Transcriptome Comparison of Defense Responses in the Rice Variety ‘Jao Hom Nin’ Regarding Two Blast Resistant Genes, Pish and Pik


Abstract
:1. Introduction
2. Results
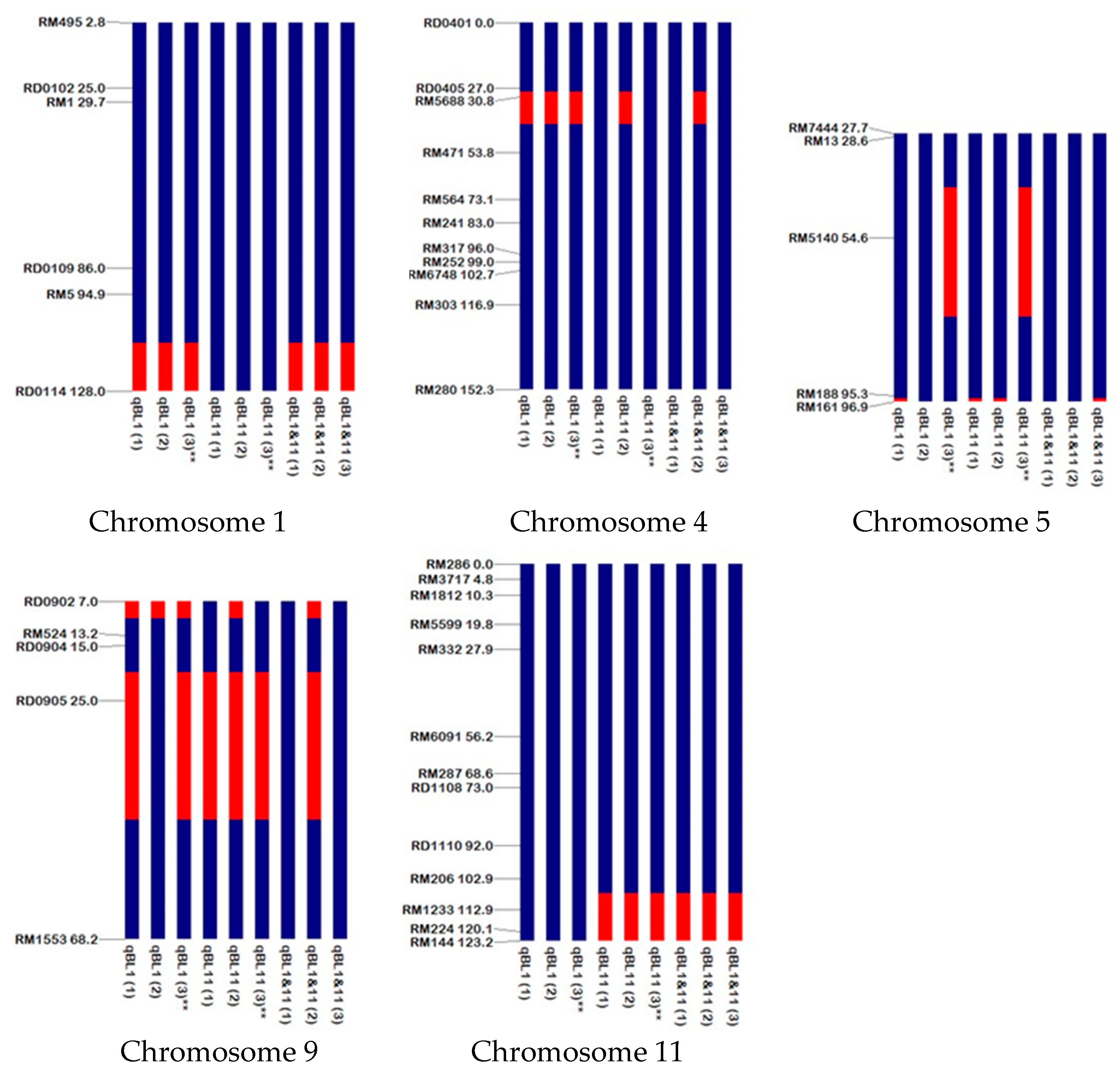
2.1. Graphical Genotyping of RD6 Backcross Inbred Lines
2.2. Transcriptome Sequencing Data of Rice Containing Pish and Pik Genes upon Rice Blast Infection
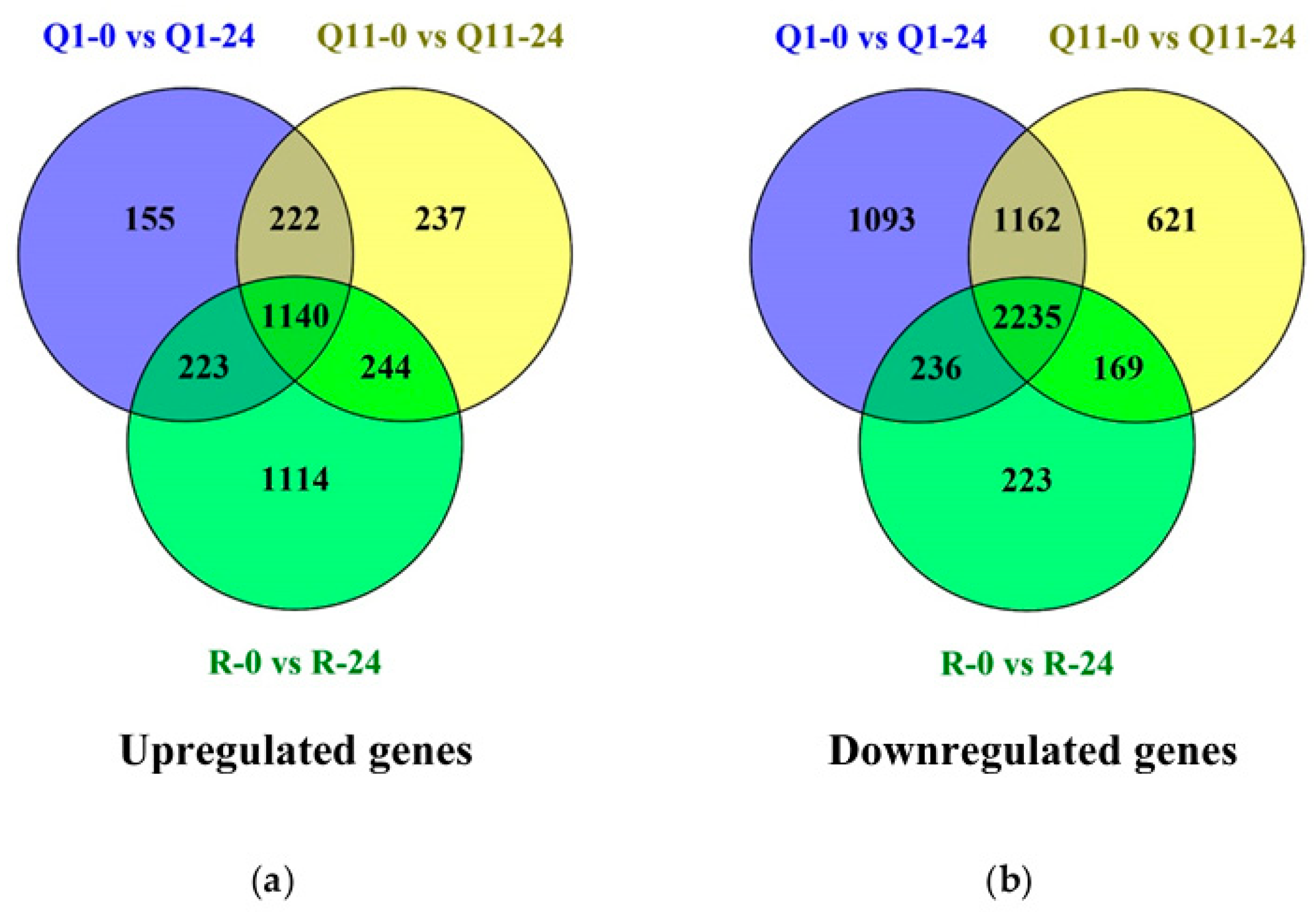
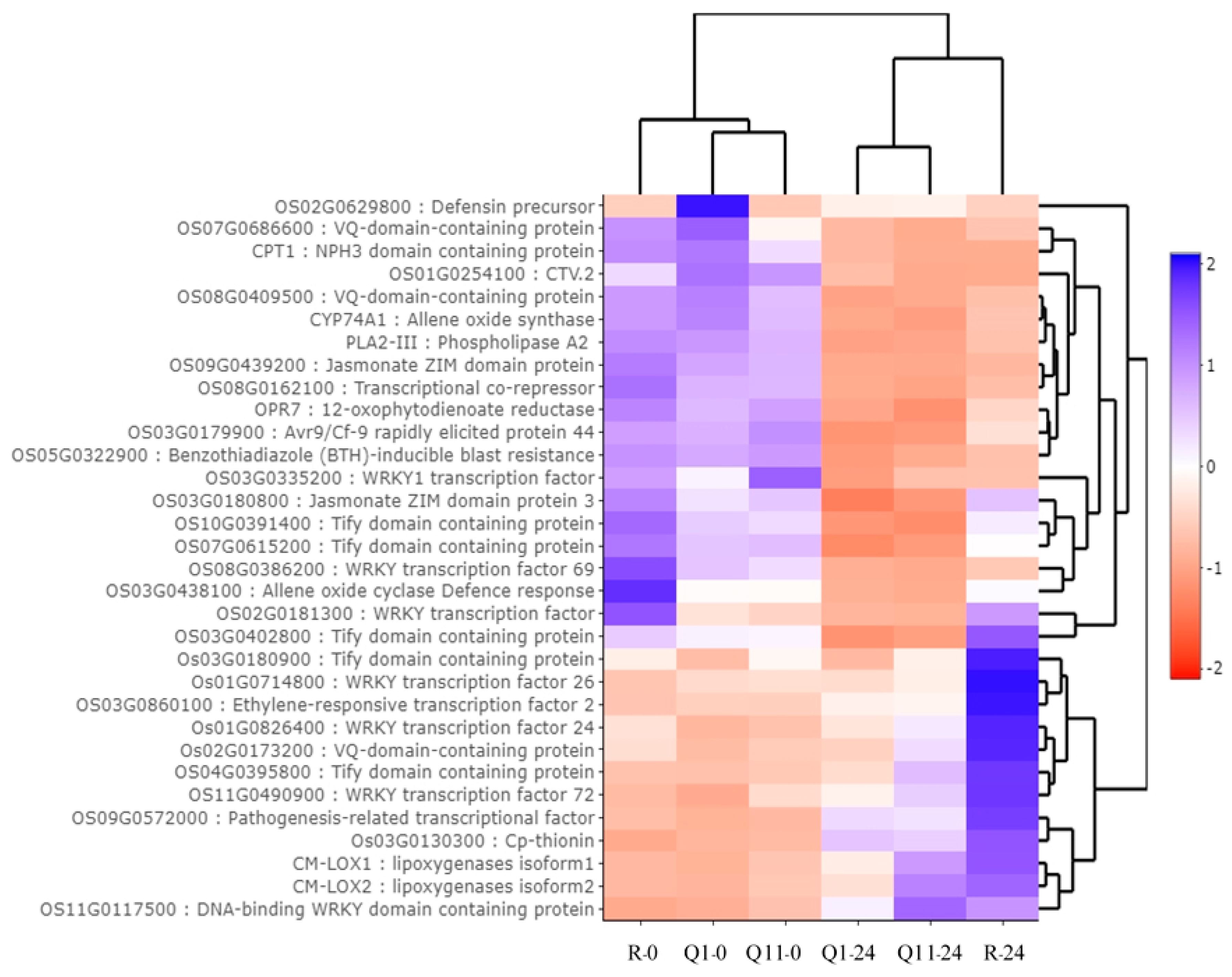
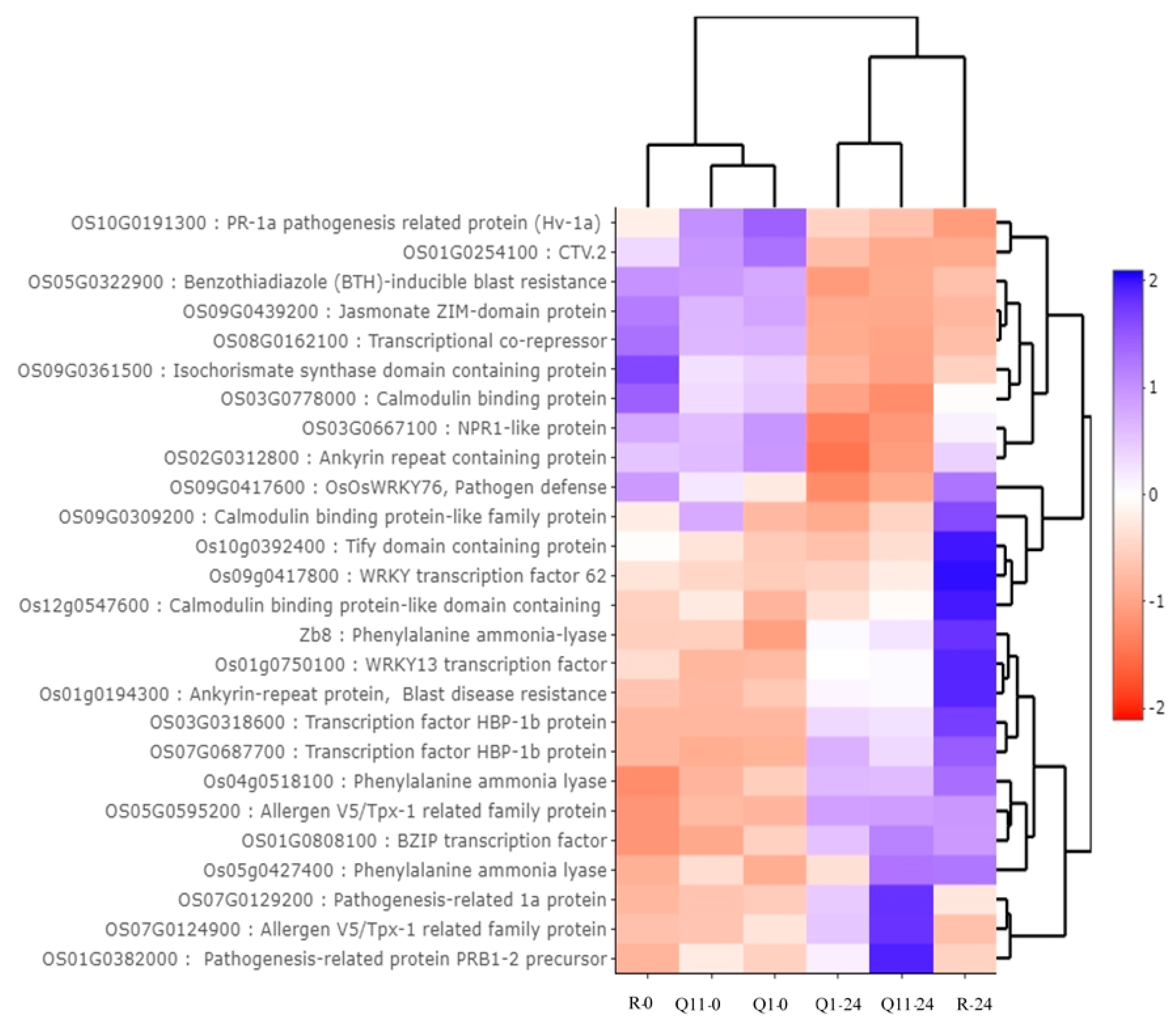
2.3. Identification of Differently Expressed Genes (DEGs) between 0 and 24 H After Innoculation in the Pish-Containing Rice (Q1) and the Pik-Containing Rice (Q11)
2.4. Identification of Gene Ontology Using agriGO
2.5. Identification of Biological Pathway Using Plant Reactome
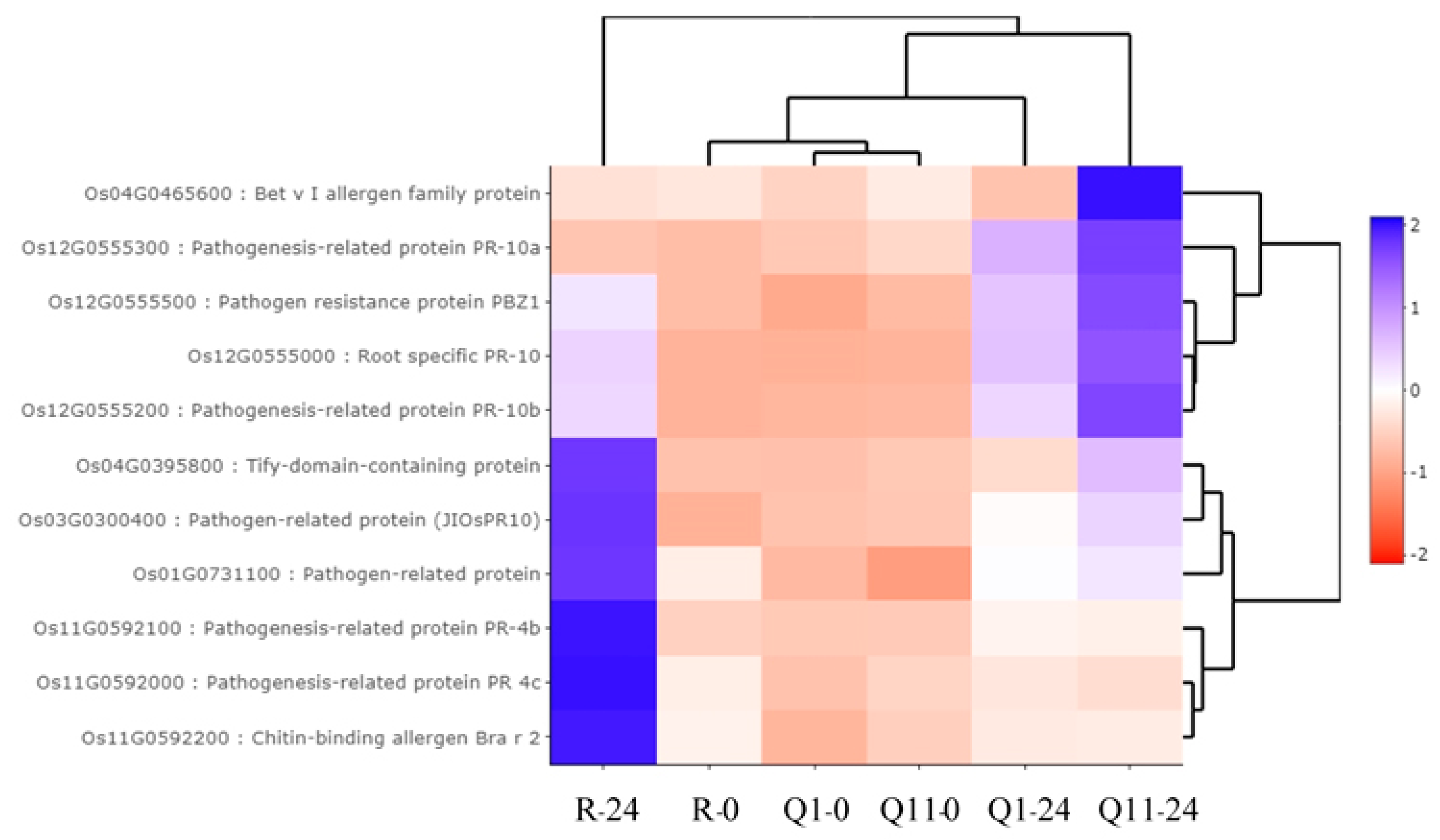
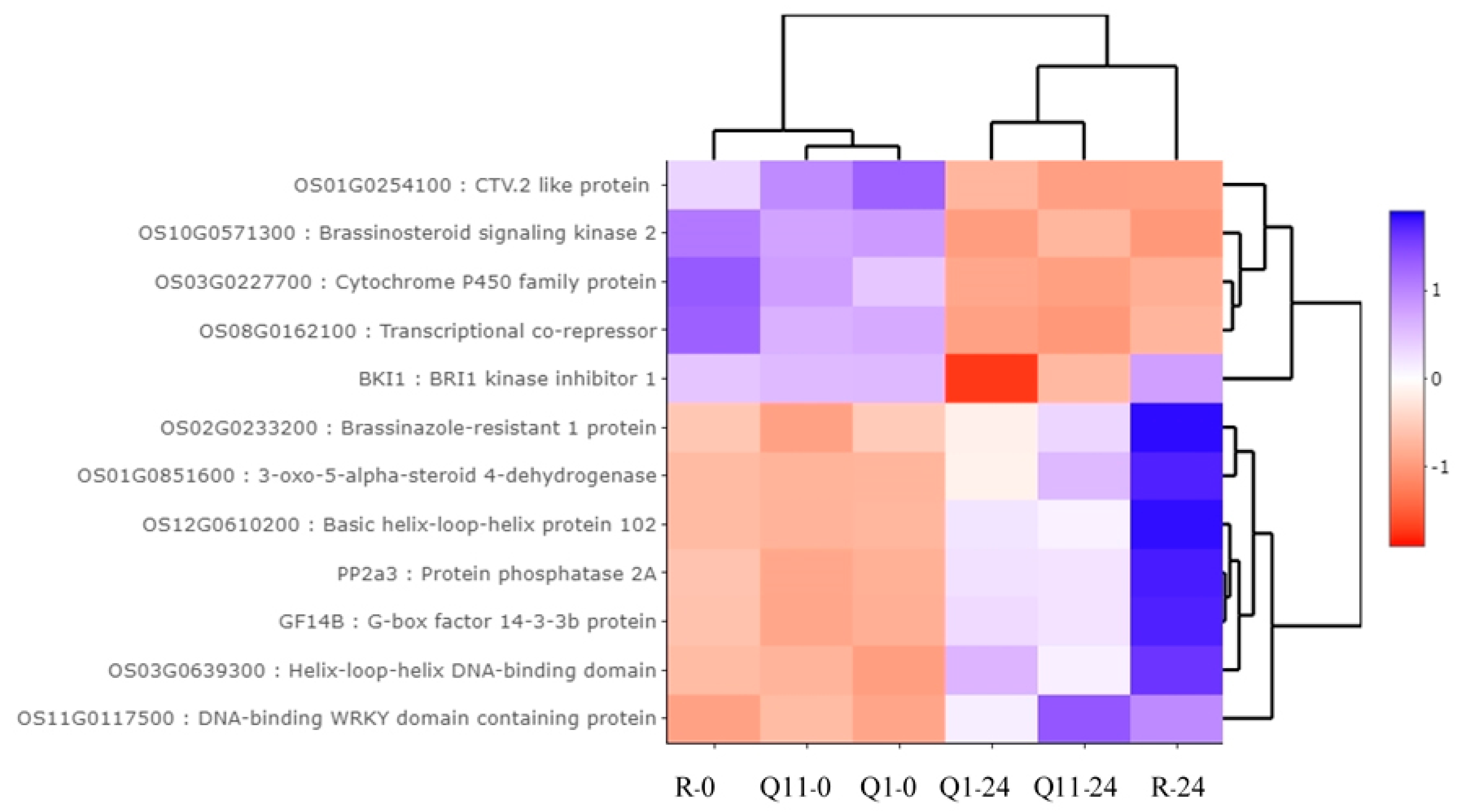
2.6. Validation of RNA-Seq Results by a Quantitative Real-Time PCR (qRT-PCR)
3. Discussion
4. Materials and Methods
4.1. Rice Materials
4.2. Screening of Polymorphic Markers in JHN and RD6
4.3. Graphical Genotyping
4.4. Rice Blast Fungus Inoculation
4.5. Transcriptome Experiment and Data Analysis and Validation of Gene Expression Profiling
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lin, H.C.; Fukushima, Y. Rice cultivation methods and their sustainability aspects: Organic and conventional rice production in industrialized tropical monsoon Asia with a dual cropping system. Sustainability 2015, 8, 529. [Google Scholar] [CrossRef] [Green Version]
- Zibaee, A. Rice: Importance and Future. J. Rice Res. 2013, 1. [Google Scholar] [CrossRef] [Green Version]
- Ansari, M.U.R.; Shaheen, T.; Bukhari, S.A.; Husnain, T. Genetic improvement of rice for biotic and abiotic stress tolerance. Turk. J. Bot. 2015, 39, 911–919. [Google Scholar] [CrossRef]
- Khush, G.S.; Jena, K.K. Current Status and Future Prospects for Research on Blast Resistance in Rice (Oryza sativa L.). In Advances in Genetics, Genomics and Control of Rice Blast Disease; Wang, G.L., Valent, B., Eds.; Springer: Berlin, Germany, 2009; pp. 1–11. [Google Scholar]
- Nalley, L.; Tsiboe, F.; Durand-Morat, A.; Shew, A.; Thoma, G. Economic and environmental impact of rice blast pathogen (Magnaporthe oryzae) alleviation in the United States. PLoS ONE 2016, 11, e0167295. [Google Scholar] [CrossRef]
- Miah, G.; Rafii, M.Y.; Ismail, M.R.; Puteh, A.B.; Rahim, H.A.; Islam, K.N.; Latif, M.A. A review of microsatellite markers and their applications in rice breeding programs to improve blast disease resistance. Int. J. Mol. 2013, 14, 22499–22528. [Google Scholar] [CrossRef] [Green Version]
