Mapping the Potential Global Range of the Brown Marmorated Stink Bug, Halyomorpha halys, with Particular Reference to New Zealand


Abstract
:1. Introduction
2. Materials and Methods
3. Results
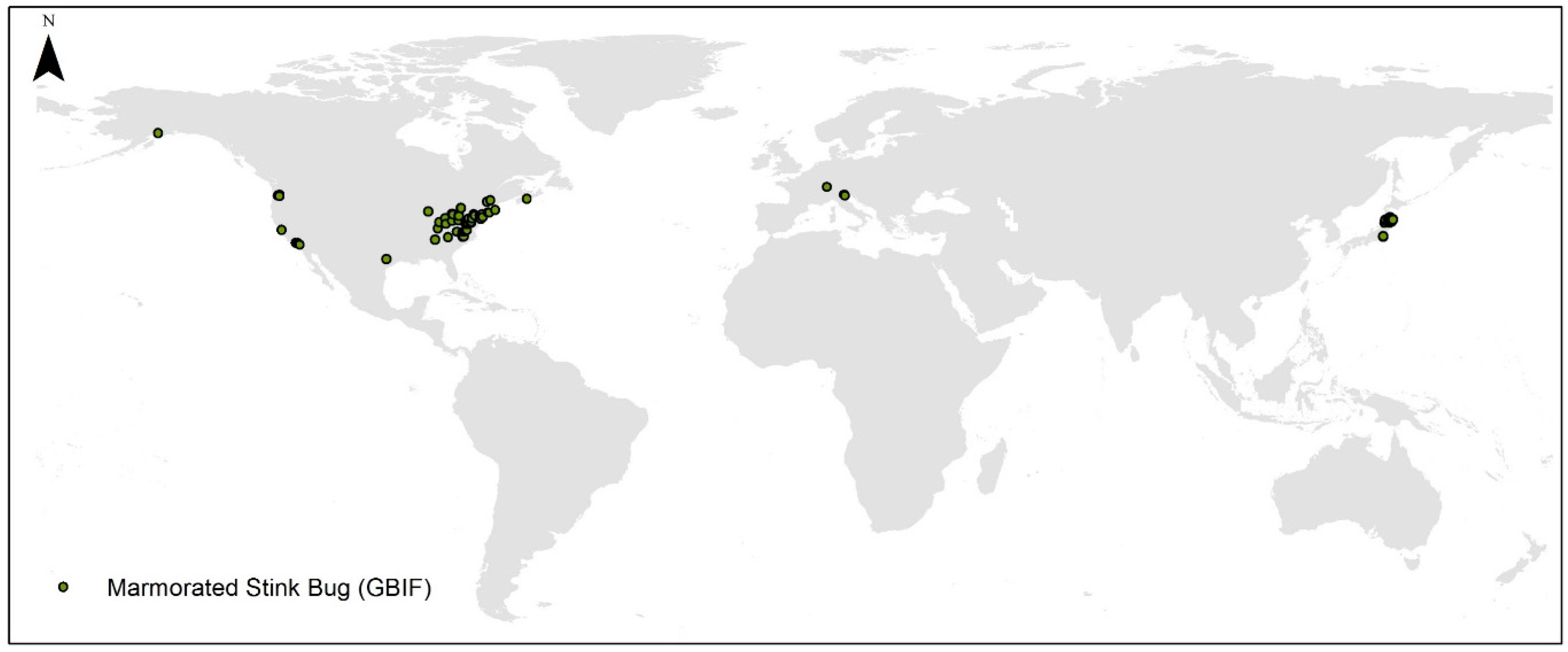
3.1. Occurrence Data
3.2. Model Evaluation
