Variation of Growth-to-Ripening Time Interval Induced by Abscisic Acid and Synthetic Auxin affecting Transcriptome and Flavor Compounds in Cabernet Sauvignon Grape Berry


 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
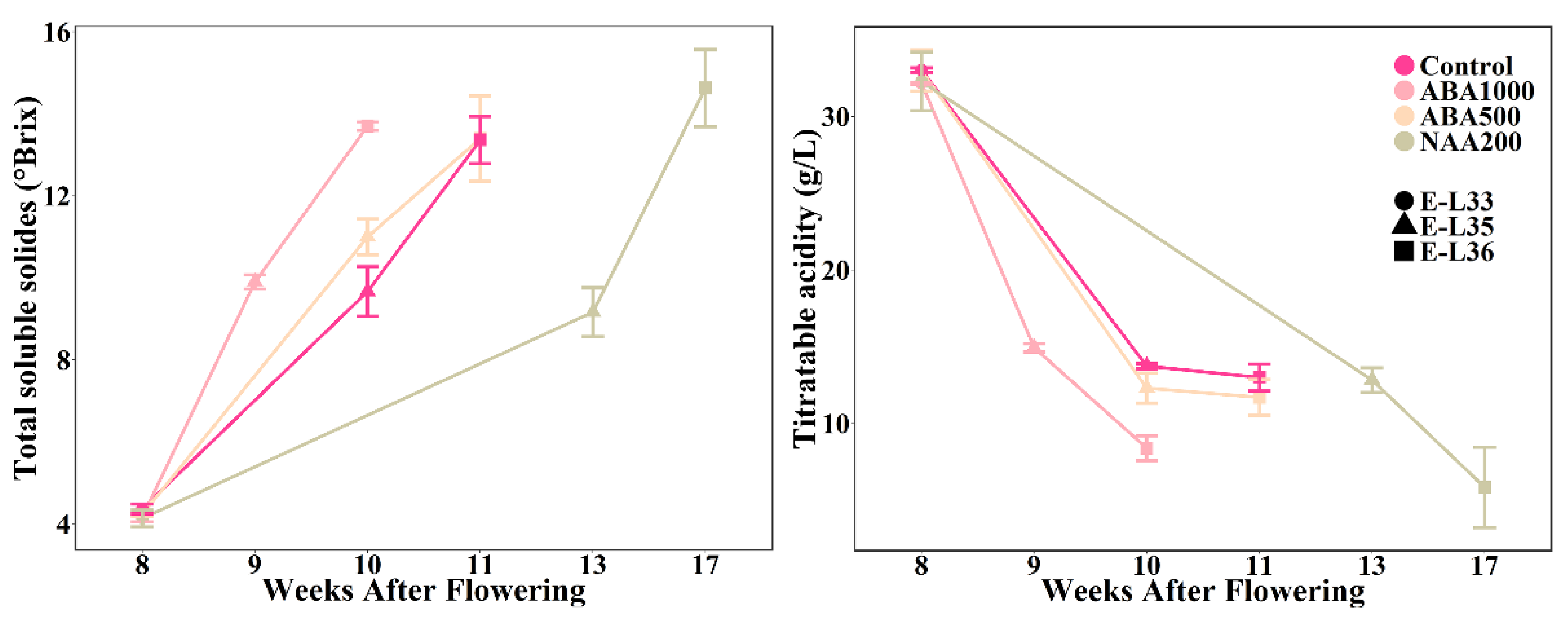
2.1. Variation in Timing and Duration of Ripening by ABA and NAA Treatments
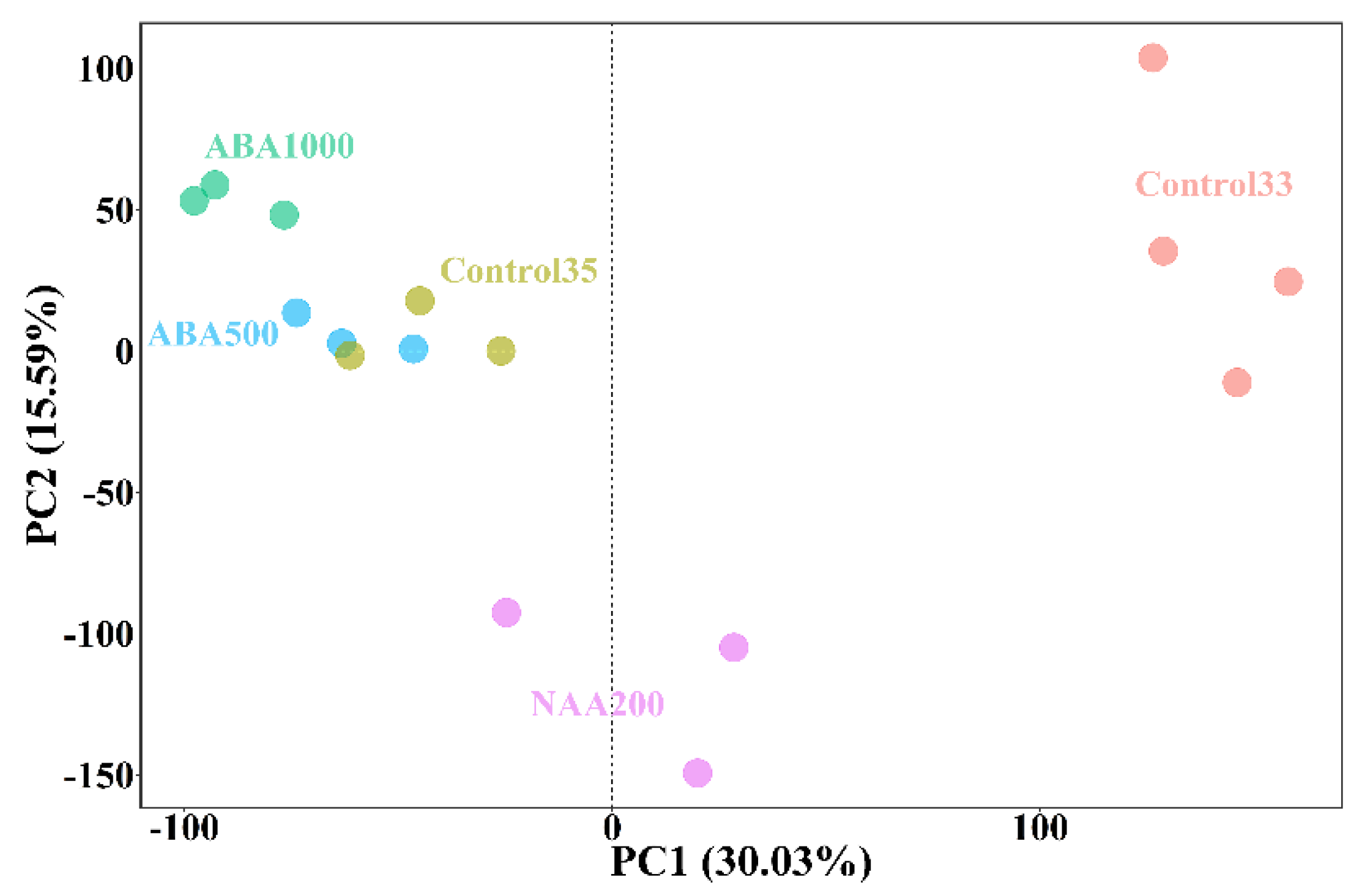
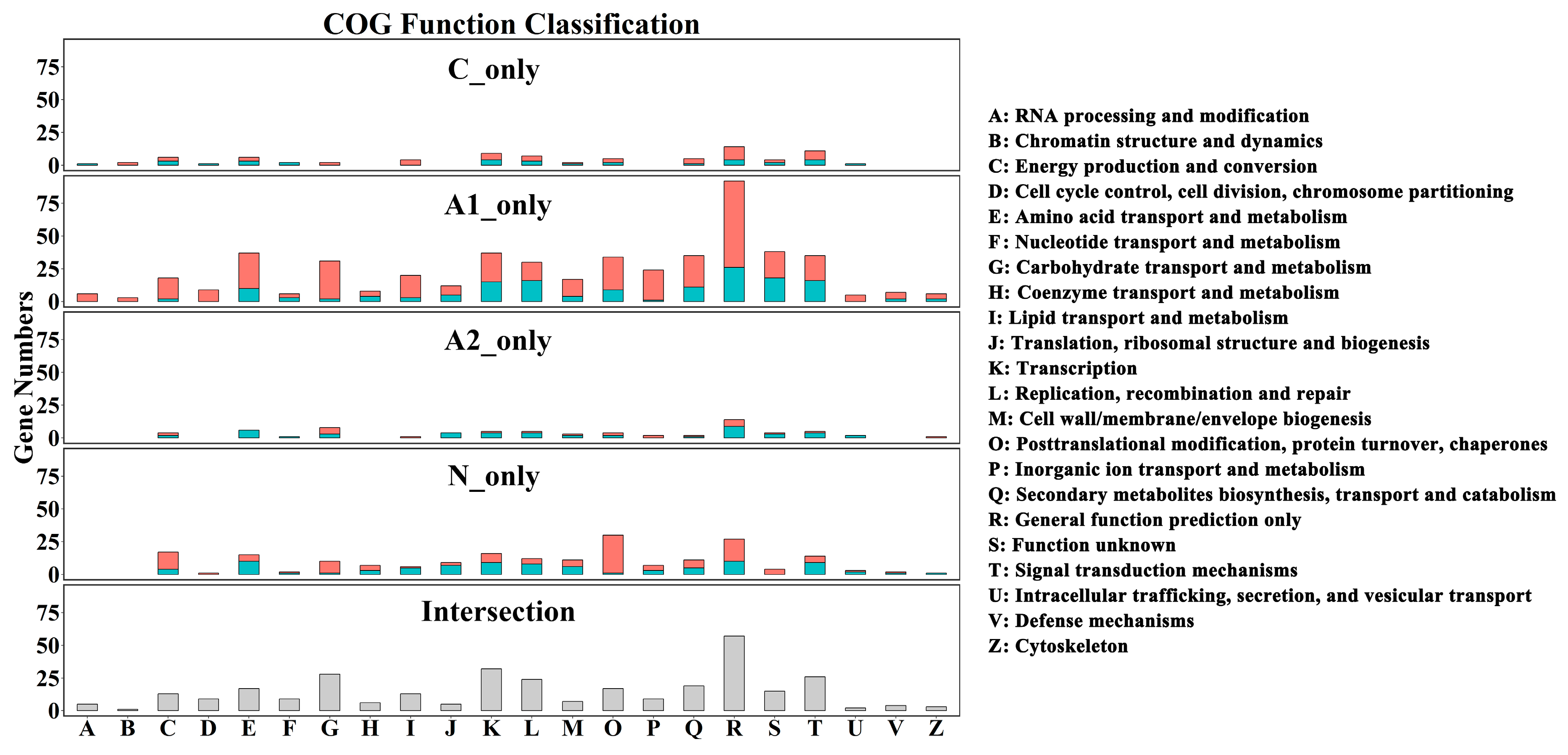
2.2. Transcriptomic Analysis of Gene Expression Profiling
2.3. Genes Associated with Growth-To-Ripening Transition and Ripening Initiation
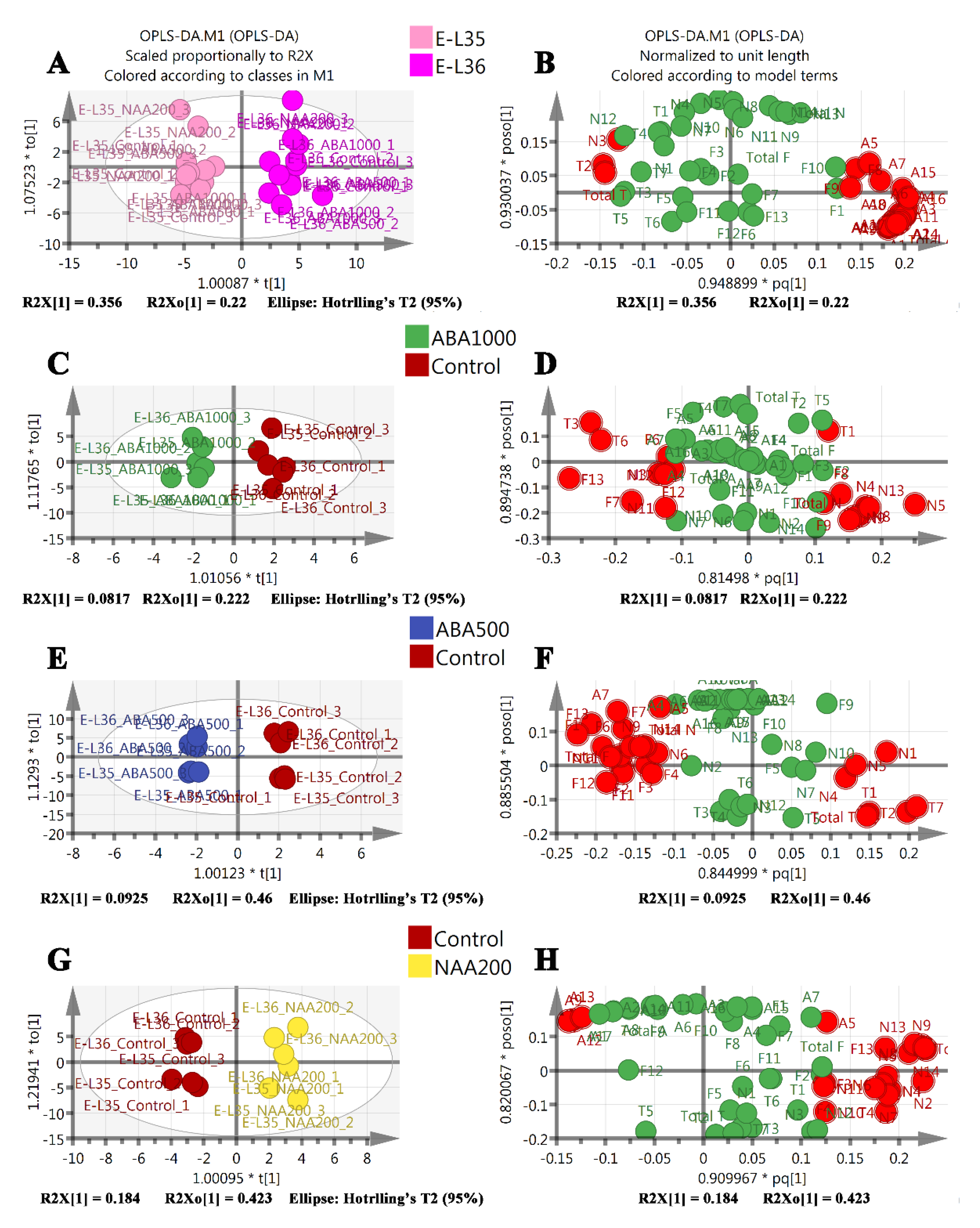
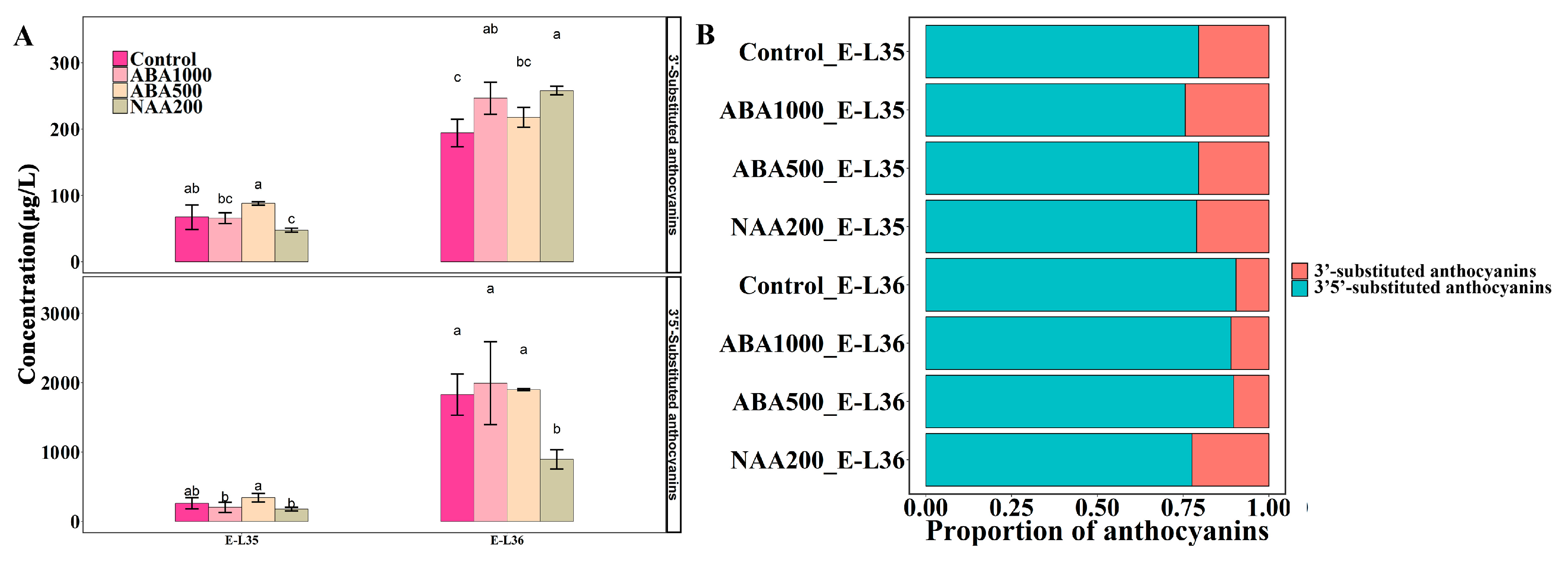
2.4. Identification of Development-Specific and Treatment-Specific Metabolites
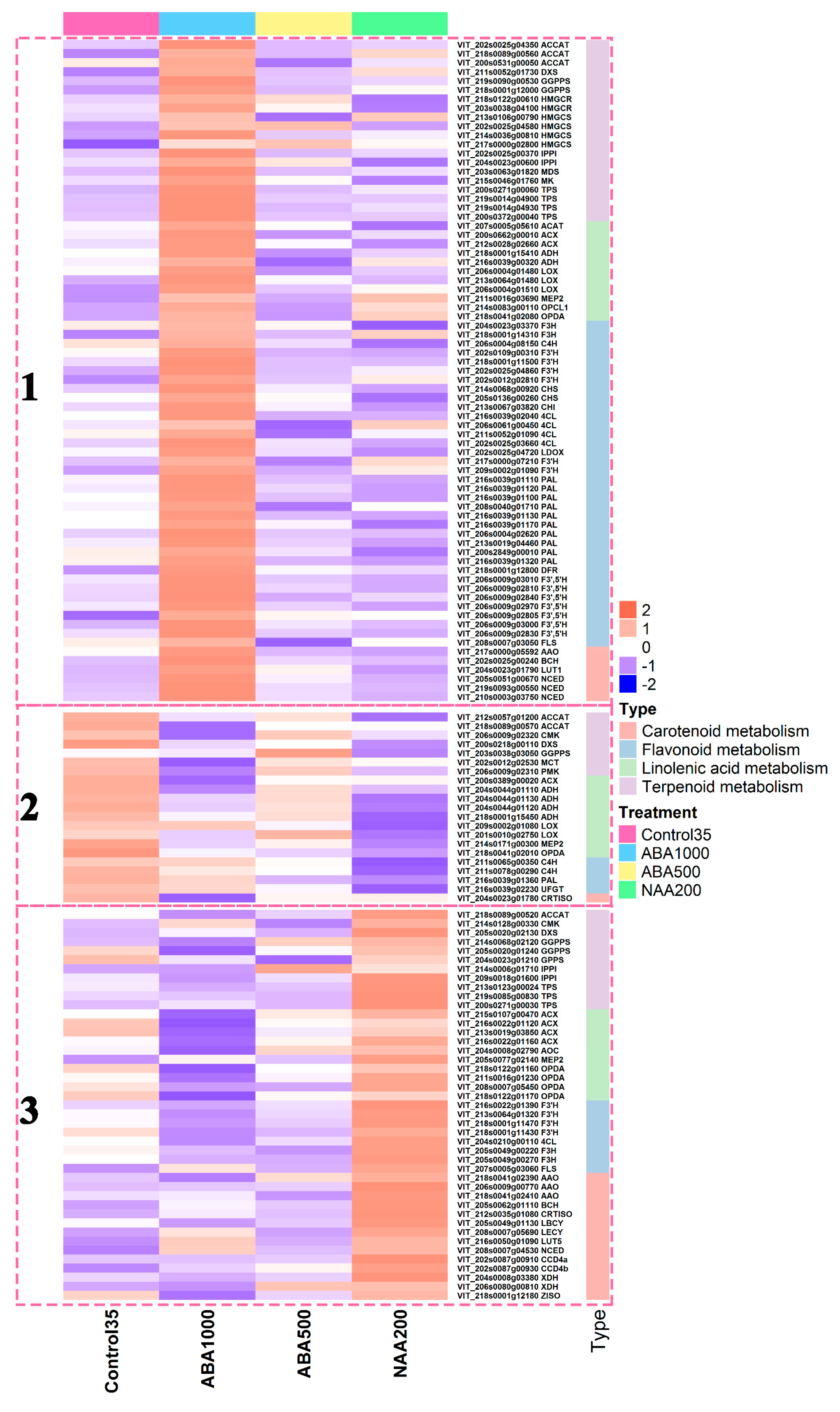
2.5. Berry Transcriptome Analysis Supports the Metabolic Plasticity
2.6. Validation by Quantitative Real-Time PCR
3. Discussion
4. Materials and Methods
4.1. ABA and NAA Application in Vineyard and Grape Berry Sampling
4.2. Physicochemical Index Analysis
4.3. RNA Extraction and Sequencing
4.4. Quantitative Real-Time PCR
4.5. Determination of Anthocyanins
4.6. Volatile Compounds Determination
4.7. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ABA | Abscisic acid |
| NAA | α-Naphthaleneacetic acid |
| SA | Salicylic acid |
| CL | Changli |
| GT | Gaotai |
| IAA | Indole-3-acetic acid |
| BTOA | Benzothiazole-2-oxyacetic acid |
| DEG | Differentially expressed genes |
| PE | Pectinesterase |
| PG | Polygalacturonase |
| PL | Pectate lyase |
| NCED | 9-Cis-epoxycarotenoid dioxygenase |
| SUC | Sucrose transporter |
| SUS | Sucrose synthase |
| MDH | Malate dehydrogenase |
| PYR/PYL | Pyrabactin resistance/pyrabactin-like |
| PP2C | Type 2C protein phosphates |
| TGA | TGACG motif-binding factor |
| BRI1 | Brassinosteroid-insensitive1 |
| BR | Brassinosteroid |
| BAK1 | BRI1 associated receptor Kinase 1 |
| AUX1 | Auxin influx carrier protein |
| AUX/IAA | Auxin-responsive proteins |
