
Climate Change, Ecosystem Processes and Biological Diversity Responses in High Elevation Communities


{kind=link}
{kind=link}
{kind=link}
Abstract
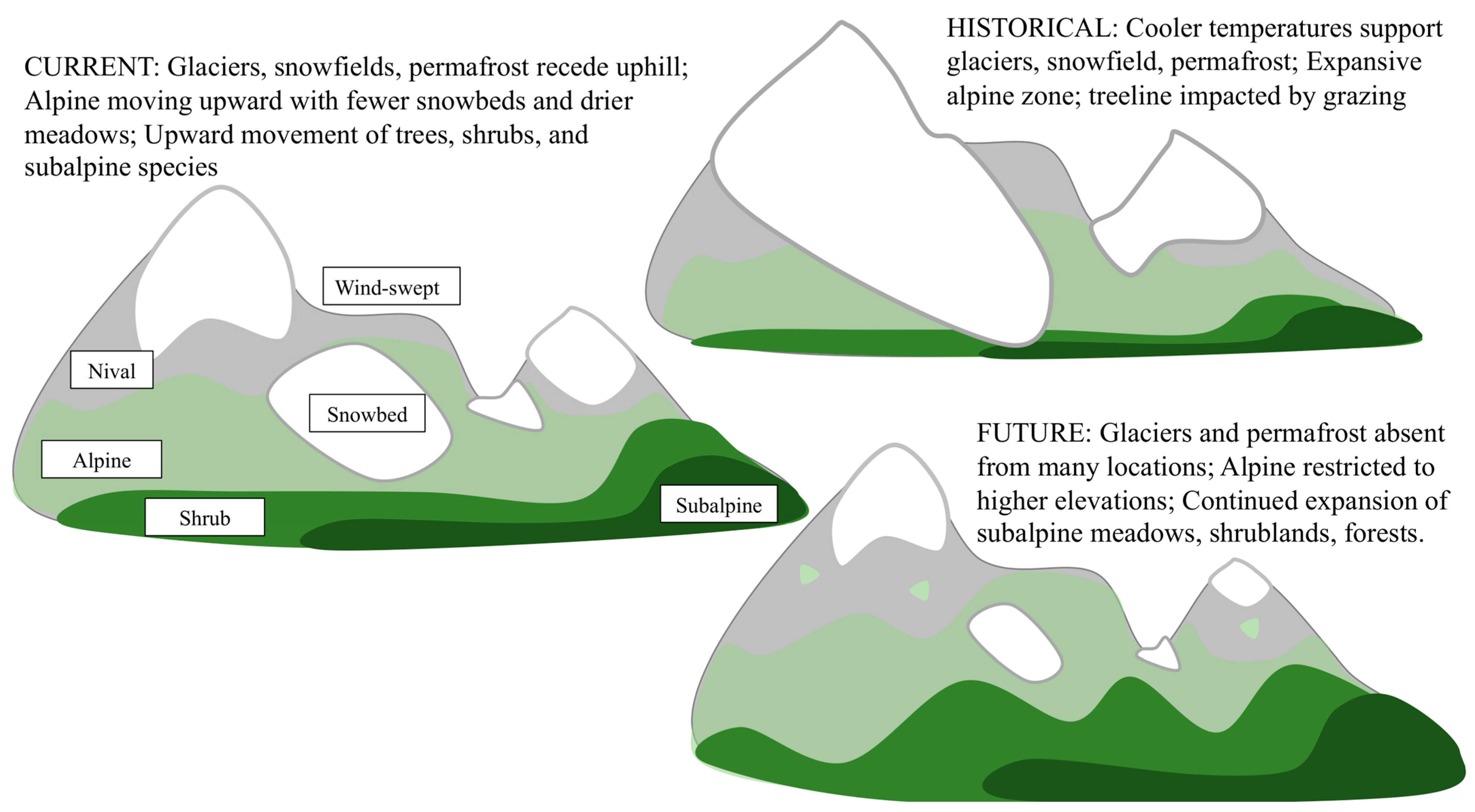
:1. Introduction
2. Identifying and Quantifying the Components of Climate Change for High Elevation Species
3. Patterns of Species Richness and Vulnerability to Climate Change
4. How Warming Will Change Local Plant Community Composition
5. Herbaceous Species Moving Uphill
6. Shrublandification and Tree Encroachment
7. The Increasing Importance of Fire
8. Conclusions
- High elevation systems in those areas of the planet experiencing the most rapid change;
- Those species whose current climate regimes will no longer exist due to either shifts to non-analogue climate conditions or lack of additional habitat at the highest elevations;
- Those species adapted to snowbed communities and short growing seasons;
- Those species in moister habitats more vulnerable to woody species invasions.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Verrall, B.; Pickering, C.M. Alpine Vegetation in the Context of Climate Change: A Global Review of Past Research and Future Directions. Sci. Total Environ. 2020, 748, 141344. [Google Scholar] [CrossRef] [PubMed]
- Catalan, J.; Ninot, J.M.; Aniz, M.M. (Eds.) High Mountain Conservation in a Changing World; Advances in Global Change Research; Springer International Publishing: Cham, Switzerland, 2017; Volume 62, ISBN 978-3-319-55981-0. [Google Scholar]
- Wilson, J.B.; Peet, R.K.; Dengler, J.; Pärtel, M. Plant Species Richness: The World Records. J. Veg. Sci. 2012, 23, 796–802. [Google Scholar] [CrossRef]
- Nogués-Bravo, D.; Araújo, M.B.; Errea, M.P.; Martínez-Rica, J.P. Exposure of Global Mountain Systems to Climate Warming during the 21st Century. Glob. Environ. Chang. 2007, 17, 420–428. [Google Scholar] [CrossRef]
- Grimm, N.B.; Chapin, F.S.; Bierwagen, B.; Gonzalez, P.; Groffman, P.M.; Luo, Y.; Melton, F.; Nadelhoffer, K.; Pairis, A.; Raymond, P.A.; et al. The Impacts of Climate Change on Ecosystem Structure and Function. Front. Ecol. Environ. 2013, 11, 474–482. [Google Scholar] [CrossRef] [Green Version]
- Mountain Research Initiative EDW Working Group Elevation-Dependent Warming in Mountain Regions of the World. Nat. Clim. Chang. 2015, 5, 424–430. [CrossRef] [Green Version]
- Rumpf, S.B.; Hülber, K.; Klonner, G.; Moser, D.; Schütz, M.; Wessely, J.; Willner, W.; Zimmermann, N.E.; Dullinger, S. Range Dynamics of Mountain Plants Decrease with Elevation. Proc. Natl. Acad. Sci. USA 2018, 115, 1848–1853. [Google Scholar] [CrossRef] [Green Version]
- Wershow, S.T.; DeChaine, E.G. Retreat to Refugia: Severe Habitat Contraction Projected for Endemic Alpine Plants of the Olympic Peninsula. Am. J. Bot. 2018, 105, 760–778. [Google Scholar] [CrossRef] [Green Version]
- Dolezal, J.; Jandova, V.; Macek, M.; Mudrak, O.; Altman, J.; Schweingruber, F.H.; Liancourt, P. Climate Warming Drives Himalayan Alpine Plant Growth and Recruitment Dynamics. J. Ecol. 2021, 109, 179–190. [Google Scholar] [CrossRef]
- Suggitt, A.J.; Wilson, R.J.; Isaac, N.J.B.; Beale, C.M.; Auffret, A.G.; August, T.; Bennie, J.J.; Crick, H.Q.P.; Duffield, S.; Fox, R.; et al. Extinction Risk from Climate Change Is Reduced by Microclimatic Buffering. Nat. Clim. Chang. 2018, 8, 713–717. [Google Scholar] [CrossRef]
- Graae, B.J.; Vandvik, V.; Armbruster, W.S.; Eiserhardt, W.L.; Svenning, J.-C.; Hylander, K.; Ehrlén, J.; Speed, J.D.M.; Klanderud, K.; Bråthen, K.A.; et al. Stay or Go—How Topographic Complexity Influences Alpine Plant Population and Community Responses to Climate Change. Perspect. Plant Ecol. Evol. Syst. 2018, 30, 41–50. [Google Scholar] [CrossRef] [Green Version]