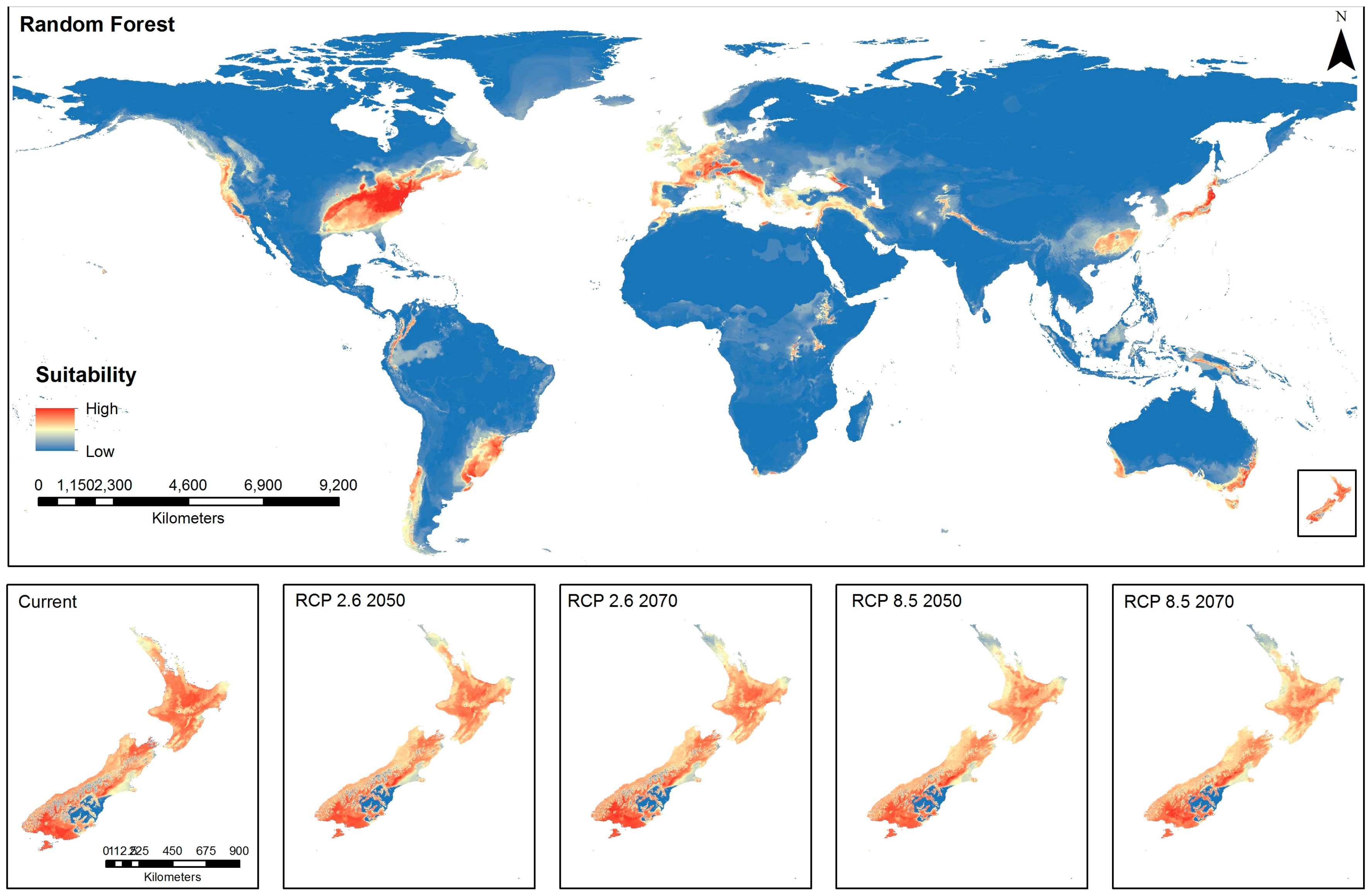
3.3. Spatial Distribution Using Random Forest (RF) Model
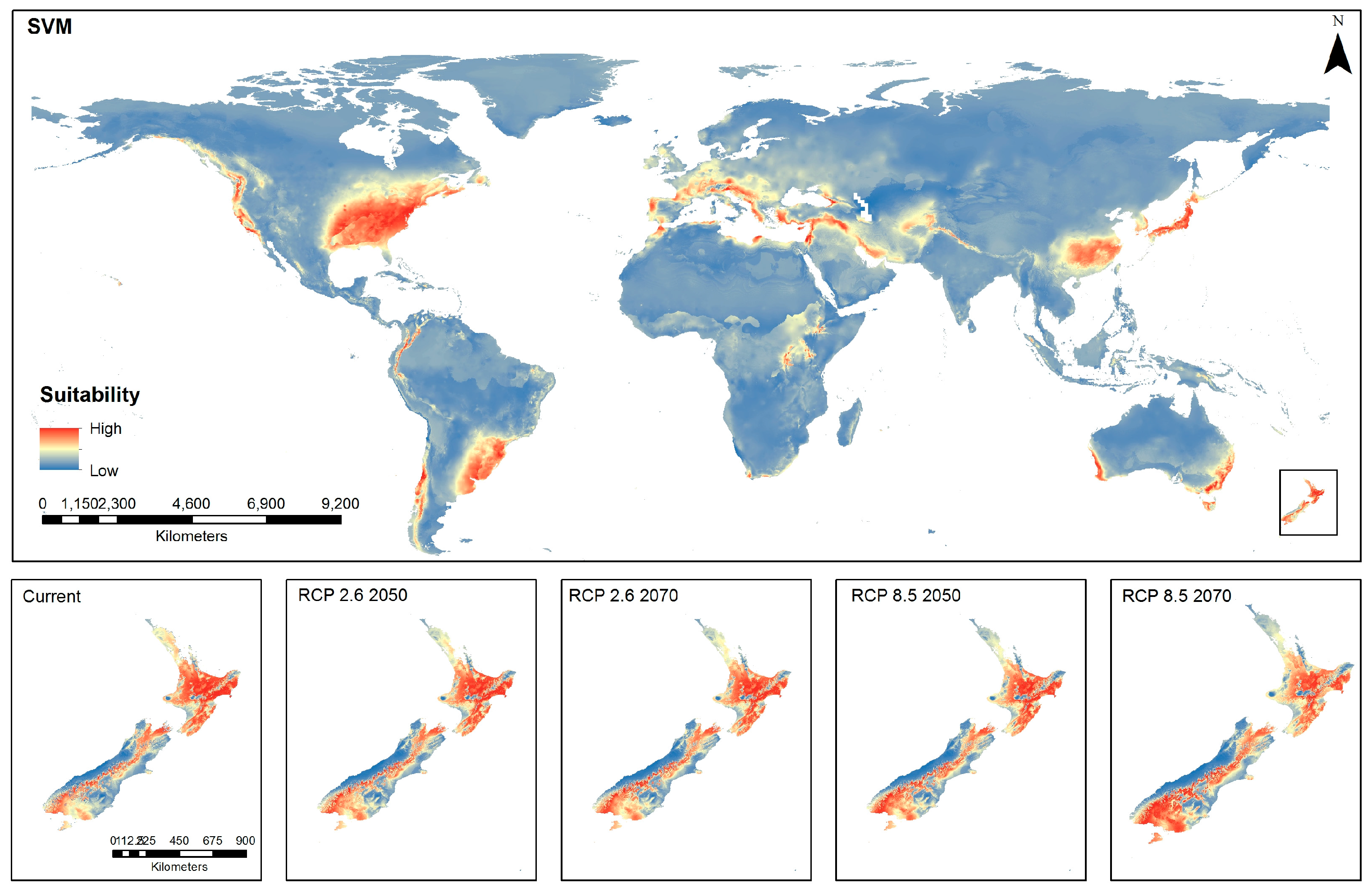
3.4. Spatial Distribution Using Support Vector Machine (SVM) Model
3.5. Spatial Distribution Using Maxent Model
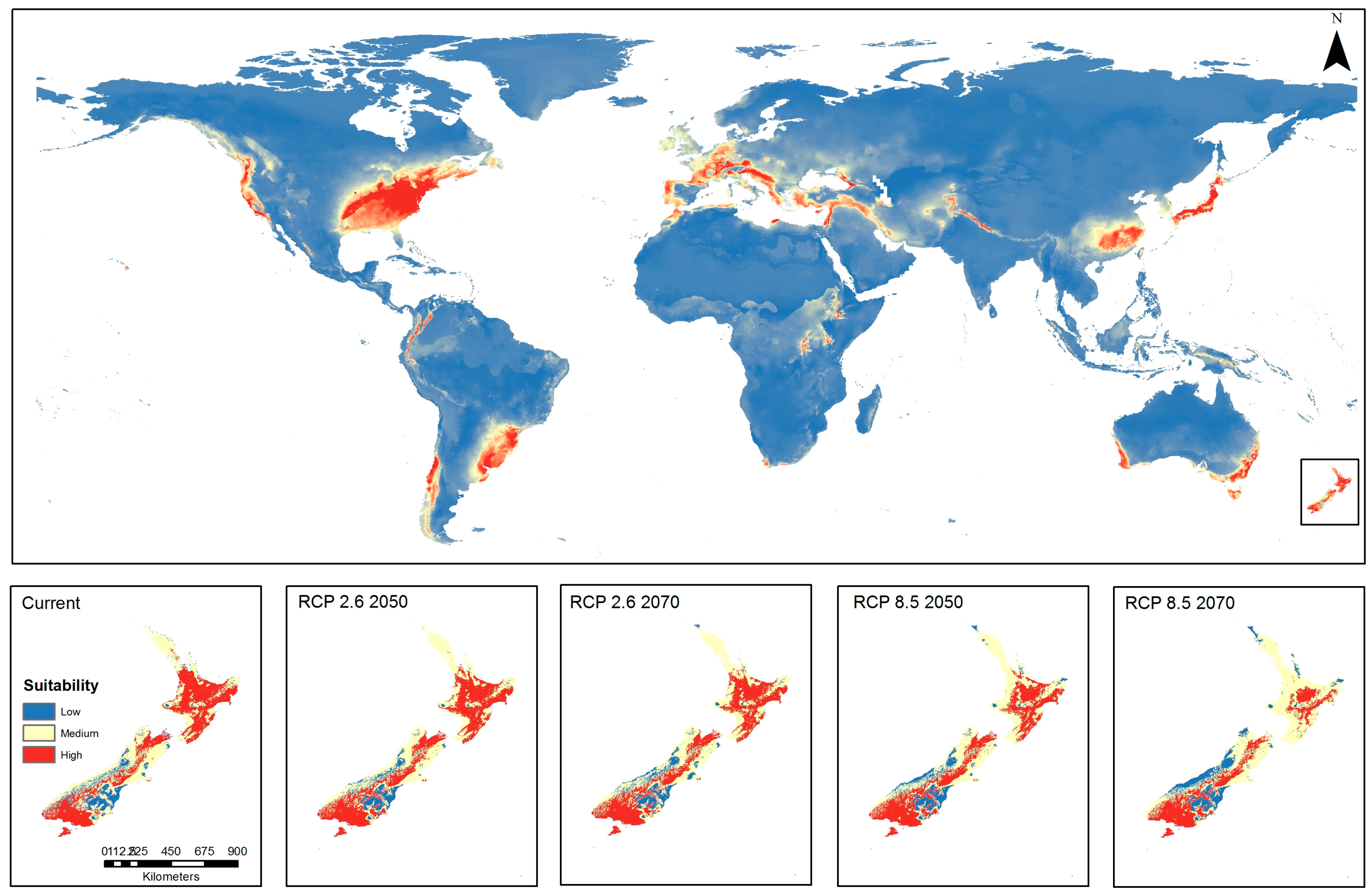
