Leaf Water Relations in Lime Trees Grown under Shade Netting and Open-Air


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Meteorological Conditions
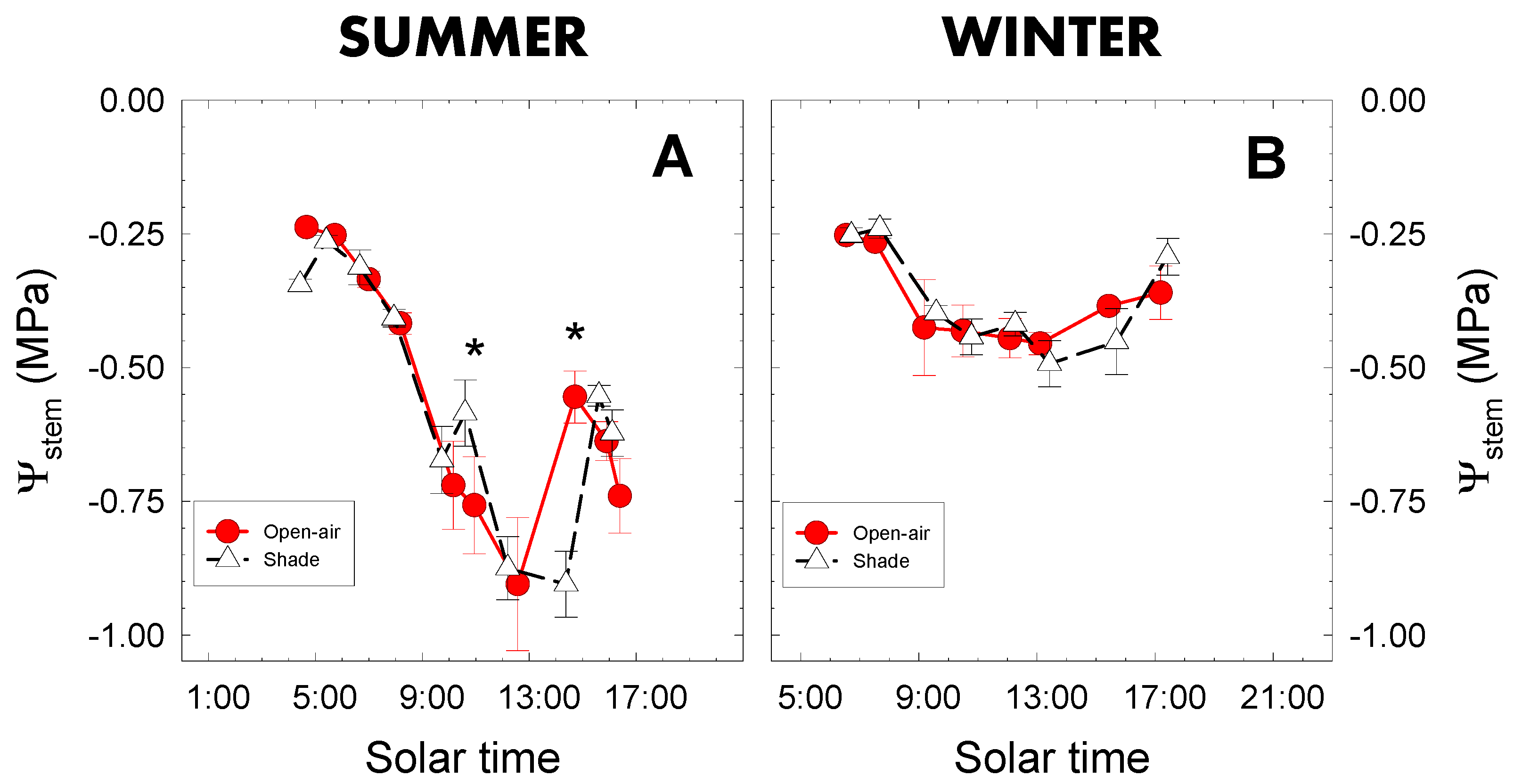
2.2. Stem Water Potential
2.3. Gas Exchange
