
Climate Variability Shifting Immigrated Rice Planthoppers in Taiwan

 ,
,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. Results
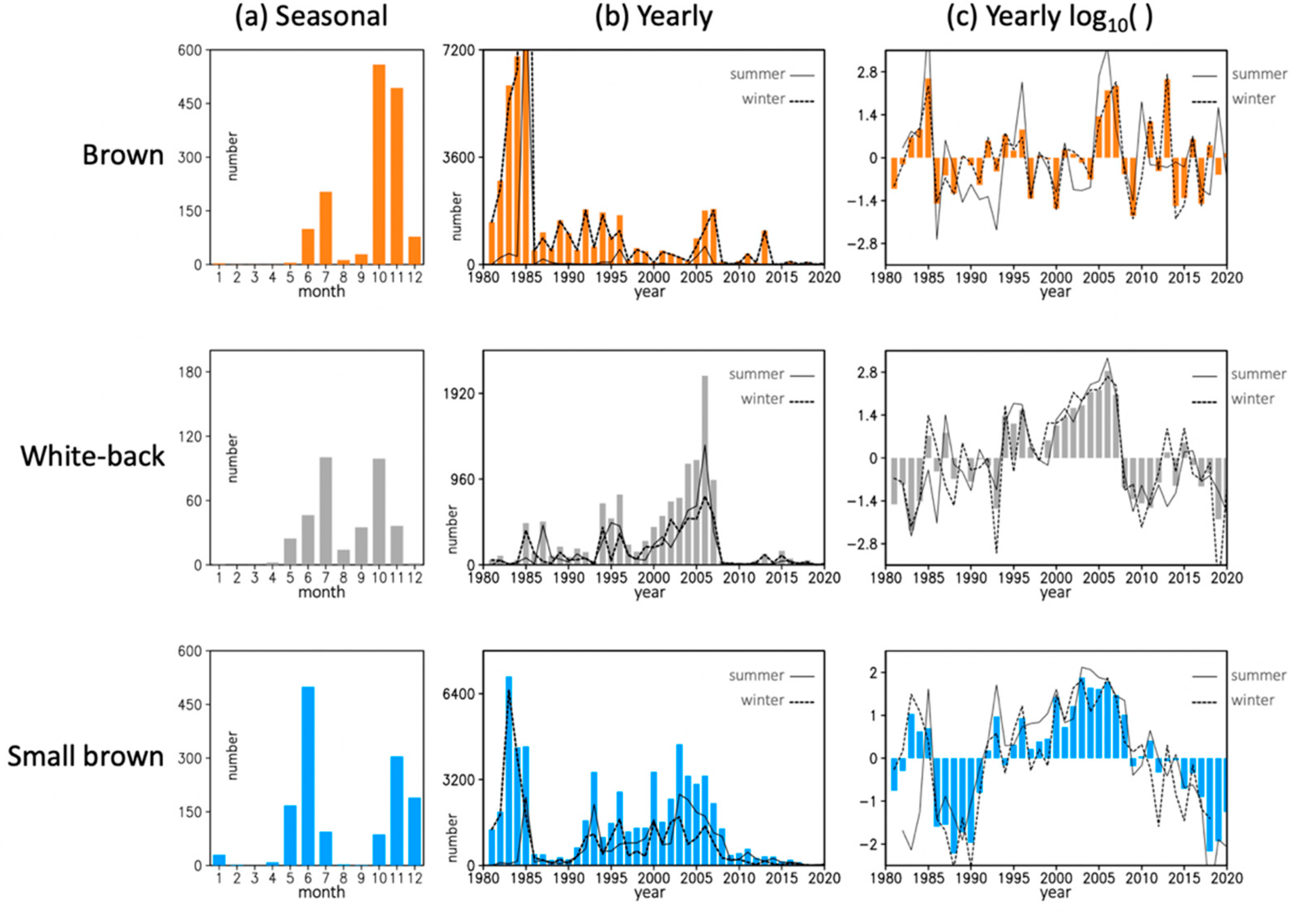
3.1. Pest Population Change
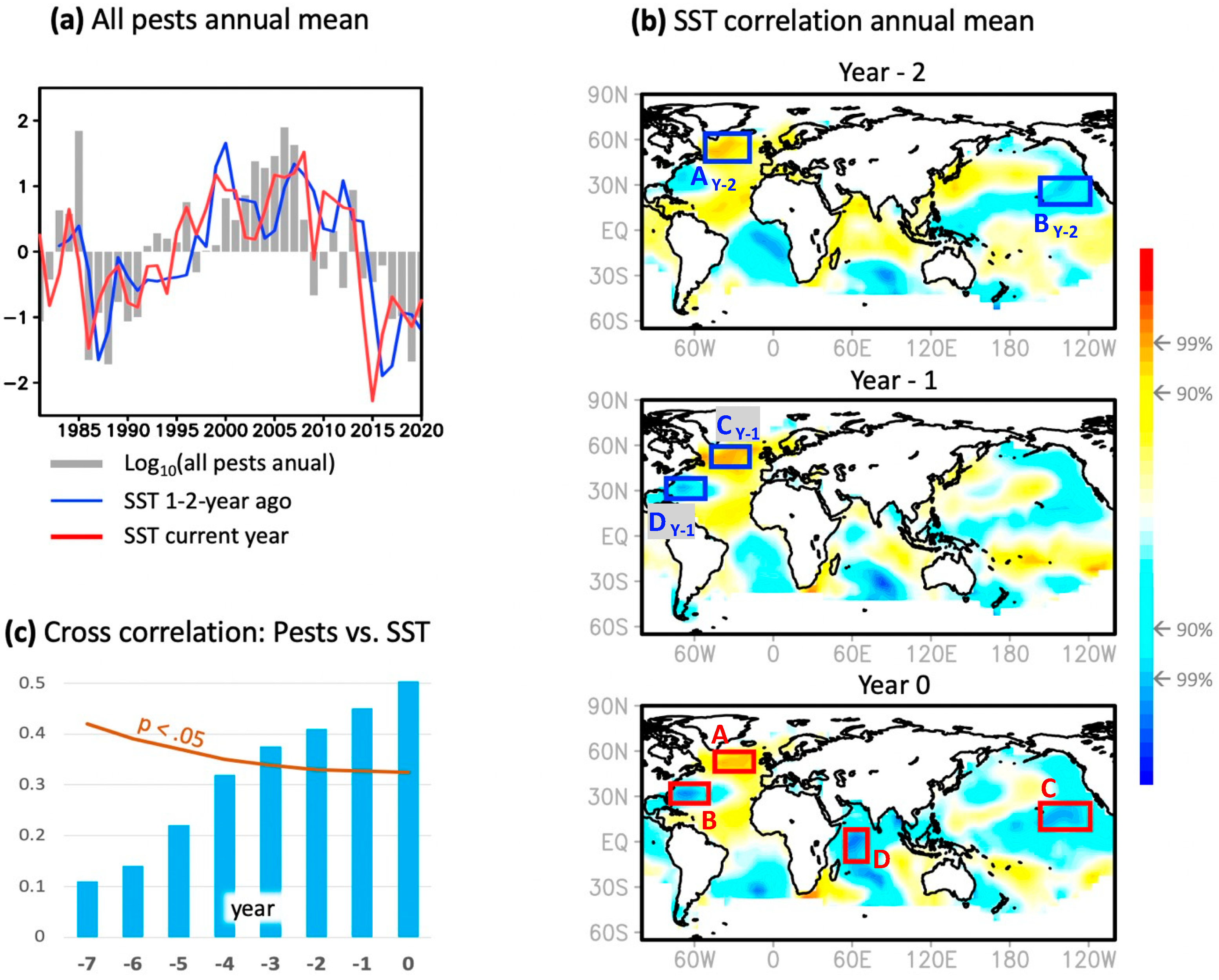
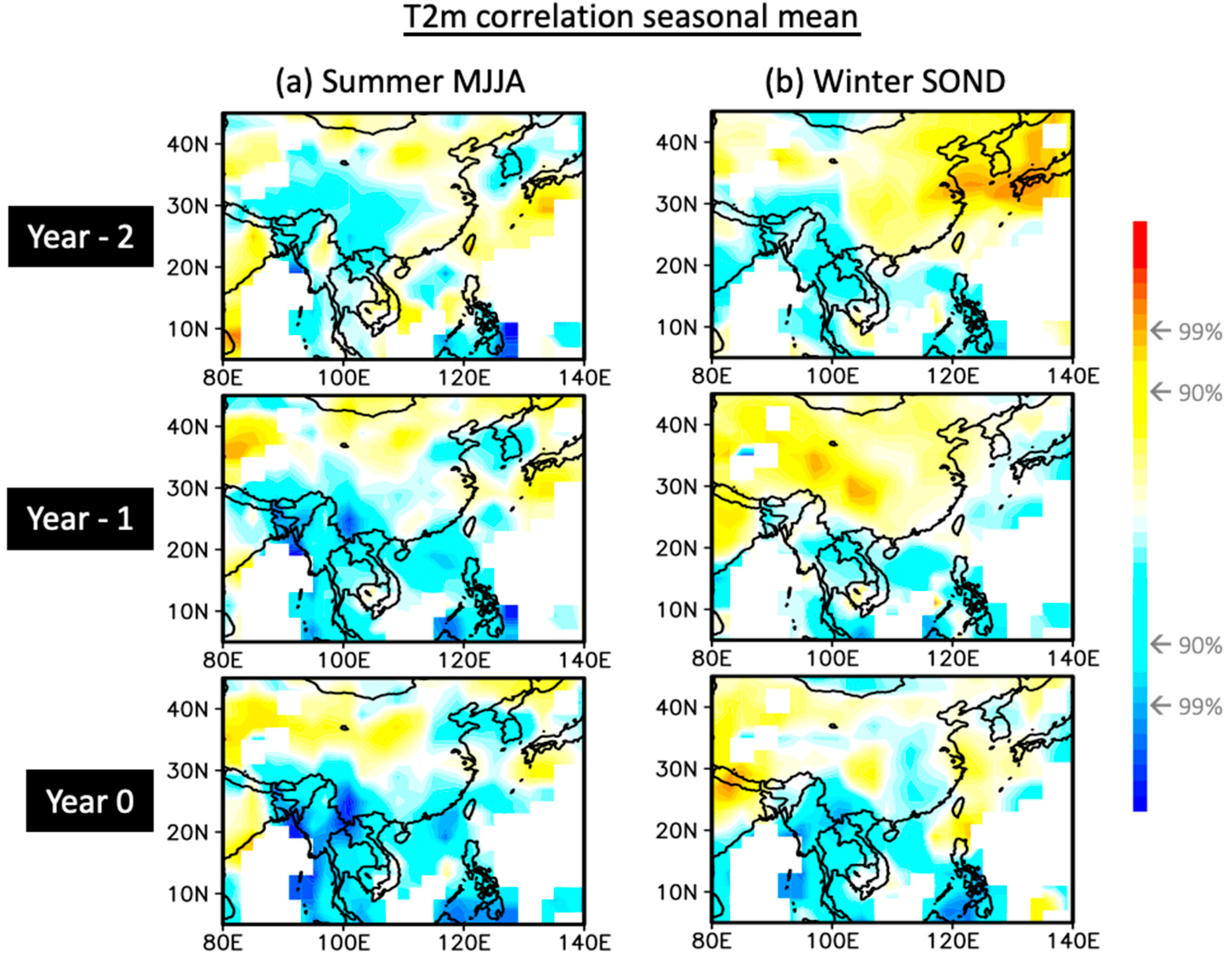
3.2. Climate Variability Connection
4. Discussion
4.1. Climate Dynamics
4.2. Implication on Insect Pests
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, C.C.; Cheng, C.H.; Chen, C.C.; Wang, S.S.; Chu, Y.I. Immigrations of planthoppers from oversea to Taiwan in 1987. Chin. J. Entomol. 1989, 9, 1–11. [Google Scholar]
- Cheng, C.H. Studies on population dynamics and forecasting of population abundance of brown planthopper, Nilaparvata lugens in Chia-nan area. Chin. J. Entomol. 1990, 10, 1–25. [Google Scholar]
- Huang, S.H.; Cheng, C.H.; Chen, C.N.; Wu, W.J.; Otuka, A. Estimating the immigration source of rice planthoppers, Nilaparvata lugens (Stål) and Sogatella furcifera (Horváth) (Homoptera: Delphacidae), in Taiwan. Appl. Entomol. Zool. 2010, 45, 521–531. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.H.; Cheng, C.H.; Chen, C.N.; Wu, W.J. The trend of occurrence and present status of brown planthopper and white-backed planthopper in Taiwan. In Proceedings of the International Workshop on Forecasting and Management of Rice Planthoppers in East Asia: Their Ecology and Genetics, Kumamoto, Japan, 4–5 December 2007; pp. 27–35. [Google Scholar]
- Otuka, A.; Huang, S.H.; Sanada-Morimura, S.; Matsumura, M. Migration analysis of Nilaparvata lugens (Hemiptera: Delphacidae) from the Philippines to Taiwan under typhoon-induced windy conditions. Appl. Entomol. Zool. 2012, 47, 263–271. [Google Scholar] [CrossRef]