3.6. Ensemble Prediction
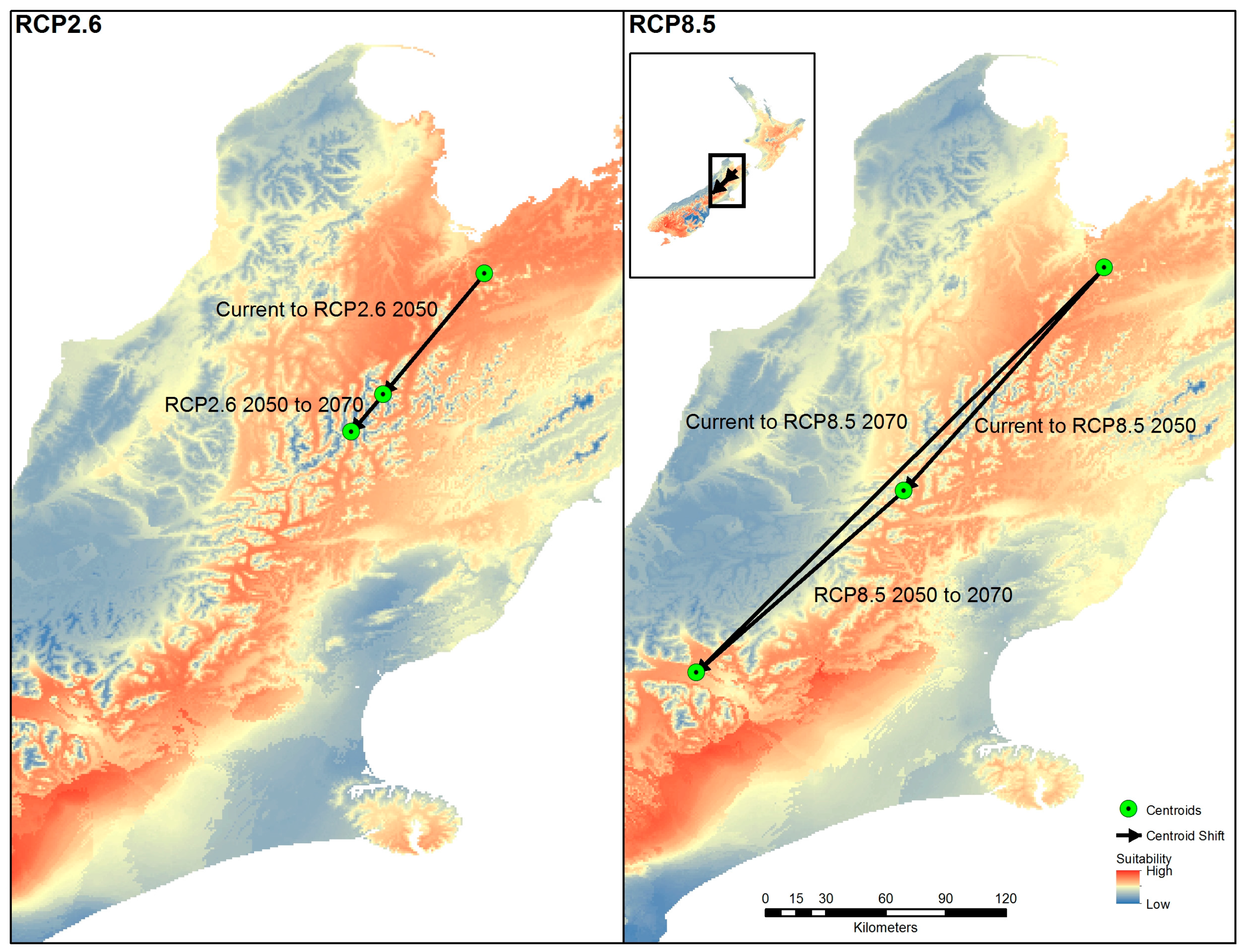
3.7. Centroid Shift of Environmental Suitability for BMSB in New Zealand as Determined Using the Ensemble Model
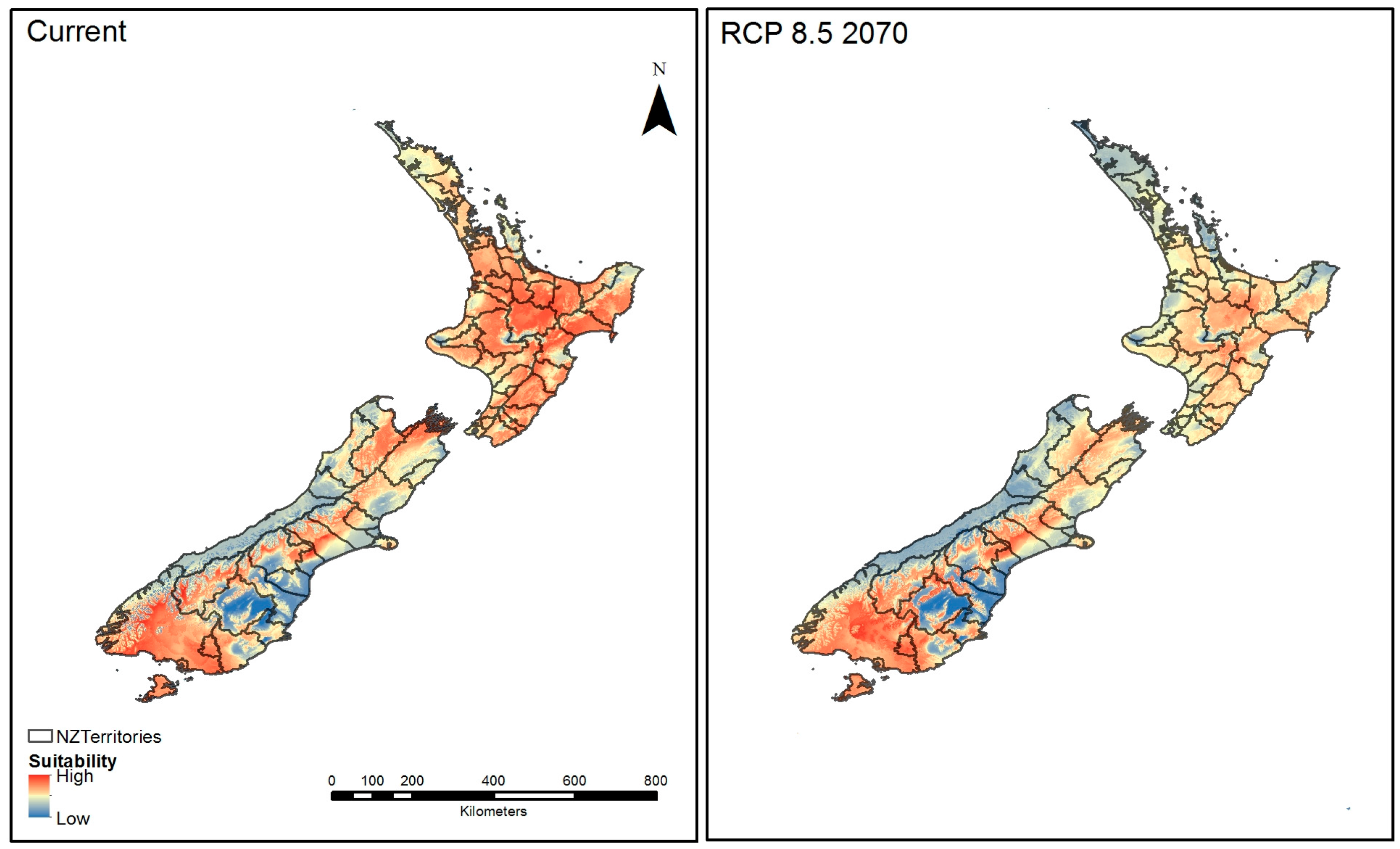
3.8. Ensembled Model Prediction Map Combined with New Zealand Official Territories
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ministry of Primary Industries (MPI). Protection and Response: Brown Marmorated Stink Bug. 2017. Available online: https://www.mpi.govt.nz/protection-and-response/responding/alerts/brown-marmorated-stink-bug/ (accessed on 6 March 2017).