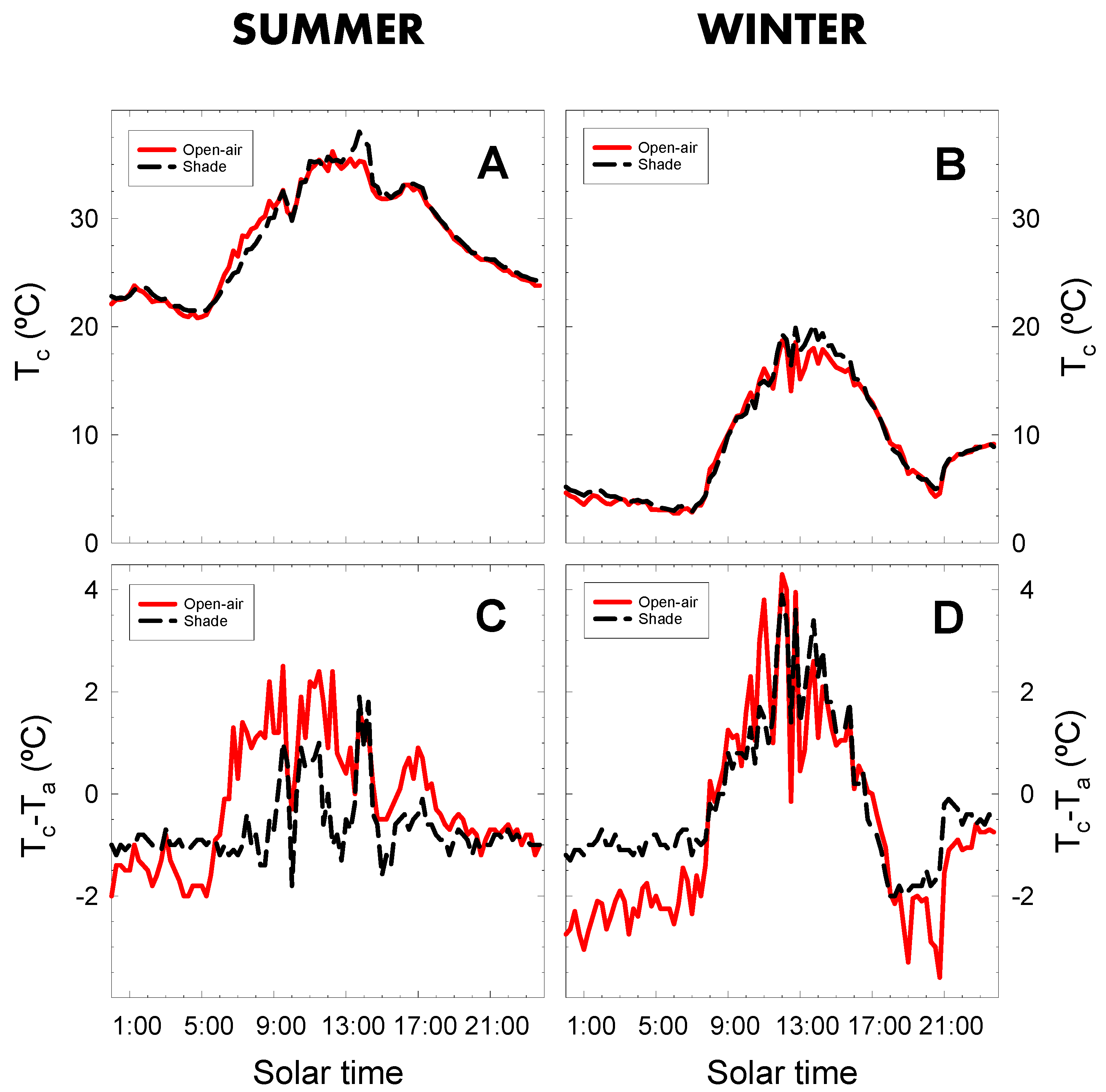
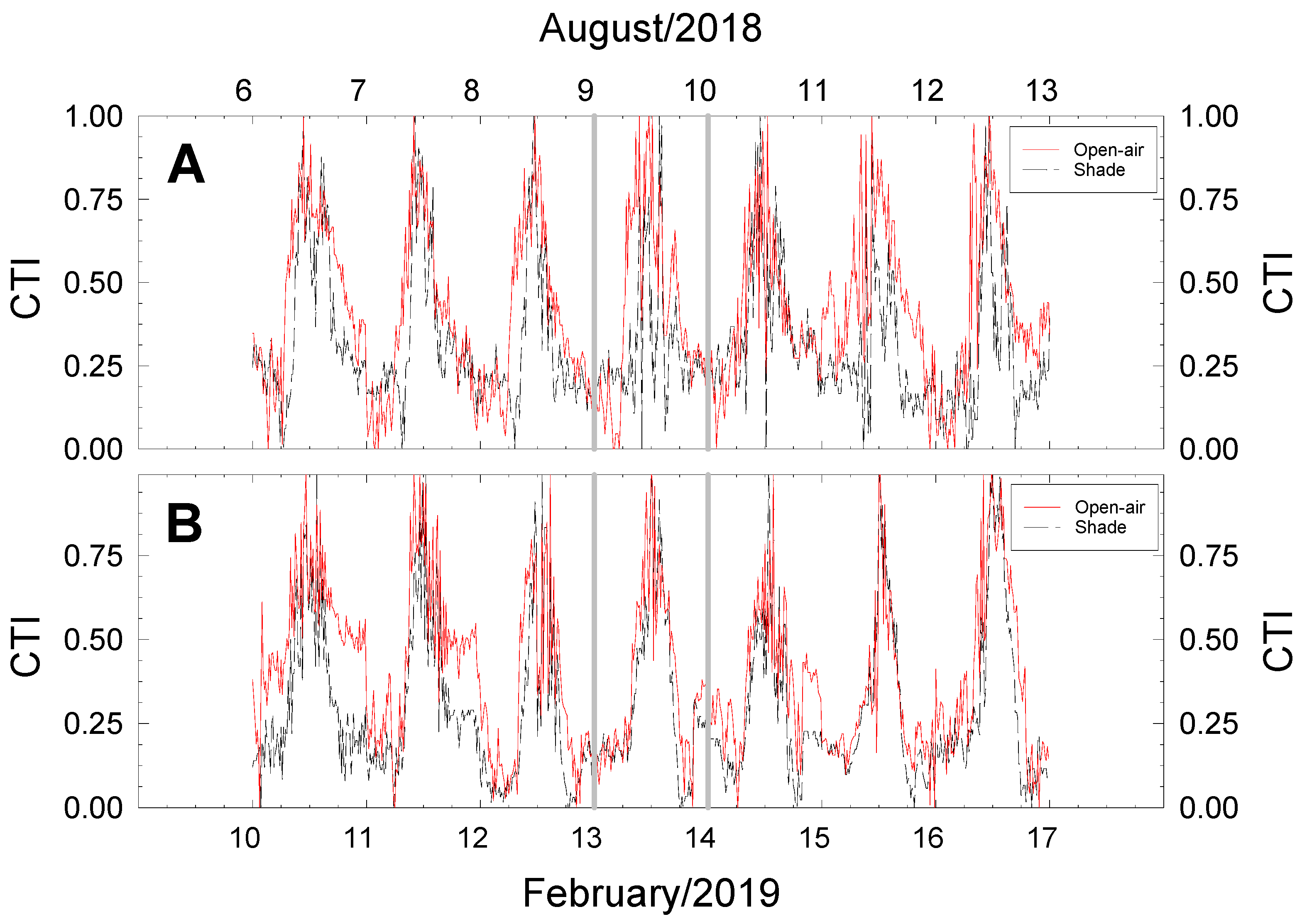
2.4. Canopy Temperature
3. Materials and Methods
3.1. Plant Material and Experimental Conditions
3.2. Cropping Conditions
3.3. Measurements
3.4. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Food and Agriculture Organization of the United Nations (FAO). Citrus Fruit: Fresh and Processed. Statistical Bulletin. 2017. Available online: http://www.fao.org/economic/est/est-commodities/citricos/es (accessed on 17 December 2019).
