Mars Regolith Simulant Ameliorated by Compost as in situ Cultivation Substrate Improves Lettuce Growth and Nutritional Aspects

 ,
,  , , ,
, , ,  ,
,  ,
,  ,
, 

Abstract
:1. Introduction
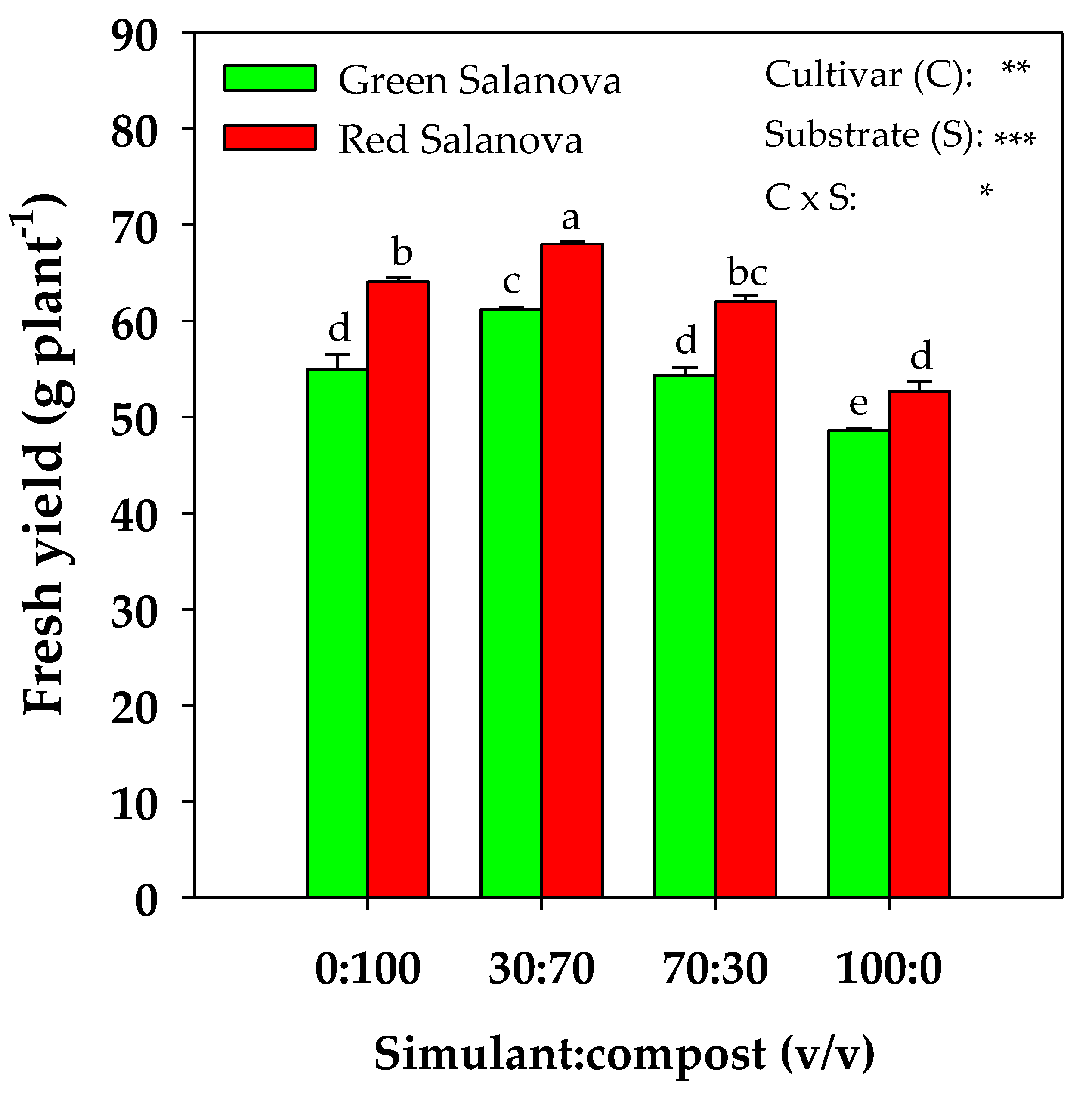
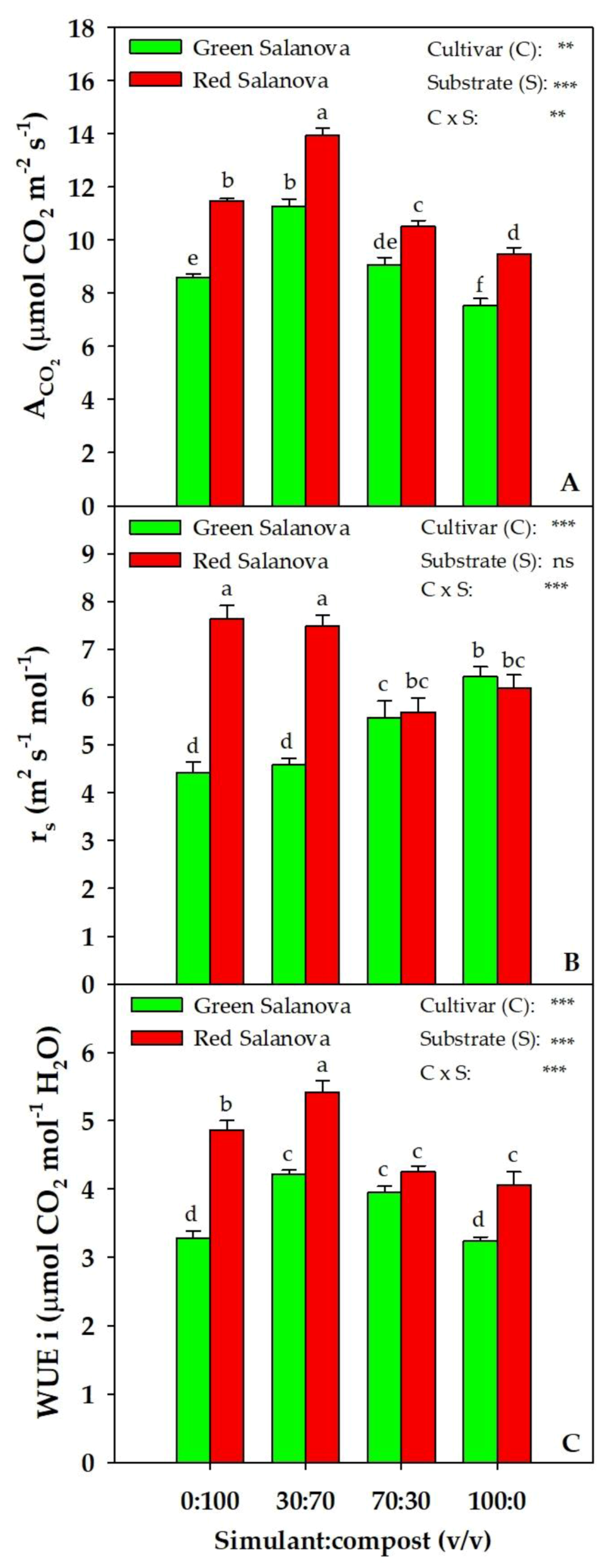
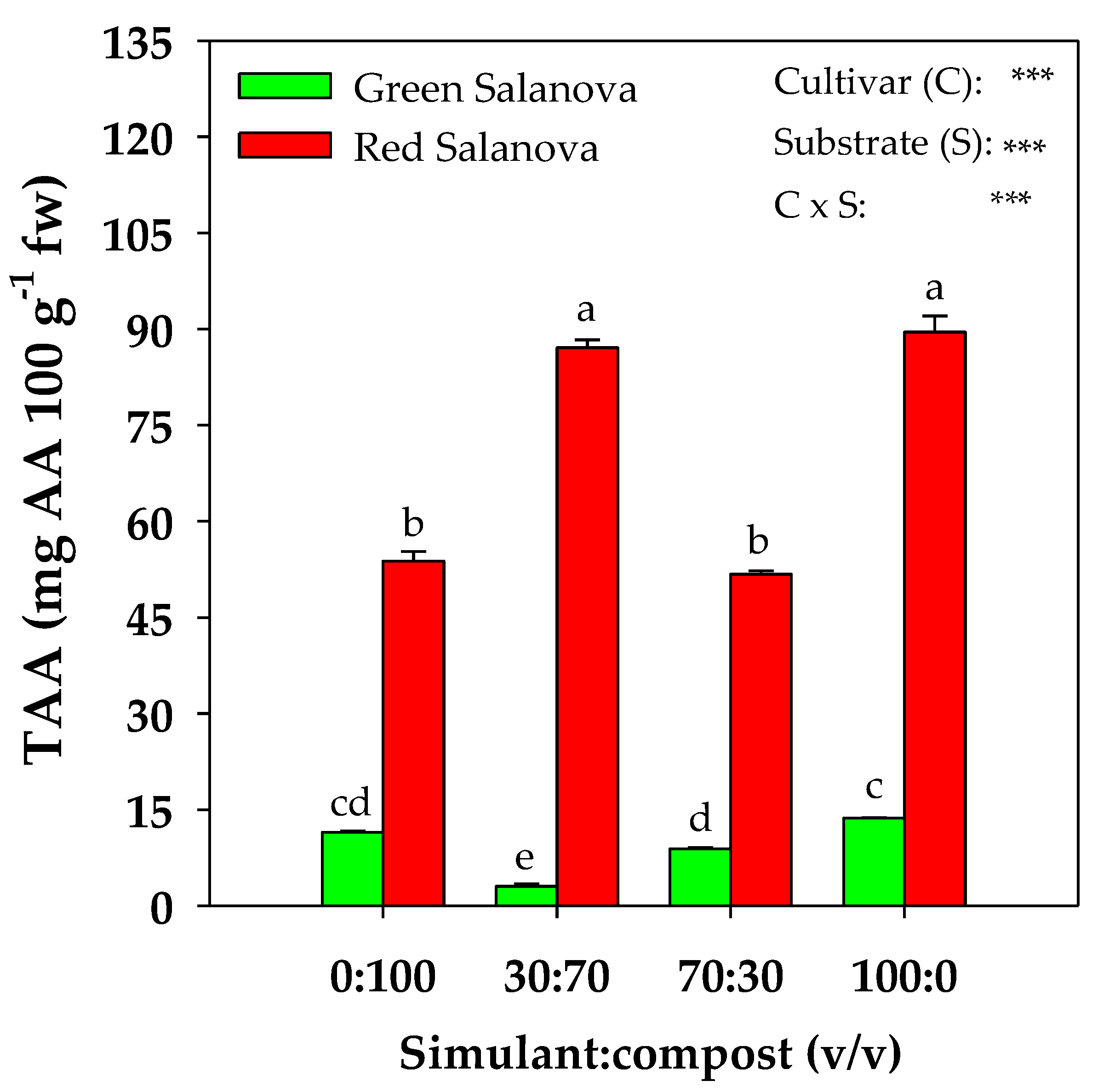
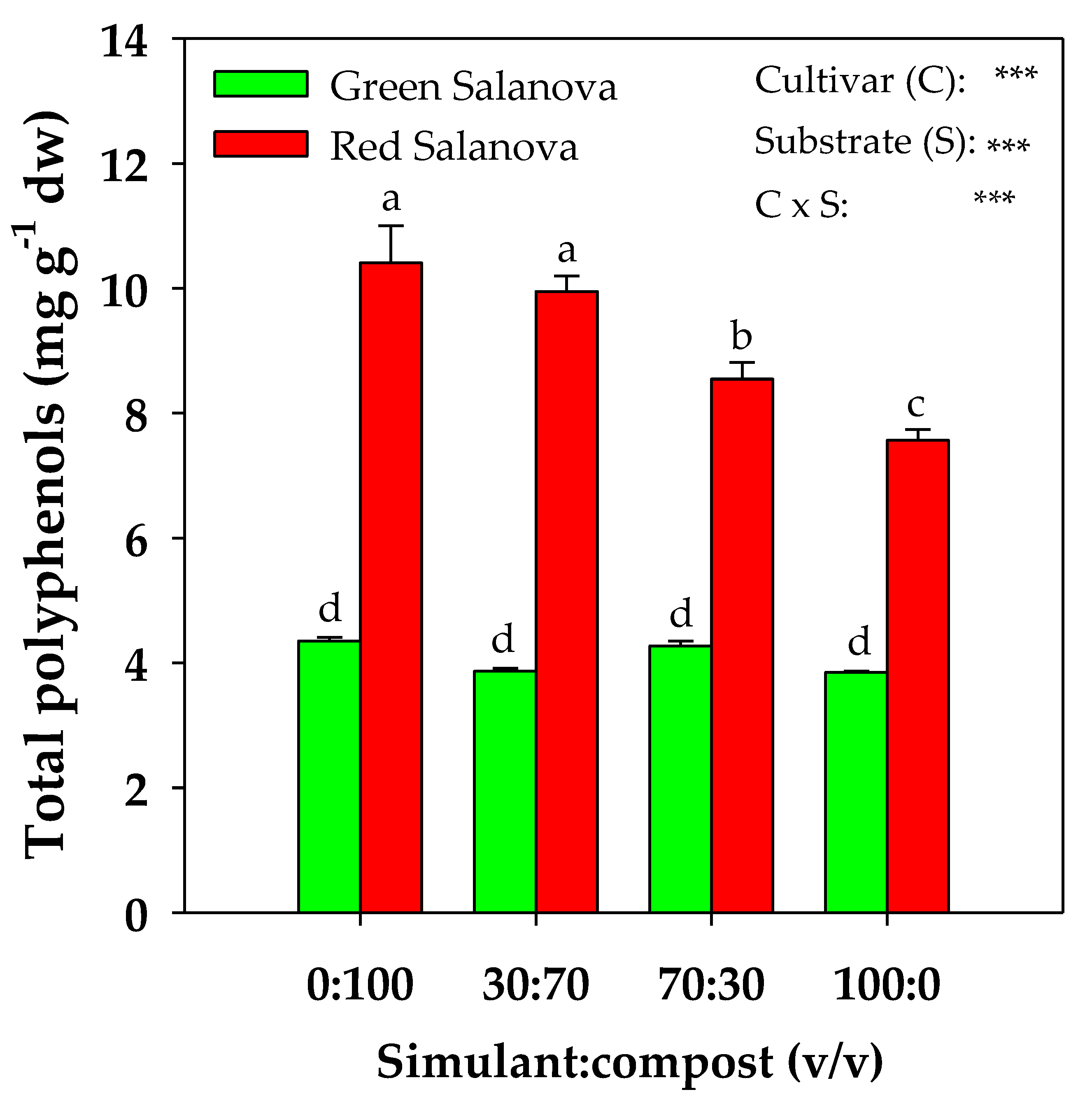
2. Results
2.1. Yield and Physiological Parameters
2.2. Shoots and Roots Mineral Composition
2.3. Total Ascorbic Acid, Total Chlorophyll and Carotenoids Content
2.4. Polyphenols Content Profile
3. Discussion
4. Materials and Methods
4.1. Plant Growth Conditions and Experimental Design
4.2. Leaf Gas Exchange
4.3. Fresh Biomass and Sampling
4.4. Total Nitrogen, Nitrate and Mineral Content
4.5. Total Chlorophyll and Total Ascorbic Acid Content
4.6. Carotenoids Quantification by HPLC-DAD and Polyphenols Analysis by UHPLC-Q-Orbitrap HRMS
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Menezes, A.A.; Cumbers, J.; Hogan, J.A.; Arkin, A.P. Towards synthetic biological approaches to resource utilization on space missions. J. R. Soc. Interface 2015, 12, 20140715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Llorente, B.; Williams, T.; Goold, H. The multiplanetary future of plant synthetic biology. Genes 2018, 9, 348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyen, F.E.; Hecht, M.H.; Hoffman, J.A.; MOXIE Team. Thermodynamic model of Mars oxygen ISRU experiment (MOXIE). Acta Astronaut. 2016, 129, 82–87. [Google Scholar] [CrossRef] [Green Version]
- Verseux, C.; Baque, M.; Lehto, K.; de Vera, J.P.P.; Rothschild, L.J.; Billi, D. Sustainable life support on Mars–the potential roles of cyanobacteria. Int. J. Astrobiol. 2016, 15, 65–92. [Google Scholar] [CrossRef] [Green Version]
- Benaroya, H.; Metzger, P.; Muscatello, A. Special issue: In situ resource utilization. J. Aerospace Eng. 2013, 26, 1–4. [Google Scholar] [CrossRef]
- Loader, C.A.; Garland, J.L.; Levine, L.H.; Cook, K.L.; Mackowiak, C.L.; Vivenzio, H.R. Direct recycling of human hygiene water into hydroponic plant growth systems. Life Support Biosph. Sci. 1999, 6, 141–152. [Google Scholar]
- Paradiso, R.; De Micco, V.; Buonomo, R.; Aronne, G.; Barbieri, G.; De Pascale, S. Soilless cultivation of soybean for Bioregenerative Life Support Systems: A literature review and the experience of the MEL i SSA P roject–F ood characterisation P hase I. Plant Biol. 2014, 16, 69–78. [Google Scholar] [CrossRef]
- Fu, Y.; Li, L.; Xie, B.; Dong, C.; Wang, M.; Jia, B.; Shao, L.; Dong, Y.; Deng, S.; Liu, H.; et al. How to establish a Bioregenerative Life Support System for long-term crewed missions to the Moon or Mars. Astrobiology 2016, 16, 925–936. [Google Scholar] [CrossRef]
- El-Nakhel, C.; Giordano, M.; Pannico, A.; Carillo, P.; Fusco, G.M.; Pascale, S.D.; Rouphael, Y. Cultivar-specific performance and qualitative descriptors for butterhead Salanova lettuce produced in closed soilless cultivation as a candidate salad crop for human Life Support in Space. Life 2019, 9, 61. [Google Scholar] [CrossRef] [Green Version]
- Bates, S.; Gushin, V.; Bingham, G.; Vinokhodova, A.; Marquit, J.; Sychev, V. Plants as countermeasures: A review of the literature and application to habitation systems for humans living in isolated or extreme environments. Habitation 2009, 12, 33–40. [Google Scholar] [CrossRef]
- Koga, K.; Iwasaki, Y. Psychological and physiological effect in humans of touching plant foliage-using the semantic differential method and cerebral activity as indicators. J. Physiol. Anthropol. 2013, 32, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Odeh, R.; Guy, C.L. Gardening for therapeutic people-plant interactions during long-duration space missions. Open Agric. 2017, 2, 1–13. [Google Scholar] [CrossRef]