- The Pennsylvanian State University. PennState College of Agriculture Insect Advice from Extension: Brown Marmorated Stink Bug, Halyomorpha halys. 2017. Available online: http://ento.psu.edu/extension/factsheets/brown-marmorated-stink-bug (accessed on 19 September 2017).
- Hoebeke, E.R.; Carter, M.E. Halyomorpha halys (Stål) (Heteroptera: Pentatomidea): A polyphagous plant pest from Asia newly detected in North America. Proc. Entomol. Soc. Wash. 2003, 105, 225–237. [Google Scholar]
- Northern Invasive Pest Management Center. Where is BMSB? State-By-State. 2017. Available online: http://www.stopbmsb.org/where-is-bmsb/ (accessed on 27 March 2017).
- Xu, J.; Fonseca, D.M.; Hamilton, G.C.; Hoelmer, K.A.; Nielsen, A.L. Tracing the origin of US brown marmorated stink bugs. Halyomorpha halys. Biol. Invasions 2014, 16, 153–166. [Google Scholar] [CrossRef]
- Arnold, K. Halyomorpha halys (Stål, 1855) a stink bug species newly detected among the European fauna (Insecta: Heteroptera, Pentatomidae, Pentatominae, cappaeini). Mitteilungen Thuringer Entomol. 2009, 16, 19. [Google Scholar]
- Wermelinger, B.; Wyniger, D.; Forster, B. First records of an invasive bug in Europe: Halyomorpha halys (Heteroptera: Pentatomidae), a new pest on woody ornamentals and fruit trees? Mitteilungen Schweizerischen Entomol. Gesellschaft 2008, 81, 1–8. [Google Scholar]
- Heckmann, R. First evidence of Halyomorpha halys (Stål, 1855) (Heteroptera: Pentatomidae) in Germany. Heteropteron H 2012, 36, 17–18. [Google Scholar]
- Callot, H.; Brua, C. Halyomorpha halys (Stål, 1855), the marmorated stink bug, new species for the fauna of France (Heteroptera: Pentatomidae). LÕEntomologiste 2013, 69, 69–71. [Google Scholar]
- Pansa, M.G.; Asteggiano, L.; Costamagna, C.; Vittone, G.; Tavella, L. First discovery of Halyomorpha halys in peach orchards in Piedmont. (Primo ritrovamento di Halyomorpha halys nei pescheti piemontesi.). Informatore Agrario 2013, 69, 60–61. [Google Scholar]
- Vétek, G.; Papp, V.; Haltrich, A.; Rédei, D. First record of the brown marmorated stink bug, Halyomorpha halys (Hemiptera: Heteroptera: Pentatomidae), in Hungary, with description of the genitalia of both sexes. Zootaxa 2014, 3780, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Harris, A.C. Halyomorpha halys (Hemiptera: Pentomidae) and Protaetia brevitaris (Coleoptera: Scarabaeidae: Cetoniinae) intercepted in Dunedin. Weta 2010, 40, 42–44. [Google Scholar]
- Kiwi Fruit Vine Health (KVH). Brown Marmorated Stink Bug—KVH Activity Update, March 2016. Available online: http://www.kvh.org.nz/vdb/document/102656 (accessed on 29 March 2017).
- Landcare Research. Brown Marmorated Stink Bug—Halyomorpha halys (Stål, 1855). 2017. Available online: https://www.landcareresearch.co.nz/resources/identification/animals/pentatomidae/factsheets/halyomorpha-halys (accessed on 15 August 2017).
- Funayama, K. Importance of apple fruits as food for the brown marmorated stink bug, Halyomorpha halys (Stål) (Heteroptera: Pentatomidae). Appl. Entomol. Zool. 2004, 39, 617–623. [Google Scholar] [CrossRef]
- Rice, K.B.; Bergh, C.J.; Bergmann, E.J.; Biddinger, D.J.; Dieckhoff, C.; Dively, G.; Fraser, H.; Gariepy, T.; Hamilton, G.; Haye, T.; et al. Biology, ecology, and management of brown marmorated stink bug (Hemiptera: Pentatomidae). J. Integr. Pest Manag. 2014, 5, A1–A13. [Google Scholar] [CrossRef]
- Leskey, T.C.; Hamilton, G.C.; Nielsen, A.L.; Olk, D.F.; Rodriguez-Saona, C.; Bergh, J.C.; Herbert, D.A.; Kuhar, T.P.; Pfeiffer, D.; Dively, G.P.; et al. Pest status of the brown marmorated stink bug, Halyomorpha halys in the USA. Outlooks Pest Manag. 2012, 23, 218–226. [Google Scholar] [CrossRef]
- Minnesota Department of Agriculture. Brown Marmorated Stink Bug. 2017. Available online: https://www.mda.state.mn.us/plants/insects/stinkbug.aspx (accessed on 3 April 2017).
- Leskey, T.; Hamilton, G.C. Brown Marmorated Stink Bug Working Group Meeting. 2010. Available online: https://projects.ipmcenters.org/Northeastern/FundedProjects/ReportFiles/Pship2010/Pship2010-Leskey-ProgressReport-237195-Meeting-2010_11_17.pdf (accessed on 4 April 2017).
- Bergmann, E.; Bernhard, K.M.; Bernon, G.; Bickerton, M.; Gill, S.; Gonzales, C.; Hamilton, G.C.; Hedstrom, C.; Kamminga, K.; Koplinka-Loehr, C.; et al. Host Plants of the Brown Marmorated Stink Bug in the U.S. 2016. Available online: http://www.stopbmsb.org/where-is-bmsb/host-plants/ (accessed on 3 April 2017).