- Oldfather, M.F.; Kling, M.M.; Sheth, S.N.; Emery, N.C.; Ackerly, D.D. Range Edges in Heterogeneous Landscapes: Integrating Geographic Scale and Climate Complexity into Range Dynamics. Glob. Chang. Biol. 2020, 26, 1055–1067. [Google Scholar] [CrossRef]
- Elsen, P.R.; Tingley, M.W. Global Mountain Topography and the Fate of Montane Species under Climate Change. Nat. Clim. Chang. 2015, 5, 772–776. [Google Scholar] [CrossRef]
- Alexander, J.M.; Diez, J.M.; Levine, J.M. Novel Competitors Shape Species’ Responses to Climate Change. Nature 2015, 525, 515–518. [Google Scholar] [CrossRef]
- Ackerly, D.D.; Kling, M.M.; Clark, M.L.; Papper, P.; Oldfather, M.F.; Flint, A.L.; Flint, L.E. Topoclimates, Refugia, and Biotic Responses to Climate Change. Front. Ecol. Environ. 2020, 18, 288–297. [Google Scholar] [CrossRef]
- Alexander, J.M.; Chalmandrier, L.; Lenoir, J.; Burgess, T.I.; Essl, F.; Haider, S.; Kueffer, C.; McDougall, K.; Milbau, A.; Nuñez, M.A.; et al. Lags in the Response of Mountain Plant Communities to Climate Change. Glob. Chang. Biol. 2018, 24, 563–579. [Google Scholar] [CrossRef] [PubMed]
- Körner, C. Alpine Plant Life: Functional Plant Ecology of High Mountain Ecosystems; Springer: Berlin/Heidelberg, Germany, 2003; ISBN 978-3-540-00347-2. [Google Scholar]
- Testolin, R.; Attorre, F.; Jiménez-Alfaro, B. Global Distribution and Bioclimatic Characterization of Alpine Biomes. Ecography 2020, 43, 779–788. [Google Scholar] [CrossRef] [Green Version]
- Malanson, G.P.; Resler, L.M.; Butler, D.R.; Fagre, D.B. Mountain Plant Communities: Uncertain Sentinels? Prog. Phys. Geogr. Earth Environ. 2019, 43, 521–543. [Google Scholar] [CrossRef]
- Grabherr, G.; Gottfried, M.; Pauli, H. Climate Change Impacts in Alpine Environments. Geogr. Compass 2010, 4, 1133–1153. [Google Scholar] [CrossRef] [Green Version]
- Leopold, M.; Lewis, G.; Dethier, D.; Caine, N.; Williams, M.W. Cryosphere: Ice on Niwot Ridge and in the Green Lakes Valley, Colorado Front Range. Plant Ecol. Divers. 2015, 8, 625–638. [Google Scholar] [CrossRef]
- Wang, H.; Liu, H.; Cao, G.; Ma, Z.; Li, Y.; Zhang, F.; Zhao, X.; Zhao, X.; Jiang, L.; Sanders, N.J.; et al. Alpine Grassland Plants Grow Earlier and Faster but Biomass Remains Unchanged over 35 Years of Climate Change. Ecol. Lett. 2020, 23, 701–710. [Google Scholar] [CrossRef] [Green Version]
- Ozturk, M. (Ed.) Climate Change Impacts on High-Altitude Ecosystems; Springer: Berlin, Germany, 2015; ISBN 978-3-319-12858-0. [Google Scholar]
- Llambí, L.D.; Rada, F. Ecological Research in the Tropical Alpine Ecosystems of the Venezuelan Páramo: Past, Present and Future. Plant Ecol. Divers. 2019, 12, 519–538. [Google Scholar] [CrossRef]
- Beniston, M.; Farinotti, D.; Stoffel, M.; Andreassen, L.M.; Coppola, E.; Eckert, N.; Fantini, A.; Giacona, F.; Hauck, C.; Huss, M.; et al. The European Mountain Cryosphere: A Review of Its Current State, Trends, and Future Challenges. Cryosphere 2018, 12, 759–794. [Google Scholar] [CrossRef] [Green Version]
- Formica, A.; Farrer, E.C.; Ashton, I.W.; Suding, K.N. Shrub Expansion Over the Past 62 Years in Rocky Mountain Alpine Tundra: Possible Causes and Consequences. Arct. Antarct. Alp. Res. 2014, 46, 616–631. [Google Scholar] [CrossRef] [Green Version]
- Crawford, J.T.; Hinckley, E.S.; Neff, J.C. Long-Term Trends in Acid Precipitation and Watershed Elemental Export From an Alpine Catchment of the Colorado Rocky Mountains, USA. J. Geophys. Res. Biogeosci. 2020, 125. [Google Scholar] [CrossRef]
- Giersch, J.J.; Hotaling, S.; Kovach, R.P.; Jones, L.A.; Muhlfeld, C.C. Climate-Induced Glacier and Snow Loss Imperils Alpine Stream Insects. Glob. Chang. Biol. 2017, 23, 2577–2589. [Google Scholar] [CrossRef]