- Chunxiao, X.; Hong, L. Crop candidates for the bioregenerative life support systems in China. Acta Astronaut. 2008, 63, 1076–1080. [Google Scholar] [CrossRef]
- Wheeler, R.M. Agriculture for space: People and places paving the way. Open Agric. 2017, 2, 14–32. [Google Scholar] [CrossRef]
- Kuang, A.; Xiao, Y.; McClure, G.; Musgrave, M.E. Influence of microgravity on ultrastructure and storage reserves in seeds of Brassica rapa L. Ann. Bot. 2000, 85, 851–859. [Google Scholar] [CrossRef] [Green Version]
- Meinen, E.; Dueck, T.; Kempkes, F.; Stanghellini, C. Growing fresh food on future space missions: Environmental conditions and crop management. Sci. Hortic. 2018, 235, 270–278. [Google Scholar] [CrossRef]
- Hoff, J.E.; Howe, J.M.; Mitchell, C.A. Nutritional and Cultural Aspects of Plant Species Selection for a Controlled Ecological Life Support System; NASA Technical Report; Purdue University: West Lafayette, IN, USA, 1982; Doc ID: 19820016109. [Google Scholar]
- Baslam, M.; Morales, F.; Garmendia, I.; Goicoechea, N. Nutritional quality of outer and inner leaves of green and red pigmented lettuces (Lactuca sativa L.) consumed as salads. Sci. Hortic. 2013, 151, 103–111. [Google Scholar] [CrossRef]
- Rouphael, Y.; Kyriacou, M.; Vitaglione, P.; Giordano, M.; Pannico, A.; Colantuono, A.; De Pascale, S. Genotypic variation in nutritional and antioxidant profile among iceberg lettuce cultivars. Acta Sci. Pol.-Hortoru. 2017, 16, 37–45. [Google Scholar] [CrossRef]
- Kim, D.E.; Shang, X.; Assefa, A.D.; Keum, Y.S.; Saini, R.K. Metabolite profiling of green, green/red, and red lettuce cultivars: Variation in health beneficial compounds and antioxidant potential. Food Res. Int. 2018, 361–370. [Google Scholar] [CrossRef]
- Giordano, M.; El-Nakhel, C.; Pannico, A.; Kyriacou, M.C.; Stazi, S.R.; De Pascale, S.; Rouphael, Y. Iron biofortification of red and green pigmented lettuce in closed soilless cultivation impacts crop performance and modulates mineral and bioactive composition. Agronomy 2019, 9, 290. [Google Scholar] [CrossRef] [Green Version]
- Della Penna, D. Nutritional genomics: Manipulating plant micronutrients to improve human health. Science 1999, 285, 375–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.J.; Moon, Y.; Tou, J.C.; Mou, B.; Waterland, N.L. Nutritional value, bioactive compounds and health benefits of lettuce (Lactuca sativa L.). J. Food Compos. Anal. 2016, 49, 19–34. [Google Scholar] [CrossRef]
- Kennedy, D.O.; Wightman, E.L. Herbal extracts and phytochemicals: Plant secondary metabolites and the enhancement of human brain function. Adv. Nutr. 2011, 2, 32–50. [Google Scholar] [CrossRef] [PubMed]
- Rouphael, Y.; Kyriacou, M.C. Enhancing quality of fresh vegetables through salinity eustress and biofortification applications facilitated by soilless cultivation. Front. Plant Sci. 2018, 9, 1254. [Google Scholar] [CrossRef] [PubMed]
- Rouphael, Y.; Kyriacou, M.C.; Petropoulos, S.A.; De Pascale, S.; Colla, G. Improving vegetable quality in controlled environments. Sci. Hortic. 2018, 234, 275–289. [Google Scholar] [CrossRef]
- El-Nakhel, C.; Pannico, A.; Kyriacou, M.C.; Giordano, M.; De Pascale, S.; Rouphael, Y. Macronutrient deprivation eustress elicits differential secondary metabolites in red and green-pigmented butterhead lettuce grown in a closed soilless system. J. Sci. Food Agric. 2019, 99, 6962–6972. [Google Scholar] [CrossRef]
