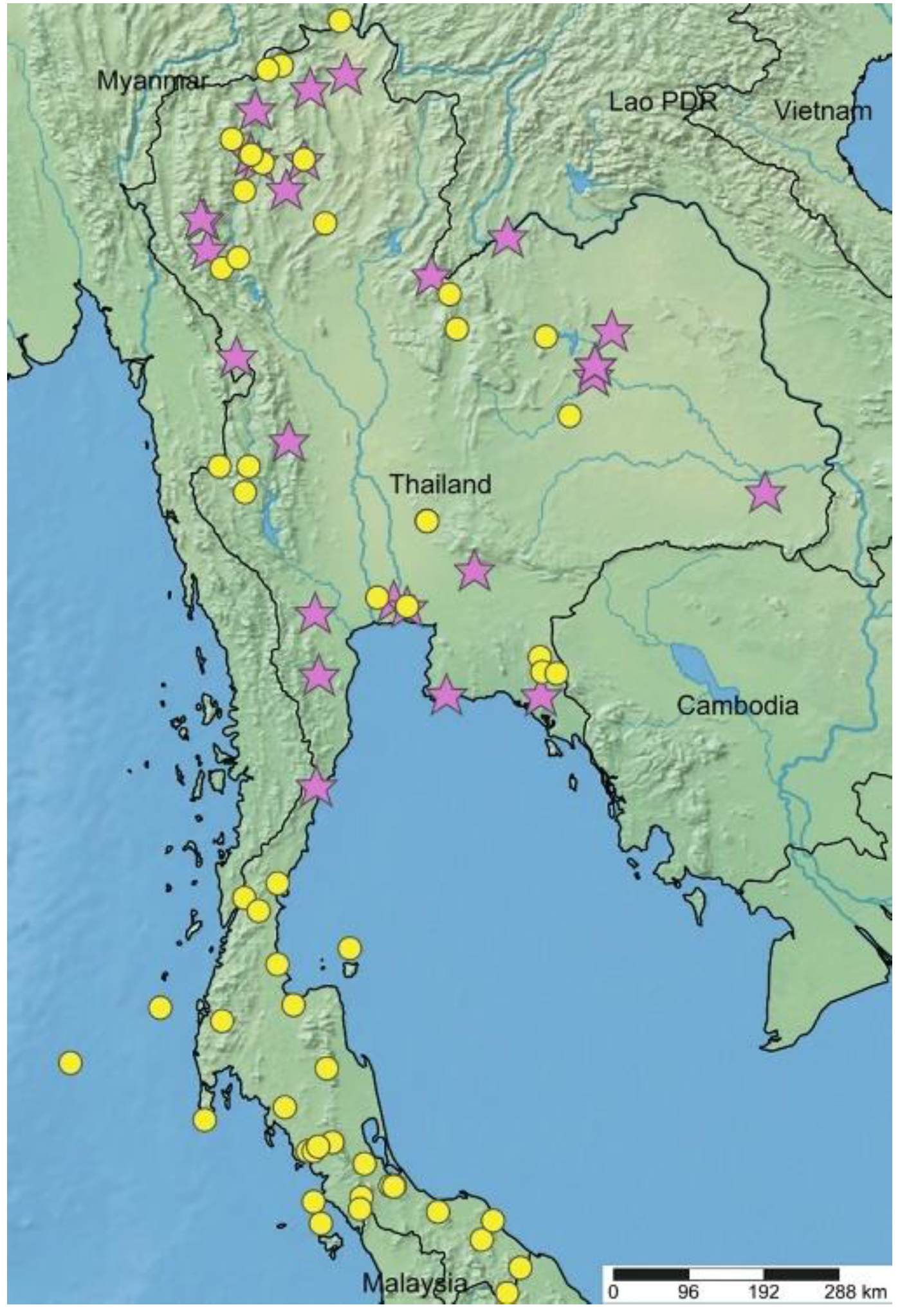
2.1. Macroscopic Morphology and Distribution
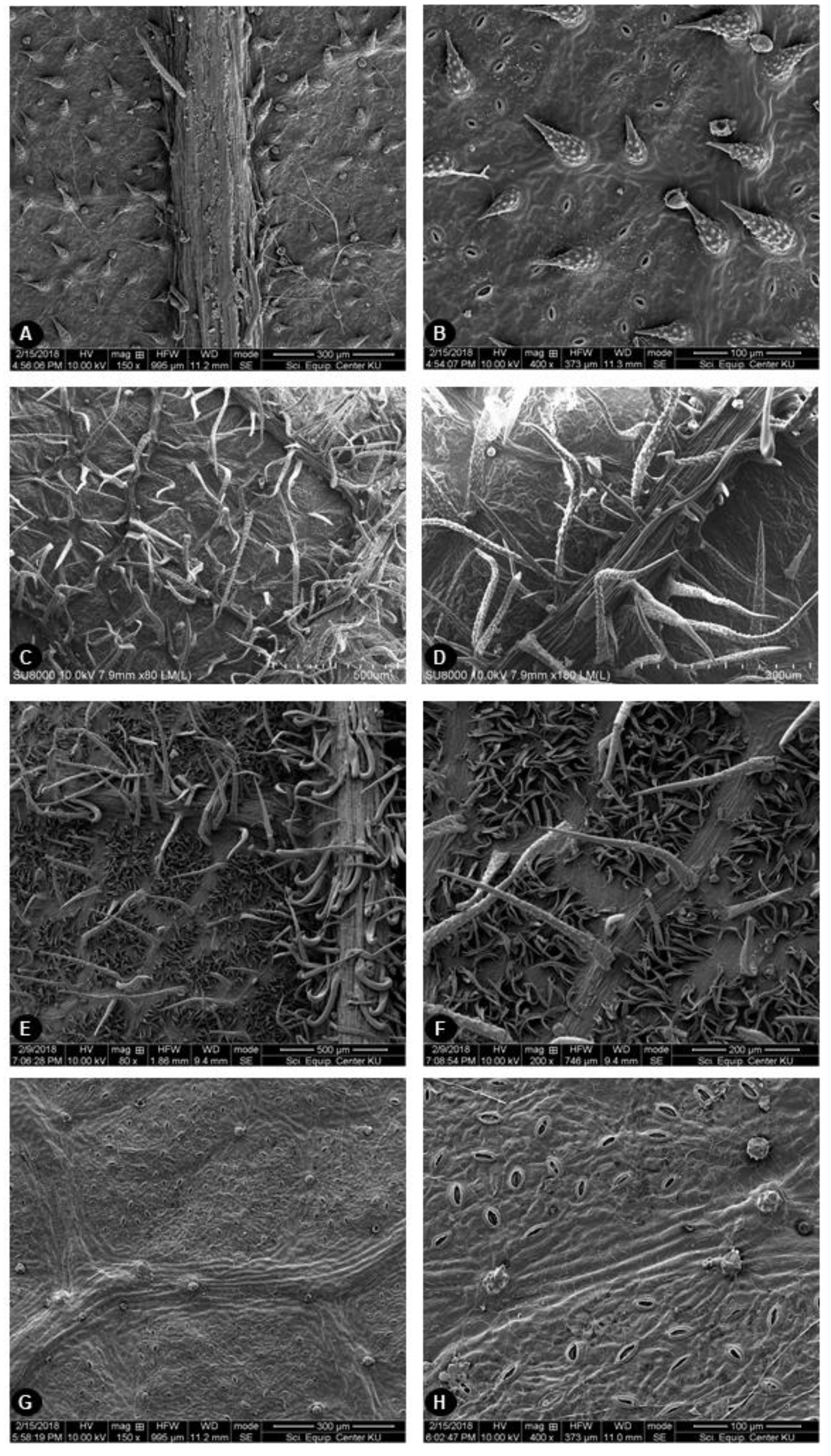
2.2. Leaf Surface Morphology
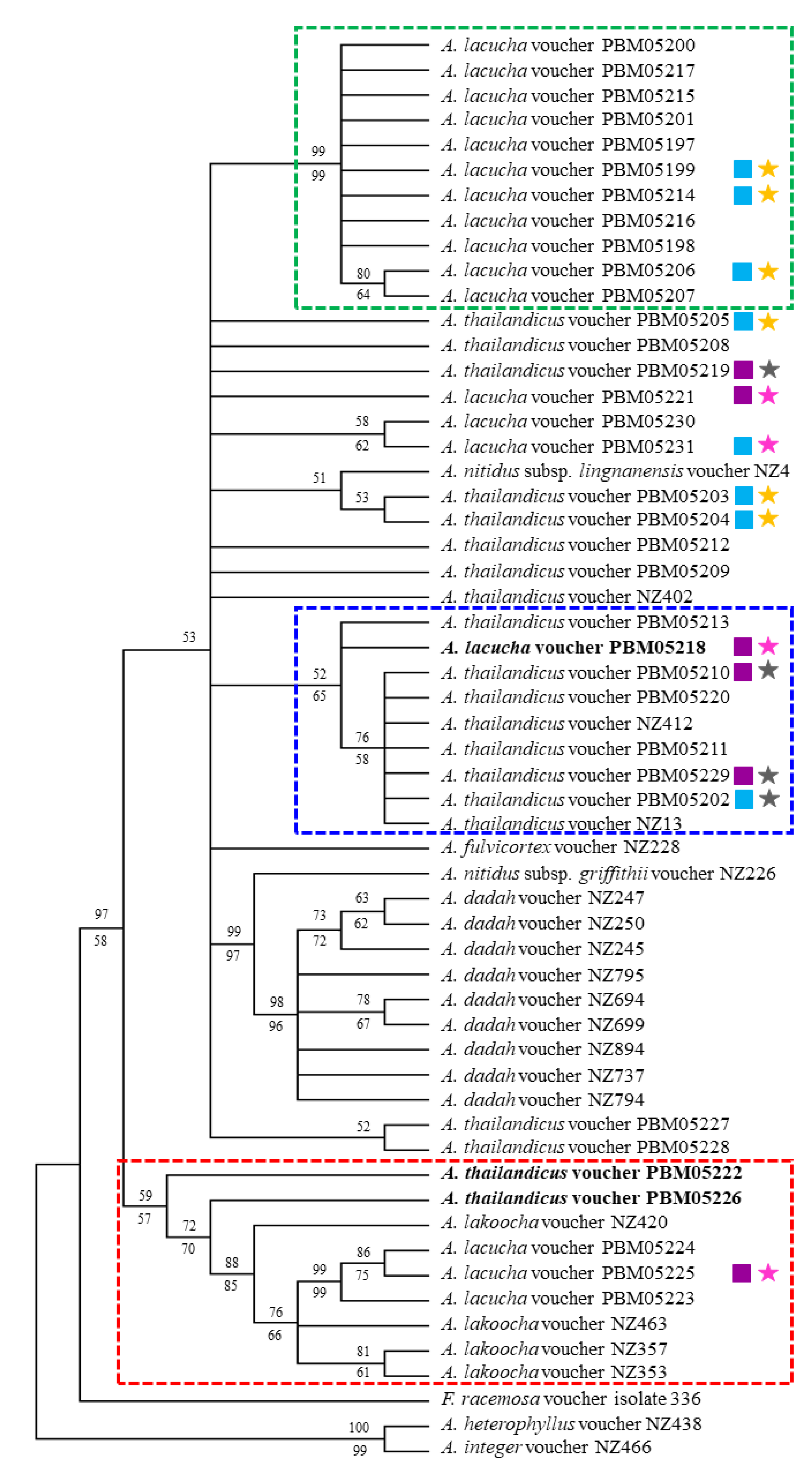
2.3. Phylogenetics
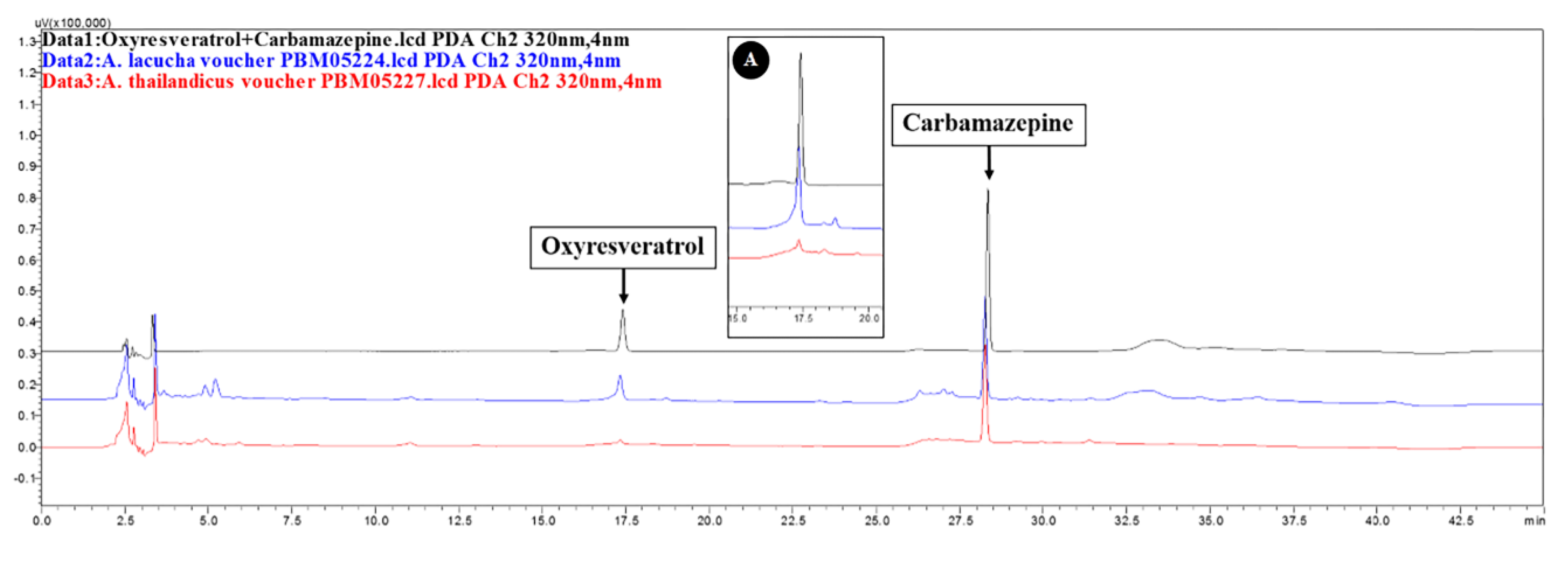
2.4. Qualitative Phytochemistry
3. Discussion
4. Materials and Methods
4.1. Taxonomic Study
4.2. Scanning Electron Microscopy (SEM)
4.3. DNA Extraction, Amplification and Sequencing
4.4. Sequence Alignment and Phylogenetic Analysis
4.5. Qualitative Phytochemical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zhang, Q.; Onstein, R.E.; Little, S.A.; Sauquet, H. Estimating divergence times and ancestral breeding systems in Ficus and Moraceae. Ann. Bot. 2018, 123, 191–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zerega, N.J.C.; Nur Supardi, M.N.; Motley, T.J. Phylogeny and Recircumscription of Artocarpeae (Moraceae) with a Focus on Artocarpus. Syst. Bot. 2010, 35, 766–782. [Google Scholar] [CrossRef] [Green Version]
- Datwyler, S.L.; Weiblen, G.D. On the origin of the fig: Phylogenetic relationships of Moraceae from ndhF sequences. Am. J. Bot. 2004, 91, 767–777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zerega, N.J.C.; Clement, W.L.; Datwyler, S.L.; Weiblen, G.D. Biogeography and divergence times in the mulberry family (Moraceae). Mol. Phylogenet. Evol. 2005, 37, 402–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clement, W.L.; Weiblen, G.D. Morphological evolution in the mulberry family (Moraceae). Syst. Bot. 2009, 34, 530–552. [Google Scholar] [CrossRef]
- Williams, E.W.; Gardner, E.M.; Harris, R., III; Chaveerach, A.; Pereira, J.T.; Zerega, N.J.C. Out of Borneo: Biogeography, phylogeny and divergence date estimates of Artocarpus (Moraceae). Ann. Bot. 2017, 119, 611–627. [Google Scholar] [CrossRef] [Green Version]