- Grennan, A.K. Plant response to bacterial pathogens. Overlap between innate and Gene-for-Gene defense response. Plant Physiol. 2006, 142, 809–811. [Google Scholar] [CrossRef] [Green Version]
- Gururani, M.A.; Venkatesh, J.; Upadhayaya, C.P.; Nookaraju, A.; Pandey, S.K.; Park, S.W. Plant disease resistance genes: Current status and future directions. Physiol. Mol. Plant Pathol. 2012, 78, 51–65. [Google Scholar] [CrossRef]
- Prell, H.H.; Day, P. Plant-Fungal Pathogen Interaction: A Classical and Molecular View; Springer: Berlin/Heidelberg, Germany, 2001. [Google Scholar]
- Gachomo, E.W.; Shonukan, O.O.; Kotchoni, S.O. The molecular initiation and subsequent acquisition of disease resistance in plants. Afr. J. Biotechnol. 2003, 2, 26–32. [Google Scholar]
- Tanweer, F.A.; Rafii, M.Y.; Sijam, K.; Rahim, H.A.; Ahmed, F.; Latif, M.A. Current advance methods for the identification of blast resistance genes in rice. Comptes Rendus Biol. 2015, 338, 321–334. [Google Scholar] [CrossRef]
- Wang, X.; Lee, S.; Wang, J.; Ma, J.; Bianco, T.; Jia, Y. Current Advances on Genetic Resistance to Rice Blast Disease. In Rice-Germplasm, Genetics and Improvement; Yan, W.G., Ed.; IntechOpen: London, UK, 2014; pp. 195–217. [Google Scholar]
- Li, W.; Chern, M.; Yin, J.; Wang, J.; Chen, X. Recent advances in broad-spectrum resistance to the rice blast disease. Curr. Opin. Plant Biol. 2019, 50, 114–120. [Google Scholar] [CrossRef]
- Takahashi, A.; Hayashi, N.; Miyao, A.; Hirochika, H. Unique features of the rice blast resistance Pish locus revealed by large scale retrotransposon-tagging. BMC Plant Biol. 2010, 10, 175. [Google Scholar] [CrossRef] [Green Version]
- Kanzaki, H.; Yoshida, K.; Saitoh, H.; Fujidaki, K.; Hirabuchi, A.; Alaux, L.; Fournier, E.; Tharreau, D.; Terauchi, R. Arms race co-evolution of Magnaporthe oryzae AVR-Pik and rice Pik genes driven by their physical interactions. Plant J. 2012, 72, 894–907. [Google Scholar] [CrossRef]
- Corwin, J.A.; Kliebenstein, D.J. Quantitative Resistance: More than Just Perception of a Pathogen. Plant Cell. 2017, 29, 655–665. [Google Scholar] [CrossRef] [Green Version]
- Jiang, G.L. Molecular Markers and Marker-Assisted Breeding in Plants. In Plant Breeding from Laboratories to Fields; Andersen, S.B., Ed.; IntechOpen: London, UK, 2013; pp. 45–83. [Google Scholar]
- Lema, M. Marker assisted selection in comparison to conventional plant breeding: Review article. Agric. Res. Technol. Open Access J. 2018, 14, 1–10. [Google Scholar] [CrossRef] [Green Version]