- McCollom, T.M.; Robbins, M.; Moskowitz, B.; Berquó, T.S.; Jöns, N.; Hynek, B.M. Experimental study of acid sulfate alteration of basalt and implications for sulfate deposits on Mars. J. Geophys. Res. Planets 2013, 118, 577–614. [Google Scholar] [CrossRef]
- Zeng, X.; Li, X.; Wang, S.; Li, S.; Spring, N.; Tang, H.; Li, Y.; Feng, J. JMSS-1: A new Martian soil simulant. Earth Planets Space 2015, 67. [Google Scholar] [CrossRef] [Green Version]
- Filiberto, J. Geochemistry of Martian basalts with constraints on magma genesis. Chem. Geol. 2017, 466, 1–14. [Google Scholar] [CrossRef]
- Cannon, K.M.; Britt, D.T.; Smith, T.M.; Fritsche, R.F.; Batcheldor, D. Mars global simulant MGS-1: A rocknest-based open standard for basaltic martian regolith simulants. Icarus 2019, 317, 470–478. [Google Scholar] [CrossRef] [Green Version]
- Benison, K.C.; LaClair, D.; Walker, J. Physical sedimentology experiments with sulfuric acid solutions: Implications for Mars? Earth Planet. Sci. Lett. 2008, 270, 330–337. [Google Scholar] [CrossRef]
- Eigenbrode, J.L.; Summons, R.E.; Steele, A.; Freissinet, C.; Millan, M.; Navarro-González, R.; Sutter, B.; McAdam, A.C.; Franz, H.B.; Glavin, D.P.; et al. Organic matter preserved in 3-billion-year-old mudstones at Gale crater, Mars. Science 2018, 360, 1096–1101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilrain, M.R.; Hogan, J.A.; Cowan, R.M.; Finstein, M.S.; Logendra, L.S. Preliminary study of greenhouse grown Swiss chard in mixtures of compost and Mars regolith simulant. SAE Tech. Pap. 1999. [Google Scholar] [CrossRef]
- Mortley, D.G.; Aglan, H.A.; Bonsi, C.K.; Hill, W.A. Growth of sweetpotato in lunar and mars simulants. SAE Tech. Pap. 2000. [Google Scholar] [CrossRef]
- Wamelink, G.W.; Frissel, J.Y.; Krijnen, W.H.; Verwoert, M.R.; Goedhart, P.W. Can plants grow on Mars and the moon: A growth experiment on Mars and moon soil simulants. PLoS ONE 2014, 9. [Google Scholar] [CrossRef]
- Caporale, A.G.; Vingiani, S.; Palladino, M.; El-Nakhel, C.; Duri, L.G.; Pannico, A.; Rouphael, Y.; De Pascale, S.; Adamo, P. Geo-mineralogical characterisation of Mars simulant MMS-1 and appraisal of substrate physico-chemical properties and crop performance obtained with variable green compost amendment rates. Sci. Total Environ. 2020, 720, 137543. [Google Scholar] [CrossRef]
- Rouphael, Y.; Petropoulos, S.A.; El Nakhel, C.; Pannico, A.; Kyriacou, M.C.; Giordano, M.; Troise, A.D.; Vitaglione, P.; De Pascale, S. Reducing energy requirements in future Bioregenerative Life Support Systems (BLSSs): Performance and bioactive composition of diverse lettuce genotypes grown under optimal and suboptimal light conditions. Front. Plant Sci. 2019, 10, 1305. [Google Scholar] [CrossRef]
- Di Massa, G.; Emmerich, J.C.; Morrow, R.C.; Bourget, C.M.; Mitchell, C.A. Plant-growth lighting for space life support: A review. Gravit. Space Biol. 2006, 19, 19–30. [Google Scholar]
- Caporale, A.G.; Adamo, P.; Azam, S.M.G.G.; Rao, M.A.; Pigna, M. May humic acids or mineral fertilisation mitigate arsenic mobility and availability to carrot plants (Daucus carota L.) in a volcanic soil polluted by as from irrigation water? Chemosphere 2018, 193, 464–471. [Google Scholar] [CrossRef]
- Ouni, Y.; Mateos-Naranjo, E.; Lakhdar, A.; Andrades-Moreno, L.; Abdelly, C.; Barhoumi, Z. Municipal solid waste compost application improves the negative impact of saline soil in two forage species. Commun. Soil Sci. Plan. 2014, 45, 1421–1434. [Google Scholar] [CrossRef]
