Τhe Nematicidal Potential of Bioactive Streptomyces Strains Isolated from Greek Rhizosphere Soils Tested on Arabidopsis Plants of Varying Susceptibility to Meloidogyne spp.

 ,
,  ,
,  , ,
, , 

Abstract
:1. Introduction
2. Results
2.1. Antimicrobial Activity
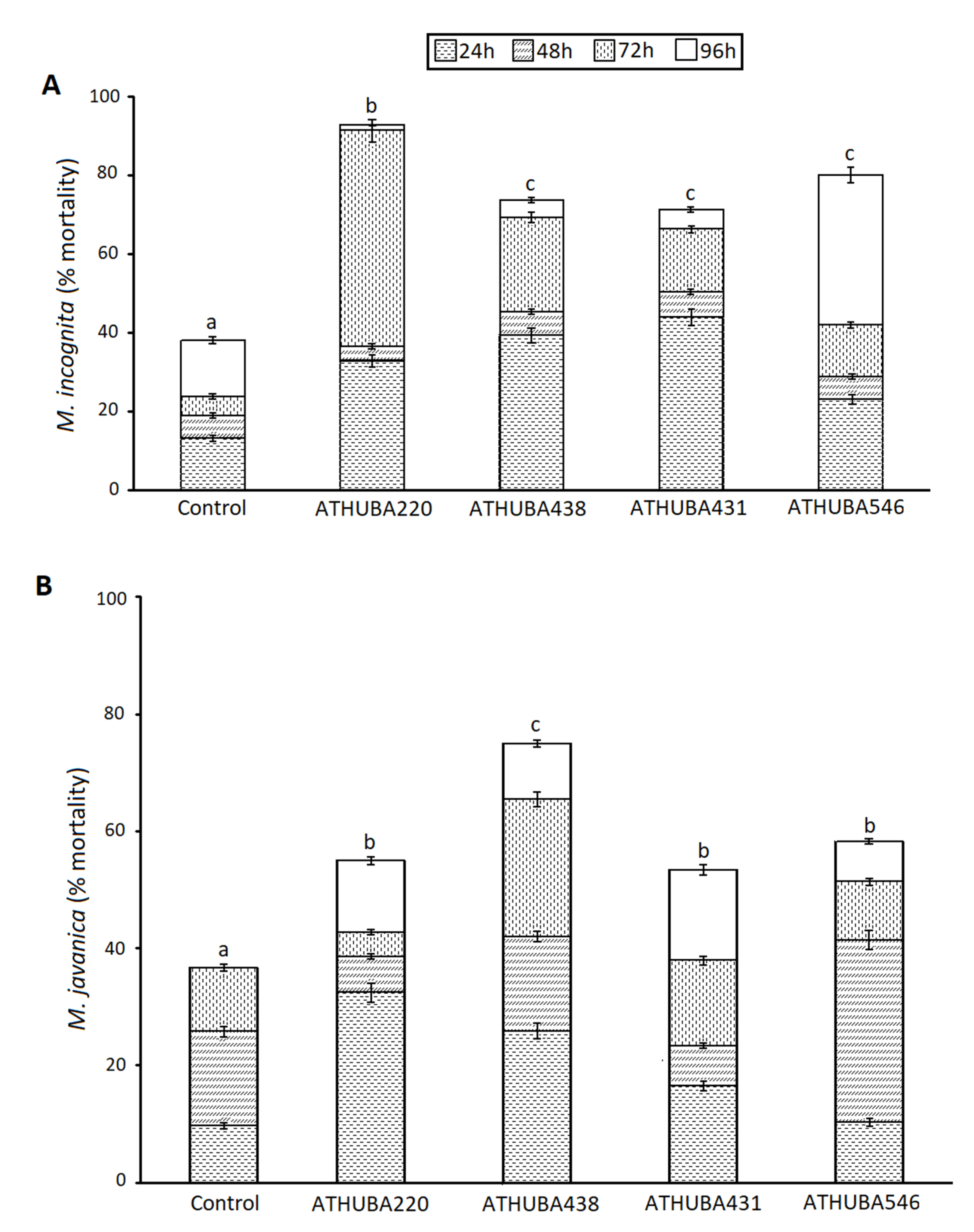
2.2. Nematicidal Activity of Streptomycetes Isolates
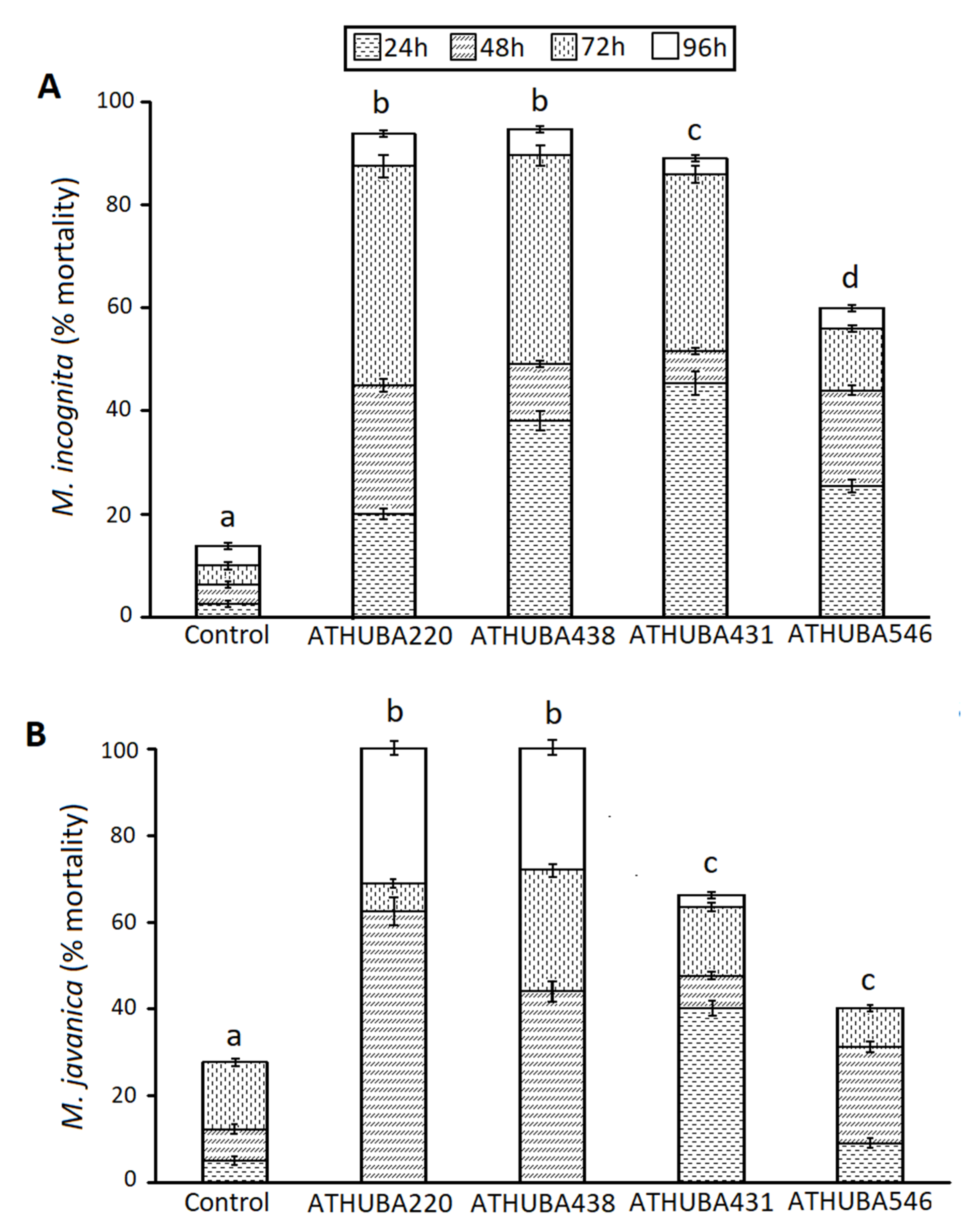
2.3. Nematode Biological Cycle Arrest
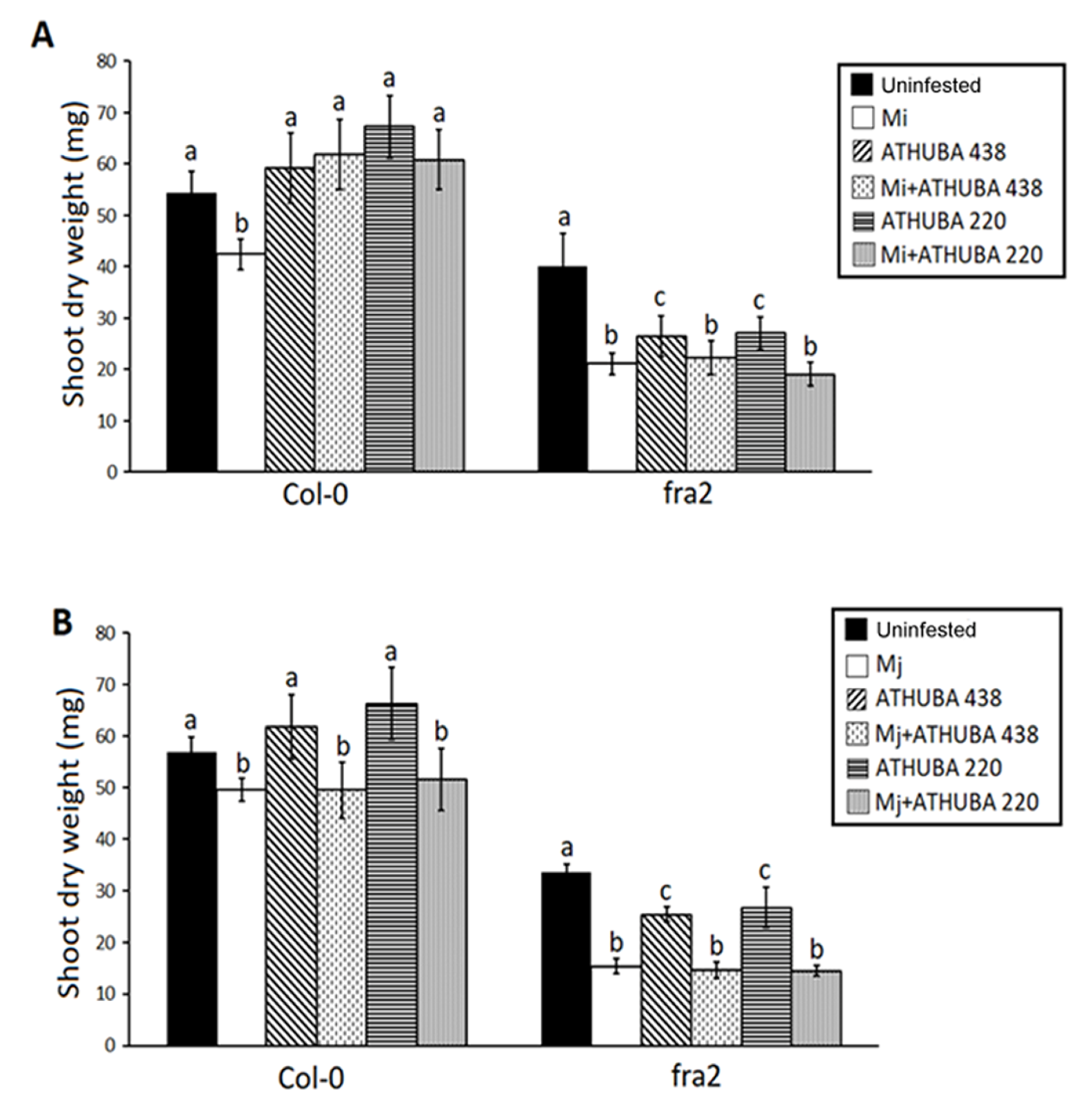
2.4. Growth Promotion
3. Discussion
4. Materials and Methods
4.1. Microbial Strain Isolation and Nematode Rearing
4.2. Streptomyces Growth Media and Growth Conditions
4.3. Preliminary Screening for Antimicrobial Activity
4.4. Screening of Streptomyces Isolates for Nematicidal Activity
4.5. DNA Amplification and Sequencing of 16S rDNA Gene
4.6. Biological Cycle Arrest
4.7. Shoot Dry Weight Recording
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rashad, F.M.; Fathy, H.M.; El-Zayat, A.S.; Elghonaimy, A.M. Isolation and characterization of multifunctional Streptomyces species with antimicrobial, nematicidal and phytohormone activities from marine environments in Egypt. Microbiol. Res. 2015, 175, 34–47. [Google Scholar] [CrossRef] [PubMed]
- Ruanpanun, P.; Chamswarng, C. Potential of actinomycetes isolated from earthworm castings in controlling root-knot nematode Meloidogyne incognita. J. Gen. Plant Pathol. 2016, 82, 43–50. [Google Scholar] [CrossRef]
- Alam, M.; Dahrni, S.; Khaliq, A.; Srivastava, S.K.; Samad, A.; Gupta, M.K. A promising strain of Streptomyces sp. with agricultural traits for growth-promotion and disease management. Indian J. Exp. Microbiol. 2012, 50, 559–568. [Google Scholar]
- Back, M.A.; Haydock, P.P.J.; Jenkinson, P. Disease complexes involving plant parasitic nematodes and soilborne pathogens. Plant Pathol. 2002, 51, 683–697. [Google Scholar] [CrossRef]
- Sun, M.H.; Gao, L.; Shi, Y.X.; Li, B.J.; Liu, X.Z. Fungi and actinomycetes associated with Meloidogyne spp. eggs and females in China and their biocontrol potential. J. Invertebr. Pathol. 2006, 93, 22–28. [Google Scholar] [CrossRef]
- Gallego, S.; Devers-Lamrani, M.; Rousidou, K.; Karpouzas, D.G.; Martin-Laurent, F. Assessment of the effects of oxamyl on the bacterial community of an agricultural soil exhibiting enhanced biodegradation. Sci. Total Environ. 2019, 651, 1189–1198. [Google Scholar] [CrossRef]