- van Eck, H.J.; Vos, P.G.; Valkonen, J.P.T.; Uitdewilligen, J.G.A.M.L.; Lensing, H.; de Vetten, N.; Visser, R.G.F. Graphical genotyping as a method to map Ny (o,n)sto and Gpa5 using a reference panel of tetraploid potato cultivars. Theor. Appl. Genet. 2017, 130, 515–528. [Google Scholar] [CrossRef] [Green Version]
- Dong, Z.C.; Yan, C. Transcriptomics: Advance and Approaches. Sci. China Life Sci. 2013, 56, 960–967. [Google Scholar] [CrossRef] [Green Version]
- Manthioni, S.M.; Beló, A.; Rizzo, C.J.; Dean, R.A.; Donofrio, N.M. Transcriptome profiling of the rice blast fungus during invasive plant infection and in vitro stress. BMC Genom. 2011, 12, 49. [Google Scholar]
- Wang, Y.; Kwon, S.J.; Wu, J.; Choi, J.; Lee, Y.H.; Agrawal, G.K.; Tamogami, S.; Rakwal, R.; Park, S.R.; Kim, B.G.; et al. Transcriptome analysis of early responsive genes in rice during Magnaporthe oryzae infection. Plant Pathol. J. 2014, 30, 343–354. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.K.; Rai, A.K.; Kanwar, S.S.; Chand, D.; Singh, N.K.; Sharma, T.R. The single functional blast resistance gene Pi54 activate a complex defence mechanism in rice. J. Exp. Bot. 2011, 63, 757–772. [Google Scholar] [CrossRef] [Green Version]
- Wolf, J.B.W. Principles of transcriptome analysis and gene expression quantification: An RNA-seq tutorial. Mol. Ecol. Resour. 2013, 13, 559–572. [Google Scholar] [CrossRef]
- Nagalakshmi, U.; Waern, K.; Snyder, M. RNA-Seq: A method for comprehensive transcriptome analysis. Curr. Protoc. Mol. Biol. 2010, 89, 4–11. [Google Scholar]
- Bagnaresi, P.; Biselli, C.; Orru, L.; Urso, S.; Crispino, L.; Abbruscato, P.; Piffanelli, P.; Lupotto, E.; Cattivelli, L.; Vale, G. Comparative transcriptome profiling of the early response to Magnaporthe oryzae in durable resistant vs susceptible rice (Oryza sativa L.) genotypes. PLoS ONE 2012, 7, e51609. [Google Scholar] [CrossRef] [Green Version]
- Noenplab, A.; Vanavichit, A.; Toojinda, T.; Sirithunya, P.; Tragoonrung, S.; Sriprakkhon, S.; Vongsaprom, C. QTL mapping for leaf and neck blast resistance in Khao Dawk Mali105 and Jao Hom Nin recombinant inbred lines. Sci. Asia 2006, 32, 133–142. [Google Scholar] [CrossRef]
- Sreewongchai, T.; Toojinda, T.; Thanintorn, N.; Kosawang, C.; Vanavichit, A.; Tharreau, D.; Sirithunya, P. Development of elite indica rice lines with wide spectrum of resistance to Thai blast isolates by pyramiding multiple resistance QTLs. Plant Breed. 2010, 129, 176–180. [Google Scholar] [CrossRef]