- Schuerger, A.C.; Fajardo-Cavazos, P.; Clausen, C.A.; Moores, J.E.; Smith, P.H.; Nicholson, W.L. Slow degradation of ATP in simulated martian environments suggests long residence times for the biosignature molecule on spacecraft surfaces on Mars. Icarus 2008, 194, 86–100. [Google Scholar] [CrossRef]
- Badescu, V. Mars: Prospective Energy and Material Resources; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2009. [Google Scholar] [CrossRef] [Green Version]
- Maggi, F.; Pallud, C. Martian base agriculture: The effect of low gravity on water flow, nutrient cycles, and microbial biomass dynamics. Adv. Space Res. 2010, 46, 1257–1265. [Google Scholar] [CrossRef]
- Porterfield, D.M. The biophysical limitations in physiological transport and exchange in plants grown in microgravity. J. Plant Growth Regul. 2002, 21, 177–190. [Google Scholar] [CrossRef] [PubMed]
- El-Nakhel, C.; Pannico, A.; Kyriacou, M.C.; Petropoulos, S.A.; Giordano, M.; Colla, G.; Troise, A.D.; Vitaglione, P.; De Pascale, S.; Rouphael, Y. Dataset on the organic acids, sulphate, total nitrogen and total chlorophyll contents of two lettuce cultivars grown hydroponically using nutrient solutions of variable macrocation ratios. Data Brief 2020, 29, 105135. [Google Scholar] [CrossRef]
- Abd El-Salam, M.S.; Abd El Lateef, E.M.; Tawfik, M.M.; Farrag, A.A. Effect of soil amendments on wheat (Triticum aestivum L.) yield and nutritional status in sandy calcareous saline soil. Int. J. ChemTech Res. 2016, 9, 143–153. [Google Scholar]
- Agegnehu, G.; Nelson, P.N.; Bird, M.I. Crop yield, plant nutrient uptake and soil physicochemical properties under organic soil amendments and nitrogen fertilization on Nitisols. Soil Till. Res. 2016, 160, 1–13. [Google Scholar] [CrossRef]
- Paulauskiene, A.; Danilcenko, H.; Pranckietiene, I.; Taraseviciene, Z. Effect of different fertilizers on the mineral content of pumpkin fruit. J. Elementol. 2018, 23, 1033–1042. [Google Scholar] [CrossRef]
- Kabata-Pendias, A. Trace Elements in Soils and Plants, 4th ed.; CRC Press—Taylor & Francis Group: Boca Raton, FL, USA, 2011. [Google Scholar] [CrossRef]
- El-Nakhel, C.; Petropoulos, S.A.; Pannico, A.; Kyriacou, M.C.; Giordano, M.; Colla, G.; Troise, A.D.; Vitaglione, P.; De Pascale, S.; Rouphael, Y. The bioactive profile of lettuce produced in a closed soilless system as configured by combinatorial effects of genotype and macrocation supply composition. Food Chem. 2020, 309, 125713. [Google Scholar] [CrossRef]
- Thatikunta, R.; Lakshmi, S.; Prasadini, P. Effect of organic manures on SCMR, protein content and quality in Maize. Ecol. Environ. Conserv. 2012, 18, 51–52. [Google Scholar]
- Lefsrud, M.; Kopsell, D.; Sams, C.; Wills, J.; Both, A.J. Dry matter content and stability of carotenoids in kale and spinach during drying. HortScience 2008, 43, 1731–1736. [Google Scholar] [CrossRef] [Green Version]
- Kołton, A.; Wojciechowska, R.; Długosz-Grochowska, O.; Grzesiak, W. The storage ability of lamb’s lettuce cultivated in the greenhouse under LED or HPS lamps. J. Hortic. Res. 2014, 22, 159–165. [Google Scholar] [CrossRef] [Green Version]
- Ouzounis, T.; Razi Parjikolaei, B.; Fretté, X.; Rosenqvist, E.; Ottosen, C.O. Predawn and high intensity application of supplemental blue light decreases the quantum yield of PSII and enhances the amount of phenolic acids, flavonoids, and pigments in Lactuca sativa. Front. Plant Sci. 2015, 6, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fageria, N.K. The Use of Nutrients in Crop Plants; CRC Press—Taylor & Francis Group: Boca Raton, FL, USA, 2009. [Google Scholar] [CrossRef]
