The Serine Carboxypeptidase-Like Gene SCPL41 Negatively Regulates Membrane Lipid Metabolism in Arabidopsis thaliana


Abstract
:1. Introduction
2. Results
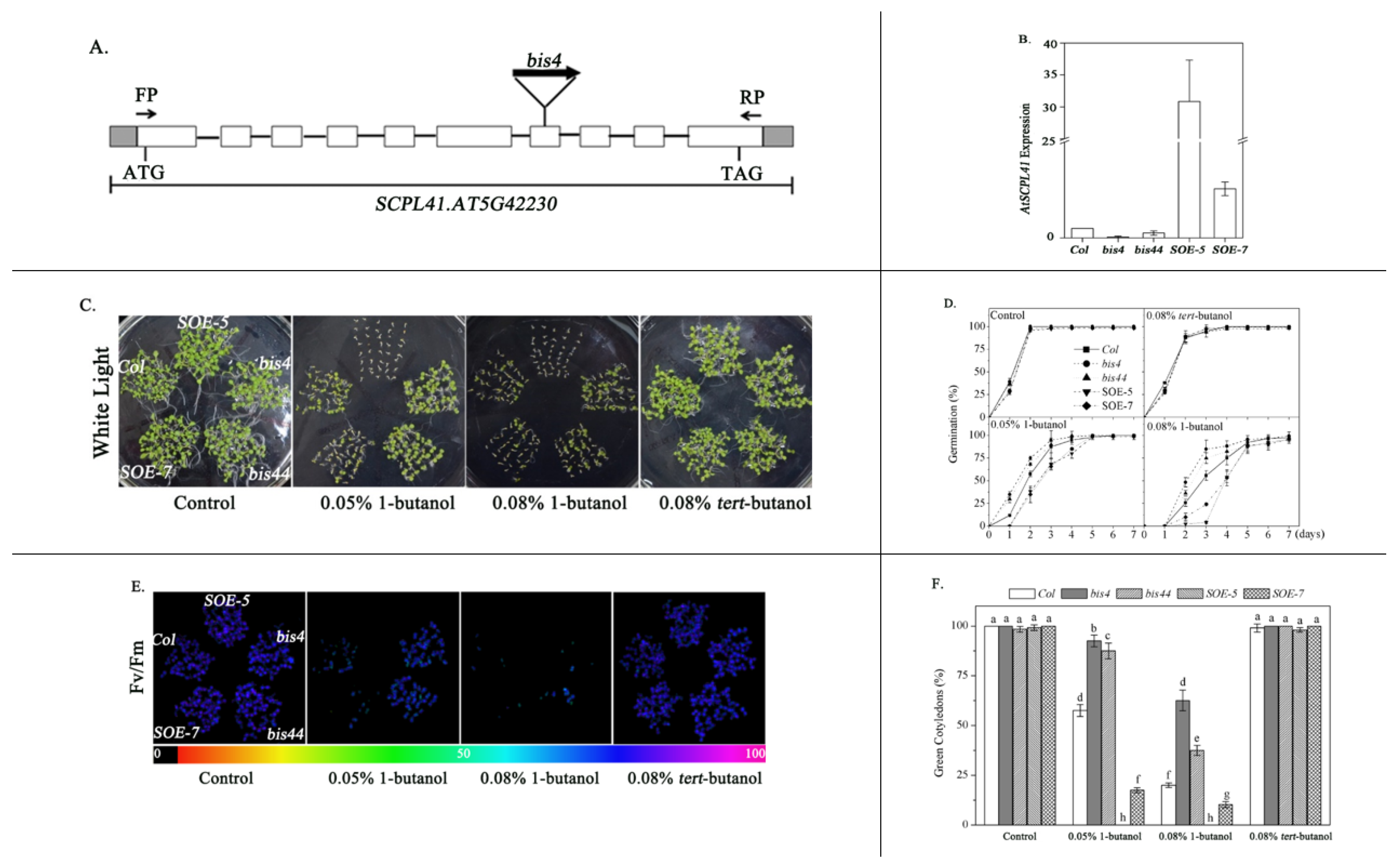
2.1. Identification of the SCPL41 Protein Involved in 1-Butanol Response
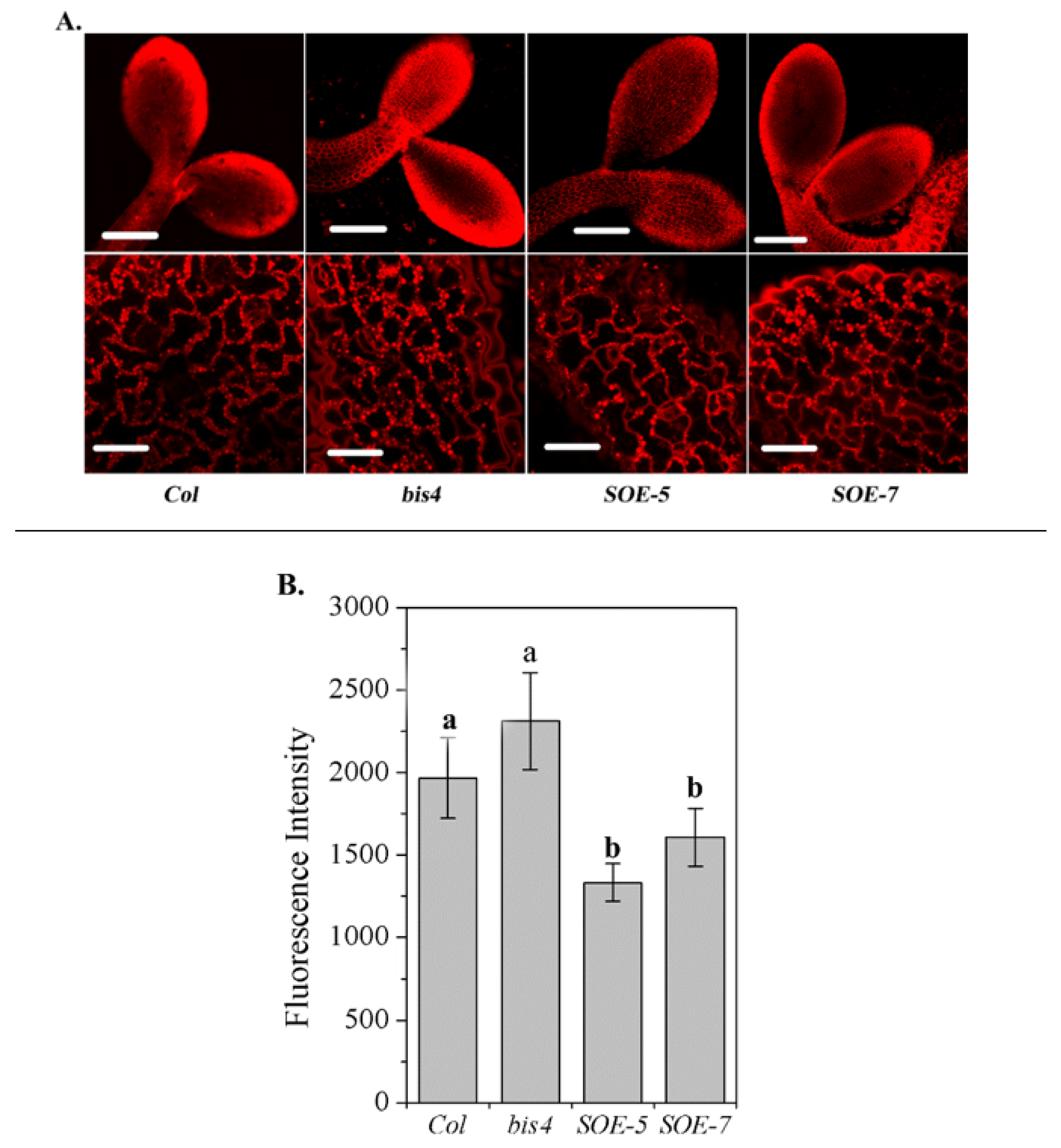
2.2. SCPL41 Had Different Effects on The Content and Composition of Lipids
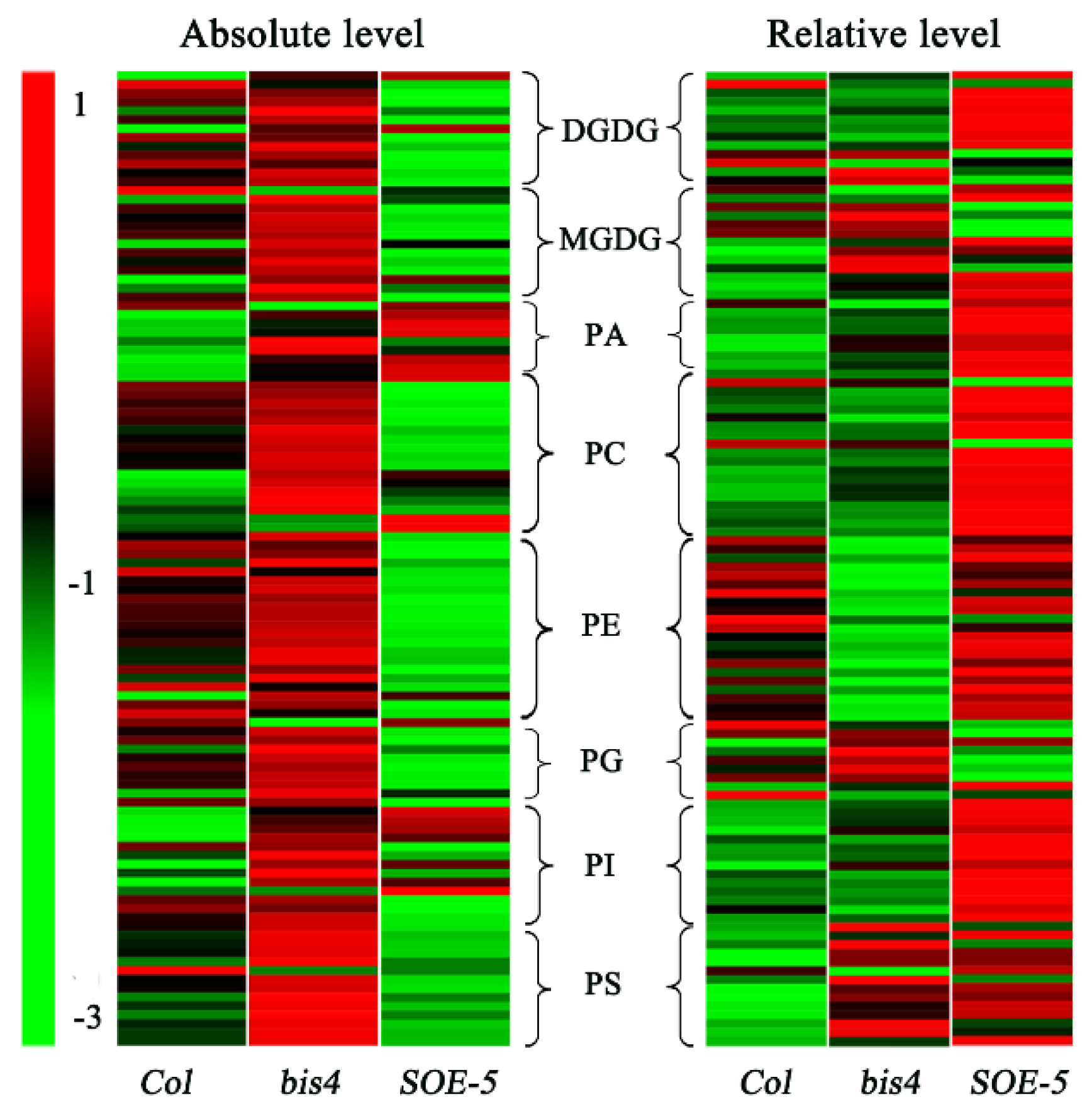
2.3. Overexpression of SCPL41 Decreased the Membrane Lipid Content, but Loss of SCPL41 Increased the Membrane Lipid Content
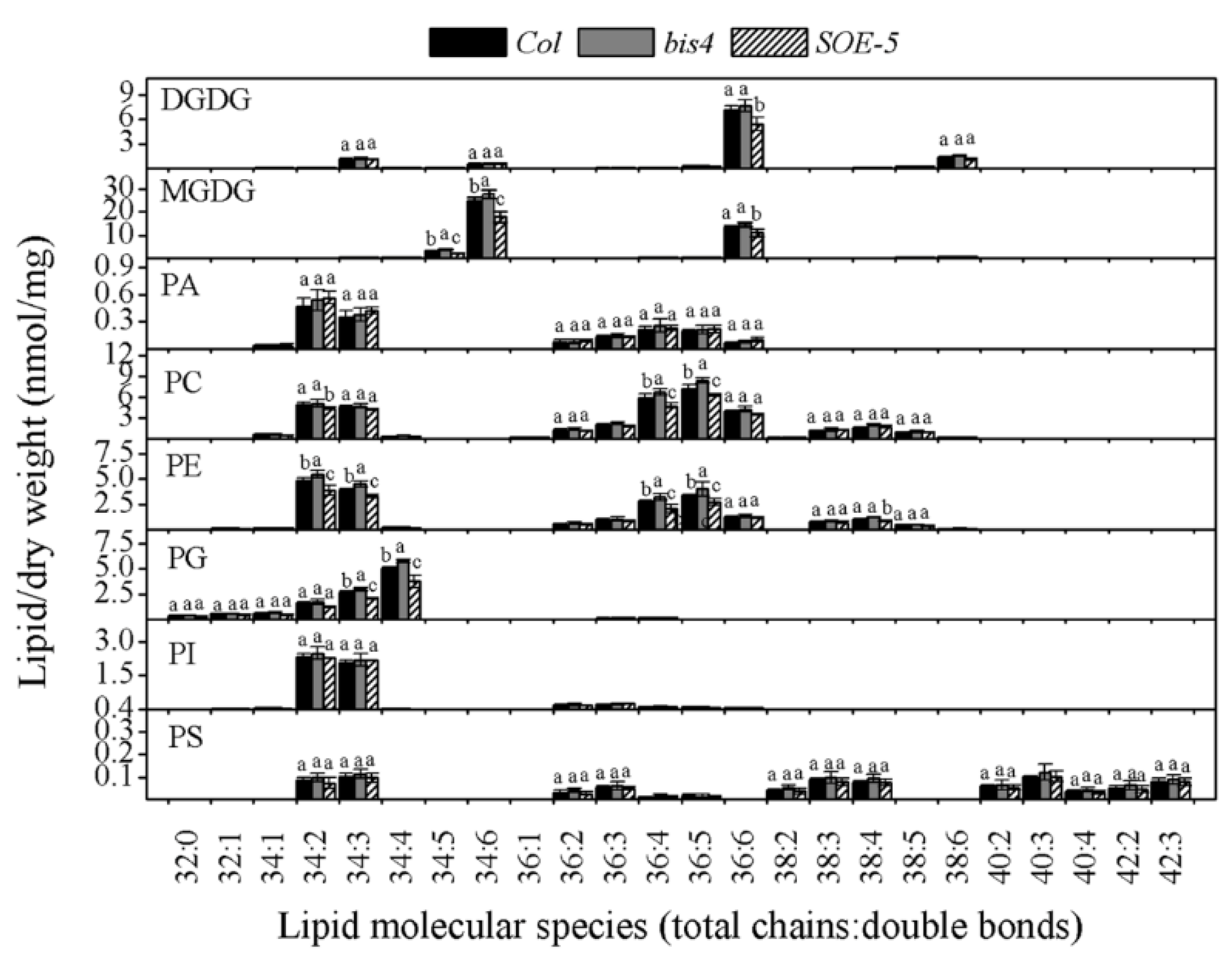
2.4. SCPL41 Had No Effect on the Composition of Lipid Classes
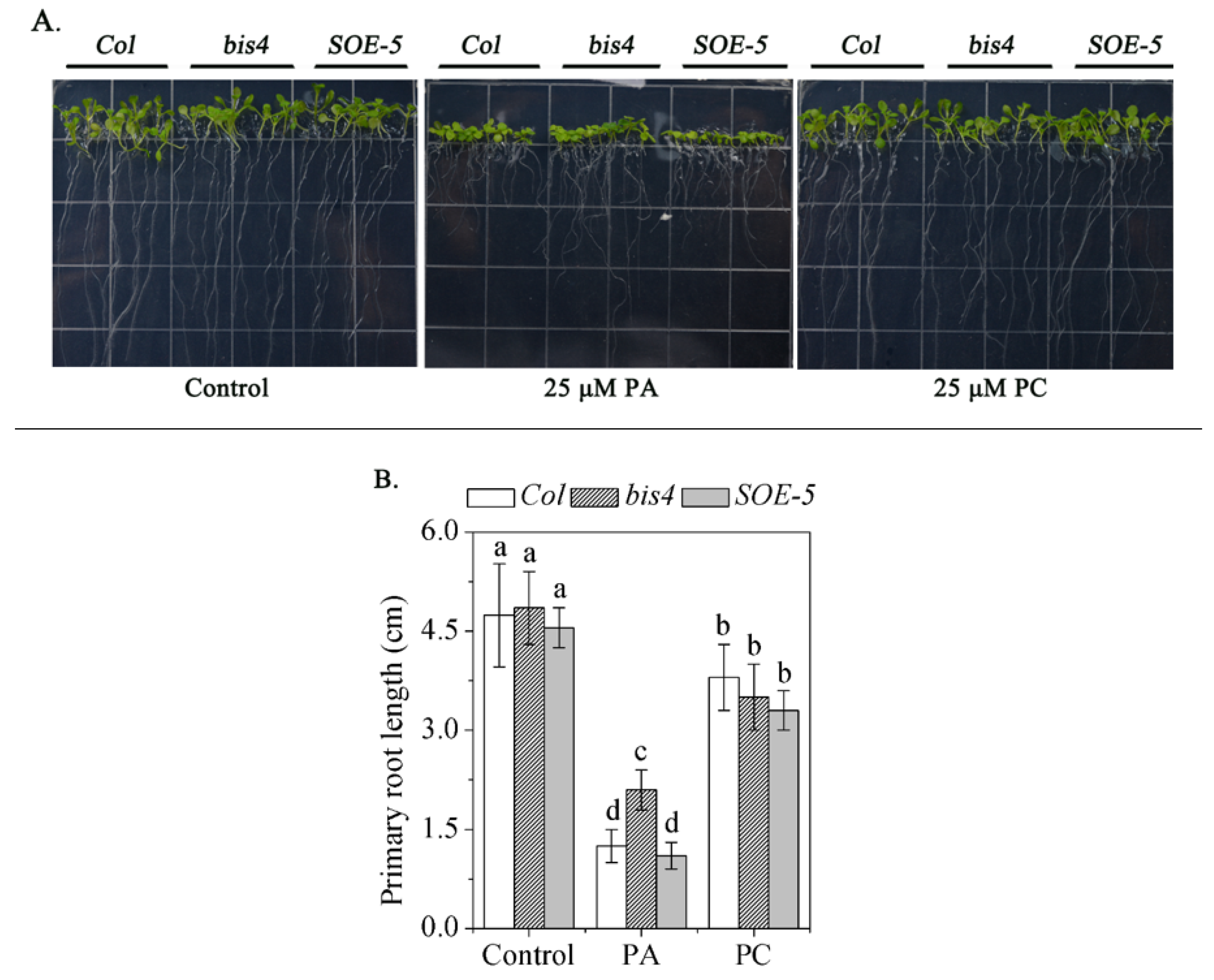
2.5. SCPL41 Had No Effect on the PA content, But Might be Involved in PA Signaling
3. Discussion
4. Material and Methods
4.1. Plant Materials and Growth Conditions
4.2. Phenotype Analysis
4.3. Transgenic Plants
4.4. Nile Red Staining
4.5. Lipid Analysis and Data Processing
4.6. RNA Isolation and Quantitative PCR
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| SCPL | serine carboxypeptidase-like |
| PG | phosphatidylglycerol |
| PC | phosphatidylcholine |
| PE | phosphatidylethanolamine |
| PI | phosphatidylinositol |
| PS | phosphatidylserine |
| PA | phosphatidic acid |
| MGDG | monogalactosyldiacylglycerol |
| DGDG | digalactosyldiacylglycerol |
| DBI | double bond index |
| ESI-MS/MS | electrospray ionization tandem mass spectrometry |
References
- Feng, Y.; Xue, Q. The serine carboxypeptidase like gene family of rice (Oryza sativa L. ssp. japonica). Funct. Integr. Genom. 2006, 6, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Mugford, S.T.; Milkowski, C. Serine carboxypeptidase-like acyltransferases from plants. Methods Enzym. 2012, 516, 279–297. [Google Scholar]
- Jiang, P.; Gao, J.; Mu, J.; Duan, L.; Gu, Y.; Han, S.; Chen, L.; Li, Y.; Yan, Y.; Li, X. Interaction between serine carboxypeptidase-like protein TtGS5 and Annexin D1 in developing seeds of Triticum timopheevi. Appl. Genet. 2020, 61, 151–162. [Google Scholar] [CrossRef] [PubMed]