- Wongsaprom, C.; Sirithunya, P.; Vanavichit, A.; Pantuwan, G.; Jongdee, B.; Sidhiwong, N.; Lanceras-Siangliw, J.; Toojinda, T. Two introgressed quantitative trait loci confer a broad-spectrum resistance to blast disease in the genetic background of the cultivar RD6 a Thai glutinous jasmine rice. Field Crops Res. 2010, 199, 245–251. [Google Scholar] [CrossRef]
- Chaipanya, C.; Telebanco-Yanoria, M.J.; Quime, B.; Longya, A.; Korinsak, S.; Korinsak, S.; Toojinda, T.; Vanavichit, A.; Jantasuriyarat, C.; Zhou, B. Dissection of broad-spectrum resistance of the Thai rice variety Jao Hom Nin conferred by two resistance genes against rice blast. Rice 2017, 10, 18. [Google Scholar] [CrossRef]
- Wu, D.H.; Wu, H.P.; Wang, C.S.; Tseng, H.Y.; Hwu, K.K. Genome-wide InDel marker system for application in rice breeding and mapping studies. Euphytica 2013, 192, 131–143. [Google Scholar]
- Tongnun, P.; Prasongmaneerut, T.; Sriwongchai, T.; Jantasuriyarat, C. Graphical genotype of KDML105xIR64 backcross lines exhibited rice blast resistance. Chiang Mai J. Sci. 2017, 44, 1322–1333. [Google Scholar]
- Zhao, K.; Wright, M.; Kimball, J.; Eizenga, G.; McClung, A.; Kovach, M.; Tyagi, W.; Ali, M.L.; Tung, C.W.; Reynolds, A.; et al. Genomic Diversity and Introgression in O. sativa Reveal the Impact of Domestication and Breeding on the Rice Genome. PLoS ONE 2010, 5, e10780. [Google Scholar]
- Kohorn, B.D.; Kohorn, S.L. The cell wall-associated kinases, WAKs, as pectin receptors. Front. Plant Sci. 2012, 3, 88. [Google Scholar]
- Magwanga, R.O.; Lu, P.; Kirungu, J.N.; Lu, H.; Wang, X.; Cai, X.; Zhou, Z.; Zhang, Z.; Salih, H.; Wang, K.; et al. Characterization of the late embryogenesis abundant (LEA) proteins family and their role in drought stress tolerance in upland cotton. BMC Genet. 2018, 19, 6. [Google Scholar]
- Vignesh, K.S.; Deepe, G.S., Jr. Metallothioneins: Emerging modulators in immunity and infection. Int. J. Mol. Sci. 2017, 18, 2197. [Google Scholar]
- Gupta, S.K.; Rai, A.K.; Kanwar, S.S.; Sharma, T.R. Comparative analysis of zinc finger proteins involved in plant disease resistance. PLoS ONE 2012, 7, e42578. [Google Scholar] [CrossRef] [Green Version]
- Ambawat, S.; Sharma, P.; Yadav, N.R.; Yadav, R.C. MYB transcription factor genes as regulators for plant responses: An overview. Physiol. Mol. Biol. Plants 2013, 19, 307–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Punja, Z.K.; Zhang, Y.Y. Plant chitinases and their roles in resistance to fungal disease. J. Nematol. 1993, 25, 526–540. [Google Scholar]
- Figueiredo, J.; Silva, M.S.; Figueiredo, A. Substilisin-like proteases in plant defense: The past, the present and beyond. Mol. Plant Pathol. 2018, 19, 1017–1028. [Google Scholar]