- Lagos, S.; Perruchon, C.; Katsoula, A.; Karpouzas, D.G. Isolation and characterization of soil bacteria able to rapidly degrade the organophosphorus nematicide fosthiazate. Lett. Appl. Microbiol. 2019, 68, 149–155. [Google Scholar] [CrossRef]
- Dong, L.Q.; Zhang, K.Q. Microbial control of plant-parasitic nematodes: A five-party interaction. Plant Soil 2006, 288, 31–45. [Google Scholar] [CrossRef]
- Laing, R.; Gillan, V.; Devaney, E. Ivermectin—Old Drug, New Tricks? Trends Parasitol. 2017, 33, 463–472. [Google Scholar] [CrossRef] [Green Version]
- Ruanpanun, P.; Laatsch, H.; Tangchitsomkid, N.; Lumyong, S. Nematicidal activity of fervenulin isolated from a nematicidal actinomycete, Streptomyces sp. CMU-MH021, on Meloidogyne incognita. World J. Microbiol. Biotechnol. 2011, 27, 1373–1380. [Google Scholar] [CrossRef] [Green Version]
- Vurukonda, S.S.P.; Giovanardi, D.; Stefani, E. Plant Growth Promoting and Biocontrol activity of Streptomyces spp. as endophytes. Int. J. Mol. Sci. 2018, 19, 952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jog, R.; Nareshkumar, G.; Rajkumar, S. Enhancing soil health and plant growth promotion by actinomycetes. In Plant Growth Promoting Actinobacteria; Gopalakrishnan, S., Sathya, A., Vijayabharathi, R., Eds.; Springer: Singapore, 2016; pp. 33–45. [Google Scholar]
- Dias, M.P.; Bastos, M.S.; Xavier, V.B.; Cassel, E.; Astarita, L.V.; Santarém, E.R. Plant growth and resistance promoted by Streptomyces spp. in tomato. Plant Physiol. Biochem. 2017, 118, 479–493. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.Y.; Li, Y.; Lai, H.; Guo, Q.; Xue, Q. Effects of two strains of Streptomyces on root-zone microbes and nematodes for biocontrol of root-knot nematode disease in tomato. Appl. Soil Ecol. 2017, 112, 34–41. [Google Scholar] [CrossRef]
- Nimnoi, P.; Pongsilp, N.; Ruanpanun, P. Monitoring the efficiency of Streptomyces galilaeus strain KPS-C004 against root knot disease and the promotion of plant growth in the plant-parasitic nematode infested soils. Biol. Control 2017, 114, 158–166. [Google Scholar] [CrossRef]
- Singh, A.; Sharma, P.; Kumari, A.; Kumar, R.; Pathak, D.V. Management of Root-Knot Nematode in Different Crops Using Microorganisms. In Plant Biotic Interactions; Springer: Cham, Switzerland, 2019; pp. 85–99. [Google Scholar]
- Subhashini, D.V.; Ramakrishnan, S.; Padmaja, K. Effect of culture filtrates of Streptomyces spp. on the mortality and hatching of root-knot nematode, Meloidogyne javanica. Indian J. Nematol. 2009, 39, 188–191. [Google Scholar]