- Martinson, H.M.; Raupp, M.J.; Shrewsbury, P.M. Invasive stink bug wounds trees, liberates sugars, and facilitates native Hymenoptera. Ann. Entomol. Soc. Am. 2013, 106, 47–52. [Google Scholar] [CrossRef]
- Inkley, D.B. Characteristics of home invasion by the brown marmorated stink bug (Hemiptera: Pentatomidae). J. Entomol. Sci. 2012, 47, 125–130. [Google Scholar] [CrossRef]
- United States Environmental Protection Agency. Brown Marmorated Stink Bug. 2017. Available online: https://www.epa.gov/safepestcontrol/brown-marmorated-stink-bug#agriculture (accessed on 3 April 2017).
- Penn State University. Insect Advice from Extension: Brown Marmorated Stink Bug. 2017. Available online: http://ento.psu.edu/extension/factsheets/brown-marmorated-stink-bug (accessed on 3 April 2017).
- Peterson, A.T.; Nakazawa, Y. Environmental data sets matter in ecological niche modelling: An example with Solenopsis Invicta and Solenopsis richteri. Glob. Ecol. Biogeogr. 2008, 17, 135–144. [Google Scholar] [CrossRef]
- Kulhanek, S.A.; Leung, B.; Ricciardi, A. Using ecological niche models to predict the abundance and impact of invasive species: Application to the common carp. Ecol. Appl. 2011, 21, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Fraser, D.L.; Aguilar, G.; Nagle, W.; Galbraith, M.; Ryall, C. The house crow (Corvus splendens): A threat to New Zealand? Int. J. Geo-Inf. 2015, 4, 725–740. [Google Scholar] [CrossRef]
- Zhu, G.; Bu, W.; Gao, Y.; Liu, G. Potential geographic distribution of brown marmorated stink bug invasion (Halyomorpha halys). PLoS ONE 2012, 7, e31246. [Google Scholar] [CrossRef] [PubMed]
- Hellmann, J.J.; Byers, J.E.; Bierwagen, B.G.; Dukes, J.S. Five potential consequences of climate change in invasive species. Conserv. Biol. 2008, 22, 534–543. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.D. Climate, climate change and range boundaries. Divers. Distrib. 2010, 16, 488–495. [Google Scholar] [CrossRef]
- Rochlin, I.; Ninivaggi, D.V.; Hutchinson, M.L.; Farajollahi, A. Climate change and range expansion of the Asian tiger mosquito (Aedes albopictus) in northeastern USA: Implications for public health practitioners. PLoS ONE 2013, 8, e60874. [Google Scholar] [CrossRef] [PubMed]
- Aguilar, G.; Blanchon, D.; Foote, H.; Pollonais, C.; Mosee, A. Queensland Fruit Fly Invasion of New Zealand: Predicting Area Suitability Under Future Climate Change Scenarios; Unitec e-Press Perspectives in Biosecurity Research Series, 2: Auckland, New Zealand, 2015; pp. 1–15. Available online: http://www.unitec.ac.nz/epress/index.php/queensland-fruit-fly-invasion-of-new-zealand-predicting-area-suitability-under-future-climate-change-scenarios/ (accessed on 24 April 2017).
- CERIS. Survey Status of Brown Marmorated Stink Bug—Halyomorpha halys (2016); Center for Environmental and Research Information Systems Purdue University: West Lafayette, IN, USA, 4 April 2016. Available online: http://pest.ceris.purdue.edu/map.php?code=IQAQQKA&year=2016 (accessed on 4 May 2016).
- Brown, J.L. SDMtoolbox: A python-based GIS toolkit for landscape genetic, biogeographic, and species distribution model analyses. Methods Ecol. Evol. 2014, 5, 694–700. [Google Scholar] [CrossRef]
- Syfert, M.M.; Smith, M.J.; Coomes, D. The effects of sampling bias and model complexity on the predictive performance of Maxent species distribution models. PLoS ONE 2013, 8, e55158. [Google Scholar] [CrossRef]
- Boria, R.A.; Olson, L.E.; Goodman, S.M.; Anderson, R.P. Spatial filtering to reduce sampling bias can improve the performance of ecological niche models. Ecol. Model. 2014, 275, 73–77. [Google Scholar] [CrossRef]
- Nazeri, M.; Kumar, L.; Jusoff, K.; Bahaman, A.R. Modelling the potential distribution of sun bear in krau wildlife reserve, Malaysia. Ecol. Inf. 2014, 20, 27–32. [Google Scholar] [CrossRef]
- Carraro, C.; Lanza, A.; Tavoni, M. All You Need to Know about the IPCC 5th Assessment Report. Mitigation of Climate Change. Review of Environment, Energy and Economics-Re3. 2014. Available online: http://www.feem.it/getpage.aspx?id=6312 (accessed on 25 March 2017).
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Münkemüller, T. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- Peterson, A.T. Ecological niche conservatism: A time-structured review of evidence. J. Biogeogr. 2011, 38, 817–828. [Google Scholar] [CrossRef]
- Rödder, D.; Lötters, S. Niche shift versus niche conservatism? Climatic characteristics of the native and invasive ranges of the Mediterranean house gecko (Hemidactylus turcicus). Glob. Ecol. Biogeogr. 2009, 18, 674–687. [Google Scholar] [CrossRef]
- Naimi, B. usdm: Uncertainty Analysis for Species Distribution Models. R Package Version 1.1-15. 2015. Available online: https://cran.r-project.org/web/packages/usdm/usdm.pdf (accessed on 23 June 2017).