- Jain, S.; Kumar, A. The pathogenesis related class 10 proteins in plant defense against biotic and abiotic stresses. APAR 2015, 2, 1–11. [Google Scholar] [CrossRef]
- Guevara-Morato, M.A.; de Lacoba, M.G.; Garcia-Luque, I.; Serra, M.T. Characterization of a pathogenesis-related protein 4 (PR-4) induced in Capsicum chinense L3 plants with dual RNase and DNase activities. J. Exp. Bot. 2010, 61, 3259–3271. [Google Scholar]
- Kim, S.G.; Kim, S.T.; Wang, Y.; Yu, S.; Choi, I.S.; Kim, Y.C.; Kim, W.T.; Agrawal, G.K.; Rakwal, R.; Kang, K.Y. The RNase activity of rice probenazole-induced protein1 (PBZ1) plays a key role in cell death in plants. Mol. Cells 2011, 31, 25–31. [Google Scholar] [CrossRef]
- Deng, X.G.; Zhu, T.; Peng, X.J.; Xi, D.H.; Guo, H.; Yin, Y.; Zhang, D.W.; Lin, H.H. Role of brassinosteroids signaling in modulating Tobacco mosaic virus resistance in Nicotiana benthamiana. Sci. Rep. 2015, 6, 20579. [Google Scholar]
- Wang, X.; Chory, J. Brassinosteroids regulate dissociation of BKI1, a negative regulator of BRI1 signaling, from the plasma membrane. Science 2006, 313, 1118–1122. [Google Scholar] [CrossRef] [PubMed]
- Vidhyasekaran, P. Brassinosteroids Signaling in Plant Immune System. In Plant Hormone Signaling Systems in Plant Innate Immunity; Vidhyasekaran, P., Ed.; Springer: Dordrecht, The Netherlands, 2014; pp. 403–444. [Google Scholar]
- Carvalhais, L.C.; Schenk, P.M.; Dennis, P.G. Jasmonic acid signaling and the plant halobiont. Curr. Opin. Microbiol. 2013, 37, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Kolomiets, M.V.; Hannapel, D.J.; Chen, H.; Tymeson, M.; Gladon, R.J. Lipoxygenase is involved in the control of potato tuber development. Plant Cell 2000, 13, 613–626. [Google Scholar] [CrossRef] [PubMed]
- Saha, G.; Park, J.I.; Kayum, M.A.; Nou, I.S. A genome-wide analysis reveals stress and hormone responsive patterns of TIFY family genes in Brassica rapa. Front. Plant Sci. 2016, 7, 936. [Google Scholar] [CrossRef] [Green Version]
- Phukan, U.J.; Jeena, G.S.; Shukla, R.K. WRKY transcription factors: Molecular regulation and stress responses in plants. Front. Plant Sci. 2016, 7, 760. [Google Scholar] [CrossRef] [Green Version]
- War, A.R.; Paulraj, M.G.; War, M.Y.; Ignacimuthu, S. Role of salicylic acid in induction of plant defense system in chickpea (Cicer arietinum L.). Plant Signal Behav. 2011, 6, 1787–1792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mouradov, A.; Mouradova, E.; Scott, K.J. Gene family encoding basic pathogenesis-related 1 proteins in barley. Plant Mol. Biol. 1994, 26, 503–507. [Google Scholar] [CrossRef] [PubMed]