- Caboni, P.; Ntalli, N.G. Botanical Nematicides, Recent Findings; ACS Symposium Series; ACS Publications: Washington, DC, USA, 2014; Volume 1172, pp. 145–157. [Google Scholar]
- Katsifas, E.A.; Giannoutsou, E.P.; Karagouni, A.D. Diversity of streptomycetes among specific Greek terrestrial ecosystems. Lett. Appl. Microbiol. 1999, 29, 48–51. [Google Scholar] [CrossRef]
- Meidani, C.; Ntalli, N.G.; Giannoutsou, E.; Adamakis, I.-D.S. Cell Wall Modifications in Giant Cells Induced by the Plant Parasitic Nematode Meloidogyne incognita in Wild-Type (Col-0) and the fra2 Arabidopsis thaliana Katanin Mutant. Int. J. Mol. Sci. 2019, 20, 5465. [Google Scholar] [CrossRef] [Green Version]
- Katsifas, E.A.; Koraki, T.G.; Karagouni, A.D. Determination of metabolic activity of Streptomycetes in soil microcosms. J. Appl. Microbiol. 2000, 89, 178–184. [Google Scholar] [CrossRef]
- Kanini, G.S.; Katsifas, E.A.; Savvides, A.L.; Hatzinikolaou, D.G.; Karagouni, A.D. Greek indigenous streptomycetes as biocontrol agents against the soil-borne fungal plant pathogen Rhizoctonia solani. J. Appl. Microbiol. 2013, 114, 1468–1479. [Google Scholar] [CrossRef]
- Kanini, G.S.; Katsifas, E.A.; Savvides, A.L.; Karagouni, A.D. Streptomyces rochei ACTA1551, an indigenous Greek isolate studied as a potential biocontrol agent against Fusarium oxysporum f. sp. lycopersici. BioMed Res. Int. 2013, 2013. [Google Scholar] [CrossRef] [Green Version]
- Alivizatos, A.S.; Pantazis, S. Preliminary Studies on Biological Control of Potato Common Scab Caused by Streptomyces sp. In Biological Control of Plant Diseases; Springer: Boston, MA, USA, 1992; pp. 85–93. [Google Scholar]
- Nikolakopoulou, T.L.; Egan, S.; van Overbeek, L.S.; Guillaume, G.; Heuer, H.; Wellington, E.M.; Karagouni, A.D. PCR detection of oxytetracycline resistance genes otr (A) and otr (B) in tetracycline-resistant Streptomycete isolates from diverse habitats. Curr. Microbiol. 2005, 51, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Wiggins, B.E.; Kinkel, L.L. Green manures and crop sequences influence alfalfa root rot and pathogen inhibitory activity among soil borne Streptomycetes. Plant Soil 2005, 268, 271–283. [Google Scholar] [CrossRef]
- Rey, T.; Dumas, B. Plenty is no plague: Streptomyces symbiosis with crops. Trends Plant Sci. 2017, 22, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Moore, J.M.; Bradshaw, E.; Seipke, R.F.; Hutchings, M.I.; McArthur, M. Use and discovery of chemical elicitors that stimulate biosynthetic gene clusters in Streptomyces bacteria. In Methods in Enzymology; Academic Press: New York, NY, USA, 2012; Volume 517, pp. 367–385. [Google Scholar]
