Development and Characterization of an Ethyl Methane Sulfonate (EMS) Induced Mutant Population in Capsicum annuum L.

 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Material and Methods
2.1. Plant Material
2.2. Mutagenesis
2.3. Construction of M1, M2, and M3 Generations
2.4. Phenotype Evaluation of M2 Plants
2.5. TILLING Experiment
3. Results
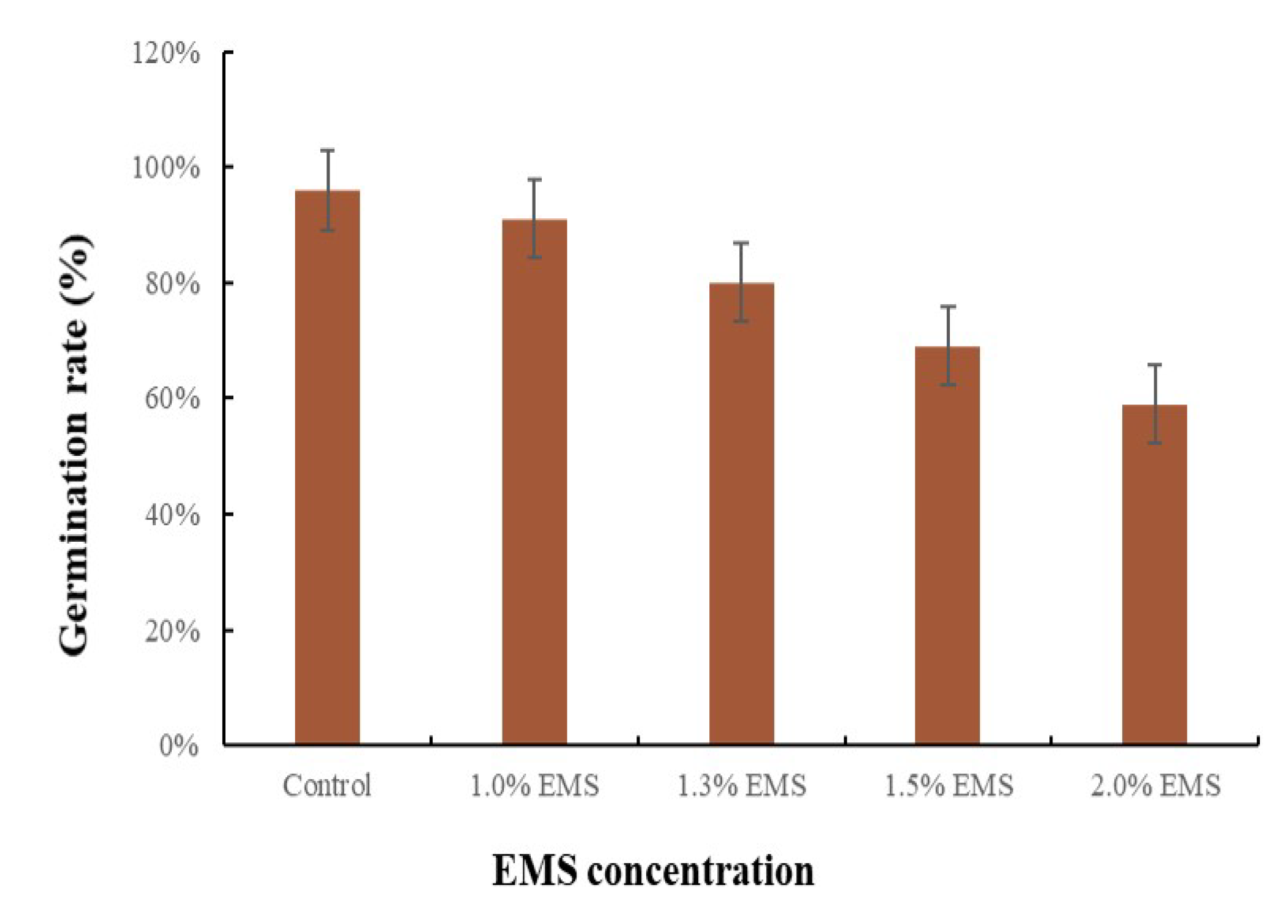
3.1. Effect of EMS dose on Seed Germination
3.2. Phenotypes of M1 Mutant Plants
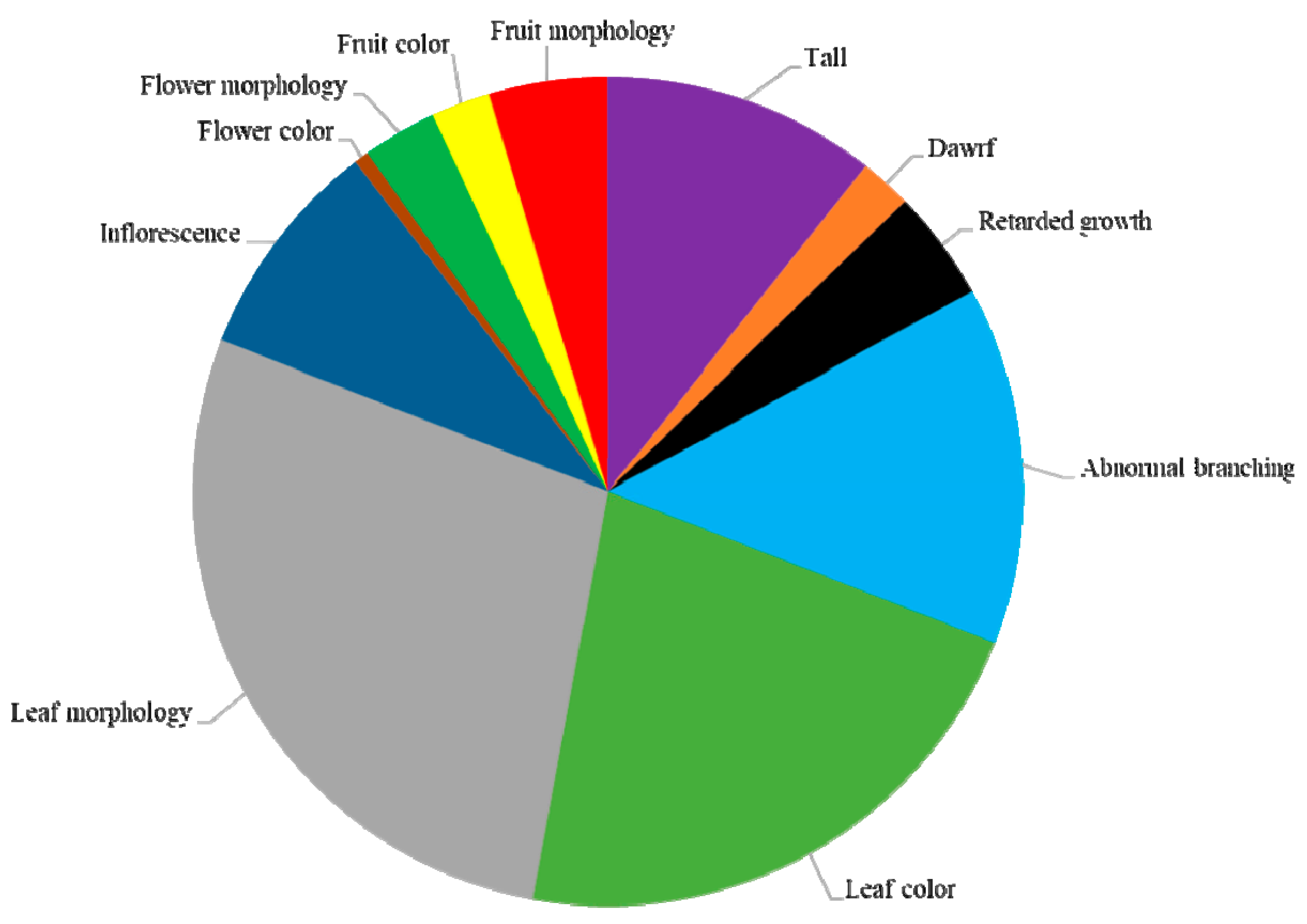
3.3. Characterization of M2 Mutant Phenotypes
3.4. Plant Growth and Habit Phenotypes
3.5. Leaf Color and Morphology Phenotypes
3.6. Flower Color and Morphology Phenotypes
3.7. Fruit Color and Morphology
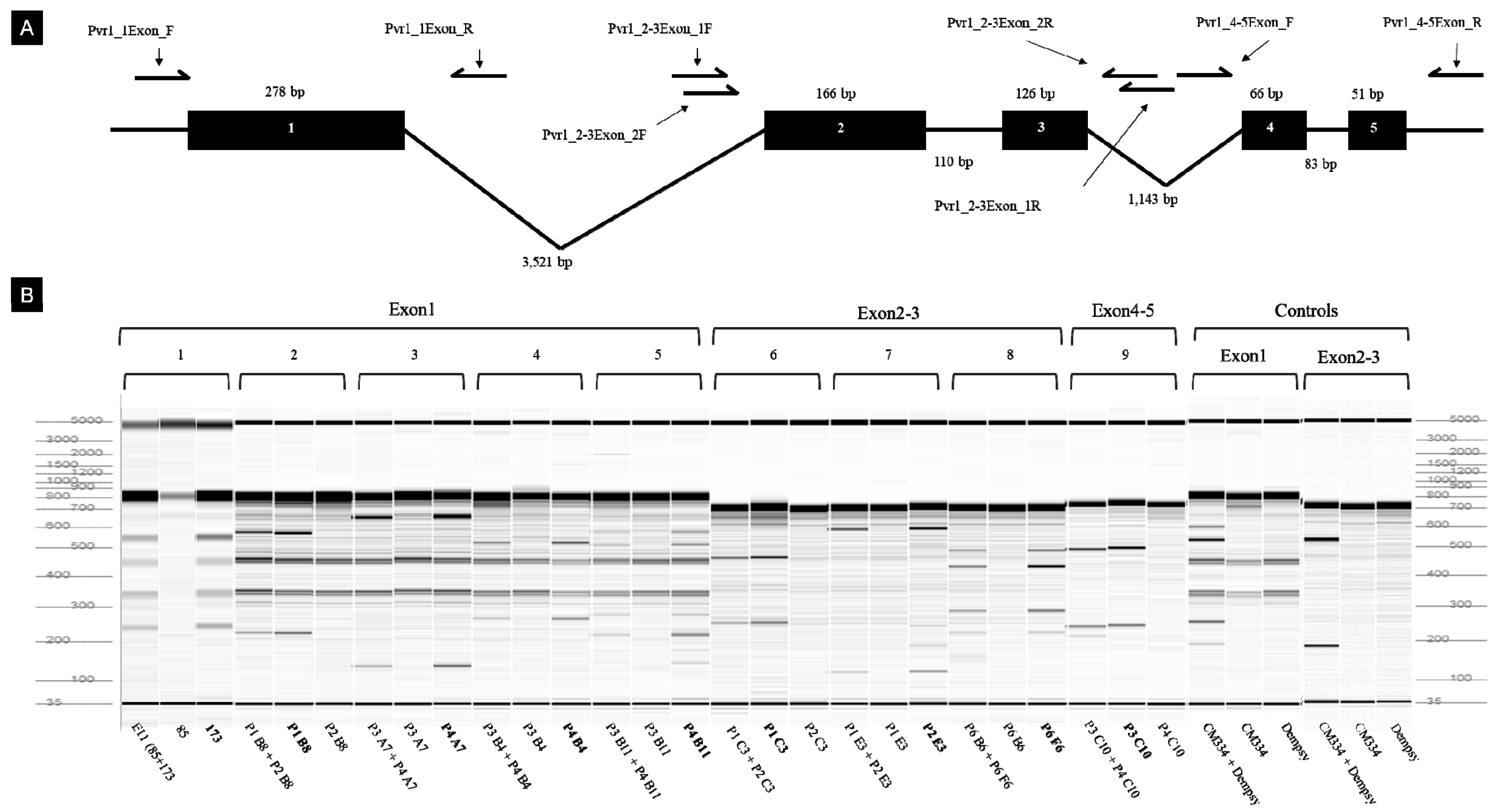
3.8. Mutation Screening and Detection by TILLING
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Bosland, P.W.; Votava, E.J.; Votava, E.M. Peppers: Vegetable and Spice Capsicums; CABI: Wallingford, UK, 2012; Volume 22. [Google Scholar]
- Siddique, M.I.; Lee, H.Y.; Ro, N.Y.; Han, K.; Venkatesh, J.; Solomon, A.M.; Patil, A.S.; Changkwian, A.; Kwon, J.K.; Kang, B.C. Identifying candidate genes for Phytophthora capsici resistance in pepper (Capsicum annuum) via genotyping-by-sequencing-based QTL mapping and genome-wide association study. Sci. Rep. 2019, 9, 1–15. [Google Scholar] [CrossRef]
- Jeong, H.J.; Kwon, J.K.; Pandeya, D.; Hwang, J.; Hoang, N.H.; Bae, J.H.; Kang, B.C. A survey of natural and ethyl methane sulfonate-induced variations of eIF4E using high-resolution melting analysis in Capsicum. Mol. Breed. 2012, 29, 349–360. [Google Scholar] [CrossRef]
- Jo, Y.D.; Kim, S.H.; Hwang, J.E.; Kim, Y.S.; Kang, H.S.; Kim, S.W.; Kwon, S.J.; Ryu, J.; Kim, J.B.; Kang, S.Y. Construction of mutation populations by gamma-ray and carbon beam irradiation in chili pepper (Capsicum annuum L.). Hortic. Environ. Biotechnol. 2016, 57, 606–614. [Google Scholar] [CrossRef]
- Lee, H.Y.; Ro, N.Y.; Jeong, H.J.; Kwon, J.K.; Jo, J.; Ha, Y.; Jung, A.; Han, J.W.; Venkatesh, J.; Kang, B.C. Genetic diversity and population structure analysis to construct a core collection from a large Capsicum germplasm. BMC Genet. 2016, 17, 142. [Google Scholar] [CrossRef] [Green Version]