| TCH4 | Xyloglucan:xyloglucosyl transferase |
| F3′H | Flavonoid-3’-hydroxylases |
| UFGT | UDP glucose flavonoid-3-O-glucosyltransferase |
| PSY | Phytoene synthase |
| CCD | Carotenoid cleavage dioxygenase |
| RPKM | Reads per kilobases per million reads |
| FDR | False discovery rate |
| KEGG | Kyoto encyclopedia of genes and genomes |
| GO | Gene ontology |
| Nr | NCBI non-redundant protein sequences |
| qRT-PCR | Quantitative real-time PCR |
References
- Fang, Y.; Qian, M.C. Quantification of selected aroma-active compounds in pinot noir wines from different grape maturities. J. Agric. Food Chem. 2006, 54, 8567–8573. [Google Scholar] [CrossRef]
- Pérez-Magariño, S.; González-San José, M.L. Polyphenols and colour variability of red wines made from grapes harvested at different ripeness grade. Food Chem. 2006, 96, 197–208. [Google Scholar] [CrossRef]
- Miranda-Lopez, R.I.T.A.; Libbey, L.M.; Watson, B.T.; McDaniel, M.R. Odor analysis of Pinot Noir wines from grapes of different maturities by a gas chromatography-olfactometry technique (Osme). J. Food Sci. 1992, 57, 985–993. [Google Scholar] [CrossRef]
- Gomez, E.; Martinez, A.; Laencina, J. Changes in volatile compounds during maturation of some grape varieties. J. Sci. Food. Agric. 1995, 67, 229–233. [Google Scholar] [CrossRef]
- Zhang, B.; He, F.; Liu, Y.; Cai, J.; Duan, C.Q. Impact of Adding Ellagic Acid to Red Grapes on the Phenolic Composition and Chromatic Quality of Cabernet Sauvignon Wines from a Warm Climate. J. Food Process Preserv. 2017, 41, e13080. [Google Scholar] [CrossRef]
- Spayd, S.E.; Tarara, J.M.; Mee, D.L.; Ferguson, J.C. Separation of Sunlight and Temperature Effects on the Composition of Vitis vinifera cv. Merlot Berries. Am. J. Enol. Viticult. 2002, 53, 171–182. [Google Scholar]
- Feng, H.; Yuan, F.; Skinkis, P.A.; Qian, M.C. Influence of cluster zone leaf removal on Pinot noir grape chemical and volatile composition. Food Chem. 2015, 173, 414–423. [Google Scholar] [CrossRef]