- Trécul, A. Mémoire sur la famille des Artocarpées. Ann. Sci. Nat. 1847, 3, 6–383. [Google Scholar]
- Jarrett, F.M. Studies in Artocarpus and allied genera, I. General considerations. J. Arnold Arbor. 1959, 40, 1–29. [Google Scholar]
- Jarrett, F.M. Studies in Artocarpus and allied genera, II. A revision of Prainea. J. Arnold Arbor. 1959, 40, 30–37. [Google Scholar]
- Berg, C.C. A New Species of Artocarpus (Moraceae) from Thailand. Blumea 2005, 50, 531–533. [Google Scholar] [CrossRef] [Green Version]
- Berg, C.C.; Pattharahirantricin, N.; Chantarasuwan, B. Moraceae. In Flora of Thailand; Santisuk, T., Larsen, K., Eds.; The Forest Herbarium, Department of National Parks, Wildlife and Plant Conservation: Bangkok, Thailand, 2011; Volume 10, pp. 475–675. [Google Scholar]
- Maneechai, S.; Likhitwitayawuid, K.; Sritularak, B.; Palanuvej, C.; Ruangrungsi, N.; Sirisa-ard, P. Quantitative Analysis of Oxyresveratrol Content in Artocarpus lakoocha and ‘Puag-Haad’. Med. Princ. Pract. 2009, 18, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, P.; Roychowdhury, S.; Engelmann, M.; Wolf, G.; Horn, T.F.W. Oxyresveratrol and resveratrol are potent antioxidants and free radical scavengers: Effect on nitrosative and oxidative stress derived from microglial cells. Nitric Oxide 2003, 9, 64–76. [Google Scholar] [CrossRef] [PubMed]
- Chung, K.O.; Kim, B.Y.; Lee, M.H.; Kim, Y.R.; Chung, H.Y.; Park, J.H.; Moon, J.O. In-vitro and in-vivo anti-inflammatory effect of oxyresveratrol from Morus alba L. J. Pharm. Pharmacol. 2003, 55, 1695–1700. [Google Scholar] [CrossRef] [PubMed]