- Solomon, A.M.; Han, K.; Lee, J.H.; Lee, H.Y.; Jang, S.; Kang, B.C. Genetic diversity and population structure of Ethiopian Capsicum germplasms. PLoS ONE 2019, 14, e0216886. [Google Scholar] [CrossRef]
- Hwang, D.; Jeong, H.J.; Kwon, J.K.; Kim, H.; Kang, S.Y.; Kang, B.C. Phenotypic variants among ethyl methanesulfonate M 2 mutant lines in Capsicum annuum. Plant Genet. Resour. 2014, 12, S141–S145. [Google Scholar] [CrossRef]
- Espina, M.J.; Ahmed, C.M.; Bernardini, A.; Adeleke, E.; Yadegari, Z.; Arelli, P.; Pantalone, V.; Taheri, A. Development and phenotypic screening of an ethyl methane sulfonate mutant population in soybean. Front. Plant Sci. 2018, 9, 394. [Google Scholar] [CrossRef] [Green Version]
- Voytas, D.F.; Gao, C. Precision genome engineering and agriculture: Opportunities and regulatory challenges. PLoS Biol. 2014, 12, e1001877. [Google Scholar] [CrossRef]
- Lu, S.; Yin, X.; Spollen, W.; Zhang, N.; Xu, D.; Schoelz, J.; Bilyeu, K.; Zhang, Z.J. Analysis of the siRNA-mediated gene silencing process targeting three homologous genes controlling soybean seed oil quality. PLoS ONE 2015, 10, e0129010. [Google Scholar] [CrossRef] [Green Version]
- Daskalov, S. Mutation breeding in pepper. IAEA 1986, 4, 25. [Google Scholar]
- Jo, Y.D.; Kim, J.B. Frequency and spectrum of radiation-induced mutations revealed by whole-genome sequencing analyses of plants. Quantum Beam Sci. 2019, 3, 7. [Google Scholar] [CrossRef] [Green Version]
- Talebi, A.B.; Talebi, A.B.; Shahrokhifar, B. Ethyl methane sulphonate (EMS) induced mutagenesis in Malaysian rice (cv. MR219) for lethal dose determination. Am. J. Plant Sci 2012, 3, 1661–1665. [Google Scholar] [CrossRef] [Green Version]
- Arisha, M.H.; Liang, B.K.; Shah, S.N.M.; Gong, Z.H.; Li, D.W. Kill curve analysis and response of first generation Capsicum annuum L. B12 cultivar to ethyl methane sulfonate. Genet. Mol. Res. 2014, 13, 10049–10061. [Google Scholar] [CrossRef]
- Piron, F.; Nicolai, M.; Minoia, S.; Piednoir, E.; Moretti, A.; Salgues, A.; Zamir, D.; Caranta, C.; Bendahmane, A. An induced mutation in tomato eIF4E leads to immunity to two potyviruses. PLoS ONE 2010, 5, e13313. [Google Scholar] [CrossRef] [Green Version]
- Gauffier, C.; Lebaron, C.; Moretti, A.; Constant, C.; Moquet, F.; Bonnet, G.; Caranta, C.; Gallois, J.L. A TILLING approach to generate broad-spectrum resistance to potyviruses in tomato is hampered by eIF4E gene redundancy. Plant J. 2016, 85, 717–729. [Google Scholar] [CrossRef] [Green Version]