- Aguilar, G.D.; Farnworth, M.J.; Winder, L. Mapping the stray domestic cat (Felis catus) population in New Zealand: Species distribution modelling with a climate change scenario and implications for protected areas. Appl. Geogr. 2015, 63, 146–154. [Google Scholar] [CrossRef]
- Aguilar, G.D.; Blanchon, D.J.; Foote, H.; Pollonais, C.W.; Mosee, A.N. A performance based consensus approach for predicting spatial extent of the Chinese windmill palm (Trachycarpus fortunei) in New Zealand under climate change. Ecol. Inf. 2017, 39, 130–139. [Google Scholar] [CrossRef]
- Duan, R.Y.; Kong, X.Q.; Huang, M.Y.; Fan, W.Y.; Wang, Z.G. The predictive performance and stability of six species distribution models. PLoS ONE 2014, 9, e112764. [Google Scholar] [CrossRef] [PubMed]
- Swets, J.A. Measuring the accuracy of diagnostic systems. Science 1988, 240, 1285–1293. [Google Scholar] [CrossRef] [PubMed]
- Peterson, A.T.; Soberon, J.; Pearson, R.G.; Anderson, R.P.; Martinez-Meyer, E.; Nakamura, M.; Araujo, M.B. Ecological Niches and Geographic Distributions; Princeton University Press: Princeton, NJ, USA, 2011; ISBN 978-0691136882. [Google Scholar]
- Cohen, E.J.M. Conceptual design and feasibility study of navy deep submergence vehicles. Nav. Eng. J. 1968, 80, 875–887. [Google Scholar] [CrossRef]
- Pearson, R.G.; Dawson, T.P.; Liu, C. Modelling species distributions in Britain: A hierarchical integration of climate and land-cover data. Ecography 2004, 27, 285–298. [Google Scholar] [CrossRef]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistics (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Araújo, M.B.; Guisan, A. Five (or so) challenges for species distribution modelling. J. Biogeogr. 2006, 33, 1677–1688. [Google Scholar] [CrossRef]
- Marmion, M.; Parviainen, M.; Luoto, M.; Heikkinen, R.K.; Thuiller, W. Evaluation of consensus methods in predictive species distribution modelling. Divers. Distrib. 2009, 5, 59–69. [Google Scholar] [CrossRef]
- Roura-Pascual, N.; Brotons, L.; Peterson, A.T.; Thuiller, W. Consensual predictions of potential distributional areas for invasive species: A case study of Argentine ants in the Iberian Peninsula. Biol. Invasions 2008, 11, 1017–1031. [Google Scholar] [CrossRef]
- Poulos, H.; Chernoff, B.; Fuller, P.; Butman, D. Ensemble forecasting of potential habitat for three invasive fishes. Aquat. Invasions 2012, 7, 59–72. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, S.; Sun, P.; Wang, T.; Wang, G.; Zhang, X.; Wang, L. Consensus forecasting of species distributions: The effects of niche model performance and niche properties. PLoS ONE 2015, 10, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Hijmans, R.J.; Elith, J. Species Distribution Modeling with R. 2016. Available online: https://cran.r-project.org/web/packages/dismo/vignettes/sdm.pdf (accessed on 25 February 2017).
- Jarnevich, C.S.; Stohlgren, T.J.; Kumar, S.; Morisette, J.T.; Holcombe, T.R. Caveats for correlative species distribution modeling. Ecol. Inf. 2015, 29, 6–15. [Google Scholar] [CrossRef]
- Liu, C.; White, M.; Newell, G. Selecting thresholds for the prediction of species occurrence with presence-only data. J. Biogeogr. 2013, 40, 778–789. [Google Scholar] [CrossRef]
- Muirhead, J.R.; Leug, B.; van Overdyk, C.; Kelly, D.W.; Nanda Kumar, K.; Marchant, K.R.; MacIsaac, H.J. Modelling local and long-distance dispersal of invasive emerald ash borer Agrilus planipennis (Coleoptera) in North America. Divers. Distrib. 2006, 12, 71–79. [Google Scholar] [CrossRef]
- Hartley, S.; Krushelnycky, P.D.; Lester, P.J. Integrated physiology, population dynamics and climate to make multi-scale predictions for the spread of an invasive insect: The Argentine ant at Haleakala National Park, Hawaii. Ecography 2010, 33, 83–94. [Google Scholar] [CrossRef]
- Dukes, J.S.; Pontius, J.; Orwig, D.; Garnas, J.R.; Rodgers, V.L.; Brazee, N.; Cooke, B.; Theoharides, K.A.; Stange, E.E.; Harrington, R.; et al. Responses of insect pests, pathogens, and invasive plant species to climate change in forests of northeastern North America: What can we predict? Can. J. For. Res. 2009, 39, 231–248. [Google Scholar] [CrossRef]
- Estay, S.A.; Lima, M.; Labra, F.A. Predicting insect pest status under climate change scenarios: Combining experimental data and population dynamics modelling. J. Appl. Entomol. 2009, 133, 491–499. [Google Scholar] [CrossRef]
- Ministry of the Environment and Statistics New Zealand. Environmental Report Series. Environment Aoteoroa; 2015. Available online: http://www.mfe.govt.nz/sites/default/files/media/Environmental%20reporting/Environment-Aotearoa-2015.pdf (accessed on 25 April 2017).
- Chen, I.-C.; Hill, J.K.; Ohlemüller, R.; Roy, D.B.; Thomas, C.D. Rapid range shifts of species associated with high levels of climate warming. Science 2011, 333, 1024–1026. [Google Scholar]
- Ministry of Primary Industries, New Zealand. Situations and Outlook for Primary Industries, December 2016. Available online: https://www.mpi.govt.nz/ (accessed on 4 April 2017).
- Sakai, A.K.; Allendorf, F.W.; Holt, J.S.; Lodge, D.M.; Molofsky, J.; With, K.A.; Baughman, S.; Cabin, R.J.; Cohen, J.E.; Ellstrand, N.C.; et al. The population biology of invasive species. Ann. Rev. Ecol. Syst. 2001, 32, 305–332. [Google Scholar] [CrossRef]
- Lockwood, J.L.; Cassey, P.; Blackburn, T. The role of propagule pressure in explaining species invasions. Trends Ecol. Evol. 2005, 20, 223–228. [Google Scholar] [CrossRef] [PubMed]
- StatsNZ. International Travel and Migration: July 2017. Available online: http://www.stats.govt.nz/browse_for_stats/population/Migration/IntTravelAndMigration_HOTPJul17.aspx (accessed on 22 August 2017).