- Likhitwitayawuid, K.; Sornsute, A.; Sritularak, B.; Ploypradith, P. Chemical transformations of oxyresveratrol (trans-2,4,3′,5′-tetrahydroxystilbene) into a potent tyrosinase inhibitor and a strong cytotoxic agent. Bioorg. Med. Chem. Lett. 2006, 16, 5650–5653. [Google Scholar] [CrossRef]
- Tengamnuay, P.; Pengrungruangwong, K.; Pheansri, I.; Likhitwitayawuid, K. Artocarpus lakoocha heartwood extract as a novel cosmetic ingredient: Evaluation of the in vitro anti-tyrosinase and in vivo skin whitening activities. Int. J. Cosmet. Sci. 2006, 28, 269–276. [Google Scholar] [CrossRef]
- Likhitwitayawuid, K. Stilbenes with tyrosinase inhibitory activity. Curr. Sci. 2008, 94, 44–52. [Google Scholar]
- Mabberley, D.J. Francis Hamilton’s Commentaries with Particular Reference to Meliaceae. Taxon 1977, 26, 529. [Google Scholar] [CrossRef]
- Charoenlarp, P.; Radomyos, P.; Harinasuta, T. Treatment of taeniasis with Puag-Haad: A crude extract of Artocarpus lakoocha wood. Southeast Asian J. Trop. Med. Public Health 1981, 12, 568–570. [Google Scholar]
- Puntumchai, A.; Kittakoop, P.; Rajviroongit, S.; Vimuttipong, S.; Likhitwitayawuid, K.; Thebtaranonth, Y. Lakoochins A and B, New Antimycobacterial Stilbene Derivatives from Artocarpus lakoocha. J. Nat. Prod. 2004, 67, 485–486. [Google Scholar] [CrossRef]
- Likhitwitayawuid, K.; Sritulalak, B.; Benchanak, K.; Lipipun, V.; Mathew, J.; Schinazi, R.F. Phenolics with antiviral activity from Millettia erythrocalyx and Artocarpus lakoocha. Nat. Prod. Res. 2005, 19, 177–182. [Google Scholar] [CrossRef]
- Chuanasa, T.; Phromjai, J.; Lipipun, V.; Likhitwitayawuid, K.; Suzuki, M.; Pramyothin, P.; Hattori, M.; Shiraki, K. Anti-herpes simplex virus (HSV-1) activity of oxyresveratrol derived from Thai medicinal plant: Mechanism of action and therapeutic efficacy on cutaneous HSV-1 infection in