- Fraser, C.M.; Rider, L.W.; Chapple, C. An expression and bioinformatics analysis of the Arabidopsis serine carboxypeptidase-like gene family. Plant Physiol. 2005, 138, 1136–1148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dal Degan, F.; Rocher, A.; Cameron-Mills, V.; von Wettstein, D. The expression of serine carboxypeptidases during maturation and germination of the barley grain. Proc. Natl. Acad. Sci. USA 1994, 91, 8209–8213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bullock, T.L.; Breddam, K.; Remington, S.J. Peptide aldehyde complexes with wheat serine carboxypeptidase II: Implications for the catalytic mechanism and substrate specificity. J. Mol. Biol. 1996, 255, 714–725. [Google Scholar] [CrossRef] [PubMed]
- Washio, K.; Ishikawa, K. Structure and expression during the germination of rice seeds of the gene for a carboxypeptidase. Plant Mol. Biol. 1992, 19, 631–640. [Google Scholar] [CrossRef]
- Washio, K.; Ishikawa, K. Cloning and sequencing of the gene for type I carboxypeptidase in rice. Biochim. Biophys. Acta 1994, 1199, 311–314. [Google Scholar] [CrossRef]
- Washio, K.; Ishikawa, K. Organ-specific and hormone-dependent expression of genes for serine carboxypeptidases during development and following germination of rice grains. Plant Physiol. 1994, 105, 1275–1280. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Tang, L.; Qiu, J.; Zhang, W.; Wang, Y.; Tong, X.; Wei, X.; Hou, Y.; Zhang, J. Serine carboxypeptidase 46 Regulates Grain Filling and Seed Germination in Rice (Oryza sativa L.). PLoS ONE 2016, 11, e0159737. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Wang, X.; Zhang, H.; Yang, Y.; Ge, X.; Song, F. A rice serine carboxypeptidase-like gene OsBISCPL1 is involved in regulation of defense responses against biotic and oxidative stress. Gene 2008, 420, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Li, A.X.; Steffens, J.C. An acyltransferase catalyzing the formation of diacylglucose is a serine carboxypeptidase-like protein. Proc. Natl. Acad. Sci. USA 2000, 97, 6902–6907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moura, D.S.; Bergey, D.R.; Ryan, C.A. Characterization and localization of a wound-inducible type I serine-carboxypeptidase from leaves of tomato plants (Lycopersicon esculentum Mill.). Planta 2001, 212, 222–230. [Google Scholar] [CrossRef] [PubMed]