- Xi-ou, X.; Wenqiu, L.; Wei, L.; Xiaoming, G.; Lingling, L.; Feiyue, M.; Yuge, L. The analysis of physiological variations in M2 generation of Solanum melongena L. Mutagenized by ethyl methane sulfonate. Front. Plant Sci. 2017, 8, 17. [Google Scholar] [CrossRef] [Green Version]
- Jabeen, N.; Mirza, B. Ethyl methane sulfonate enhances genetic variability in Capsicum annuum. Asian J. Plant Sci. 2002, 1, 425–428. [Google Scholar]
- Bosland, P.W. Inheritance of a novel flaccid mutant in Capsicum annuum. J. Hered. 2002, 93, 380–382. [Google Scholar] [CrossRef] [Green Version]
- Paran, I.; Borovsky, Y.; Nahon, S.; Cohen, O. The use of induced mutations to study shoot architecture in Capsicum. Isr. J. Plant Sci. 2007, 55, 125–131. [Google Scholar] [CrossRef]
- Arisha, M.H.; Shah, S.N.M.; Gong, Z.H.; Jing, H.; Li, C.; Zhang, H.X. Ethyl methane sulfonate induced mutations in M2 generation and physiological variations in M1 generation of peppers (Capsicum annuum L.). Front. Plant Sci. 2015, 6, 399. [Google Scholar] [CrossRef] [Green Version]
- Chaim, A.B.; Borovsky, Y.; Rao, G.U.; Gur, A.; Zamir, D.; Paran, I. Comparative QTL mapping of fruit size and shape in tomato and pepper. Isr. J. Plant Sci. 2006, 54, 191–203. [Google Scholar] [CrossRef]
- Oleykowski, C.A.; Bronson Mullins, C.R.; Godwin, A.K.; Yeung, A.T. Mutation detection using a novel plant endonuclease. Nucleic Acids Res. 1998, 26, 4597–4602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slade, A.J.; Fuerstenberg, S.I.; Loeffler, D.; Steine, M.N.; Facciotti, D. A reverse genetic, nontransgenic approach to wheat crop improvement by TILLING. Nat. Biotechnol. 2005, 23, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Till, B.J.; Reynolds, S.H.; Greene, E.A.; Codomo, C.A.; Enns, L.C.; Johnson, J.E.; Burtner, C.; Odden, A.R.; Young, K.; Taylor, N.E. Large-scale discovery of induced point mutations with high-throughput TILLING. Genome Res. 2003, 13, 524–530. [Google Scholar] [CrossRef] [Green Version]
- Henikoff, S.; Till, B.J.; Comai, L. TILLING. Traditional mutagenesis meets functional genomics. Plant Physiol. 2004, 135, 630–636. [Google Scholar] [CrossRef] [Green Version]