mice. Antivir. Res. 2008, 80, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Sasivimolphan, P.; Lipipun, V.; Likhitwitayawuid, K.; Takemoto, M.; Pramyothin, P.; Hattori, M.; Shiraki, K. Inhibitory activity of oxyresveratrol on wild-type and drug-resistant varicella-zoster virus replication in vitro. Antivir. Res. 2009, 84, 95–97. [Google Scholar] [CrossRef] [PubMed]
- Boonkla, W.; Pitchuanchom, S.; Meepowpan, P.; Thaisuchat, H.; Nuntasaen, N.; Punyanitya, S.; Pompimon, W. Aromatic compound glucopyranoside from new species Artocarpus thailandicus. Am. J. Appl. Sci. 2011, 8, 1093–1097. [Google Scholar] [CrossRef]
- Royal Botanic Garden, C.; Carey, W.; Roxburgh, W. Classis XXI. MONOECIA MONANDRIA. Hort. Bengal. 1814, 1, 65–70. Available online: https://www.biodiversitylibrary.org/bibliography/95337#/summary (accessed on 22 March 2020).
- Ragone, D. Chromosome numbers and pollen stainability of three species of Pacific Island breadfruit (Artocarpus, Moraceae). Am. J. Bot. 2001, 88, 693–696. [Google Scholar] [CrossRef]
- Jarrett, F.M. Studies in Artocarpus and allied genera. III. A revision of Artocarpus subgenus Artocarpus. J. Arnold Arbor. 1959, 40, 113–155. [Google Scholar]
- Brantjes, N.B.M. Nectar and pollination of breadfruit, Artocarpus altilis (Moraceae). Acta Bot. Neerl. 1981, 30, 345–352. [Google Scholar] [CrossRef]
- Maraci, Ö.; Özkan, H.; Bilgin, R. Phylogeny and genetic structure in the genus Secale. PLoS ONE 2018, 13, e0200825. [Google Scholar] [CrossRef]
- Anand, K.K.; Jena, S.N.; Chaudhary, L.B.; Singh, M. Conflict between morphological and molecular data: A case study of Ficus krishnae (Moraceae). Phytotaxa 2016, 247, 143–147. [Google Scholar] [CrossRef] [Green Version]