- StatsNZ. Overseas Merchandise Trade: June 2017. Available online: http://www.stats.govt.nz/browse_for_stats/industry_sectors/imports_and_exports/OverseasMerchandiseTrade_HOTPJun17.aspx (accessed on 22 August 2017).
- Wallner, A.M.; Hamilton, G.C.; Nielsen, A.L.; Hahn, N.; Green, E.J.; Rodriguez-Saona, C.R. Landscape factors facilitating the invasive dynamics and distribution of the brown marmorated stink bug, Halyomorpha halys (Hemiptera: Pentatomidae), after arrival in the United States. PLoS ONE 2014, 9, e95691. [Google Scholar] [CrossRef] [PubMed]
- New Zealand Department of Conservation (DOC) Whakawhiti Korero: Working Better by Working Together. March 2017. Available online: http://us8.campaign-archive2.com/?u=2956fc2bebc9e28a4cf526b4e&id=fe87d0f26a&e=d06cc1b4d9 (accessed on 10 April 2017).
- Simberloff, D. The role of propagule pressure in biological invasions. Ann. Rev. Ecol. Evol. Syst. 2009, 40, 81–102. [Google Scholar] [CrossRef]
- Brockerhoff, E.G.; Kimberley, H.; Liebhold, A.M.; Haack, R.A.; Carey, J.F. Predicting how altering the propagule pressure changes establishment rates of biological invaders across species pool. Ecology 2014, 95, 594–601. [Google Scholar] [CrossRef] [PubMed]
- Fraser, D.; Galbraith, M.; Adams, N.; Blanchon, D. Range expansion in the spur-winged plover (Vanellus miles novaehollandiae) in New Zealand. Notornis 2014, 61, 43–47. [Google Scholar]
- Ministry of Primary Industries. Brown Marmorated Stink Bug Risk Management—2016/17 Season (as at 20th March 2017). Available online: http://www.kvh.org.nz/vdb/document/103524 (accessed on 19 September 2017).








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Layer | Climatic Variable | Mean | Standard Error | Max | Min |
|---|---|---|---|---|---|
| * BIO1 | Annual Mean Temperature | 11.58 | 2.01 | 19.60 | 1.20 |
| * BIO2 | Mean Diurnal Range (Mean of monthly (max temp—min temp)) | 10.98 | 7.50 | 14.10 | 7.70 |
| BIO3 | Isothermality (BIO2/BIO7) (* 100) | 3.32 | 5.38 | 5.50 | 2.30 |
| BIO4 | Temperature Seasonality (standard deviation * 100) | 791.09 | 3.96 | 1000.20 | 303.20 |
| BIO5 | Max Temperature of Warmest Month | 28.65 | 4.32 | 35.20 | 16.90 |
| BIO6 | Min Temperature of Coldest Month | 4.54 | 2.11 | 8.60 | −15.50 |
| BIO7 | Temperature Annual Range (BIO5-BIO6) | 33.19 | 145.25 | 41.20 | 19.10 |
| * BIO8 | Mean Temperature of Wettest Quarter | 18.11 | 0.72 | 24.70 | −1.50 |
| * BIO9 | Mean Temperature of Driest Quarter | 4.83 | 1.56 | 22.70 | −7.10 |
| BIO10 | Mean Temperature of Warmest Quarter | 21.64 | 2.79 | 28.00 | 11.00 |
| BIO11 | Mean Temperature of Coldest Quarter | 1.30 | 7.85 | 14.30 | −8.70 |
| * BIO12 | Annual Precipitation | 1125.71 | 13.20 | 1928.00 | 315.00 |
| BIO13 | Precipitation of Wettest Month | 132.11 | 7.45 | 257.00 | 68.00 |
| BIO14 | Precipitation of Driest Month | 63.00 | 9.79 | 106.00 | 0.00 |
| * BIO15 | Precipitation Seasonality (Coefficient of Variation) | 27.40 | 2.24 | 101.00 | 7.00 |
| BIO16 | Precipitation of Wettest Quarter | 367.72 | 2.40 | 739.00 | 184.00 |
| BIO17 | Precipitation of Driest Quarter | 207.45 | 3.56 | 344.00 | 4.00 |
| BIO18 | Precipitation of Warmest Quarter | 315.41 | 28.72 | 589.00 | 9.00 |
| * BIO19 | Precipitation of Coldest Quarter | 259.65 | 4.41 | 685.00 | 105.00 |
| Algorithm | AUC | Kappa | TSS |
|---|---|---|---|
| RF | 0.98739 | 0.51379 | 0.59695 |
| SVM | 0.98488 | 0.45132 | 0.58917 |
| Maxent | 0.98970 | 0.39969 | 0.18408 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fraser, D.; Kumar, S.; Aguilar, G. Mapping the Potential Global Range of the Brown Marmorated Stink Bug, Halyomorpha halys, with Particular Reference to New Zealand. Climate 2017, 5, 75. https://doi.org/10.3390/cli5030075
Fraser D, Kumar S, Aguilar G. Mapping the Potential Global Range of the Brown Marmorated Stink Bug, Halyomorpha halys, with Particular Reference to New Zealand. Climate. 2017; 5(3):75. https://doi.org/10.3390/cli5030075
Chicago/Turabian StyleFraser, Diane, Shivani Kumar, and Glenn Aguilar. 2017. "Mapping the Potential Global Range of the Brown Marmorated Stink Bug, Halyomorpha halys, with Particular Reference to New Zealand" Climate 5, no. 3: 75. https://doi.org/10.3390/cli5030075
APA StyleFraser, D., Kumar, S., & Aguilar, G. (2017). Mapping the Potential Global Range of the Brown Marmorated Stink Bug, Halyomorpha halys, with Particular Reference to New Zealand. Climate, 5(3), 75. https://doi.org/10.3390/cli5030075