- Ministerio de Agricultura Pesca y Alimentación (MAPA). Anuario de Estadística Avanzada. 2018. Available online: https://www.mapa.gob.es/es/estadistica/temas/publicaciones/anuario-de-estadistica/2018/default.aspx (accessed on 17 December 2019).
- Consejería de Agua, Agricultura, Ganadería, P. y M. (CARM). Estadística Agrarian. 2018. Available online: https://www.carm.es/web/pagina?IDCONTENIDO=1392&IDTIPO=100&RASTRO=c1415$m1174 (accessed on 17 December 2019).
- Centro Regional de Estadística de Murcia (CREM). Estadísticas Agricultura, Ganadería, Selvicultura y Pesca. Evolución de la Producción Agrícola Según Tipo de Cultivo. 2018. Available online: http://econet.carm.es/web/crem/inicio/-/crem/sicrem/PU590/sec23.html (accessed on 8 January 2020).
- Intergovernmental Panel on Climate Change (IPCC). Special Report: Global Warming of 1.5 °C. Available online: https://www.ipcc.ch/sr15/ (accessed on 8 January 2020).
- Fernández, J.E. Plant-based methods for irrigation scheduling of woody crops. Horticulturae 2017, 3, 35. [Google Scholar] [CrossRef] [Green Version]
- Jones, H.G. Thermal imaging and infrared sensing in plant ecophysiology. Adv. Plant Ecophysiol. Tech. 2004, 41, 107–163. [Google Scholar]
- Abdelfatah, A.; Aranda, X.; Savé, R.; de Herralde, F.; Biel, C. Evaluation of the response of maximum daily shrinkage in young cherry trees submitted to water stress cycles in a greenhouse. Agric. Water Manag. 2013, 118, 150–158. [Google Scholar] [CrossRef]
- Blanco, V.; Domingo, R.; Pérez-Pastor, A.; Blaya-Ros, P.J.; Torres-Sánchez, R. Soil and plant water indicators for deficit irrigation management of field-grown sweet cherry trees. Agric. Water Manag. 2018, 208, 83–94. [Google Scholar] [CrossRef]
- Celedón, J.M.; Gil, P.M.; Ferreyra, R.; Maldonado, P.; Barrera, C. Sensitivity and variability of two plant water stress indicators: Exploring criteria for choosing a plant monitoring method for avocado irrigation management. Chil. J. Agric. Res. 2012, 72, 379–387. [Google Scholar] [CrossRef] [Green Version]
- Egea, G.; Pagán, E.; Baille, A.; Domingo, R.; Nortes, P.A.; Pérez-Pastor, A. Usefulness of establishing trunk diameter based reference lines for irrigation scheduling in almond trees. Irrig. Sci. 2009, 27, 431–441. [Google Scholar] [CrossRef]
- De la Rosa, J.M.; Conesa, M.R.; Domingo, R.; Torres, R.; Pérez-Pastor, A. Feasibility of using trunk diameter fluctuation and stem water potential reference lines for irrigation scheduling of early nectarine trees. Agric. Water Manag. 2013, 126, 133–141. [Google Scholar] [CrossRef]
- Bellvert, J.; Marsal, J.; Girona, J.; Gonzalez-Dugo, V.; Fereres, E.; Ustin, S.L.; Zarco-Tejada, P.J. Airborne thermal imagery to detect the seasonal evolution of crop water status in peach, nectarine and Saturn peach orchards. Remote Sens. 2016, 8, 39. [Google Scholar] [CrossRef] [Green Version]
- Ortuño, M.F.; García-Orellana, Y.; Conejero, W.; Ruiz-Sánchez, M.C.; Alarcón, J.J.; Torrecillas, A. Stem and leaf water potentials, gas exchange, sap flow, and trunk diameter fluctuations for detecting water stress in lemon trees. Trees 2005, 20, 1–8. [Google Scholar] [CrossRef]