- Dalin, P.; Ågren, J.; Björkman, C.; Huttunen, P.; Kärkkäinen, K. Leaf trichome formation and plant resistance to herbivory. In Induced Plant Resistance to Herbivory; Schaller, A., Ed.; Springer: Dordrecht, The Netherlands, 2008; pp. 89–105. [Google Scholar]
- War, A.R.; Paulraj, M.G.; Sharma, H.C. Mechanisms of plant defense against insect herbivores. Plant Signal Behav. 2012, 7, 1306–1320. [Google Scholar] [CrossRef] [Green Version]
- Jhonson, H.B. Plant pubescence: An ecological perspective. Bot. Rev. 1975, 41, 233–258. [Google Scholar] [CrossRef]
- Chantarasuwan, B.; Baas, P.; van Heuven, B.J.; Baider, C.; van Welzen, P.C. Leaf anatomy of Ficus subsection Urostigma (Moraceae). Bot. J. 2014, 175, 259–281. [Google Scholar] [CrossRef] [Green Version]
- Rebeiz, M.; Tsiantis, M. Enhancer evolution and the origins of morphological novelty. Curr. Opin. Genet. Dev. 2017, 45, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Das Gupta, M.; Tsiantis, M. Gene networks and the evolution of plant morphology. Curr. Opin. Plant Biol. 2018, 45, 82–87. [Google Scholar] [CrossRef] [PubMed]
- Lazarowych, N.J.; Pekos, P. Use of fingerprinting and marker compounds for identification and standardization of botanical drugs: Strategies for Applying Pharmaceutical HPLC Analysis to Herbal Products. Ther. Innov. Regul. Sci. 1998, 32, 497–512. [Google Scholar] [CrossRef]
- Giri, L.; Andola, H.; Purohit, V.K.; Rawat, M.S.M.; Rawal, R.; Bhatt, I.D. Chromatographic and spectral fingerprinting standardization of traditional medicines: An overview as modern tools. Res. J. Phytochem. 2010, 4, 234–241. [Google Scholar] [CrossRef] [Green Version]
- SimpleMappr, an Online Tool to Produce Publication-Quality Point Maps. Available online: http://www.simplemappr.net (accessed on 2 September 2019).