- Kuhn, N.; Guan, L.; Dai, Z.W.; Wu, B.-H.; Lauvergeat, V.; Gomès, E.; Li, S.-H.; Godoy, F.; Arce-Johnson, P.; Delrot, S. Berry ripening: Recently heard through the grapevine. J. Exp. Bot. 2014, 65, 4543–4559. [Google Scholar] [CrossRef] [Green Version]
- Coombe, B.G.; Hale, C.R. The hormone content of ripening grape berries and the effects of growth substance treatments. Plant Physiol. 1973, 51, 629–634. [Google Scholar] [CrossRef] [Green Version]
- Jeong, S.T.; Goto-Yamamoto, N.; Kobayashi, S.; Esaka, M.J.P.S. Effects of plant hormones and shading on the accumulation of anthocyanins and the expression of anthocyanin biosynthetic genes in grape berry skins. Plant Sci. 2004, 167, 247–252. [Google Scholar] [CrossRef]
- Wheeler, S.; Loveys, B.; Ford, C.; Davies, C. The relationship between the expression of abscisic acid biosynthesis genes, accumulation of abscisic acid and the promotion of Vitis vinifera L. Berry ripening by abscisic acid. Aust. J. Grape Wine R. 2009, 15, 195–204. [Google Scholar] [CrossRef]
- Zhang, X.; Luo, G.; Wang, R.; Wang, J.; Himelrick, D.G. Growth and developmental responses of seeded and seedless grape berries to shoot girdling. J. Am. Soc. Hortic. Sci. 2003, 128, 316–323. [Google Scholar] [CrossRef] [Green Version]
- Böttcher, C.; Harvey, K.; Forde, C.G.; Boss, P.K.; Davies, C. Auxin treatment of pre-veraison grape (Vitis vinifera L.) berries both delays ripening and increases the synchronicity of sugar accumulation. Aust. J. Grape Wine R. 2011, 17, 1–8. [Google Scholar] [CrossRef]
- Davies, C.; Boss, P.K.; Robinson, S.P. Treatment of grape berries, a nonclimacteric fruit with a synthetic auxin, retards ripening and alters the expression of developmentally regulated genes. Plant Physiol. 1997, 115, 1155–1161. [Google Scholar] [CrossRef] [Green Version]
- Koyama, K.; Sadamatsu, K.; Goto-Yamamoto, N. Abscisic acid stimulated ripening and gene expression in berry skins of the cabernet sauvignon grape. Funct. Integr. Genomic. 2010, 10, 367–381. [Google Scholar] [CrossRef] [Green Version]