- Page, D.R.; Grossniklaus, U. The art and design of genetic screens: Arabidopsis thaliana. Nat. Rev. Genet. 2002, 3, 124–136. [Google Scholar] [CrossRef]
- Østergaard, L.; Yanofsky, M.F. Establishing gene function by mutagenesis in Arabidopsis thaliana. Plant J. 2004, 39, 682–696. [Google Scholar] [CrossRef]
- Menda, N.; Semel, Y.; Peled, D.; Eshed, Y.; Zamir, D. In silico screening of a saturated mutation library of tomato. Plant J. 2004, 38, 861–872. [Google Scholar] [CrossRef]
- Martí, E.; Gisbert, C.; Bishop, G.J.; Dixon, M.S.; García-Martínez, J.L. Genetic and physiological characterization of tomato cv. Micro-Tom. J. Exp. Bot. 2006, 57, 2037–2047. [Google Scholar]
- Gady, A.L.F.; Hermans, F.W.K.; Van de Wal, M.H.B.J.; van Loo, E.N.; Visser, R.G.F.; Bachem, C.W.B. Implementation of two high through-put techniques in a novel application: Detecting point mutations in large EMS mutated plant populations. Plant Methods 2009, 5, 13. [Google Scholar] [CrossRef] [Green Version]
- Minoia, S.; Petrozza, A.; D’Onofrio, O.; Piron, F.; Mosca, G.; Sozio, G.; Cellini, F.; Bendahmane, A.; Carriero, F. A new mutant genetic resource for tomato crop improvement by TILLING technology. BMC Res. Notes 2010, 3, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, S.N.; Gong, Z.H.; Arisha, M.H.; Khan, A.; Tian, S.L. Effect of ethyl methyl sulfonate concentration and different treatment conditions on germination and seedling growth of the cucumber cultivar Chinese long (9930). Genet. Mol. Res. 2015, 14, 2440–2449. [Google Scholar] [CrossRef] [PubMed]
- Sikder, S.; Biswas, P.; Hazra, P.; Akhtar, S.; Chattopadhyay, A.; Badigannavar, A.M.; D’Souza, S.F. Induction of mutation in tomato (Solanum lycopersicum L.) by gamma irradiation and EMS. Indian J. Genet. Plant Breed. 2013, 73, 392–399. [Google Scholar] [CrossRef]
- Alcantara, T.P.; Bosland, P.W.; Smith, D.W. Ethyl methanesulfonate-induced seed mutagenesis of Capsicum annuum. J. Hered. 1996, 87, 239–241. [Google Scholar] [CrossRef] [Green Version]
- Roychowdhury, R.; Tah, J. Mutagenesis—A potential approach for crop improvement. In Crop Improvement; Springer: Berlin/Heidelberg, Germany, 2013; pp. 149–187. [Google Scholar]
- Saito, T.; Ariizumi, T.; Okabe, Y.; Asamizu, E.; Hiwasa-Tanase, K.; Fukuda, N.; Mizoguchi, T.; Yamazaki, Y.; Aoki, K.; Ezura, H. TOMATOMA: A novel tomato mutant database distributing Micro-Tom mutant collections. Plant Cell Physiol. 2011, 52, 283–296. [Google Scholar] [CrossRef] [Green Version]