- Oleksy, I.A.; Beck, W.S.; Lammers, R.W.; Steger, C.E.; Wilson, C.; Christianson, K.; Vincent, K.; Johnson, G.; Johnson, P.T.J.; Baron, J.S. The Role of Warm, Dry Summers and Variation in Snowpack on Phytoplankton Dynamics in Mountain Lakes. Ecology 2020, ecy.3132. [Google Scholar] [CrossRef]
- Oleksy, I.A.; Baron, J.S.; Beck, W.S. Nutrients and Warming Alter Mountain Lake Benthic Algal Structure and Function. Freshw. Sci. 2021, 40, 88–102. [Google Scholar] [CrossRef]
- Loria, K.A.; McKnight, D.; Ragar, D.M.; Johnson, P.T.J. The Life Aquatic in High Relief: Shifts in the Physical and Biological Characteristics of Alpine Lakes along an Elevation Gradient in the Rocky Mountains, USA. Aquat. Sci. 2020, 82, 11. [Google Scholar] [CrossRef]
- Messerli, B.; Viviroli, D.; Weingartner, R. Mountains of the World: Vulnerable Water Towers for the 21st Century. Ambio 2004, 13, 29–34. [Google Scholar] [CrossRef]
- Bandyopadhyay, J. Water Towers of the World. People Planet 1996, 5, 14–16. [Google Scholar]
- Moritz, C.; Agudo, R. The Future of Species Under Climate Change: Resilience or Decline? Science 2013, 341, 504–508. [Google Scholar] [CrossRef]
- Tramblay, Y.; Llasat, M.C.; Randin, C.; Coppola, E. Climate Change Impacts on Water Resources in the Mediterranean. Reg. Environ. Chang. 2020, 20, 83. [Google Scholar] [CrossRef]
- Inauen, N.; Körner, C.; Hiltbrunner, E. No Growth Stimulation by CO2 Enrichment in Alpine Glacier Forefield Plants. Glob. Chang. Biol. 2012, 18, 985–999. [Google Scholar] [CrossRef]
- Diaz, H.F.; Bradley, R.S. Temperature Variation during the Last Century at High Elevations Sites. Clim. Chang. 1997, 36, 253–279. [Google Scholar] [CrossRef]
- Hiebl, J.; Schöner, W. Temperature Inversions in Austria in a Warming Climate—Changes in Space and Time. metz 2018, 27, 309–323. [Google Scholar] [CrossRef]
- Collados-Lara, A.-J.; Pardo-Igúzquiza, E.; Pulido-Velazquez, D.; Jiménez-Sánchez, J. Precipitation Fields in an Alpine Mediterranean Catchment: Inversion of Precipitation Gradient with Elevation or Undercatch of Snowfall? Int. J. Clim. 2018, 38, 3565–3578. [Google Scholar] [CrossRef]
- Sevruk, B.; Ondrás, M.; Chvíla, B. The WMO Precipitation Measurement Intercomparisons. Atmos. Res. 2009, 92, 376–380. [Google Scholar] [CrossRef]
- McCullough, I.M.; Davis, F.W.; Dingman, J.R.; Flint, L.E.; Flint, A.L.; Serra-Diaz, J.M.; Syphard, A.D.; Moritz, M.A.; Hannah, L.; Franklin, J. High and Dry: High Elevations Disproportionately Exposed to Regional Climate Change in Mediterranean-Climate Landscapes. Landsc. Ecol. 2016, 31, 1063–1075. [Google Scholar] [CrossRef] [Green Version]
- McCain, C.M.; King, S.R.B.; Szewczyk, T.M. Unusually Large Upward Shifts in Cold-adapted, Montane Mammals as Temperature Warms. Ecology 2021. [Google Scholar] [CrossRef]
- Rapacciuolo, G.; Maher, S.P.; Schneider, A.C.; Hammond, T.T.; Jabis, M.D.; Walsh, R.E.; Iknayan, K.J.; Walden, G.K.; Oldfather, M.F.; Ackerly, D.D.; et al. Beyond a Warming Fingerprint: Individualistic Biogeographic Responses to Heterogeneous Climate Change in California. Glob. Chang. Biol. 2014, 20, 2841–2855. [Google Scholar] [CrossRef]
- Harsch, M.A.; HilleRisLambers, J. Climate Warming and Seasonal Precipitation Change Interact to Limit Species Distribution Shifts across Western North America. PLoS ONE 2016, 11, e0159184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ordonez, A.; Williams, J.W.; Svenning, J.-C. Mapping Climatic Mechanisms Likely to Favour the Emergence of Novel Communities. Nat. Clim. Chang. 2016, 6, 1104–1109. [Google Scholar] [CrossRef]
- Niittynen, P.; Heikkinen, R.K.; Aalto, J.; Guisan, A.; Kemppinen, J.; Luoto, M. Fine-Scale Tundra Vegetation Patterns Are Strongly Related to Winter Thermal Conditions. Nat. Clim. Chang. 2020, 10, 1143–1148. [Google Scholar] [CrossRef]