- Mehta, R.A.; Mattoo, A.K. Isolation and identification of ripening-related tomato fruit carboxypeptidase. Plant Physiol 1996, 110, 875–882. [Google Scholar] [CrossRef] [Green Version]
- Bienert, M.D.; Delannoy, M.; Navarre, C.; Boutry, M. NtSCP1 from tobacco is an extracellular serine carboxypeptidase III that has an impact on cell elongation. Plant Physiol 2012, 158, 1220–1229. [Google Scholar] [CrossRef] [Green Version]
- Cercos, M.; Urbez, C.; Carbonell, J. A serine carboxypeptidase gene (PsCP), expressed in early steps of reproductive and vegetative development in Pisum sativum, is induced by gibberellins. Plant Mol. Biol. 2003, 51, 165–174. [Google Scholar] [CrossRef]
- Mugford, S.T.; Qi, X.; Bakht, S.; Hill, L.; Wegel, E.; Hughes, R.K.; Papadopoulou, K.; Melton, R.; Philo, M.; Sainsbury, F.; et al. A serine carboxypeptidase-like acyltransferase is required for synthesis of antimicrobial compounds and disease Resistance in oats. Plant Cell 2009, 21, 2473–2484. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Lease, K.A.; Tax, F.E.; Walker, J.C. BRS1, a serine carboxypeptidase, regulates BRI1 signaling in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2001, 98, 5916–5921. [Google Scholar] [CrossRef] [Green Version]
- Shirley, A.M.; McMichael, C.M.; Chapple, C. The sng2 mutant of Arabidopsis is defective in the gene encoding the serine carboxypeptidase-like protein sinapoylglucose: Choline sinapoyltransferase. Plant J. 2001, 28, 83–94. [Google Scholar] [CrossRef]
- Shirley, A.M.; Chapple, C. Biochemical characterization of sinapoylglucose: Choline sinapoyltransferase, a serine carboxypeptidase-like protein that functions as an acyltransferase in plant secondary metabolism. J. Biol. Chem. 2003, 278, 19870–19877. [Google Scholar] [CrossRef] [Green Version]
- Parzych, K.R.; Ariosa, A.; Mari, M.; Klionsky, D.J. A newly characterized vacuolar serine carboxypeptidase, Atg42/Ybr139w, is required for normal vacuole function and the terminal steps of autophagy in the yeast Saccharomyces cerevisiae. Mol. Biol. Cell 2018, 29, 1089–1099. [Google Scholar] [CrossRef] [PubMed]