- Schrey, S.D.; Tarkka, M.T. Friends and foes: Streptomycetes as modulators of plant disease and symbiosis. Antonie Leeuwenhoek 2008, 94, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Newitt, J.T.; Prudence, S.M.M.; Hutchings, M.I.; Worsley, S.F. Biocontrol of cereal crop diseases using Streptomycetes. Pathogens 2019, 8, 78. [Google Scholar] [CrossRef] [Green Version]
- Sawhney, R.; Webster, M.J. The influence of some metabolic inhibitors on the response of susceptible/resistant cultivars of tomato to Meloidogyne Incognita. Nematologica 1979, 25, 86–93. [Google Scholar] [CrossRef] [Green Version]
- Tarkka, M.; Hampp, R. Secondary metabolites of soil streptomycetes in biotic interactions. In Secondary Metabolites in Soil Ecology; Springer: Berlin, Germany, 2008; pp. 107–126. [Google Scholar]
- Jin, N.; Xue, H.; Li, W.-J.; Wang, X.-Y.; Liu, Q.; Liu, S.-S.; Liu, P.; Zhao, J.-L.; Jian, H. Field evaluation of Streptomyces rubrogriseus HDZ-9-47 for biocontrol of Meloidogyne incognita on tomato. J. Int. Agric. 2017, 16, 1347–1357. [Google Scholar] [CrossRef]
- Oplos, C.; Eloh, K.; Spiroudi, U.-M.; Pierluigi, C.; Ntalli, N. Nematicidal weeds, Solanum nigrum and Datura stramonium. J. Nematol. 2018, 50, 317–328. [Google Scholar] [CrossRef] [Green Version]
- Ntalli, N.; Oplos, C.; Michailidis, M.; Thanasenaris, A.; Kontea, D.; Caboni, P.; Tsiropoulos, N.G.; Menkissoglu-Spiroudi, U.; Adamski, Z. Strong synergistic activity and egg hatch inhibition by (E,E)-2,4-decadienal and (E)-2-decenal in Meloidogyne species. J. Pest Sci. 2016, 89, 565–579. [Google Scholar] [CrossRef]
- Olanrewaju, O.S.; Babalola, O.O. Streptomyces: Implications and interactions in plant growth promotion. Appl. Microbiol. Biotechnol. 2019, 103, 1179–1188. [Google Scholar] [CrossRef] [Green Version]
- Gupta, R.; Singh, A.; Srivastava, M.; Shanker, K.; Pandey, R. Plant-microbe interactions endorse growth by uplifting microbial community structure of Bacopa monnieri rhizosphere under nematode stress. Microbiol. Res. 2019, 218, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Inbar, E.; Green, S.J.; Hadar, Y.; Minz, D. Competing factors of compost concentration and proximity to root affect the distribution of Streptomycetes. Microb. Ecol. 2005, 50, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Abd-Alla, M.H.; El-Sayed, E.A.; Rasmey, A.M. Indole-3-acetic acid (IAA) production by Streptomyces atrovirens isolated from rhizospheric soil in Egypt. J. Biol. Earth Sci. 2013, 3, 182–193. [Google Scholar]