- Yamamura, K.; Yokozawa, M.; Nishimori, M.; Ueda, Y.; Yokosuka, T. How to analyze long-term insect population dynamics under climate change: 50-year data of three insect pests in paddy fields. Popul. Ecol. 2006, 48, 31–48. [Google Scholar] [CrossRef]
- Cheng, C.H.; Lu, J.L. Detection of the transoceanic immigration of rice planthoppers, Nilaparvata lugens Stål and Sogatella furcifera Horváth to the southwestern Taiwan and their relative weather-conditions. Chin. J. Entomol. 1990, 10, 301–324. [Google Scholar]
- Sutherst, R.W.; Constable, F.; Finlay, K.J.; Harrington, R.; Luck, J.; Zalucki, M.P. Adapting to crop pest and pathogen risks under a changing climate. Wiley Interdiscip. Rev. Clim. Chang. 2011, 2, 220–237. [Google Scholar] [CrossRef]
- Nguyen, C.; Bahar, M.H.; Baker, G.; Andrew, N.R. Thermal tolerance limits of diamondback moth in ramping and plunging assays. PLoS ONE 2014, 9, e87535. [Google Scholar]
- Sharma, H.C.; Prabhakar, C. Impact of climate change on pest management and food security. In Integrated Pest Management: Current Concepts and Ecological Perspective; Abrol, D.P., Ed.; Academic Press: San Diego, CA, USA, 2014; pp. 23–36. [Google Scholar]
- Kaplan, A.; Cane, M.; Kushnir, Y.; Clement, A.; Blumenthal, M.; Rajagopalan, B. Analyses of global sea surface temperature 1856–1991. J. Geophys. Res. 1998, 103, 18567–18589. [Google Scholar] [CrossRef] [Green Version]
- Hersbach, H.; Bell, B. The ERA5 global reanalysis. Q. J. R. Meteorol. Soc. 2020, 146, 1999–2049. [Google Scholar] [CrossRef]