- Mugford, S.T.; Osbourn, A. Evolution of serine carboxypeptidase-like acyltransferases in the monocots. Plant Signal. Behav. 2010, 5, 193–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, A.; Li, J. Arabidopsis BRS1 is a secreted and active serine carboxypeptidase. J. Biol Chem. 2005, 280, 35554–35561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehfeldt, C.; Shirley, A.M.; Meyer, K.; Ruegger, M.O.; Cusumano, J.C.; Viitanen, P.V.; Strack, D.; Chapple, C. Cloning of the SNG1 gene of Arabidopsis reveals a role for a serine carboxypeptidase-like protein as an acyltransferase in secondary metabolism. Plant Cell 2000, 12, 1295–1306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciarkowska, A.; Ostrowski, M.; Jakubowska, A. A serine carboxypeptidase-like acyltransferase catalyzes synthesis of indole-3-acetic (IAA) ester conjugate in rice (Oryza sativa). Plant Phys. Biochem. 2018, 125, 126–135. [Google Scholar] [CrossRef]
- Fraser, C.M.; Thompson, M.G.; Shirley, A.M.; Ralph, J.; Schoenherr, J.A.; Sinlapadech, T.; Hall, M.C.; Chapple, C. Related Arabidopsis serine carboxypeptidase—Like sinapoylglucose acyltransferases display distinct but overlapping substrate Specificities. Plant Physiol. 2007, 144, 1986–1999. [Google Scholar] [CrossRef] [Green Version]
- Stehle, F.; Brandt, W.; Schmidt, J.; Milkowski, C.; Strack, D. Activities of Arabidopsis sinapoylglucose: Malate sinapoyltransferase shed light on functional diversification of serine carboxypeptidase-like acyltransferases. Phytochemistry 2008, 69, 1826–1831. [Google Scholar] [CrossRef]
- Stehle, F.; Stubbs, M.T.; Strack, D.; Milkowski, C. Heterologous expression of a serine carboxypeptidase-like acyltransferase and characterization of the kinetic mechanism. FEBS J. 2008, 275, 775–787. [Google Scholar] [CrossRef]
- Dominguez, F.; Cejudo, F.J. Patterns of starchy endosperm acidification and protease gene expression in wheat grains following germination. Plant Physiol. 1999, 119, 81–88. [Google Scholar] [CrossRef] [Green Version]
- Li, J.W.; Wang, X.M. Phospholipase D and phosphatidic acid in plant immunity. Plant Sci. 2019, 279, 45–50. [Google Scholar] [CrossRef]