- Mote, P.W.; Li, S.; Lettenmaier, D.P.; Xiao, M.; Engel, R. Dramatic Declines in Snowpack in the Western US. Npj Clim. Atmos. Sci. 2018, 1, 2. [Google Scholar] [CrossRef]
- Musselman, K.N.; Addor, N.; Vano, J.A.; Molotch, N.P. Winter Melt Trends Portend Widespread Declines in Snow Water Resources. Nat. Clim. Chang. 2021. [Google Scholar] [CrossRef]
- Kittel, T.; White, C.; Hartman, M.; Chowanski, K.; Ackerman, T.; Williams, M.; Losleben, M. Infilled Air Temperature Data for C1 Chart Recorder, 1952–2018, Daily. Environ. Data Initiat. 2019. [Google Scholar] [CrossRef]
- Kittel, T.; White, C.; Hartman, M.; Chowanski, K.; Ackerman, T.; Williams, M.; Losleben, M. Infilled Air Temperature Data for D1 Chart Recorder, 1952–2018, Daily. Environ. Data Initiat. 2019. [Google Scholar] [CrossRef]
- Bueno de Mesquita, C.P.; Tillmann, L.S.; Bernard, C.D.; Rosemond, K.C.; Molotch, N.P.; Suding, K.N. Topographic Heterogeneity Explains Patterns of Vegetation Response to Climate Change (1972–2008) across a Mountain Landscape, Niwot Ridge, Colorado. Arct. Antarct. Alp. Res. 2018, 50, e1504492. [Google Scholar] [CrossRef] [Green Version]
- Sherrod, S.K.; Seastedt, T.R.; Walker, M.D. Northern Pocket Gopher (Thomomys Talpoides) Control of Alpine Plant Community Structure. Arct. Antarct. Alp. Res. 2005, 37, 585–590. [Google Scholar] [CrossRef] [Green Version]
- Erb, L.P.; Ray, C.; Guralnick, R. On the Generality of a Climate-Mediated Shift in the Distribution of the American Pika (Ochotona Princeps). Ecology 2011, 92, 1730–1735. [Google Scholar] [CrossRef]
- Hofgaard, A.; Løkken, J.O.; Dalen, L.; Hytteborn, H. Comparing Warming and Grazing Effects on Birch Growth in an Alpine Environment—A 10-Year Experiment. Plant Ecol. Divers. 2010, 3, 19–27. [Google Scholar] [CrossRef]
- Zhang, T.; Zhang, Y.; Xu, M.; Zhu, J.; Wimberly, M.C.; Yu, G.; Niu, S.; Xi, Y.; Zhang, X.; Wang, J. Light-Intensity Grazing Improves Alpine Meadow Productivity and Adaption to Climate Change on the Tibetan Plateau. Sci. Rep. 2015, 5, 15949. [Google Scholar] [CrossRef] [PubMed]
- Ingty, T. Pastoralism in the Highest Peaks: Role of the Traditional Grazing Systems in Maintaining Biodiversity and Ecosystem Function in the Alpine Himalaya. PLoS ONE 2021, 16, e0245221. [Google Scholar] [CrossRef] [PubMed]
- Körner, C. Alpine Plant Diversity: A Global Survey and Functional Interpretations. In Arctic and Alpine Biodiversity: Patterns, Causes and Ecosystem Consequences; Ecological, Studies; Chapin, F.S., Körner, C., Eds.; Springer: Berlin/Heidelberg, Germany, 1995; Volume 113, pp. 45–62. ISBN 978-3-642-78968-7. [Google Scholar]
- Taberlet, P.; Zimmermann, N.E.; Englisch, T.; Tribsch, A.; Holderegger, R.; Alvarez, N.; Niklfeld, H.; Coldea, G.; Mirek, Z.; Moilanen, A.; et al. Genetic Diversity in Widespread Species Is Not Congruent with Species Richness in Alpine Plant Communities. Ecol. Lett. 2012, 15, 1439–1448. [Google Scholar] [CrossRef] [PubMed]
- Rixen, C.; Wipf, S. Non-equilibrium in Alpine Plant Assemblages: Shifts in Europe’s Summit Floras. In High Mountain Conservation in a Changing World; Advances in Global Change, Research; Catalan, J., Ninot, J.M., Aniz, M.M., Eds.; Springer International Publishing: Cham, Switzerland, 2017; Volume 62, pp. 285–303. ISBN 978-3-319-55981-0. [Google Scholar]
- Bruun, H.H.; Moen, J.; Virtanen, R.; Grytnes, J.; Oksanen, L.; Angerbjörn, A. Effects of Altitude and Topography on Species Richness of Vascular Plants, Bryophytes and Lichens in Alpine Communities. J. Veg. Sci. 2006, 17, 37–46. [Google Scholar] [CrossRef]