- Crawford, D.L.; Kowalski, M.; Roberts, M.A.; Merrell, G.; Deobald, L.A. Discovery, development, and commercialization of a microbial antifungal biocontrol agent, Streptomyces lydicus WYEC108: History of a decade long endeavor. Soc. Ind. Microbiol. News 2005, 55, 88–95. [Google Scholar]
- Kaur, T.; Jasrotia, S.; Ohri, P.; Manhas, R.K. Evaluation of in vitro and in vivo nematicidal potential of a multifunctional streptomycete, Streptomyces hydrogenans strain DH16 against Meloidogyne incognita. Microbiol. Res. 2016, 192, 247–252. [Google Scholar] [CrossRef]
- Bischoff, V.; Cookson, S.J.; Wu, S.; Scheible, W.R. Thaxtomin A affects CESA-complex density, expression of cell wall genes, cell wall composition, and causes ectopic lignification in Arabidopsis thaliana seedlings. J. Exp. Bot. 2009, 60, 955–965. [Google Scholar] [CrossRef] [Green Version]
- Burk, D.H.; Liu, B.; Zhong, R.; Morrison, W.H.; Ye, Z.H. A Katanin-like protein regulates normal cell wall biosynthesis and cell elongation. Plant Cell 2001, 13, 807–827. [Google Scholar] [CrossRef]
- Kabaluk, J.T.; Svircev, A.M.; Goettel, M.S.; Woo, S.G. The Use and Regulation of Microbial Pesticides in Representative Jurisdiction Worldwide; IOBC Global: Hong Kong, China, 2010; p. 99. [Google Scholar]
- Zade NS, E.; Sadeghi, A.; Moradi, P. Streptomyces strains alleviate water stress and increase peppermint (Mentha piperita) yield and essential oils. Plant Soil 2019, 434, 441–452. [Google Scholar] [CrossRef]
- Akbari, A.; Gharanjik, S.; Koobaz, P.; Sadeghi, A. Plant growth promoting Streptomyces strains are selectively interacting with the wheat cultivars especially in saline conditions. Heliyon 2020, 6, e03445. [Google Scholar] [CrossRef]
- Sadeghi, A.; Koobaz, P.; Azimi, H.; Karimi, E.; Akbari, A.R. Plant growth promotion and suppression of Phytophthora drechsleri damping-off in cucumber by cellulase-producing Streptomyces. BioControl 2017, 62, 805–819. [Google Scholar] [CrossRef]
- Hussey, R.S.; Barker, K.R. Comparison of methods of collecting inocula of Meloidogyne spp. including a new technique. Plant Dis. Rep. 1973, 57, 1025–1028. [Google Scholar]
- Atlas, R.M. Handbook of Microbiological Media; Parks, L.C., Ed.; CRC Press: Boca Raton, FL, USA, 2000. [Google Scholar]