- Testerink, C.; Larsen, P.B.; van der Does, D.; van Himbergen, J.A.; Munnik, T. Phosphatidic acid binds to and inhibits the activity of Arabidopsis CTR1. J. Exp. Bot. 2007, 58, 3905–3914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Lin, F.; Mao, T.; Nie, J.; Yan, M.; Yuan, M.; Zhang, W. Phosphatidic acid regulates microtubule organization by interacting with MAP65-1 in response to salt stress in Arabidopsis. Plant Cell 2012, 24, 4555–4576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, X.; Liu, Q.; Qiu, Y.; Fan, X.; Han, Q.; Liu, Y.; Zhang, L.; Xue, C. Identification of a novel phospholipase D with high transphosphatidylation activity and its application in synthesis of phosphatidylserine and DHA-phosphatidylserine. J. Biotechnol. 2017, 249, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Roy Choudhury, S.; Pandey, S. Phosphatidic acid binding inhibits RGS1 activity to affect specific signaling pathways in Arabidopsis. Plant J. 2017, 90, 466–477. [Google Scholar] [CrossRef] [Green Version]
- Munnik, T.; Arisz, S.A.; De Vrije, T.; Musgrave, A. G protein activation stimulates phospholipase D signaling in plant. Plant Cell 1995, 7, 2197–2210. [Google Scholar] [CrossRef]
- Siloto, R.M.; Findlay, K.; Lopez-Villalobos, A.; Yeung, E.C.; Nykiforuk, C.L.; Moloney, M.M. The accumulation of oleosins determines the size of seed oilbodies in Arabidopsis. Plant Cell 2006, 18, 1961–1974. [Google Scholar] [CrossRef] [Green Version]
- Welti, R.; Li, W.; Li, M.; Sang, Y.; Biesiada, H.; Zhou, H.E.; Rajashekar, C.B.; Williams, T.D.; Wang, X. Profiling membrane lipids in plant stress responses. Role of phospholipase D alpha in freezing-induced lipid changes in Arabidopsis. J. Biol. Chem. 2002, 277, 31994–32002. [Google Scholar] [CrossRef] [Green Version]
- Dormann, P.; Benning, C. Galactolipids rule in seed plants. Trends Plant Sci. 2002, 7, 112–118. [Google Scholar] [CrossRef]
- Wang, G.; Ryu, S.; Wang, X. Plant phospholipases: An overview. Methods Mol. Biol. 2012, 861, 123–137. [Google Scholar]
- Guo, L.; Mishra, G.; Taylor, K.; Wang, X. Phosphatidic acid binds and stimulates Arabidopsis sphingosine kinases. J. Biol. Chem. 2011, 286, 13336–13345. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Qu, Y.; Wang, Q.; Song, P.; Wang, P.; Jia, Q.; Guo, J. Arabidopsis phospholipase D alpha 1-derived phosphatidic acid regulates microtubule organization and cell development under microtubule-interacting drugs treatment. J. Plant Res. 2017, 130, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.C.; Guo, L.; Wang, X. Phosphatidic acid binds to cytosolic glyceraldehyde-3-phosphate dehydrogenase and promotes its cleavage in Arabidopsis. J. Biol. Chem. 2013, 288, 11834–11844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Streb, J.W.; Maltby, K.M.; Kitchen, C.M.; Miano, J.M. Cloning of a novel retinoid-inducible serine carboxypeptidase from vascular smooth muscle cells. J. Biol. Chem. 2001, 276, 34175–34181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Granat, S.J.; Wilson, K.A.; Tan-Wilson, A.L. New serine carboxypeptidase in mung bean seedling cotyledons. J. Plant Physiol. 2003, 160, 1263–1266. [Google Scholar] [CrossRef]
- Devaiah, S.P.; Roth, M.R.; Baughman, E.; Li, M.; Tamura, P.; Jeannotte, R.; Welti, R.; Wang, X. Quantitative profiling of polar glycerolipid species from organs of wild-type Arabidopsis and a PHOSPHOLIPASE Dα1 knockout mutant. Phytochemistry 2006, 67, 1907–1924. [Google Scholar] [CrossRef]
- Gut, H.; Matile, P. Breakdown of galactolipids in senescent barley leaves. Botanica Acta 1989, 102, 31–36. [Google Scholar] [CrossRef]
- Yang, S.F.; Freer, S.; Benson, A.A. Transphosphatidylation by phospholipase D. J. Biol. Chem. 1967, 242, 477–484. [Google Scholar]
- Baumert, A.; Milkowski, C.; Schmidt, J.; Nimtz, M.; Wray, V.; Strack, D. Formation of a complex pattern of sinapate esters in Brassica napus seeds, catalyzed by enzymes of a serine carboxypeptidase-like acyltransferase family? Phytochemistry 2005, 66, 1334–1345. [Google Scholar] [CrossRef]
- Ciarkowska, A.; Ostrowski, M.; Jakubowska, A. Abiotic stress and phytohormones affect enzymic activity of 1-O-(indole-3-acetyl)-β-d-glucose: Myo-inositol indoleacetyl transferase from rice (Oryza sativa). J. Plant Phys. 2016, 205, 93–96. [Google Scholar] [CrossRef]
- Gardiner, J.; Collings, D.A.; Harper, J.D.; Marc, J. The effects of the phospholipase D-antagonist 1-butanol on seedling development and microtubule organisation in Arabidopsis. Plant Cell Physiol. 2003, 44, 687–696. [Google Scholar] [CrossRef] [Green Version]