- Lippert, L.F.; Bergh, B.O.; Cook, A.A. Three variegated seedling mutants in the pepper: Multiple allelism indicated in crossing studies. J. Hered. 1964, 55, 79–83. [Google Scholar] [CrossRef]
- Fridborg, I.; Kuusk, S.; Moritz, T.; Sundberg, E. The Arabidopsis dwarf mutant shi exhibits reduced gibberellin responses conferred by overexpression of a new putative zinc finger protein. Plant Cell 1999, 11, 1019–1031. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Bosland, P.W. The genes of Capsicum. HortScience 2006, 41, 1169–1187. [Google Scholar] [CrossRef]
- Wu, Z.; Zhang, X.; He, B.; Diao, L.; Sheng, S.; Wang, J.; Guo, X.; Su, N.; Wang, L.; Jiang, L. A chlorophyll-deficient rice mutant with impaired chlorophyllide esterification in chlorophyll biosynthesis. Plant Physiol. 2007, 145, 29–40. [Google Scholar] [CrossRef] [Green Version]
- Coschigano, K.T.; Melo-Oliveira, R.; Lim, J.; Coruzzi, G.M. Arabidopsis gls mutants and distinct Fd-GOGAT genes: Implications for photorespiration and primary nitrogen assimilation. Plant Cell 1998, 10, 741–752. [Google Scholar] [CrossRef] [Green Version]
- Murchie, E.H.; Chen, Y.Z.; Hubbart, S.; Peng, S.; Horton, P. Interactions between senescence and leaf orientation determine in situ patterns of photosynthesis and photoinhibition in field-grown rice. Plant Physiol. 1999, 119, 553–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yanofsky, M.F.; Ma, H.; Bowman, J.L.; Drews, G.N.; Feldmann, K.A.; Meyerowitz, E.M. The protein encoded by the Arabidopsis homeotic gene agamous resembles transcription factors. Nature 1990, 346, 35–39. [Google Scholar] [CrossRef]
- Shifriss, C. Male sterility in pepper (Capsicum annuum L.). Euphytica 1997, 93, 83–88. [Google Scholar] [CrossRef]
- Siddique, M.I.; Wai, K.P.P.; Mo, H.S.; Yoo, H.J.; Jang, K.S.; Jeon, S.G.; Hwang, J.E.; Kim, B.S. Resistance to Phytophthora capsici, restorer-of-fertility genotype for cytoplasmic male sterility and chemical quality components of breeding lines developed for improvement of subicho, a land race of pepper in Yeongyang. Hort. Sci. Technol. 2017, 35, 758–768. [Google Scholar]
- Burton, G.W.; Hanna, W.W. Stable cytoplasmic male-sterile mutants induced in Tift 23DB1 pearl millet with mitomycin and streptomycin 1. Crop Sci. 1982, 22, 651–652. [Google Scholar] [CrossRef]