- Raghava, R.K.V.; Mani, P.; Satyanarayana, B.; Raghava, R.T. Purification and structural elucidation of three bioactive compounds isolated from Streptomyces coelicoflavus BC 01 and their biological activity. 3 Biotech 2017, 7, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, H.; Hauser, M.; Naider, F.; Becker, J.M. Halo Assay for Toxic Peptides and Other Compounds in Microorganisms. Bio-Protocol 2016, 6, e2025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, R.A.; Li, J.; Allenby, N.E.; Errington, J.; Hayles, J.; Nurse, P. Screening and purification of natural products from actinomycetes that affect the cell shape of fission yeast. J. Cell Sci. 2017, 130, 3173–3185. [Google Scholar] [CrossRef] [Green Version]
- BLAST Sequence. Available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 30 May 2020).
- Wellington, E.M.; Cresswell, N.; Saunders, V.A. Growth and survival of streptomycete inoculants and extent of plasmid transfer in sterile and nonsterile soil. Appl. Environ. Microbiol. 1990, 56, 1413–1419. [Google Scholar] [CrossRef] [Green Version]
- Adamakis, I.D.S.; Panteris, E.; Eleftheriou, E.P. Tungsten disrupts root growth in Arabidopsis thaliana by PIN targeting. J. Plant Physiol. 2014, 171, 1174–1187. [Google Scholar] [CrossRef]
- Bybd, D.W., Jr.; Kirkpatrick, T.; Barker, K. An improved technique for clearing and staining plant tissues for detection of nematodes. J. Nematol. 1983, 15, 142. [Google Scholar]
- Huang, P.; de-Bashan, L.; Crocker, T.; Kloepper, J.W.; Bashan, Y. Evidence that fresh weight measurement is imprecise for reporting the effect of plant growth-promoting (rhizo) bacteria on growth promotion of crop plants. Biol. Fertil. Soils 2017, 53, 199–208. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Streptomyces sp. | Aspergillus niger DSM 1957 | Aspergillus nidulans LA1 | Fusarium oxysporum DSM 62059 | Saccharomyces cerevisiae DSM 70449 | Cyberlinderasaturnus Minter ATHUM 2576 | Escherichia coli NEB DH 5a | Pseudomonas aeruginosa ATCC 15442 | Pseudomonas fluorescens DSM 50090 | Acinetobacter radioresistens DSM 6976 | Bacillus subtilis DSM 10 | Micrococcus luteus DSM 1790 | Lactobacillus fermentum ATCC 9338 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ATHUBa 127 | 4.8 ± 0.2 | 0 | 0 | 7.3 ± 0.5 | 12.2 ± 1.0 | 0 | 0 | 16.0 ± 1.1 | 2.3 ± 0.2 | 3.6 ± 0.4 | 13.0 ± 0.6 | 4.0 ± 0.3 |
| ATHUBa 220 | 6.2 ± 0.4 | 9.0 ± 0.9 | 12.2 ± 0.8 | 16.8 ± 1.3 | 13.7 ± 0.9 | 2.2 ± 0.2 | 3.2 ± 0.2 | 2.2 ± 0.2 | 12.3 ± 0.6 | 3.6 ± 0.3 | 2.2 ± 0.1 | 20.3 ± 1.8 |