- Johnson, P.L.; Billings, W.D. The Alpine Vegetation of the Beartooth Plateau in Relation to Cryopedogenic Processes and Patterns. Ecol. Monogr. 1962, 32, 105–135. [Google Scholar] [CrossRef]
- Smithers, B.V.; Oldfather, M.F.; Koontz, M.J.; Bishop, J.; Bishop, C.; Nachlinger, J.; Sheth, S.N. Community Turnover by Composition and Climatic Affinity across Scales in an Alpine System. Am. J. Bot. 2020, 107, 239–249. [Google Scholar] [CrossRef] [Green Version]
- Sproull, G.J.; Quigley, M.F.; Sher, A.; González, E. Long-Term Changes in Composition, Diversity and Distribution Patterns in Four Herbaceous Plant Communities along an Elevational Gradient. J. Veg. Sci. 2015, 26, 552–563. [Google Scholar] [CrossRef]
- Kazakis, G.; Ghosn, D.; Vogiatzakis, I.N.; Papanastasis, V.P. Vascular Plant Diversity and Climate Change in the Alpine Zone of the Lefka Ori, Crete. Biodivers. Conserv. 2007, 16, 1603–1615. [Google Scholar] [CrossRef]
- Clarke, C.L.; Edwards, M.E.; Gielly, L.; Ehrich, D.; Hughes, P.D.M.; Morozova, L.M.; Haflidason, H.; Mangerud, J.; Svendsen, J.I.; Alsos, I.G. Persistence of Arctic-Alpine Flora during 24,000 Years of Environmental Change in the Polar Urals. Sci. Rep. 2019, 9, 19613. [Google Scholar] [CrossRef] [Green Version]
- Lamprecht, A.; Semenchuk, P.R.; Steinbauer, K.; Winkler, M.; Pauli, H. Climate Change Leads to Accelerated Transformation of High-Elevation Vegetation in the Central Alps. New Phytol. 2018, 220, 447–459. [Google Scholar] [CrossRef] [PubMed]
- Salick, J.; Fang, Z.; Hart, R. Rapid Changes in Eastern Himalayan Alpine Flora with Climate Change. Am. J. Bot. 2019, 106, 520–530. [Google Scholar] [CrossRef]
- Spasojevic, M.J.; Bowman, W.D.; Humphries, H.C.; Seastedt, T.R.; Suding, K.N. Changes in Alpine Vegetation over 21 Years: Are Patterns across a Heterogeneous Landscape Consistent with Predictions? Ecosphere 2013, 4, art117. [Google Scholar] [CrossRef]
- Huxley, J.D.; Spasojevic, M.J. Area Not Geographic Isolation Mediates Biodiversity Responses of Alpine Refugia to Climate Change. Front. Ecol. Evol. 2021, in press. [Google Scholar] [CrossRef]
- Scharnagl, K.; Johnson, D.; Ebert-May, D. Shrub Expansion and Alpine Plant Community Change: 40-Year Record from Niwot Ridge, Colorado. Plant Ecol. Divers. 2019, 12, 407–416. [Google Scholar] [CrossRef]
- Pauli, H.; Gottfried, M.; Dullinger, S.; Abdaladze, O.; Akhalkatsi, M.; Alonso, J.L.B.; Coldea, G.; Dick, J.; Erschbamer, B.; Calzado, R.F.; et al. Recent Plant Diversity Changes on Europe’s Mountain Summits. Science 2012, 336, 353–355. [Google Scholar] [CrossRef] [Green Version]
- Engler, R.; Randin, C.F.; Thuiller, W.; Dullinger, S.; Zimmermann, N.E.; Araújo, M.B.; Pearman, P.B.; Le Lay, G.; Piedallu, C.; Albert, C.H.; et al. 21st Century Climate Change Threatens Mountain Flora Unequally across Europe: CLIMATE CHANGE IMPACTS ON MOUNTAIN FLORAE. Glob. Chang. Biol. 2011, 17, 2330–2341. [Google Scholar] [CrossRef]
- Lesica, P.; Crone, E.E. Arctic and Boreal Plant Species Decline at Their Southern Range Limits in the Rocky Mountains. Ecol. Lett. 2017, 20, 166–174. [Google Scholar] [CrossRef]
- Compagnoni, A.; Levin, S.; Childs, D.Z.; Harpole, S.; Paniw, M.; Römer, G.; Burns, J.H.; Che-Castaldo, J.; Rüger, N.; Kunstler, G.; et al. Herbaceous Perennial Plants with Short Generation Time Have Stronger Responses to Climate Anomalies than Those with Longer Generation Time. Nat. Commun. 2021, 12, 1824. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.T.; Wadgymar, S.M. Climate Change Disrupts Local Adaptation and Favours Upslope Migration. Ecol. Lett. 2020, 23, 181–192. [Google Scholar] [CrossRef]
- Körner, C. The Use of ‘Altitude’ in Ecological Research. Trends Ecol. Evol. 2007, 22, 569–574. [Google Scholar] [CrossRef]