- García-Orellana, Y.; Ortuño, M.F.; Conejero, W.; Ruiz-Sánchez, M.C. Diurnal variations in water relations of deficit irrigated lemon trees during fruit growth period. Spanish J. Agric. Res. 2013, 11, 137–145. [Google Scholar] [CrossRef] [Green Version]
- Manja, K.; Aoun, M. The use of nets for tree fruit crops and their impact on the production: A review. Sci. Hortic. 2019, 246, 110–122. [Google Scholar] [CrossRef]
- Mupambi, G.; Anthony, B.M.; Layne, D.R.; Musacchi, S.; Serra, S.; Schmidt, T.; Kalcsits, L.A. The influence of protective netting on tree physiology and fruit quality of apple: A review. Sci. Hortic. 2018, 236, 60–72. [Google Scholar] [CrossRef]
- Mditshwa, A.; Magwaza, L.S.; Tesfay, S.Z. Shade netting on subtropical fruit: Effect on environmental conditions, tree physiology and fruit quality. Sci. Hortic. 2019, 256, 108556. [Google Scholar] [CrossRef]
- Alarcón, J.J.; Ortuño, M.F.; Nicolás, E.; Navarro, A.; Torrecillas, A. Improving water-use efficiency of young lemon trees by shading with aluminised-plastic nets. Agric. Water Manag. 2006, 82, 387–398. [Google Scholar] [CrossRef]
- Incesu, M.; Yeşiloǧlu, T.; Çimen, B.; Yilmaz, B. Effects of nursery shading on plant growth, chlorophyll content and PSII in ‘Lane Late’ navel orange seedlings. Acta Hortic. 2016, 1130, 301–305. [Google Scholar] [CrossRef]
- Zhou, K.; Jerszurki, D.; Sadka, A.; Shlizerman, L.; Rachmilevitch, S.; Ephrath, J. Effects of photoselective netting on root growth and development of young grafted orange trees under semi-arid climate. Sci. Hortic. 2018, 238, 272–280. [Google Scholar] [CrossRef]
- Medina, C.L.; Souza, R.P.; Machado, E.C.; Ribeiro, R.V.; Silva, J.A.B. Photosynthetic response of citrus grown under reflective aluminized polypropylene shading nets. Sci. Hortic. 2002, 96, 115–125. [Google Scholar] [CrossRef]
- Jifon, J.L.; Syvertsen, J.P. Effects of moderate shade on citrus leaf gas exchange, fruit yield, and quality. Proc. Fla. State Hortic. Soc. 2001, 114, 117–181. [Google Scholar]
- Blakey, R.J.; van Rooyen, Z.; Köhne, J.S.; Malapana, K.C.; Mazhawu, E.; Tesfay, S.Z.; Savage, M.J. Growing Avocados Under Shade netting. Progress Report-Year 2. S. Afr. Avocado Grow. Assos. Yearb. 2016, 38, 80–83. [Google Scholar]
- Tinyane, P.P.; Soundy, P.; Sivakumar, D. Growing ‘Hass’ avocado fruit under different coloured shade netting improves the marketable yield and affects fruit ripening. Sci. Hortic. 2018, 230, 43–49. [Google Scholar] [CrossRef]
- Alaphilippe, A.; Capowiez, Y.; Simon, S.; Saudreau, M.; Caruso, S.; Vergani, S. Codling moth exclusion netting: An overview of French and Italian experiences. IOBC WPRS Bull. 2016, 112, 31–35. [Google Scholar]
- Kührt, U.; Samietz, J.; Dorn, S. Effect of plant architecture and hail nets on temperature of codling moth habitats in apple orchards. Entomol. Exp. Appl. 2006, 118, 245–259. [Google Scholar] [CrossRef]
- Gimeno, V.; Simón, I.; Martínez, V.; Lidón, V.; Shahid, M.A.; Garcia-Sanchez, F. Effect of shade screen on production, fruit quality and growth parameters of ‘Fino 49′ lemon trees grafted on Citrus macrophylla and sour orange. Acta Hortic. 2015, 1065, 1845–1852. [Google Scholar] [CrossRef]
- Hunsche, M.; Blanke, M.M.; Noga, G. Does the microclimate under hail nets influence micromorphological characteristics of apple leaves and cuticles? J. Plant Physiol. 2010, 167, 974–980. [Google Scholar] [CrossRef]