| ATHUBa 309 | 2.0 ± 0.1 | 0 | 0 | 7.3 ± 0.5 | 23.0 ± 1.1 | 0 | 0 | 2.6 ± 0.1 | 0 | 0 | 4.0 ± 0.3 | 4.0 ± 0.4 |
| ATHUBa 371 | 0 | 0 | 2.2 ± 0.1 | 4.0 ± 0.2 | 0 | 2.3 ± 0.1 | 0 | 6.2 ± 0.5 | 12.3 ± 0.9 | 6.2 ± 0.4 | 2.3 ± 0.1 | 2.6 ± 0.2 |
| ATHUBa 393 | 0 | 0 | 2.6 ± 0.2 | 4.0 ± 0.3 | 0 | 4.0 ± 0.4 | 0 | 4.0 ± 0.4 | 6.3 ± 0.5 | 6.2 ± 0.4 | 2.6 ± 0.2 | 4.8 ± 0.3 |
| ATHUBa 405 | 0 | 0 | 3.6 ± 0.3 | 6.2 ± 0.4 | 0 | 4.0 ± 0.3 | 0 | 1.2 ± 0.1 | 5.8 ± 0.4 | 4.0 ± 0.4 | 6.2 ± 0.5 | 4.0 ± 0.2 |
| ATHUBa 421 | 0 | 0 | 2.9 ± 0.2 | 4.0 ± 0.4 | 0 | 2.6 ± 0.2 | 0 | 4.8 ± 0.2 | 2.6 ± 0.2 | 3.6 ± 0.3 | 0 | 2.6 ± 0.2 |
| ATHUBa 431 | 4.0 ± 0.4 | 2.6 ± 0.2 | 8.4 ± 0.6 | 9.0± 0.9 | 12.2 ± 0.7 | 6.2 ± 0.5 | 3.6 ± 0.3 | 12.2 ± 0.6 | 25.0 ± 1.7 | 9.0 ± 0.6 | 20.2 ± 1.4 | 6.2 ± 0.5 |
| ATHUBa 438 | 2.2 ± 0.1 | 1.9 ± 0.1 | 6.2 ± 0.6 | 6.8 ± 0.7 | 12.2 ± 0.8 | 7.8 ± 0.6 | 6.3 ± 0.4 | 10.9 ± 0.9 | 25.0 ± 1.5 | 5.3 ± 0.4 | 19.4 ± 1.3 | 6.2 ± 0.6 |
| ATHUBa 501 | 6.2 ± 0.4 | 4.0 ± 0.3 | 2.6 ± 0.0 | 1.4 ± 0.1 | 2.6 ± 0.2 | 0 | 0 | 2.3 ± 0.1 | 6.8 ± 0.5 | 5.3 ± 0.4 | 9.0 ± 0.5 | 16.0 ± 1.1 |
| ATHUBa 526 | 2.2 ± 0.2 | 2.2 ± 0.1 | 2.0 ± 0.2 | 2.6 ± 0.2 | 1.7 ± 0.2 | 0 | 0 | 2.0 ± 0.1 | 4.0 ± 0.4 | 0 | 1.2 ± 0.1 | 0 |
| ATHUBa 527 | 2.2 ± 0.2 | 2.2 ± 0.2 | 1.7 ± 0.2 | 2.9 ± 0.2 | 3.2 ± 0.3 | 0 | 0 | 2.6 ± 0.2 | 2.9 ± 0.3 | 0 | 3.2 ± 0.3 | 0 |
| ATHUBa 531 | 3.6 ± 0.4 | 2.3 ± 0.2 | 1.4 ± 0.1 | 2.9 ± 0.2 | 1.9 ± 0.2 | 0 | 0 | 4.0 ± 0.2 | 4.0 ± 0.3 | 1.4 ± 0.1 | 2.6 ± 0.2 | 0 |
| ATHUBa 546 | 0 | 12.2 ± 1.0 | 0 | 4.0 ± 0.2 | 0 | 0 | 0 | 4.0 ± 0.3 | 0 | 26.0 ± 1.6 | 9.0 ± 0.5 | 49.0 ± 3.2 |
| ATHUBa 576 | 0 | 4.0 ± 0.2 | 0 | 2.0 ± 0.2 | 2.6 ± 0.2 | 0 | 0 | 5.8 ± 0.6 | 2.6 ± 0.2 | 2.6 ± 0.2 | 6.2 ± 0.6 | 2.3 ± 0.1 |
| ATHUBa 133 | 3.6 ± 0.2 | 0 | 0 | 7.3 ± 0.7 | 12.2 ± 0.8 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| ATHUBa 178 | 0 | 0 | 0 | 9.0 ± 0.9 | 12.9 ± 0.9 | 0 | 0 | 0 | 0 | 0 | 1.4 ± 0.1 | 0 |
| ATHUBa 181 | 0 | 0 | 2.0 ± 0.1 | 9.0 ± 0.8 | 12.2 ± 0.7 | 0 | 0 | 0 | 0 | 0 | 1.0 ± 0.2 | 0 |
| ATHUBa 183 | 2.2 ± 0.1 | 0 | 3.6 ± 0.4 | 10.2 ± 1.0 | 12.2 ± 0.6 | 0 | 0 | 0 | 0 | 0 | 1.2 ± 0.1 | 0 |
| ATHUBa 187 | 1.4 ± 0.1 | 0 | 1.9 ± 0.1 | 9.0 ± 0.6 | 12.2 ± 1.1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| ATHUBa 200 | 6.8 ± 0.5 | 0 | 0 | 16.0 ± 0.9 | 16.0 ± 1.1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| ATHUBa 201 | 6.2± 0.5 | 0 | 0 | 15.2 ± 1.1 | 12.3 ± 0.7 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| ATHUBa 262 | 1.7 ± 0.1 | 0 | 0 | 6.8 ± 0.7 | 10.2 ± 1.0 | 0 | 0 | 0 | 0 | 0 | 0 | 1.4 ± 0.1 |