- Elsen, P.R.; Monahan, W.B.; Merenlender, A.M. Topography and Human Pressure in Mountain Ranges Alter Expected Species Responses to Climate Change. Nat. Commun. 2020, 11, 1974. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.W.; Jackson, S.T. Novel Climates, No-Analog Communities, and Ecological Surprises. Front. Ecol. Evol. 2007, 5, 475–482. [Google Scholar] [CrossRef]
- Inouye, D.W. Effects of Climate Change on Alpine Plants and Their Pollinators. Ann. N. Y. Acad. Sci. 2020, 1469, 26–37. [Google Scholar] [CrossRef] [PubMed]
- Dullinger, S.; Gattringer, A.; Thuiller, W.; Moser, D.; Zimmermann, N.E.; Guisan, A.; Willner, W.; Plutzar, C.; Leitner, M.; Mang, T.; et al. Extinction Debt of High-Mountain Plants under Twenty-First-Century Climate Change. Nat. Clim. Chang. 2012, 2, 619–622. [Google Scholar] [CrossRef]
- Niskanen, A.K.J.; Niittynen, P.; Aalto, J.; Väre, H.; Luoto, M. Lost at High Latitudes: Arctic and Endemic Plants under Threat as Climate Warms. Divers. Distrib. 2019, 25, 809–821. [Google Scholar] [CrossRef] [Green Version]
- Nagy, L.; Grabherr, G. The Biology of Alpine Habitats; The Biology of Habitats Series; Oxford University Press: Oxford, UK; New York, NY, USA, 2009; ISBN 978-0-19-856703-5. [Google Scholar]
- Litaor, M.I.; Williams, M.; Seastedt, T.R. Topographic Controls on Snow Distribution, Soil Moisture, and Species Diversity of Herbaceous Alpine Vegetation, Niwot Ridge, Colorado. J. Geophys. Res. 2008, 113, G000419. [Google Scholar] [CrossRef] [Green Version]
- Winkler, D.E.; Butz, R.J.; Germino, M.J.; Reinhardt, K.; Kueppers, L.M. Snowmelt Timing Regulates Community Composition, Phenology, and Physiological Performance of Alpine Plants. Front. Plant Sci. 2018, 9, 1140. [Google Scholar] [CrossRef] [Green Version]
- Winkler, D.E.; Chapin, K.J.; Kueppers, L.M. Soil Moisture Mediates Alpine Life Form and Community Productivity Responses to Warming. Ecology 2016, 97, 1553–1563. [Google Scholar] [CrossRef] [Green Version]
- Anthelme, F.; Cavieres, L.A.; Dangles, O. Facilitation among Plants in Alpine Environments in the Face of Climate Change. Front. Plant Sci. 2014, 5. [Google Scholar] [CrossRef] [Green Version]
- Bowman, W.D.; Murgel, J.; Blett, T.; Porter, E. Nitrogen Critical Loads for Alpine Vegetation and Soils in Rocky Mountain National Park. J. Environ. Manag. 2012, 103, 165–171. [Google Scholar] [CrossRef]
- Bowman, W.D.; Theodose, T.A.; Schardt, J.C.; Conant, R.T. Constraints of Nutrient Availability on Primary Production in Two Alpine Tundra Communities. Ecology 1993, 74, 2085–2097. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Klein, J.A.; Winkler, D.E.; Peng, A.; Lazarus, B.E.; Germino, M.J.; Suding, K.N.; Smith, J.G.; Kueppers, L.M. Warming of Alpine Tundra Enhances Belowground Production and Shifts Community towards Resource Acquisition Traits. Ecosphere 2020, 11, e03270. [Google Scholar] [CrossRef]
- Dong, Z.; Driscoll, C.T.; Johnson, S.L.; Campbell, J.L.; Pourmokhtarian, A.; Stoner, A.M.K.; Hayhoe, K. Projections of Water, Carbon, and Nitrogen Dynamics under Future Climate Change in an Old-Growth Douglas-Fir Forest in the Western Cascade Range Using a Biogeochemical Model. Sci. Total Environ. 2019, 656, 608–624. [Google Scholar] [CrossRef]
- Knowles, J.F.; Blanken, P.D.; Lawrence, C.R.; Williams, M.W. Evidence for Non-Steady-State Carbon Emissions from Snow-Scoured Alpine Tundra. Nat. Commun. 2019, 10, 1306. [Google Scholar] [CrossRef] [Green Version]
- Morelli, T.L.; Daly, C.; Dobrowski, S.Z.; Dulen, D.M.; Ebersole, J.L.; Jackson, S.T.; Lundquist, J.D.; Millar, C.I.; Maher, S.P.; Monahan, W.B.; et al. Managing Climate Change Refugia for Climate Adaptation. PLoS ONE 2016, 11, e0159909. [Google Scholar] [CrossRef] [Green Version]