- Fanasca, S.; Colla, G.; Maiani, G.; Venneria, E.; Rouphael, Y.; Azzini, E.; Saccardo, F. Changes in antioxidant content of tomato fruits in response to cultivar and nutrient solution composion. J. Agric. Food Chem. 2006, 54, 4319–4325. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.; Jiang, Y.; Zhao, H.; Guo, D.; He, L.; Liu, F.; Zhou, Q.; Nandwani, D.; Hui, D.; Yu, J. Electrical conductivity of nutrient solution influenced photosynthesis, quality, and antioxidant enzyme activity of pakchoi (Brassica campestris L. ssp. Chinensis) in a hydroponic system. PLoS ONE 2018, 13, e0202090. [Google Scholar] [CrossRef] [PubMed]
- Taghipour, S.; Rahimi, A.; Zartoshti, M.R.; Arslan, Y. The effect of micronutrients on antioxidant properties of thyme (Thymus vulgaris L.) under humic acid using condition. YYU J. Agric. Sci. 2017, 27, 589–600. [Google Scholar] [CrossRef]
- Sousa, C.; Valentao, P.; Range, J.; Lopes, G.; Pereira, J.A.; Ferreres, F.; Seabra, R.M.; Andrade, P.B. Influence of two fertilization regimens on the amounts of organic acids and phenolic compounds of tronchuda cabbage (Brassica oleracea L. Var. costata DC). J. Agric. Food Chem. 2005, 53, 9128–9132. [Google Scholar] [CrossRef]
- Saikia, L.R.; Upadhyaya, S. Antioxidant activity, phenol and flavonoid content of A.racemosus Willd. a medicinal plant grown using different organic manures. Res. J. Pharm. Biol. Chem. Sci. 2011, 2, 457–463. [Google Scholar]
- Aminifard, M.; Aroiee, H.; Azizi, M.; Nemati, H.; Jaafar, H. Effect of compost on antioxidant components and fruit quality of sweet pepper (Capsicum annuum L.). J. Cent. Eur. Agric. 2013, 14, 47–56. [Google Scholar] [CrossRef] [Green Version]
- Luján-Hidalgo, M.C.; Gómez-Hernández, D.E.; Villalobos-Maldonado, J.J.; Abud-Archila, M.; Montes-Molina, J.A.; Enciso-Saenz, S.; Ruiz-Valdiviezo, V.M.; Gutiérrez-Miceli, F.A. Effects of vermicompost and vermiwash on plant, phenolic content, and anti-oxidant activity of Mexican pepperleaf (Piper auritum Kunth) cultivated in phosphate rock potting media. Compost Sci. Util. 2017, 25, 95–101. [Google Scholar] [CrossRef]
- Schiavon, M.; Pizzeghello, D.; Muscolo, A.; Vaccaro, S.; Francioso, O.; Nardi, S. High molecular size humic substances enhance phenylpropanoid metabolism in maize (Zea mays L.). J. Chem. Ecol. 2010, 36, 662–669. [Google Scholar] [CrossRef]
- Canellas, L.P.; Olivares, F.L.; Aguiar, N.O.; Jones, D.L.; Nebbioso, A.; Mazzei, P.; Piccolo, A. Humic and fulvic acids as biostimulants in horticulture. Sci. Hortic. 2015, 196, 15–27. [Google Scholar] [CrossRef]
- Neocleous, D.; Koukounaras, A.; Siomos, A.S.; Vasilakakis, M. Assessing the salinity effects on mineral composition and nutritional quality of green and red ‘Baby’ lettuce. J. Food Qual. 2014, 37, 1–8. [Google Scholar] [CrossRef]
- Rapisarda, P.; Tomaino, A.; Lo Cascio, R.; Bonina, F.; De Pasqual, A.; Saija, A. Antioxidant effectiveness as influenced by phenolic content of fresh orange juices. J. Agric. Food Chem. 1999, 47, 4718–4723. [Google Scholar] [CrossRef] [PubMed]
- Rouphael, Y.; Cardarelli, M.; Lucini, L.; Rea, E.; Colla, G. Nutrient solution concentration affects growth, mineral composition, phenolic acids, and flavonoids in leaves of artichoke and cardoon. HortScience 2012, 47, 1424–1429. [Google Scholar] [CrossRef] [Green Version]
- Carillo, P.; Colla, G.; El-Nakhel, C.; Bonini, P.; D’Amelia, L.; Dell’Aversana, E.; Pannico, A.; Giordano, M.; Sifola, M.I.; Kyriacou, M.C.; et al. Biostimulant application with a tropical plant extract enhances Corchorus olitorius adaptation to sub-optimal nutrient regimens by improving physiological parameters. Agronomy 2019, 9, 249. [Google Scholar] [CrossRef] [Green Version]