- Solomakhin, A.; Blanke, M. The microclimate under coloured hailnets affects leaf and fruit temperature, leaf anatomy, vegetative and reproductive growth as well as fruit colouration in apple. Ann. Appl. Biol. 2010, 156, 121–136. [Google Scholar] [CrossRef]
- García-Sánchez, F.; Simón, I.; Lidón, V.; Manera, F.J.; Simón-Grao, S.; Pérez-Pérez, J.G.; Gimeno, V. Shade screen increases the vegetative growth but not the production in ‘Fino 49′ lemon trees grafted on Citrus macrophylla and Citrus aurantium L. Sci. Hortic. 2015, 194, 175–180. [Google Scholar] [CrossRef]
- Domingo, R.; Ruiz-Sánchez, M.C.; Sánchez-Blanco, M.J.; Torrecillas, A. Water relations, growth and yield of Fino lemon trees under regulated deficit irrigation. Irrig. Sci. 1996, 16, 115–123. [Google Scholar] [CrossRef]
- Goldhamer, D.A.; Intrigliolo, D.S.; Castel, J.R.; Fereres, E. Citrus. In Crop Yield Response to Water; Steduto, P., Hsiao, T.C., Fereres, E., Raes, D., Eds.; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2012; pp. 316–331. ISBN 978-92-5-107274-5. [Google Scholar]
- Sánchez-Blanco, M.J.; Torrecillas, A.; Del Amor, F.; Leon, A. The water relations of Verna lemon trees from flowering to the end of rapid fruit growth. Biol. Plant. 1990, 32, 357–363. [Google Scholar] [CrossRef]
- Ruiz-Sánchez, M.C.; Domingo, R.; Pérez-Pastor, A. Daily variations in water relations of apricot trees under different irrigation regimes. Biol. Plant. 2007, 51, 735–740. [Google Scholar] [CrossRef]
- Conejero, W.; Mellisho, C.D.; Ortuño, M.F.; Galindo, A.; Pérez-Sarmiento, F.; Torrecillas, A. Establishing maximum daily trunk shrinkage and midday stem water potential reference equations for irrigation scheduling of early maturing peach trees. Irrig. Sci. 2011, 29, 299–309. [Google Scholar] [CrossRef]
- Fereres, E.; Goldhamer, D. Suitability of stem diameter variations and water potential as indicators for irrigation scheduling of almond trees. J. Hortic. Sci. Biotechnol. 2003, 78, 139–144. [Google Scholar] [CrossRef]
- McCutchan, H.; Shackel, K. Stem-water potential as a sensitive indicator of water stress in prune trees (Prunus domestica L. cv. French). J. Am. Soc. Hortic. Sci. 1992, 117, 607–611. [Google Scholar] [CrossRef] [Green Version]
- Naor, A. Irrigation Scheduling and Evaluation of Tree Water Status in Deciduous Orchards. In Horticultural Reviews; Wiley-Blackwell, Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2010; Volume 32, pp. 111–165. ISBN 9780470767986. [Google Scholar]
- Abrisqueta, I.; Conejero, W.; Valdés-Vela, M.; Vera, J.; Ortuño, M.F.; Ruiz-Sánchez, M.C. Stem water potential estimation of drip-irrigated early-maturing peach trees under Mediterranean conditions. Comput. Electron. Agric. 2015, 114, 7–13. [Google Scholar] [CrossRef]
- Brakke, M.; Allen, L.H. Gas exchange of Citrus seedlings at different temperatures, vapor-pressure deficits, and soil water contents. J. Am. Soc. Hortic. Sci. 1995, 120, 497–504. [Google Scholar] [CrossRef] [Green Version]
- De Freitas, S.T.; do Amarante, C.V.T.; Dandekar, A.M.; Mitcham, E.J. Shading affects flesh calcium uptake and concentration, bitter pit incidence and other fruit traits in ‘Greensleeves’ apple. Sci. Hortic. 2013, 161, 266–272. [Google Scholar] [CrossRef]