- Ziliotto, F.; Corso, M.; Rizzini, F.M.; Rasori, A.; Botton, A.; Bonghi, C. Grape berry ripening delay induced by a pre-véraison naa treatment is paralleled by a shift in the expression pattern of auxin- and ethylene-related genes. BMC Plant Biol. 2012, 12, 185. [Google Scholar] [CrossRef] [Green Version]
- Ferrara, G.; Mazzeo, A.; Matarrese, A.M.S.; Pacucci, C.; Punzi, R.; Faccia, M.; Trani, A.; Gambacorta, G. Application of abscisic acid (s-aba) and sucrose to improve colour, anthocyanin content and antioxidant activity of cv. Crimson seedless grape berries. Aust. J. Grape Wine R. 2015, 21, 18–29. [Google Scholar] [CrossRef]
- Böttcher, C.; Boss, P.K.; Davies, C. Delaying Riesling grape berry ripening with a synthetic auxin affects malic acid metabolism and sugar accumulation, and alters wine sensory characters. Funct. Plant Biol. 2012, 39, 745–753. [Google Scholar] [CrossRef]
- Strauss, C.R.; Wilson, B.; Gooley, P.R.; Williams, P.J. Role of monoterpenes in grape and wine flavor. In Biogeneration of Aromas; Parliament, T.H., Croteau, R., Eds.; American Chemical Society: Washington, DC, USA, 1986; pp. 222–242. [Google Scholar]
- Crupi, P.; Coletta, A.; Antonacci, D. Analysis of carotenoids in grapes to predict norisoprenoid varietal aroma of wines from Apulia. J. Agric. Food Chem. 2010, 58, 9647–9656. [Google Scholar] [CrossRef]
- Loscos, N.; Hernandez-Orte, P.; Cacho, J.; Ferreira, V. Release and formation of varietal aroma compounds during alcoholic fermentation from nonfloral grape odorless flavor precursors fractions. J. Agric. Food Chem. 2007, 55, 6674–6684. [Google Scholar] [CrossRef]
- Boss, P.K.; Davies, C. Molecular biology of sugar and anthocyanin accumulation in grape berries. In Molecular Biology and Biotechnology of the Grapevine; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2001; pp. 1–33. [Google Scholar]
- Coombe, B.G. Research on development and ripening of the grape berry. Am. J. Enol. Viticult. 1992, 43, 101–110. [Google Scholar]