- Bremner, J.M. Nitrogen—Total. In Methods of Soil Analysis, Agronomy Monograph; Black, C.A., Evans, D.D., White, I.L., Ensminger, L.E., Clark, F.E., Eds.; American Society of Agronomy: Madison, WI, USA, 1965; Part 2; pp. 1149–1178. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G.; Giordano, M.; El-Nakhel, C.; Kyriacou, M.C.; De Pascale, S. Foliar applications of a legume-derived protein hydrolysate elicit dose-dependent increases of growth, leaf mineral composition, yield and fruit quality in two greenhouse tomato cultivars. Sci. Hortic. 2017, 226, 353–360. [Google Scholar] [CrossRef]
- Lichtenhaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. In Proceedings of the Biochemical Society Transactions 603rd Meeting, Liverpool, UK, 1 October 1983; Volume 11, pp. 591–592. [Google Scholar] [CrossRef] [Green Version]
- Kampfenkel, K.; Vanmontagu, M.; Inze, D. Extraction and determination of ascorbate and dehydroascorbate from plant tissue. Anal. Biochem. 1995, 225, 165–167. [Google Scholar] [CrossRef]
- Kyriacou, M.C.; El-Nakhel, C.; Graziani, G.; Pannico, A.; Soteriou, G.A.; Giordano, M.; Ritieni, A.; De Pascale, S.; Rouphael, Y. Functional quality in novel food sources: Genotypic variation in the nutritive and phytochemical composition of thirteen microgreens species. Food Chem. 2019, 277, 107–118. [Google Scholar] [CrossRef]
- Kim, H.J.; Fonseca, J.M.; Choi, J.H.; Kubota, C.; Kwon, D.Y. Salt in irrigation water affects the nutritional and visual properties of romaine lettuce (Lactuca sativa L.). J. Agric. Food Chem. 2008, 56, 3772–3776. [Google Scholar] [CrossRef]
- Kyriacou, M.C.; El Nakhel, C.; Pannico, A.; Graziani, G.; Soteriou, G.; Giordano, M.; Zarrelli, A.; Ritieni, A.; De Pascale, S.; Rouphael, Y. Genotype-specific modulatory effects of select spectral bandwidths on the nutritive and phytochemical composition of microgreens. Front. Plant Sci. 2019, 10, 1501. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variance | NO3 (g kg−1 dw) | PO4 (g kg−1 dw) | K (g kg−1 dw) | Ca (g kg−1 dw) | Mg (g kg−1 dw) | Na (g kg−1 dw) | Cl (g kg−1 dw) | SO4 (g kg−1 dw) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Shoot | Root | Shoot | Root | Shoot | Root | Shoot | Root | Shoot | Root | Shoot | Root | Shoot | Root | Shoot | Root | |
| Cultivar (C) | ||||||||||||||||
| Green Salanova | 27.6 | 42.9 a | 9.0 | 5.7 | 64.7 | 50.6 | 7.1 | 6.2 | 2.5 | 2.8 | 1.0 | 5.9 | 3.3 | 2.2 | 1.5 b | 8.9 |
| Red Salanova | 30.4 | 28.8 b | 10.4 | 7.5 | 71.7 | 44.6 | 6.2 | 6.0 | 2.5 | 2.9 | 1.0 | 5.2 | 3.1 | 1.9 | 2.5 a | 9.4 |
| Simulant:compost (v:v) (S) | ||||||||||||||||
| 0:100 | 29.4 a | 33.3 ab | 11.2 a | 8.7 a | 82.7 a | 69.2 a | 4.8 c | 5.6 ab | 2.2 b | 2.2 c | 0.8 b | 2.0 c | 6.9 a | 2.8 a | 2.2 a | 9.6 ab |
| 30:70 | 32.5 a | 24.9 b | 11.9 a | 7.3 a | 75.2 b | 48.4 b | 6.4 b | 6.7 a | 2.4 b | 2.5 bc | 0.8 b | 2.7 bc | 2.0 b | 1.7 b | 2.3 a | 9.3 b |
| 70:30 | 32.4 a | 43.6 a | 9.5 b | 7.3 a | 69.5 c | 57.4 b | 6.9 b | 6.6 a | 2.3 b | 3.0 b | 0.9 b | 4.8 b | 2.0 b | 2.0 b | 2.0 a | 11.1 a |
| 100:0 | 21.7 b | 41.6 a | 6.2 c | 2.9 b | 45.2 d | 15.5 c | 8.5 a | 5.3 b | 3.2 a | 3.9 a | 1.7 a | 12.8 a | 1.7 b | 1.9 b | 1.4 b | 6.6 c |
| C × S | ||||||||||||||||
| Green Salanova × 0:100 | 28.0 | 37.3 | 10.6 | 6.9 | 80.2 | 77.6 | 5.5 | 5.6 | 2.3 | 2.2 c | 0.9 | 1.9 | 7.5 | 3.1 | 1.7 | 10.1 |
| Green Salanova × 30:70 | 32.6 | 30.7 | 11.2 | 6.1 | 71.3 | 48.0 | 6.4 | 6.6 | 2.3 | 2.2 c | 0.8 | 2.6 | 1.9 | 1.5 | 1.7 | 8.8 |
| Green Salanova × 70:30 | 30.4 | 52.1 | 8.3 | 6.6 | 64.6 | 59.0 | 7.3 | 6.6 | 2.3 | 2.6 bc | 1.0 | 5.0 | 2.2 | 2.2 | 1.6 | 10.5 |