| ATHUBa 307 | 0 | 0 | 0 | 6.2 ± 0.5 | 12.2 ± 0.7 | 0 | 0 | 0 | 0 | 0 | 1.2 ± 0.1 | 0 |
| ATHUBa 309 | 1.9 ± 0.1 | 0 | 0 | 7.3 ± 0.7 | 23.0 ± 1.4 | 0 | 0 | 2.6 ± 0.1 | 0 | 0 | 4.0 ± 0.3 | 4.1 ± 0.2 |
| ATHUBa 358 | 0 | 0 | 4.0 ± 0.3 | 4.8 ± 0.3 | 9.6 ± 0.7 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Ref. | Type of Ecosystem | |
|---|---|---|
| S. colombiensisATHUBA 431 | [19] | Rhizosphere of Pinus brutia |
| S. colombiensisATHUBA 438 | [19] | Rhizosphere of Pinus brutia |
| S. youssoufensisATHUBA 546 | [Present work] | Rhizosphere of Olea europea |
| S. monomyciniATHUBA 220 | [19] | Rhizosphere of evergreen woody shrubs |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meidani, C.; Savvidis, A.; Lampropoulou, E.; Sagia, A.; Katsifas, E.; Monokrousos, N.; Hatzinikolaou, D.G.; Karagouni, A.D.; Giannoutsou, E.; Adamakis, I.-D.S.; et al. Τhe Nematicidal Potential of Bioactive Streptomyces Strains Isolated from Greek Rhizosphere Soils Tested on Arabidopsis Plants of Varying Susceptibility to Meloidogyne spp. Plants 2020, 9, 699. https://doi.org/10.3390/plants9060699
Meidani C, Savvidis A, Lampropoulou E, Sagia A, Katsifas E, Monokrousos N, Hatzinikolaou DG, Karagouni AD, Giannoutsou E, Adamakis I-DS, et al. Τhe Nematicidal Potential of Bioactive Streptomyces Strains Isolated from Greek Rhizosphere Soils Tested on Arabidopsis Plants of Varying Susceptibility to Meloidogyne spp. Plants. 2020; 9(6):699. https://doi.org/10.3390/plants9060699
Chicago/Turabian StyleMeidani, Christianna, Alexandros Savvidis, Evaggelia Lampropoulou, Aggeliki Sagia, Efstathios Katsifas, Nikolaos Monokrousos, Dimitris G. Hatzinikolaou, Amalia D. Karagouni, Eleni Giannoutsou, Ioannis-Dimosthenis S. Adamakis, and et al. 2020. "Τhe Nematicidal Potential of Bioactive Streptomyces Strains Isolated from Greek Rhizosphere Soils Tested on Arabidopsis Plants of Varying Susceptibility to Meloidogyne spp." Plants 9, no. 6: 699. https://doi.org/10.3390/plants9060699
APA StyleMeidani, C., Savvidis, A., Lampropoulou, E., Sagia, A., Katsifas, E., Monokrousos, N., Hatzinikolaou, D. G., Karagouni, A. D., Giannoutsou, E., Adamakis, I. -D. S., & Ntalli, N. G. (2020). Τhe Nematicidal Potential of Bioactive Streptomyces Strains Isolated from Greek Rhizosphere Soils Tested on Arabidopsis Plants of Varying Susceptibility to Meloidogyne spp. Plants, 9(6), 699. https://doi.org/10.3390/plants9060699