- Fortes, A.M.; Teixeira, R.T.; Agudelo-Romero, P. Complex interplay of hormonal signals during grape berry ripening. Molecules. 2015, 20, 9326–9343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnold, W.N. β-fructofuranosidase from grape berries. BBA Enzymol. Biol. Oxid. 1965, 110, 134–147. [Google Scholar] [CrossRef]
- Wold, S.; Sjöström, M.; Eriksson, L. PLS-regression: A basic tool of chemometrics. Chemometr. Intell. Lab. 2001, 58, 109–130. [Google Scholar] [CrossRef]
- Kotseridis, Y.; Baumes, R.L.; Bertrand, A.; Skouroumounis, G.K. Quantitative determination of β-ionone in red wines and grapes of bordeaux using a stable isotope dilution assay. J. Chromatogr. A. 1999, 848, 317–325. [Google Scholar] [CrossRef]
- Pineau, B.; Barbe, J.C.; Van Leeuwen, C.; Dubourdieu, D. Which impact for β-damascenone on red wines aroma? J. Agric. Food Chem. 2007, 55, 4103–4108. [Google Scholar] [CrossRef]
- Lashbrooke, J.G.; Young, P.R.; Dockrall, S.J.; Vasanth, K.; Vivier, M.A. Functional characterisation of three members of the Vitis vinifera L. Carotenoid cleavage dioxygenase gene family. BMC Plant Biol. 2013, 13, 156. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Tian, L.; Liu, H.; Pan, Q.H.; Zhan, J.; Huang, W. Sugars induce anthocyanin accumulation and flavanone 3-hydroxylase expression in grape berries. Plant Growth Regul. 2009, 58, 251–260. [Google Scholar] [CrossRef]
- Villalobos-González, L.; Peña-Neira, A.; Ibáñez, F.; Pastenes, C. Long-term effects of abscisic acid (ABA) on the grape berry phenylpropanoid pathway: Gene expression and metabolite content. Plant Physiol. Bioch. 2016, 105, 213–223. [Google Scholar] [CrossRef]
- Castellarin, S.D.; Di Gaspero, G.; Marconi, R.; Nonis, A.; Peterlunger, E.; Paillard, S.; Adam-Blondon, A.F.; Testolin, R. Colour variation in red grapevines (Vitis vinifera L.): Genomic organisation, expression of flavonoid 3′-hydroxylase, flavonoid 3′, 5′-hydroxylase genes and related metabolite profiling of red cyanidin-/blue delphinidin-based anthocyanins in berry skin. BMC Genomics 2006, 7, 12. [Google Scholar] [CrossRef]