| Green Salanova × 100:0 | 19.5 | 51.5 | 5.8 | 3.2 | 42.6 | 17.8 | 9.3 | 5.9 | 3.2 | 4.3 a | 1.5 | 14.0 | 1.7 | 2.2 | 1.1 | 6.2 |
| Red Salanova × 0:100 | 30.9 | 29.2 | 11.8 | 10.6 | 85.3 | 60.8 | 4.2 | 5.6 | 2.1 | 2.3 c | 0.8 | 2.1 | 6.4 | 2.4 | 2.8 | 9.1 |
| Red Salanova × 30:70 | 32.4 | 19.1 | 12.8 | 8.6 | 79.2 | 48.9 | 6.3 | 6.9 | 2.6 | 2.8 bc | 0.7 | 2.8 | 2.2 | 1.8 | 2.9 | 9.7 |
| Red Salanova × 70:30 | 34.4 | 35.0 | 10.8 | 8.0 | 74.5 | 55.9 | 6.5 | 6.7 | 2.3 | 3.3 b | 0.7 | 4.5 | 1.9 | 1.8 | 2.4 | 11.8 |
| Red Salanova × 100:0 | 23.8 | 31.7 | 6.5 | 2.7 | 47.8 | 13.1 | 7.7 | 4.7 | 3.3 | 3.4 b | 1.9 | 11.6 | 1.8 | 1.7 | 1.7 | 7.1 |
| Significance | ||||||||||||||||
| Cultivar (C) | ns | ** | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | *** | ns |
| Substrate (S) | * | * | *** | *** | *** | *** | *** | * | *** | *** | ** | *** | *** | ** | *** | *** |
| C × S | ns | ns | ns | ns | ns | ns | ns | ns | ns | * | ns | ns | ns | ns | ns | ns |
| Source of Variance | Total N | Nitrate | Total Chlorophyll | Lutein | β-Carotene |
|---|---|---|---|---|---|
| (g 100g−1 dw) | (mg kg−1 fw) | (mg 100g−1 fw) | (mg kg−1 dw) | (mg kg−1 dw) | |
| Cultivar (C) | |||||
| Green Salanova | 3.9 | 1488 | 10.3 b | 85.5 b | 262.4 b |
| Red Salanova | 4.0 | 1528 | 21.8 a | 249.5 a | 511.2 a |
| Simulant:compost (v:v) (S) | |||||
| 0:100 | 3.9 | 1542 | 15.4 | 170.3 b | 386.9 |
| 30:70 | 4.0 | 1609 | 14.6 | 164.0 b | 379.3 |
| 70:30 | 3.9 | 1637 | 16.7 | 199.4 a | 437.3 |
| 100:0 | 3.8 | 1244 | 17.6 | 136.2 c | 343.6 |
| C × S | |||||
| Green Salanova × 0:100 | 3.9 | 1486 | 10.6 | 88.7 | 271.0 |
| Green Salanova × 30:70 | 4.0 | 1670 | 10.2 | 88.8 | 262.7 |
| Green Salanova × 70:30 | 3.9 | 1591 | 9.9 | 112.4 | 295.2 |
| Green Salanova × 100:0 | 3.7 | 1205 | 10.5 | 52.1 | 220.7 |
| Red Salanova × 0:100 | 4.0 | 1598 | 20.1 | 251.9 | 502.8 |
| Red Salanova × 30:70 | 4.0 | 1548 | 19.0 | 239.3 | 495.8 |
| Red Salanova × 70:30 | 4.0 | 1682 | 23.5 | 286.3 | 579.3 |
| Red Salanova × 100:0 | 3.9 | 1283 | 24.7 | 220.3 | 466.6 |
| Significance | |||||
| Cultivar (C) | ns | ns | *** | *** | *** |
| Substrate (S) | ns | ns | ns | ** | ns |
| C × S | ns | ns | ns | ns | ns |
| Source of Variance | Chlorogenic Acid | Caffeic Acid Hexoside | Caffeic Acid | Luteolin-7-Oglucoside | Apigenin Malonil Glucoside | Coumaroyl Quinic Acid | Coumaric Acid | Feruloyl Quinic Acid | Quercetin-3-O-Galactoside | Dicaffeoylquinic Acid | Quercetin-3-O-Glucuronide | Quercetin-3-O-Glucoside | Feruloylglycoside | Kaempferol-7-O-Glucoside | Rutin | Quercetin Malonylglucoside | Kaempferolo-3-O-Rutinoside | Feruloyltartaric Acid | Caffeoylferuloylquinic Acid |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (µg g−1 dw) | |||||||||||||||||||
| Cultivar (C) | |||||||||||||||||||
| Green Salanova | 330 b | 9.7 | 15.1 b | 4.1 b | 64.8 a | 420.6 b | 9.5 a | 17.8 b | 7.7 b | nd | 69.3 a | 7.4 b | 10.0 a | 4.1 b | 814.0 b | 614 b | 51.8 b | 1064 a | 571 b |
| Red Salanova | 4156 a | 6.9 | 57.9 a | 8.4 a | 24.0 b | 746.7 a | 6.8 b | 25.8 a | 40.1 a | 90.0 | 52.8 b | 34.6 a | 7.3 b | 9.2 a | 866.3 a | 1276 a | 73.3 a | 978 b | 656 a |
| Simulant:compost (v/v) (S) | |||||||||||||||||||
| 0:100 | 2437 a | 12.6 a | 34.2 b | 5.7 c | 100.8 a | 534.1 c | 8.0 b | 21.5 b | 23.2 b | 134.9 | 76.3 a | 25.3 a | 8.2 c | 5.4 c | 943.0 a | 1335 a | 67.4 a | 1039 a | 634 a |
| 30:70 | 2534 a | 6.8 c | 46.4 a | 6.6 b | 26.0 b | 620.8 b | 7.7 b | 23.7 a | 34.4 a | 73.5 | 73.6 a | 21.3 c | 9.5 b | 6.8 b | 846.2 b | 914 b | 60.3 b | 1015 b | 618 b |
| 70:30 | 2345 a | 7.4 b | 48.3 a | 7.5 a | 32.4 b | 502.9 c | 9.2 a | 25.1 a | 22.7 b | 84.0 | 60.2 b | 22.8 b | 10.8 a | 9.1 a | 808.1 c | 774 b | 62.0 b | 1014 b | 606 c |