- Kinoshita, T.; Takahashi, M.E.; Mikami, T. Cytoplasmic mutation of male sterility induced by chemical mutagens in sugar beets. Proc. Jpn. Acad. Ser. B 1982, 58, 319–322. [Google Scholar] [CrossRef] [Green Version]
- Jeong, H.B.; Kang, M.Y.; Jung, A.; Han, K.; Lee, J.H.; Jo, J.; Lee, H.Y.; An, J.W.; Kim, S.; Kang, B.C. Single-molecule real-time sequencing reveals diverse allelic variations in carotenoid biosynthetic genes in pepper (Capsicum spp.). Plant Biotechnol. J. 2019, 17, 1081–1093. [Google Scholar] [CrossRef] [Green Version]
- Till, B.J.; Zerr, T.; Comai, L.; Henikoff, S. A protocol for TILLING and Ecotilling in plants and animals. Nat. Protoc. 2006, 1, 2465. [Google Scholar] [CrossRef] [Green Version]
- Amri-Tiliouine, W.; Laouar, M.; Abdelguerfi, A.; Jankowicz-Cieslak, J.; Jankuloski, L.; Till, B.J. Genetic variability induced by gamma rays and preliminary results of low-cost TILLING on M2 generation of Chickpea (Cicer arietinum L.). Front. Plant Sci. 2018, 9, 1568. [Google Scholar] [CrossRef] [Green Version]
- Sega, G.A. A review of the genetic effects of ethyl methanesulfonate. Mutat. Res. Rev. Genet. Toxicol. 1984, 134, 113–142. [Google Scholar] [CrossRef]
- Kurowska, M.; Daszkowska-Golec, A.; Gruszka, D.; Marzec, M.; Szurman, M.; Szarejko, I.; Maluszynski, M. TILLING-a shortcut in functional genomics. J. Appl. Genet. 2011, 52, 371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, H.; Howell, T.; Nitcher, R.; Missirian, V.; Watson, B.; Ngo, K.J.; Lieberman, M.; Fass, J.; Uauy, C.; Tran, R.K. Discovery of rare mutations in populations: TILLING by sequencing. Plant Physiol. 2011, 156, 1257–1268. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Abernathy, B.; Zeng, Y.; Ozias-Akins, P. TILLING by sequencing to identify induced mutations in stress resistance genes of peanut (Arachis hypogaea). BMC Genom. 2015, 16, 157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, P.; Reddaiah, B.; Salava, H.; Upadhyaya, P.; Tyagi, K.; Sarma, S.; Datta, S.; Malhotra, B.; Thomas, S.; Sunkum, A. Next-generation sequencing (NGS)-based identification of induced mutations in a doubly mutagenized tomato (Solanum lycopersicum) population. Plant J. 2017, 92, 495–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5ʹ->3ʹ) |