- Motes, C.M.; Pechter, P.; Yoo, C.M.; Wang, Y.S.; Chapman, K.D.; Blancaflor, E.B. Differential effects of two phospholipase D inhibitors, 1-butanol and N-acylethanolamine, on in vivo cytoskeletal organization and Arabidopsis seedling growth. Protoplasma 2005, 226, 109–123. [Google Scholar] [CrossRef] [PubMed]
- Dhonukshe, P.; Laxalt, A.M.; Goedhart, J.; Gadella, T.W.J.; Munnik, T. Phospholipase D activation correlates with microtubule reorganization in living plant cells. Plant Cell 2003, 15, 2666–2679. [Google Scholar] [CrossRef] [Green Version]
- Bonfig, K.; Schreiber, U.; Gabler, A.; Roitsch, T.; Berger, S. Infection with virulent and avirulent P. syringae strains differentially affects photosynthesis and sink metabolism in Arabidopsis leaves. Planta 2006, 225, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Tao, F.; Li, W. Lipid profiling demonstrates that suppressing Arabidopsis phospholipase Dδ retards ABA-promoted leaf senescence by attenuating lipid degradation. PLoS ONE 2013, 8, e65687. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Li, W. Phospholipase D antagonist 1-butanol inhibited the mobilization of triacylglycerol during seed germination in Arabidopsis. Plant Divers. 2018, 40, 292–298. [Google Scholar] [CrossRef]
- Rawyler, A.; Pavelic, D.; Gianinazzi, C.; Oberson, J.; Braendle, R. Membrane lipid integrity relies on a threshold of ATP production rate in Potato cell cultures submitted to anoxia. Plant Physiol. 1999, 120, 293–300. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lipid Classes | Lipids/Dry Weight (nmol/mg) | Ratio | |||
|---|---|---|---|---|---|
| Col | bis4 | SOE-5 | bis4/Col | SOE-5/Col | |
| PG | 10.88 ± 0.45 b | 12.28 ± 0.39 a | 8.44 ± 0.79 c | 112.84 | 77.57 |
| PC | 35.14 ± 2.21 b | 38.70 ± 2.44 a | 31.43 ± 0.45 c | 110.13 | 89.43 |
| PE | 21.02 ± 0.95 b | 23.85 ± 1.33 a | 17.41 ± 1.72 c | 113.46 | 82.87 |
| PI | 5.44 ± 0.32 a | 5.79 ± 0.69 a | 5.53 ± 0.17 a | - | - |
| PA | 1.55 ± 0.25 a | 1.74 ± 0.34 a | 1.81 ± 0.15 a | - | - |
| PS | 0.88 ± 0.06 a | 1.02 ± 0.25 a | 0.80 ± 0.17 a | - | - |
| MGDG | 44.33 ± 2.11 b | 49.43 ± 2.76 a | 33.75 ± 4.54 c | 111.50 | 76.15 |
| DGDG | 11.42 ± 0.80 a | 12.09 ± 1.08 a | 9.20 ± 1.09 b | - | 80.58 |
| Total lipids/dry weight (nmol/mg) | |||||
| Total lipids | 127.26 ± 2.37 b | 146.89 ± 8.11 a | 108.97 ± 7.65 c | 115.43 | 83.02 |