- Chantarasuwan, B.; Berg, C.C.; Kjellberg, F.; Rønsted, N.; Garcia, M.; Baider, C.; van Welzen, P.C. A New Classification of Ficus Subsection Urostigma (Moraceae) Based on Four Nuclear DNA Markers (ITS, ETS, G3pdh, and ncpGS), Morphology and Leaf Anatomy. PLoS ONE 2015, 10, e0128289. [Google Scholar] [CrossRef]
- Baldwin, B.G.; Markos, S. Phylogenetic utility of the external transcribed spacer (ETS) of 18S-26S rDNA: Congruence of ETS and ITS trees of Calycadenia (Compositae). Mol. Phylogenet Evol. 1998, 10, 449–463. [Google Scholar] [CrossRef]
- Taberlet, P.; Gielly, L.; Pautou, G.; Bouvet, J. Universal primers for amplification of three non-coding regions of chloroplast DNA. Plant Mol. Biol. 1991, 17, 1105–1109. [Google Scholar] [CrossRef]
- Sang, T.; Crawford, D.J.; Stuessy, T.F. Chloroplast DNA phylogeny, reticulate evolution and biogeography of Paeonia (Paeoniaceae). Am. J. Bot. 1997, 84, 1120–1136. [Google Scholar] [CrossRef] [Green Version]
- Chenna, R.; Sugawara, H.; Tadashi, K.; Lopez, R.; Gibson, T.J.; Higgins, D.G.; Thompson, J.D. Multiple sequence alignment with the Clustal series of programs. Nucleic Acids Res. 2003, 31, 3497–3500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000; pp. 1–333. [Google Scholar]
- Tamura, K. Estimation of the number of nucleotide substitutions when there are strong transition-transversion and G + C-content biases. Mol. Biol. Evol. 1992, 9, 678–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, M. A simple method for estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Zhang, J.; Chen, G.; Lu, Z.; Wang, X.; Sha, N.; Shao, B.; Li, P.; Guo, D.A. High performance liquid chromatographic method for the determination and pharmacokinetic studies of oxyresveratrol and resveratrol in rat plasma after oral administration of Smilax china extract. Biomed. Chromatogr. 2008, 22, 421–427. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characters | A. lacucha | A. thailandicus |
|---|---|---|
| Indumentum of the areoles on the lower surface of lamina | Absent | Present |
| Shape and size of staminate inflorescence | Mostly subglobose 0.6–2 cm diam. | Mostly obovoid, cylindrical, or clavate 0.2–0.4 cm diam. |
| Size of pistillate inflorescence | 1.5 cm diam. | 0.7–1 cm diam. |
| The length of peduncle of staminate inflorescence | 0.2–2 cm | Sessile or up to 0.1 cm |
| The length of peduncle of pistillate inflorescence | 0.2–4 cm | Up to 0.7 cm |
| Taxon | Floristic Region | Locality | PBM Number | Collector Number |
|---|---|---|---|---|
| A. lacucha | Central | Saraburi, Chaloem Phra Kiat, lat 14.67633° N lon 100.8868° E | 05197 | Aneklaphakij 2 |
| Central | Saraburi, Chaloem Phra Kiat, lat 14.67217° N lon 100.887° E | 05198 | Aneklaphakij 4 | |
| Central | Saraburi, Chaloem Phra Kiat, lat 14.66953° N lon 100.8898° E | 05199 | Aneklaphakij 5 | |
| Central | Nakhon Pathom, Phutthamonthon, lat 13.79072° N lon 100.317° E | 05200 | Aneklaphakij 6 | |
| Central | Nakhon Pathom, Phutthamonthon, lat 13.79072° N lon 100.317° E | 05201 | Aneklaphakij 7 | |
| North-eastern | Khon Kaen, Wiang Kao, lat 16.79369° N lon 102.2577° E | 05206 | Aneklaphakij 14 | |
| North-eastern | Khon Kaen, Wiang Kao, lat 16.79369° N lon 102.2577° E | 05207 | Aneklaphakij 15 | |