- Cunningham, F.X., Jr.; Gantt, E. Genes and enzymes of carotenoid biosynthesis in plants. Annu. Rev. Plant Phys. 1998, 49, 557–583. [Google Scholar] [CrossRef]
- Hannah, L.; Roehrdanz, P.R.; Ikegami, M.; Shepard, A.V.; Shaw, M.R.; Tabor, G.; Zhi, L.; Marquet, P.A.; Hijmans, R.J. Climate change, wine, and conservation. Proc. Natl. Acad. Sci. USA 2013, 110, 6907–6912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coombe, B.G. Growth stages of the grapevine: Adoption of a system for identifying grapevine growth stages. Aust. J. Grape Wine R. 1995, 1, 104–110. [Google Scholar] [CrossRef]
- Gao, Y.; Li, X.X.; Han, M.M.; Yang, X.F.; Li, Z.; Wang, J.; Pan, Q.H. Rain-shelter cultivation modifies carbon allocation in the polyphenolic and volatile metabolism of Vitis vinifera L. Chardonnay grapes. PLoS ONE 2016, 11, e0156117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reid, K.E.; Olsson, N.; Schlosser, J.; Peng, F.; Lund, S.T. An optimized grapevine RNA isolation procedure and statistical determination of reference genes for real-time RT-PCR during berry development. BMC Plant Biol. 2006, 6, 27. [Google Scholar] [CrossRef] [Green Version]
- Downey, M.O.; Harvey, J.S.; Robinson, S.P. Synthesis of flavonols and expression of flavonol synthase genes in the developing grape berries of shiraz and chardonnay (Vitis vinifera L.). Aust. J. Grape Wine R. 2003, 9, 110–121. [Google Scholar] [CrossRef]
- Perrone, I.; Pagliarani, C.; Lovisolo, C.; Chitarra, W.; Roman, F.; Schubert, A. Recovery from water stress affects grape leaf petiole transcriptome. Planta 2012, 235, 1383–1396. [Google Scholar] [CrossRef]
- Bogs, J.; Downey, M.O.; Harvey, J.S.; Ashton, A.R.; Tanner, G.J.; Robinson, S.P. Proanthocyanidin synthesis and expression of genes encoding leucoanthocyanidin reductase and anthocyanidin reductase in developing grape berries and grapevine leaves. Plant Physiol. 2005, 139, 652–663. [Google Scholar] [CrossRef] [Green Version]