- Germanà, C.; Continella, A.; Tribulato, E. Net shading influence on floral induction on citrus trees. Acta Hortic. 2003, 614, 527–533. [Google Scholar] [CrossRef]
- Ribeiro, R.V.; Machado, E.C.; Santos, M.G.; Oliveira, R.F. Photosynthesis and water relations of well-watered orange plants as affected by winter and summer conditions. Photosynthetica 2009, 47, 215–222. [Google Scholar] [CrossRef]
- Zekri, M.; Parsons, L.R. Water relations of grapefruit trees in response to drip, microsprinkler, and overhead sprinkler irrigation. J. Am. Soc. Hortic. Sci. 1988, 113, 819–823. [Google Scholar]
- Levy, Y.; Kaufmann, M.R. Cycling of leaf conductance in citrus exposed to natural and controlled environments. Can. J. Bot. 1979, 54, 2215–2218. [Google Scholar] [CrossRef]
- Ben Mimoun, M.; Longuenesse, J.J.; Genard, M. Pmax as related to leaf: Fruit ratio and fruit assimilate demand in peach. J. Hortic. Sci. 1996, 71, 767–775. [Google Scholar] [CrossRef]
- Layne, D.R.; Flore, J.A. Physiological responses of Prunus cerasus to whole-plant source manipulation. Leaf gas exchange, chlorophyll fluorescence, water relations and carbohydrate concentrations. Physiol. Plant. 1993, 88, 44–51. [Google Scholar] [CrossRef]
- Romero-Trigueros, C.; Bayona, J.M.; Nortes, P.A.; Alarcón, J.J.; Nicolás, E. Determination of crop water stress index by infrared thermometry in grapefruit trees irrigated with saline reclaimed water combined with deficit irrigation. Remote Sens. 2019, 11, 757. [Google Scholar] [CrossRef] [Green Version]
- Jackson, R.D.; Idso, S.B.; Reginato, R.J.; Pinter, P.J. Canopy temperature as a crop water stress indicator. Water Resour. Res. 1981, 17, 1133–1138. [Google Scholar] [CrossRef]
- Wang, D.; Gartung, J. Infrared canopy temperature of early-ripening peach trees under postharvest deficit irrigation. Agric. Water Manag. 2010, 97, 1787–1794. [Google Scholar] [CrossRef]
- Vera, J.; Conejero, W.; Conesa, M.R.; Ruiz-Sánchez, M.C. Irrigation factor approach based on soil water content: A nectarine orchard case study. Water 2019, 11, 589. [Google Scholar] [CrossRef] [Green Version]
- Hsiao, T.C. Measurements of plant water status. In Irrigation of Agricultural Crops (Monograph No. 30); Stewart, B.A., Nielsen, D.R., Eds.; American Society of Agronomy: Madison, WI, USA, 1990; pp. 243–279. [Google Scholar]








© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mira-García, A.B.; Conejero, W.; Vera, J.; Ruiz-Sánchez, M.C. Leaf Water Relations in Lime Trees Grown under Shade Netting and Open-Air. Plants 2020, 9, 510. https://doi.org/10.3390/plants9040510
Mira-García AB, Conejero W, Vera J, Ruiz-Sánchez MC. Leaf Water Relations in Lime Trees Grown under Shade Netting and Open-Air. Plants. 2020; 9(4):510. https://doi.org/10.3390/plants9040510
Chicago/Turabian StyleMira-García, Ana Belén, Wenceslao Conejero, Juan Vera, and María Carmen Ruiz-Sánchez. 2020. "Leaf Water Relations in Lime Trees Grown under Shade Netting and Open-Air" Plants 9, no. 4: 510. https://doi.org/10.3390/plants9040510
APA StyleMira-García, A. B., Conejero, W., Vera, J., & Ruiz-Sánchez, M. C. (2020). Leaf Water Relations in Lime Trees Grown under Shade Netting and Open-Air. Plants, 9(4), 510. https://doi.org/10.3390/plants9040510