| North-eastern | Khon Kaen, Phon, lat 15.88319° N lon 102.5332° E | 05214 | Aneklaphakij 23 | |
| North-eastern | Khon Kaen, Phon, lat 15.88319° N lon 102.5332° E | 05215 | Aneklaphakij 24 | |
| North-eastern | Khon Kaen, Phon, lat 15.88319° N lon 102.5332° E | 05216 | Aneklaphakij 25 | |
| North-eastern | Khon Kaen, Phon, lat 15.88319° N lon 102.5332° E | 05217 | Aneklaphakij 26 | |
| North-eastern | Khon Kaen, Phon, lat 15.88319° N lon 102.5332° E | 05218 | Aneklaphakij 27 | |
| Central | Bangkok, Prawet, lat 13.68917° N lon 100.6596° E | 05221 | Aneklaphakij 31 | |
| Northern | Chiang Mai, Mae Rim, lat 18.89464° N lon 98.85864° E | 05223 | Aneklaphakij 36 | |
| Northern | Chiang Mai, Mae Rim, lat 18.89456° N lon 98.85869° E | 05224 | Aneklaphakij 37 | |
| Northern | Chiang Mai, Mae Rim, lat 18.89133° N lon 98.86214° E | 05225 | Aneklaphakij 38 | |
| Peninsular | Songkhla, Hat Yai, lat 7.008417° N lon 100.5059° E | 05230 | Aneklaphakij 44 | |
| Peninsular | Songkhla, Hat Yai, lat 7.008528° N lon 100.5062° E | 05231 | Aneklaphakij 47 | |
| A. thailandicus | South-western | Phetchaburi, Kaeng Krachan, lat 12.9025° N lon 99.64325° E | 05202 | Aneklaphakij 9 |
| Eastern | Ubon Ratchathani, Samrong, lat 15.01044° N lon 104.7846° E | 05203 | Aneklaphakij 10 | |
| Eastern | Ubon Ratchathani, Samrong, lat 15.01044° N lon 104.7846° E | 05204 | Aneklaphakij 11 | |
| South-eastern | Prachin Buri, Mueang Prachin Buri, lat 14.10967° N lon 101.4367° E | 05205 | Aneklaphakij 13 | |
| North-eastern | Khon Kaen, Mueang Khon Kaen, lat 16.4695° N lon 102.8271° E | 05208 | Aneklaphakij 17 | |
| North-eastern | Khon Kaen, Mueang Khon Kaen, lat 16.47194° N lon 102.8218° E | 05209 | Aneklaphakij 18 | |
| North-eastern | Khon Kaen, Mueang Khon Kaen, lat 16.47706° N lon 102.8258° E | 05210 | Aneklaphakij 19 | |
| North-eastern | Khon Kaen, Mueang Khon Kaen, lat 16.47706° N lon 102.8258° E | 05211 | Aneklaphakij 20 | |
| North-eastern | Khon Kaen, Mueang Khon Kaen, lat 16.47975° N lon 102.8276° E | 05212 | Aneklaphakij 21 | |
| North-eastern | Khon Kaen, Mueang Khon Kaen, lat 16.47447° N lon 102.8254° E | 05213 | Aneklaphakij 22 | |
| Central | Bangkok, Prawet, lat 13.68853° N lon 100.6606° E | 05219 | Aneklaphakij 28 | |
| Central | Bangkok, Prawet, lat 13.68853° N lon 100.6606° E | 05220 | Aneklaphakij 30 | |
| Northern | Chiang Mai, Mae Rim, lat 18.89742° N lon 98.85928° E | 05222 | Aneklaphakij 33 | |
| Northern | Chiang Mai, Mae Rim, lat 18.89136° N lon 98.8625° E | 05226 | Aneklaphakij 39 | |
| Northern | Chiang Mai, Mae Rim, lat 18.88869° N lon 98.86169° E | 05227 | Aneklaphakij 40 | |
| Northern | Chiang Mai, Mae Rim, lat 18.88869° N lon 98.86169° E | 05228 | Aneklaphakij 41 | |
| Northern | Chiang Mai, Mae Rim, lat 18.89° N lon 98.85842° E | 05229 | Aneklaphakij 42 |
| Region | Primer Sequence | References |
|---|---|---|
| ITS | Forward: 5’–AACAAGGTTTCCGTAGGTGA–3’ Reverse: 5’–TATGCTTAAAYTCAGCGGGT–3’ | [2] |
| ETS | Forward: 5’–GACCCTTGGTTCCTGTGTTGC–3’ Reverse: 5’–ACTTACACATGCATGGCTTAATCT–3’ | [40,41] |
| trnL-F | primer c: 5’–CGAAATCGGTAGACGCTACG–3’ primer d: 5’–GGGGATAGAGGGACTTGAAC–3’ primer e: 5’–GGTTCAAGTCCCTCTATCCC–3’ primer f: 5’–ATTTGAACTGGTGACACGAG–3’ | [42] |