- Barrows, C.W.; Ramirez, A.R.; Sweet, L.C.; Morelli, T.L.; Millar, C.I.; Frakes, N.; Rodgers, J.; Mahalovich, M.F. Validating Climate-change Refugia: Empirical Bottom-up Approaches to Support Management Actions. Front. Ecol. Environ. 2020, 18, 298–306. [Google Scholar] [CrossRef]
- Lawler, J.J.; Ackerly, D.D.; Albano, C.M.; Anderson, M.G.; Dobrowski, S.Z.; Gill, J.L.; Heller, N.E.; Pressey, R.L.; Sanderson, E.W.; Weiss, S.B. The Theory behind, and the Challenges of, Conserving Nature’s Stage in a Time of Rapid Change: Conserving Nature’s Stage in a Time of Rapid Change. Conserv. Biol. 2015, 29, 618–629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engler, R.; Randin, C.F.; Vittoz, P.; Czáka, T.; Beniston, M.; Zimmermann, N.E.; Guisan, A. Predicting Future Distributions of Mountain Plants under Climate Change: Does Dispersal Capacity Matter? Ecography 2009, 32, 34–45. [Google Scholar] [CrossRef] [Green Version]
- Saladin, B.; Pellissier, L.; Graham, C.H.; Nobis, M.P.; Salamin, N.; Zimmermann, N.E. Rapid Climate Change Results in Long-Lasting Spatial Homogenization of Phylogenetic Diversity. Nat. Commun. 2020, 11, 4663. [Google Scholar] [CrossRef]
- Gao, M.; Piao, S.; Chen, A.; Yang, H.; Liu, Q.; Fu, Y.H.; Janssens, I.A. Divergent Changes in the Elevational Gradient of Vegetation Activities over the Last 30 Years. Nat. Commun. 2019, 10, 2970. [Google Scholar] [CrossRef]
- Chapin, F.S.; Walker, L.R.; Fastie, C.L.; Sharman, L.C. Mechanisms of Primary Succession Following Deglaciation at Glacier Bay, Alaska. Ecol. Monogr. 1994, 64, 149–175. [Google Scholar] [CrossRef]
- Jurasinski, G.; Kreyling, J. Upward Shift of Alpine Plants Increases Floristic Similarity of Mountain Summits. J. Veg. Sci. 2007, 18, 711–718. [Google Scholar] [CrossRef]
- Steinbauer, M.J.; Grytnes, J.-A.; Jurasinski, G.; Kulonen, A.; Lenoir, J.; Pauli, H.; Rixen, C.; Winkler, M.; Bardy-Durchhalter, M.; Barni, E.; et al. Accelerated Increase in Plant Species Richness on Mountain Summits Is Linked to Warming. Nature 2018, 556, 231–234. [Google Scholar] [CrossRef] [PubMed]
- Bueno de Mesquita, C.P.; Brigham, L.M.; Sommers, P.; Porazinska, D.L.; Farrer, E.C.; Darcy, J.L.; Suding, K.N.; Schmidt, S.K. Evidence for Phosphorus Limitation in High-Elevation Unvegetated Soils, Niwot Ridge, Colorado. Biogeochemistry 2020, 147, 1–13. [Google Scholar] [CrossRef]
- Bueno de Mesquita, C.P.; Sartwell, S.A.; Schmidt, S.K.; Suding, K.N. Growing-season Length and Soil Microbes Influence the Performance of a Generalist Bunchgrass beyond Its Current Range. Ecology 2020. [Google Scholar] [CrossRef]
- Kulonen, A.; Imboden, R.A.; Rixen, C.; Maier, S.B.; Wipf, S. Enough Space in a Warmer World? Microhabitat Diversity and Small-Scale Distribution of Alpine Plants on Mountain Summits. Divers. Distrib. 2018, 24, 252–261. [Google Scholar] [CrossRef] [Green Version]
- Harsch, M.A.; Hulme, P.E.; McGlone, M.S.; Duncan, R.P. Are Treelines Advancing? A Global Meta-Analysis of Treeline Response to Climate Warming. Ecol. Lett. 2009, 12, 1040–1049. [Google Scholar] [CrossRef]
- Capers, R.S.; Stone, A.D. After 33 Years, Trees More Frequent and Shrubs More Abundant in Northeast U.S. Alpine Community. Arct. Antarct. Alp. Res. 2011, 43, 495–502. [Google Scholar] [CrossRef] [Green Version]
- Bowling, D.R.; Logan, B.A.; Hufkens, K.; Aubrecht, D.M.; Richardson, A.D.; Burns, S.P.; Anderegg, W.R.L.; Blanken, P.D.; Eiriksson, D.P. Limitations to Winter and Spring Photosynthesis of a Rocky Mountain Subalpine Forest. Agric. For. Meteorol. 2018, 252, 241–255. [Google Scholar] [CrossRef]
- Wundram, D.; Pape, R.; Löffler, J. Alpine Soil Temperature Variability at Multiple Scales. Arct. Antarct. Alp. Res. 2010, 42, 117–128. [Google Scholar] [CrossRef]