|---|---|
| Pvr1_Exon1_F | AAAGCACACAGCACCAACAA |
| Pvr1_Exon1_R | CAACCATAAATATACCCCGAGAA |
| Pvr1_Exon2-3_1F | GCGCTTTGAGTTATCGTACA |
| Pvr1_Exon2-3_1R | AGAGTGAGGCGAAGTTTGAG |
| Pvr1_Exon2-3_2F | GAGTTATCGTACAACTTGGACTGTG |
| Pvr1_Exon2-3_2R | GATATAACTGATGACTTCCCCTCTT |
| Pvr1_Exon4-5_F | GGTGATGTCGTTTTGTCTCA |
| Pvr1_Exon4-5_R | TCTGTGATTTGATGTGTGCTC |
| Treatments | Total No. of Seeds | Germinated Seeds | Seed Germination (%) | Surviving Seedlings | Surviving up to Seed Harvest |
|---|---|---|---|---|---|
| EMS | 9500 | 6620 | 70% | 4210 | 3996 |
| Control | 200 | 175 | 87% | 165 | 157 |
| Categories | Number of Plants | Percentage (%) |
|---|---|---|
| Plant height (tall v) | 45 | 1.12% |
| Plant height (dwarf w) | 18 | 0.51% |
| Albino x | 23 | 0.61% |
| No flower y | 231 | 5.81% |
| Sterile z | 41 | 1.02% |
| Class | Sub-Class | Description | No. of Mutant Lines | Rate (%) | Segregation Mutant: Wild Type * |
|---|---|---|---|---|---|
| Plant growth | Tall | >18 cm | 37 | 9.9 | 3:7 |
| Dwarf | <10 cm | 7 | 1.9 | 3:7 | |
| Retarded growth | No visible stem | 15 | 4.0 | 3:7 | |
| Abnormal branching | Spreading multi-whorled canopy | 48 | 12.9 | 3:7 | |
| Leaf | Color | Mottled pale-green | 18 | 4.8 | 2:8 |
| Light-green | 6 | 1.6 | 3:7 | ||
| Mottled-yellow | 7 | 1.9 | 2:8 | ||
| Dark-green | 13 | 3.5 | 3:7 | ||
| Pale-green | 29 | 7.8 | 3:7 | ||
| Silver-green | 3 | 0.8 | 1:9 | ||
| Morphology | Leathery texture | 8 | 2.2 | 2:8 | |
| Undulation | 5 | 1.3 | 2:8 | ||
| Upward curling | 7 | 1.1 | 3:7 | ||
| Downward curling | 4 | 1.9 | 3:7 | ||
| Bushy | 3 | 0.8 | 2:8 | ||
| Obovate shape | 2 | 0.5 | 1:9 | ||
| Long petiole | 22 | 5.9 | 3:7 | ||
| Fused | 1 | 0.3 | 2:8 | ||
| Prolific | 11 | 3.0 | 2:8 | ||
| Large size | 19 | 5.1 | 3:7 | ||
| Scabrous | 12 | 3.2 | 1:9 | ||
| Narrow | 9 | 2.4 | 1:9 | ||
| Flower | Inflorescence | Flowerless | 27 | 7.3 | 4:6 |
| Twin flower | 3 | 0.8 | 1:9 | ||
| Sterile | 21 | 5.6 | 2:8 | ||
| Organ color | Pink anther | 1 | 0.3 | 1:9 | |
| Morphology | Filamentous stamen | 1 | 0.3 | 3:7 | |
| AGAMOUS type | 4 | 1.1 | 3:7 | ||
| Fasciculation type | 1 | 0.3 | 2:8 | ||
| Swelled ovary with short stamen | 3 | 0.8 | 2:8 | ||
| Shell type | 1 | 0.3 | 1:9 | ||
| Fruits | Color | Orange mature color | 7 | 1.9 | 2:8 |