- Cookson, S.J.; Clemente Moreno, M.J.; Hevin, C.; Nyamba Mendome, L.Z.; Delrot, S.; Trossat-Magnin, C.; Ollat, N. Graft union formation in grapevine induces transcriptional changes related to cell wall modification, wounding, hormone signalling, and secondary metabolism. J. Exp. Bot. 2013, 64, 2997–3008. [Google Scholar] [CrossRef]
- Deluc, L.G.; Grimplet, J.; Wheatley, M.D.; Tillett, R.L.; Quilici, D.R.; Osborne, C.; Schooley, D.A.; Schlauch, K.A.; Cushman, J.C.; Cramer, G.R. Transcriptomic and metabolite analyses of cabernet sauvignon grape berry development. BMC Genomics 2007, 8, 429. [Google Scholar] [CrossRef] [Green Version]
- Rattanakon, S.; Ghan, R.; Gambetta, G.A.; Deluc, L.G.; Schlauch, K.A.; Cramer, G.R. Abscisic acid transcriptomic signaling varies with grapevine organ. BMC Plant Biol. 2016, 16, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.; Fan, X.; Zhang, Y.; Jiang, J.; Sun, H.; Liu, C. Transcriptome analysis of genes involved in anthocyanins biosynthesis and transport in berries of black and white spine grapes (vitis davidii). Hereditas 2016, 153, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azuma, A.; Yakushiji, H.; Koshita, Y.; Kobayashi, S. Flavonoid biosynthesis-related genes in grape skin are differentially regulated by temperature and light conditions. Planta 2012, 236, 1067–1080. [Google Scholar] [CrossRef] [PubMed]
- Sun, R.Z.; Cheng, G.; Li, Q.; He, Y.N.; Wang, Y.; Lan, Y.B.; Li, S.Y.; Zhu, Y.R.; Song, W.F.; Zhang, X.; et al. Light-induced variation in phenolic compounds in cabernet sauvignon grapes (Vitis vinifera L.) involves extensive transcriptome reprogramming of biosynthetic enzymes, transcription factors, and phytohormonal regulators. Front. Plant Sci. 2017, 8, 547. [Google Scholar] [CrossRef] [Green Version]