- Llorca, C.M.; Potschin, M.; Zentgraf, U. bZIPs and WRKYs: Two large transcription factor families executing two different functional strategies. Front. Plant Sci. 2014, 5, 169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, M.L.; Zhang, Y.H.; Meng, Q.L.; Shi, F.M.; Liu, J.; Li, Y.C.; Wang, Z.Y. Cloning and function analysis of pathogenesis related protein gene HaPR1 from sunflower (Helianthus annuus). AAS 2015, 41, 1819–1827. [Google Scholar]
- Miche, L.; Battistomni, F.; Gemmer, S.; Belghazi, M.; Hurek, B.R. Upregulation of jasmonate-inducible defense proteins and differential colonization of roots of Oryza sativa cultivars with the endophyte Azoarcus sp. Mol. Plant Microbe Interact. 2006, 19, 502–511. [Google Scholar] [CrossRef] [Green Version]
- Kawahara, Y.; Oono, Y.; Kanamori, H.; Matsumoto, T.; Itoh, T.; Minami, E. Simultaneous RNA-seq analysis of a mixed transcriptome of rice and blast fungus interaction. PLoS ONE 2012, 7, e49423. [Google Scholar]
- Zhang, Y.; Zhao, J.; Li, Y.; Yuan, Z.; He, H.; Yang, H.; Qu, H.; Ma, C.; Qu, S. Transcriptome analysis highlights defense and signaling pathways mediated by rice pi21 gene with partial resistance to Magnaporthe oryzae. Front. Plant Sci. 2016, 7, 1834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Berloo, R. Computer note. GGT: Software for the display of graphical genotypes. J. Hered 1999, 90, 328–329. [Google Scholar] [CrossRef] [Green Version]
- Roumen, E.; Levy, M.; Notteghem, J.L. Characterisation of the european pathogen population of Magnaporthe grisea by DNA fingerprinting and pathotype analysis. Eur. J. Plant Pathol. 1997, 103, 363–371. [Google Scholar] [CrossRef]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential Gene and Transcript Expression Analysis of RNA-seq Experiments with TopHat and Cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Son, K.; Yu, S.; Shin, W.; Han, K.; Kang, K. A Simple Guideline to Assess the Characteristics of RNA-Seq Data. Hindawi 2018, 2018, 2906292. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Unique to Q1 | |||
| No. | Gene ID | Gene Description | Log2(FC) |
| 1 | OS06G0573500 | Hypothetical conserved gene | 6.439 |
| 2 | OS11G0440866 | Non-protein coding transcript | 5.958 |
| 3 | LEA3 | Late embryogenesis abundant (LEA) protein | 5.354 |
| 4 | OS03G0573750 | Conserved hypothetical protein | 5.141 |
| 5 | OS08G0190100 | Germin-like protein 8-11, disease resistance | 5.043 |
| 6 | OS01G0899800 | Pathogenesis-related transcriptional factor | 4.965 |