- Jones, P.D.; Moberg, A. Hemispheric and Largescale Surface Air Temperature Variations: An extensive Revision and an Update to 2001. J. Clim. 2003, 16, 206–223. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.H. The occurrence and control of the rice brown planthopper (Nilaparvata lugens Stål) in Taiwan. In Brown Planthopper; Indonesian Institute of Science: Bali, Indonesia, 1978; pp. 103–133. [Google Scholar]
- Newman, M.; Alexander, M.A.; Ault, T.R.; Cobb, K.M.; Deser, C.; Di Lorenzo, E.; Mantua, N.J.; Miller, A.J.; Minobe, S.; Nakamura, H.; et al. The Pacific decadal oscillation, revisited. J. Clim. 2016, 29, 4399–4427. [Google Scholar] [CrossRef] [Green Version]
- Knight, J.R.; Folland, C.K.; Scaife, A.A. Climate impacts of the Atlantic multidecadal oscillation. Geophys. Res. Lett. 2006, 33, 747. [Google Scholar] [CrossRef] [Green Version]
- Weisheimer, A.; Schaller, N.; O’Reilly, C.; MacLeod, D.A.; Palmer, T. Atmospheric seasonal forecasts of the twentieth century: Multi-decadal variability in predictive skill of the winter North Atlantic Oscillation (NAO) and their potential value for extreme event attribution. Q. J. R. Meteorol. Soc. 2017, 143, 917–926. [Google Scholar] [CrossRef] [PubMed]
- McCabe, G.J.; Palecki, M.A.; Betancourt, J.L. Pacific and Atlantic Ocean influences on multidecadal drought frequency in the United States. Proc. Natl. Acad. Sci. USA 2004, 101, 4136–4141. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.H.; Tsai, P.C.; Huang, W.R.; Wang, S.Y. Winter–summer contrast of the 1990s decadal change in relation to Afro–Asian monsoons. Clim. Dyn. 2022. [Google Scholar] [CrossRef]
- Kanoppaiah, V.; Sujayanad, G.K. Impact of climate change on population dynamics of insect pests. World J. Agric. Sci. 2012, 8, 240–246. [Google Scholar]
- Huang, W.-R.; Wang, S.-Y.; Guan, B. Decadal fluctuations in the western Pacific recorded by long precipitation records in Taiwan. Clim. Dyn. 2017, 50, 1597–1608. [Google Scholar] [CrossRef] [Green Version]