| Lipid Classes | Lipid (mol % of total lipids) | ||
|---|---|---|---|
| Col | bis4 | SOE-5 | |
| PG | 8.41 ± 0.31 a | 8.61 ± 0.38 a | 7.85 ± 0.26 b |
| PC | 26.72 ± 0.74 a | 26.79 ± 0.99 a | 28.91 ± 2.12 a |
| PE | 16.01 ± 0.42 a | 14.47 ± 2.72 a | 15.98 ± 0.99 a |
| PI | 4.14 ± 0.13 b | 4.24 ± 0.34 ab | 5.01 ± 0.41 a |
| PA | 1.18 ± 0.19 b | 1.28 ± 0.23 b | 1.66 ± 0.11 a |
| PS | 0.67 ± 0.04 a | 0.74 ± 0.11 a | 0.73 ± 0.14 a |
| MGDG | 33.72 ± 0.36 a | 34.55 ± 1.25 a | 30.81 ± 2.14 b |
| DGDG | 8.70 ± 0.31 a | 8.88 ± 0.31 a | 8.42 ± 0.43 a |
| Lipid ratio | |||
| PC/PE | 1.67 ± 0.06 a | 1.93 ± 0.53 a | 1.82 ± 0.18 a |
| DGDG/MGDG | 0.26 ± 0.01 a | 0.26 ± 0.01 a | 0.27 ± 0.01 a |
| Galactolipids/Phospholipids | 0.74 ± 0.02 a | 0.77 ± 0.04 a | 0.65 ± 0.07 b |
| Lipid Class | DBI (double bond index) | ||
|---|---|---|---|
| Col | bis4 | SOE-5 | |
| DGDG | 6.48 ± 0.02 b | 6.51 ± 0.01 a | 6.39 ± 0.04 c |
| MGDG | 6.84 ± 0.01 a | 6.84 ± 0.01 a | 6.83 ± 0.01 a |
| PG | 2.98 ± 0.04 a | 2.97 ± 0.04 a | 2.94 ± 0.07 a |
| PC | 3.81 ± 0.02 a | 3.82 ± 0.02 a | 3.80 ± 0.01 a |
| PE | 3.43 ± 0.02 a | 3.44 ± 0.02 a | 3.43 ± 0.02 a |
| PI | 2.59 ± 0.01 a | 2.58 ± 0.01 a | 2.58 ± 0.01 a |
| PS | 2.70 ± 0.02 a | 2.69 ± 0.05 a | 2.75 ± 0.04 a |
| PA | 3.06 ± 0.02 a | 3.11 ± 0.06 a | 3.10 ± 0.09 a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, J.; Li, W.-q.; Jia, Y.-x. The Serine Carboxypeptidase-Like Gene SCPL41 Negatively Regulates Membrane Lipid Metabolism in Arabidopsis thaliana. Plants 2020, 9, 696. https://doi.org/10.3390/plants9060696
Chen J, Li W-q, Jia Y-x. The Serine Carboxypeptidase-Like Gene SCPL41 Negatively Regulates Membrane Lipid Metabolism in Arabidopsis thaliana. Plants. 2020; 9(6):696. https://doi.org/10.3390/plants9060696
Chicago/Turabian StyleChen, Juan, Wei-qi Li, and Yan-xia Jia. 2020. "The Serine Carboxypeptidase-Like Gene SCPL41 Negatively Regulates Membrane Lipid Metabolism in Arabidopsis thaliana" Plants 9, no. 6: 696. https://doi.org/10.3390/plants9060696
APA StyleChen, J., Li, W. -q., & Jia, Y. -x. (2020). The Serine Carboxypeptidase-Like Gene SCPL41 Negatively Regulates Membrane Lipid Metabolism in Arabidopsis thaliana. Plants, 9(6), 696. https://doi.org/10.3390/plants9060696