| 7 | OS09G0109600 | Conserved hypothetical protein | 4.552 |
| 8 | OS04G0617900 | Similar to germin-like protein subfamily 1 | 4.445 |
| 9 | OS10G0180800 | Wall-associated kinase | 4.400 |
| 10 | OS12G0571000 | Metallothionein-like protein type 1 | 4.150 |
| 11 | RAB16D | Dehydrin RAB 16D | 3.984 |
| 12 | OS12G0623400 | Conserved hypothetical protein | 3.839 |
| 13 | OS03G0575500 | Conserved hypothetical protein | 3.826 |
| 14 | OS01G0223000 | Lipase | 3.764 |
| 15 | OS10G0418900 | Hypothetical conserved gene | 3.666 |
| 16 | OS03G0223301 | Hypothetical gene | 3.490 |
| 17 | OS01G0757200 | GA 2-oxidase3, GA metabolism | 3.482 |
| 18 | OS10G0147400 | Similar to Auxin influx carrier protein | 3.300 |
| 19 | OS02G0269650 | Hypothetical gene | 3.238 |
| 20 | OS02G0627700 | Similar to ATCNGC15 | 3.168 |
| Unique to Q11 | |||
| No. | Gene ID | Gene Description | Log2(FC) |
| 1 | OS05G0454200 | Guanine nucleotide exchange factors for Rop | 6.649 |
| 2 | OS09G0240200 | Zinc finger, B-box domain containing protein | 5.339 |
| 3 | OS02G0514150 | Hypothetical conserved gene | 5.186 |
| 4 | OS05G0261001 | Non-protein coding transcript | 4.767 |
| 5 | EPlOSAG00000006021 | n.a. | 4.695 |
| 6 | OS08G0335600 | Protein of unknown function DUF568 | 4.681 |
| 7 | OS02G0747900 | Atypical basic helix–loop–helix protein | 4.495 |
| 8 | OS08G0486300 | Similar to P-type R2R3 Myb protein | 4.407 |
| 9 | MADS5 | MADS-box transcription factor | 4.323 |
| 10 | OS07G0600000 | Conserved hypothetical protein | 4.185 |
| 11 | OS11G0134950 | Hypothetical protein | 4.090 |
| 12 | OS02G0572000 | Hypothetical protein | 4.087 |
| 13 | OS11G0701400 | Chitinase (EC 3.2.1.14) III C10150-rice | 4.069 |
| 14 | OS04G0465300 | A member of the GAST (gibberellin (GA)-Stimulated Transcript) | 3.980 |
| 15 | OS01G0725400 | Uncharacterized protein family UPF0497 | 3.952 |
| 16 | OS10G0107866 | Non-protein coding transcript | 3.926 |
| 17 | OS06G0711900 | Bifunctional inhibitor/plant lipid transfer protein | 3.911 |
| 18 | OS12G0157066 | Hypothetical protein | 3.809 |
| 19 | OS10G0463200 | Esterase, SGNH hydrolase-type domain containing protein. | 3.764 |
| 20 | OS01G0728100 | Lipase, GDSL-domain-containing protein. | 3.714 |
| Top 10 Upregulated DEGs of Q1 | ||||
| No. | Gene ID | Gene Description | Log2FC | |
| Q1 | Q11 | |||
| 1 | OS10G0150800 | Protein of unknown function DUF1210 family protein | 6.738 | 6.007 |
| 2 | OS02G0185900 | Hypothetical conserved gene | 6.532 | 6.011 |
| 3 | OS10G0150700 | Protein of unknown function DUF1210 family protein. | 6.310 | 6.179 |
| 4 | OS05G0375466 | Non-protein coding transcript | 6.152 | 7.593 |