- Guan, B.T.; Wright, W.; Chung, C.-H.; Chang, S.-T. ENSO and PDO strongly influence Taiwan spruce height growth. For. Ecol. Manag. 2012, 267, 50–57. [Google Scholar] [CrossRef]
- Lo, T.T.; Hsu, H.H. The early 1950s regime shift in temperature in Taiwan and East Asia. Clim. Dyn. 2008, 31, 449–461. [Google Scholar] [CrossRef]
- Hong, C.-C.; Hsu, H.-H.; Chia, H.-H.; Wu, C.-Y. Decadal relationship between the North Atlantic Oscillation and cold surge frequency in Taiwan. Geophys. Res. Lett. 2008, 35, L24707. [Google Scholar] [CrossRef]
- Liu, C.H. Analysis on the major factors causing the eruptive occurrence of the brown planthopper in Taiwan. Chin. J. Entomol. 1988, 8, 119–130. [Google Scholar]
- Cheng, C.H. Studies on integrated control of brown planthopper, Nilaparvata lugens (Stål) in Taiwan. In Ecology and Control of the Brown Planthopper; Plant Protection Center: Taichung, Taiwan, 1984; pp. 149–167. [Google Scholar]
- Otuka, A.; Dudhia, J.; Watanabe, T.; Furuno, A. A new trajectory analysis method for migratory planthoppers, Sogatella furcifera (Horváth) (Homoptera: Delphacidae) and Nilaparvata lugens (Stål), using an advanced weather forecast model. Agric. For. Entomol. 2005, 7, 1–9. [Google Scholar] [CrossRef]
- Otuka, A.; Matsumura, M.; Watanabe, T.; Dihn, T.V. A migration analysis for rice planthoppers, Sogatella furcifera (Horváth) and Nilaparvata lugens (Stål) (Homoptera: Delphacidae), emigrating from northern Vietnam from April to May. Appl. Entomol. Zool. 2008, 43, 527–534. [Google Scholar] [CrossRef] [Green Version]
- Sogawa, K.; Watanabe, T.; Tsurumachi, M. Emigration areas and meteorological factors inducing overseas migration of the brown planthopper. Proc. Assoc. Pl. Prot. Kyushu 1988, 34, 79–82. [Google Scholar] [CrossRef]