| 100:0 | 1658 b | 6.4 c | 17.1 c | 5.1 d | 18.5 c | 676.6 a | 7.8 b | 16.8 c | 15.2 c | 67.5 | 34.1 c | 14.6 d | 6.1 d | 5.4 c | 763.3 d | 757 b | 60.5 b | 1016 b | 596 d |
| C × S | |||||||||||||||||||
| Green Salanova × 0:100 | 138 d | 17.8 a | 6.1 f | 4.5 e | 175.5 a | 372.3 e | 9.5 b | 15.3 c | 6.2 f | nd | 101.3 a | 5.8 f | 8.7 c | 4.2 d | 883.8 b | 865 | 61.2 b | 1099 a | 577 e |
| Green Salanova × 30:70 | 241 d | 6.7 b | 15.6 de | 3.9 fg | 30.4 bc | 351.5 e | 8.9 c | 19.6 b | 7.0 f | nd | 74.9 b | 6.5 ef | 11.0 b | 3.9 d | 782.1 d | 631 | 47.2 c | 1051 b | 573 ef |
| Green Salanova × 70:30 | 639 d | 7.5 b | 26.0 c | 4.3 ef | 31.9 b | 396.0 e | 10.7 a | 21.3 b | 10.3 e | nd | 68.3 b | 10.2 d | 12.7 a | 4.4 d | 825.9 c | 535 | 48.8 c | 1054 b | 569 f |
| Green Salanova × 100:0 | 302 d | 7 b | 12.7 ef | 3.6 g | 21.6 cd | 562.4 d | 8.9 c | 15.1 c | 7.2 f | nd | 32.5 d | 7.1 e | 7.5 d | 3.9 d | 764.4 d | 424 | 49.8 c | 1054 b | 566 f |
| Red Salanova × 0:100 | 4735 a | 7.4 b | 62.4 b | 7.0 c | 26.1 bc | 696.0 c | 6.5 e | 27.8 a | 40.3 b | 134.9 a | 51.2 c | 44.8 a | 7.7 d | 6.7 c | 1002.1 a | 1805 | 73.6 a | 980 c | 692 a |
| Red Salanova × 30:70 | 4826 a | 7 b | 77.1 a | 9.2 b | 21.7 cd | 890.2 a | 6.6 e | 27.9 a | 61.9 a | 73.5 c | 72.2 b | 36.1 b | 8.0 d | 9.6 b | 910.3 b | 1196 | 73.4 a | 979 c | 664 b |
| Red Salanova × 70:30 | 4050 b | 7.3 b | 70.7 a | 10.7 a | 32.9 b | 609.8 d | 7.6 d | 29.0 a | 35.1 c | 84.0 b | 52.1 c | 35.5 b | 8.8 c | 13.7 a | 790.4 d | 1013 | 75.1 a | 975 c | 644 c |
| Red Salanova × 100:0 | 3014 c | 5.8 c | 21.5 cd | 6.5 d | 15.4 e | 790.8 b | 6.6 e | 18.6 b | 23.2 d | 67.5 c | 35.7 d | 22.0 c | 4.7 e | 6.9 c | 762.3 d | 1090 | 71.1 a | 978 c | 625 d |
| Significance | |||||||||||||||||||
| Cultivar (C) | *** | ns | *** | *** | * | *** | *** | *** | *** | na | ns | *** | ** | *** | ns | *** | *** | *** | *** |
| Substrate (S) | ** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** |
| C × S | *** | *** | *** | *** | *** | *** | * | ** | *** | *** | *** | *** | *** | *** | *** | ns | *** | ** | *** |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duri, L.G.; El-Nakhel, C.; Caporale, A.G.; Ciriello, M.; Graziani, G.; Pannico, A.; Palladino, M.; Ritieni, A.; De Pascale, S.; Vingiani, S.; et al. Mars Regolith Simulant Ameliorated by Compost as in situ Cultivation Substrate Improves Lettuce Growth and Nutritional Aspects. Plants 2020, 9, 628. https://doi.org/10.3390/plants9050628
Duri LG, El-Nakhel C, Caporale AG, Ciriello M, Graziani G, Pannico A, Palladino M, Ritieni A, De Pascale S, Vingiani S, et al. Mars Regolith Simulant Ameliorated by Compost as in situ Cultivation Substrate Improves Lettuce Growth and Nutritional Aspects. Plants. 2020; 9(5):628. https://doi.org/10.3390/plants9050628
Chicago/Turabian StyleDuri, Luigi G., Christophe El-Nakhel, Antonio G. Caporale, Michele Ciriello, Giulia Graziani, Antonio Pannico, Mario Palladino, Alberto Ritieni, Stefania De Pascale, Simona Vingiani, and et al. 2020. "Mars Regolith Simulant Ameliorated by Compost as in situ Cultivation Substrate Improves Lettuce Growth and Nutritional Aspects" Plants 9, no. 5: 628. https://doi.org/10.3390/plants9050628
APA StyleDuri, L. G., El-Nakhel, C., Caporale, A. G., Ciriello, M., Graziani, G., Pannico, A., Palladino, M., Ritieni, A., De Pascale, S., Vingiani, S., Adamo, P., & Rouphael, Y. (2020). Mars Regolith Simulant Ameliorated by Compost as in situ Cultivation Substrate Improves Lettuce Growth and Nutritional Aspects. Plants, 9(5), 628. https://doi.org/10.3390/plants9050628