| trnH-psbA | Forward: 5’–CGCGCATGGTGGATTCACAAATC–3’ Reverse: 5’–GTTATGCATGAACGTAATGCTC–3’ | [43] |
| PBM Number | ITS | ETS | trnL-F | trnH-psbA |
|---|---|---|---|---|
| 05197 | MK850217 | MK873914 | MK873937 | MK873960 |
| 05198 | MK850218 | MK873915 | MK873938 | MK873961 |
| 05199 | MH801139 | MH807231 | MH817052 | MH817064 |
| 05200 | MK850219 | MK873916 | MK873939 | MK873962 |
| 05201 | MK850220 | MK873917 | MK873940 | MK873963 |
| 05202 | MH801140 | MH807232 | MH817053 | MH817065 |
| 05203 | MH801141 | MH807233 | MH817054 | MH817066 |
| 05204 | MK850221 | MK873918 | MK873941 | MK873964 |
| 05205 | MH801142 | MH807234 | MH817055 | MH817067 |
| 05206 | MH801143 | MH807235 | MH817056 | MH817068 |
| 05207 | MK850222 | MK873919 | MK873942 | MK873965 |
| 05208 | MK850223 | MK873920 | MK873943 | MK873966 |
| 05209 | MK850224 | MK873921 | MK873944 | MK873967 |
| 05210 | MH801144 | MH807236 | MH817057 | MH817069 |
| 05211 | MK850225 | MK873922 | MK873945 | MK873968 |
| 05212 | MK850226 | MK873923 | MK873946 | MK873969 |
| 05213 | MK850227 | MK873924 | MK873947 | MK873970 |
| 05214 | MK850228 | MK873925 | MK873948 | MK873971 |
| 05215 | MK850229 | MK873926 | MK873949 | MK873972 |
| 05216 | MK850230 | MK873927 | MK873950 | MK873973 |
| 05217 | MK850231 | MK873928 | MK873951 | MK873974 |
| 05218 | MH801145 | MH807237 | MH817058 | MH817070 |
| 05219 | MH801146 | MH807238 | MH817059 | MH817071 |
| 05220 | MK850232 | MK873929 | MK873952 | MK873975 |
| 05221 | MH801147 | MH807239 | MH817060 | MH817072 |
| 05222 | MK850233 | MK873930 | MK873953 | MK873976 |
| 05223 | MK850234 | MK873931 | MK873954 | MK873977 |
| 05224 | MK850235 | MK873932 | MK873955 | MK873978 |
| 05225 | MH801148 | MH807240 | MH817061 | MH817073 |
| 05226 | MK850236 | MK873933 | MK873956 | MK873979 |
| 05227 | MK850237 | MK873934 | MK873957 | MK873980 |
| 05228 | MK850238 | MK873935 | MK873958 | MK873981 |
| 05229 | MH801149 | MH807241 | MH817062 | MH817074 |
| 05230 | MK850239 | MK873936 | MK873959 | MK873982 |
| 05231 | MH801150 | MH807242 | MH817063 | MH817075 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aneklaphakij, C.; Bunsupa, S.; Sirichamorn, Y.; Bongcheewin, B.; Satitpatipan, V. Taxonomic Notes on the ‘Mahat’ (Artocarpus lacucha and A. thailandicus, Moraceae) Species Complex in Thailand. Plants 2020, 9, 391. https://doi.org/10.3390/plants9030391
Aneklaphakij C, Bunsupa S, Sirichamorn Y, Bongcheewin B, Satitpatipan V. Taxonomic Notes on the ‘Mahat’ (Artocarpus lacucha and A. thailandicus, Moraceae) Species Complex in Thailand. Plants. 2020; 9(3):391. https://doi.org/10.3390/plants9030391
Chicago/Turabian StyleAneklaphakij, Chaiwat, Somnuk Bunsupa, Yotsawate Sirichamorn, Bhanubong Bongcheewin, and Veena Satitpatipan. 2020. "Taxonomic Notes on the ‘Mahat’ (Artocarpus lacucha and A. thailandicus, Moraceae) Species Complex in Thailand" Plants 9, no. 3: 391. https://doi.org/10.3390/plants9030391
APA StyleAneklaphakij, C., Bunsupa, S., Sirichamorn, Y., Bongcheewin, B., & Satitpatipan, V. (2020). Taxonomic Notes on the ‘Mahat’ (Artocarpus lacucha and A. thailandicus, Moraceae) Species Complex in Thailand. Plants, 9(3), 391. https://doi.org/10.3390/plants9030391