- Sogawa, K. The monsoon-dependent migration of rice planthoppers in east Asia. In Migration and Management of Insect Pests of Rice in Monsoon Asia; China National Rice Research Institute: Hangzhou, China, 1997; pp. 217–230. [Google Scholar]
- Zanchettin, D.; Gaetan, C.; Arisido, W.M.; Modali, K.; Toniazzo, T.; Keenlyside, N.; Rubino, A. Structural decomposition of decadal climate prediction errors: A Bayesian approach. Sci. Rep. 2017, 7, 12862. [Google Scholar] [CrossRef] [Green Version]
- Stenseth, N.C. How to control pest species: Application of models from the theory of island biogeography in formulating pest control strategies. J. Appl. Ecol. 1981, 18, 773–794. [Google Scholar] [CrossRef]
- Horii, A.; McCue, P.; Shetty, K. Enhancement of seed vigour following insecticide and phenolic elicitor treatment. Biores. Tech. 2007, 98, 623–632. [Google Scholar] [CrossRef]
- Fand, B.B.; Kanble, A.L.; Kumar, M. Will climate change pose serious threat to crop pest management: A critical review. Int. J. Sci. Res. Publ. 2012, 2, 1–14. [Google Scholar]
- Ford, K.A.; Casida, J.E.; Chandranb, D. Neonicotinoid insecticides induce salicylate associated plant defense responses. Proc. Natl. Acad. Sci. USA 2010, 10, 17527–17532. [Google Scholar] [CrossRef] [Green Version]
- Kuang, Y.H.; Fang, Y.F.; Lin, S.C.; Tsai, S.F.; Yang, Z.W.; Li, C.P.; Huang, S.H.; Hechanova, S.L.; Jena, K.K.; Chuang, W.P. The impact of climate change on the resistance of rice near-isogenic lines with resistance genes against brown planthopper. Rice 2021, 14, 64. [Google Scholar] [CrossRef] [PubMed]