| 5 | OS08G0189850 | Germin-like protein 8-9, disease resistance | 5.874 | 5.836 |
| 6 | OS09G0319800 | Terpene synthase-like domain containing protein | 5.842 | 8.216 |
| 7 | OS04G0664900 | Similar to H1005F08.5 protein | 5.596 | 7.617 |
| 8 | OS10G0150400 | Protein of unknown function DUF1210 family protein | 5.515 | 4.796 |
| 9 | OS02G0827400 | Similar to predicted protein | 5.428 | 3.844 |
| 10 | OS09G0315000 | Similar to mpv17/PMP22 family protein | 5.363 | 4.639 |
| Top 10 Upregulated DEGs of Q11 | ||||
| No. | Gene ID | Gene Description | Log2FC | |
| Q1 | Q11 | |||
| 1 | OS09G0319800 | Terpene synthase-like domain containing protein | 5.842 | 8.216 |
| 2 | OS11G0701500 | Similar to class III chitinase homologue (OsChib3H-g) | 3.739 | 7.891 |
| 3 | OS04G0664900 | Similar to H1005F08.5 protein | 5.596 | 7.617 |
| 4 | OS05G0375466 | Non-protein coding transcript | 6.152 | 7.593 |
| 5 | OS10G0150700 | Protein of unknown function DUF1210 family protein | 6.31 | 6.179 |
| 6 | OS02G0185900 | Hypothetical conserved gene | 6.532 | 6.011 |
| 7 | OS10G0150800 | Protein of unknown function DUF1210 family protein | 6.738 | 6.007 |
| 8 | OS08G0189850 | Germin-like protein 8-9, disease resistance | 5.874 | 5.836 |
| 9 | OS07G0124900 | Allergen V5/Tpx-1-related family protein | 1.646 | 5.750 |
| 10 | OS03G0291200 | Protein of unknown function DUF231, plant-domain-containing protein | 3.485 | 5.408 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ngernmuen, A.; Suktrakul, W.; Kate-Ngam, S.; Jantasuriyarat, C. Transcriptome Comparison of Defense Responses in the Rice Variety ‘Jao Hom Nin’ Regarding Two Blast Resistant Genes, Pish and Pik. Plants 2020, 9, 694. https://doi.org/10.3390/plants9060694
Ngernmuen A, Suktrakul W, Kate-Ngam S, Jantasuriyarat C. Transcriptome Comparison of Defense Responses in the Rice Variety ‘Jao Hom Nin’ Regarding Two Blast Resistant Genes, Pish and Pik. Plants. 2020; 9(6):694. https://doi.org/10.3390/plants9060694
Chicago/Turabian StyleNgernmuen, Athipat, Worrawit Suktrakul, Sureeporn Kate-Ngam, and Chatchawan Jantasuriyarat. 2020. "Transcriptome Comparison of Defense Responses in the Rice Variety ‘Jao Hom Nin’ Regarding Two Blast Resistant Genes, Pish and Pik" Plants 9, no. 6: 694. https://doi.org/10.3390/plants9060694
APA StyleNgernmuen, A., Suktrakul, W., Kate-Ngam, S., & Jantasuriyarat, C. (2020). Transcriptome Comparison of Defense Responses in the Rice Variety ‘Jao Hom Nin’ Regarding Two Blast Resistant Genes, Pish and Pik. Plants, 9(6), 694. https://doi.org/10.3390/plants9060694