- He, J.J.; Liu, Y.X.; Pan, Q.H.; Cui, X.Y.; Duan, C.Q. Different anthocyanin profiles of the skin and the pulp of yan73 (muscat hamburg× alicante bouschet) grape berries. Molecules 2010, 15, 1141–1153. [Google Scholar] [CrossRef]
- Li, S.Y.; He, F.; Zhu, B.Q.; Xing, R.R.; Reeves, M.J.; Duan, C.Q. A systematic analysis strategy for accurate detection of anthocyanin pigments in red wines. Rapid Commun. Mass Spectrom. 2016, 30, 1619–1626. [Google Scholar] [CrossRef]
- Lan, Y.B.; Qian, X.; Yang, Z.J.; Xiang, X.F.; Yang, W.X.; Liu, T.; Zhu, B.Q.; Pan, Q.H.; Duan, C.Q. Striking changes in volatile profiles at sub-zero temperatures during over-ripening of ‘beibinghong’ grapes in northeastern china. Food Chem. 2016, 212, 172–182. [Google Scholar] [CrossRef]










© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, L.; Ren, Z.-Y.; Wang, Y.; Fu, Y.-Q.; Li, Y.; Meng, N.; Pan, Q.-H. Variation of Growth-to-Ripening Time Interval Induced by Abscisic Acid and Synthetic Auxin affecting Transcriptome and Flavor Compounds in Cabernet Sauvignon Grape Berry. Plants 2020, 9, 630. https://doi.org/10.3390/plants9050630
He L, Ren Z-Y, Wang Y, Fu Y-Q, Li Y, Meng N, Pan Q-H. Variation of Growth-to-Ripening Time Interval Induced by Abscisic Acid and Synthetic Auxin affecting Transcriptome and Flavor Compounds in Cabernet Sauvignon Grape Berry. Plants. 2020; 9(5):630. https://doi.org/10.3390/plants9050630
Chicago/Turabian StyleHe, Lei, Zhi-Yuan Ren, Yu Wang, Ya-Qun Fu, Yue Li, Nan Meng, and Qiu-Hong Pan. 2020. "Variation of Growth-to-Ripening Time Interval Induced by Abscisic Acid and Synthetic Auxin affecting Transcriptome and Flavor Compounds in Cabernet Sauvignon Grape Berry" Plants 9, no. 5: 630. https://doi.org/10.3390/plants9050630
APA StyleHe, L., Ren, Z. -Y., Wang, Y., Fu, Y. -Q., Li, Y., Meng, N., & Pan, Q. -H. (2020). Variation of Growth-to-Ripening Time Interval Induced by Abscisic Acid and Synthetic Auxin affecting Transcriptome and Flavor Compounds in Cabernet Sauvignon Grape Berry. Plants, 9(5), 630. https://doi.org/10.3390/plants9050630