- Andrew, N.R.; Hill, S.J. Effect of climate change on insect pest management. In Environmental Pest Management: Challenges for Agronomists, Ecologists, Economists and Policymakers; Coll, M., Wajnberg, E., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2017; pp. 197–223. [Google Scholar]
- Sharma, H.C.; Srivastava, C.P.; Durairaj, C.; Gowda, C.L.L. Pest management in grain legumes and climate change. In Climate Change and Management of Cool Season Grain Legume Crops; Yadav, S.S., McNeil, D.L., Redden, R., Patil, S.A., Eds.; Springer Science and Business Media: Dordrecht, The Netherlands, 2010; pp. 115–140. [Google Scholar]
- Yadav, D.S.; Subhash, C.; Selvraj, K. Agroecological zoning of brown plant hopper, (Nilaparvata lugens (Stal.)) incidence on rice (Oryza sativa (L.)). J. Sci. Ind. Res. 2010, 69, 818–822. [Google Scholar]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, S.-H.; Lai, P.-Y.; Hwang, S.-Y.; Borhara, K.; Huang, W.-R.; Wang, S.-Y. Climate Variability Shifting Immigrated Rice Planthoppers in Taiwan. Climate 2022, 10, 71. https://doi.org/10.3390/cli10050071
Huang S-H, Lai P-Y, Hwang S-Y, Borhara K, Huang W-R, Wang S-Y. Climate Variability Shifting Immigrated Rice Planthoppers in Taiwan. Climate. 2022; 10(5):71. https://doi.org/10.3390/cli10050071
Chicago/Turabian StyleHuang, Shou-Horng, Pau-Yu Lai, Shaw-Yhi Hwang, Krishna Borhara, Wan-Ru Huang, and Shih-Yu Wang. 2022. "Climate Variability Shifting Immigrated Rice Planthoppers in Taiwan" Climate 10, no. 5: 71. https://doi.org/10.3390/cli10050071
APA StyleHuang, S. -H., Lai, P. -Y., Hwang, S. -Y., Borhara, K., Huang, W. -R., & Wang, S. -Y. (2022). Climate Variability Shifting Immigrated Rice Planthoppers in Taiwan. Climate, 10(5), 71. https://doi.org/10.3390/cli10050071