| Two different mature colors (red and yellow) | 1 | 0.3 | 2:8 | ||
| Morphology | Round shape | 5 | 1.3 | 3:7 | |
| Cylindrical shape | 2 | 0.5 | 1:9 | ||
| Oval shape | 3 | 0.8 | 2:8 | ||
| Multiple fruit on a single calyx | 1 | 0.3 | 1:9 | ||
| Pyramid shape | 4 | 1.1 | 2:8 | ||
| Two shapes on one plant | 1 | 0.3 | 2:8 | ||
| Total | 372 | 100 |
| Mutant | Position of Mutation Detection | Mutant Population | Mutation Confirmed by Sanger Sequencing | Mutation Type |
|---|---|---|---|---|
| 173 | Pvr1_1Exon1 | Micro-Pep | No variation | - |
| P1B8 | Pvr1_1Exon1 | Yuwol-cho | 546th bp | Intron mutation |
| P4A7 | Pvr1_1Exon1 | Yuwol-cho | 628th bp | Intron mutation |
| P4B4 | Pvr1_1Exon1 | Yuwol-cho | 504th bp | Intron mutation |
| P4B11 | Pvr1_1Exon1 | Yuwol-cho | No variation | - |
| P1C3 | Pvr1_2-3Exon2-3 | Yuwol-cho | No variation | - |
| P2E3 | Pvr1_2-3Exon2-3 | Yuwol-cho | No variation | - |
| P6F6 | Pvr1_2-3Exon2-3 | Micro-Pep | No variation | - |
| P3C10 | Pvr1_4-5Exon4-5 | Yuwol-cho | 5261st bp | Intron mutation |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siddique, M.I.; Back, S.; Lee, J.-H.; Jo, J.; Jang, S.; Han, K.; Venkatesh, J.; Kwon, J.-K.; Jo, Y.D.; Kang, B.-C. Development and Characterization of an Ethyl Methane Sulfonate (EMS) Induced Mutant Population in Capsicum annuum L. Plants 2020, 9, 396. https://doi.org/10.3390/plants9030396
Siddique MI, Back S, Lee J-H, Jo J, Jang S, Han K, Venkatesh J, Kwon J-K, Jo YD, Kang B-C. Development and Characterization of an Ethyl Methane Sulfonate (EMS) Induced Mutant Population in Capsicum annuum L. Plants. 2020; 9(3):396. https://doi.org/10.3390/plants9030396
Chicago/Turabian StyleSiddique, Muhammad Irfan, Seungki Back, Joung-Ho Lee, Jinkwan Jo, Siyoung Jang, Koeun Han, Jelli Venkatesh, Jin-Kyung Kwon, Yeong Deuk Jo, and Byoung-Cheorl Kang. 2020. "Development and Characterization of an Ethyl Methane Sulfonate (EMS) Induced Mutant Population in Capsicum annuum L." Plants 9, no. 3: 396. https://doi.org/10.3390/plants9030396
APA StyleSiddique, M. I., Back, S., Lee, J. -H., Jo, J., Jang, S., Han, K., Venkatesh, J., Kwon, J. -K., Jo, Y. D., & Kang, B. -C. (2020). Development and Characterization of an Ethyl Methane Sulfonate (EMS) Induced Mutant Population in Capsicum annuum L. Plants, 9(3), 396. https://doi.org/10.3390/plants9030396