- Garcia, R.A.; Cabeza, M.; Rahbek, C.; Araujo, M.B. Multiple Dimensions of Climate Change and Their Implications for Biodiversity. Science 2014, 344, 1247579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seddon, A.W.R.; Macias-Fauria, M.; Long, P.R.; Benz, D.; Willis, K.J. Sensitivity of Global Terrestrial Ecosystems to Climate Variability. Nature 2016, 531, 229–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laiolo, P.; Dondero, F.; Ciliento, E.; Rolando, A. Consequences of Pastoral Abandonment for the Structure and Diversity of the Alpine Avifauna. J. Appl. Ecol. 2004, 41, 294–304. [Google Scholar] [CrossRef]
- Benedict, J.B. Rates of Tree-Island Migration, Colorado Rocky Mountains, USA. Ecology 1984, 65, 820–823. [Google Scholar] [CrossRef]
- Fitzpatrick, O.T.; Blade, M.; Fishback, L.A.; Kershaw, G.P.; Muffly, J.; Mamet, S.D. Snowpack, Tree Size, and Ecological Legacies Promote Seedling Establishment in Tree Islands at the Treeline. Ecosystems 2020, 23, 1714–1725. [Google Scholar] [CrossRef]
- Jabis, M.D.; Germino, M.J.; Kueppers, L.M. Colonisation of the Alpine Tundra by Trees: Alpine Neighbours Assist Late-Seral but Not Early-Seral Conifer Seedlings. Plant Ecol. Divers. 2020, 13, 209–224. [Google Scholar] [CrossRef]
- Wilson, S.D.; Nilsson, C. Arctic Alpine Vegetation Change over 20 Years. Glob. Chang. Biol. 2009, 15, 1676–1684. [Google Scholar] [CrossRef] [Green Version]
- Fazlioglu, F.; Wan, J.S.H. Warming Matters: Alpine Plant Responses to Experimental Warming. Clim. Chang. 2021, 164, 56. [Google Scholar] [CrossRef]
- Jin, Y.; Xu, J.; He, H.; Li, M.-H.; Tao, Y.; Zhang, Y.; Hu, R.; Gao, X.; Bai, Y.; Wang, H.; et al. The Changbai Alpine Shrub Tundra Will Be Replaced by Herbaceous Tundra under Global Climate Change. Plants 2019, 8, 370. [Google Scholar] [CrossRef] [Green Version]
- Wheeler, J.A.; Hoch, G.; Cortés, A.J.; Sedlacek, J.; Wipf, S.; Rixen, C. Increased Spring Freezing Vulnerability for Alpine Shrubs under Early Snowmelt. Oecologia 2014, 175, 219–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, X.; Liang, E.; Wang, Y.; Babst, F.; Camarero, J.J. Mountain Treelines Climb Slowly despite Rapid Climate Warming. Glob. Ecol. Biogeogr. 2021, 30, 305–315. [Google Scholar] [CrossRef]
- Hansson, A.; Dargusch, P.; Shulmeister, J. A Review of Modern Treeline Migration, the Factors Controlling It and the Implications for Carbon Storage. J. Mt. Sci. 2021, 18, 291–306. [Google Scholar] [CrossRef]
- Dobrowski, S.Z.; Parks, S.A. Climate Change Velocity Underestimates Climate Change Exposure in Mountainous Regions. Nat. Commun. 2016, 7, 12349. [Google Scholar] [CrossRef] [Green Version]
- Andrus, R.A.; Harvey, B.J.; Rodman, K.C.; Hart, S.J.; Veblen, T.T. Moisture Availability Limits Subalpine Tree Establishment. Ecology 2018, 99, 567–575. [Google Scholar] [CrossRef]
- Hufft, R.A.; DePrenger-Levin, M.E.; Levy, R.A.; Islam, M.B. Using Herbarium Specimens to Select Indicator Species for Climate Change Monitoring. Biodivers. Conserv. 2018, 27, 1487–1501. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seastedt, T.R.; Oldfather, M.F. Climate Change, Ecosystem Processes and Biological Diversity Responses in High Elevation Communities. Climate 2021, 9, 87. https://doi.org/10.3390/cli9050087
Seastedt TR, Oldfather MF. Climate Change, Ecosystem Processes and Biological Diversity Responses in High Elevation Communities. Climate. 2021; 9(5):87. https://doi.org/10.3390/cli9050087
Chicago/Turabian StyleSeastedt, Timothy R., and Meagan F. Oldfather. 2021. "Climate Change, Ecosystem Processes and Biological Diversity Responses in High Elevation Communities" Climate 9, no. 5: 87. https://doi.org/10.3390/cli9050087
APA StyleSeastedt, T. R., & Oldfather, M. F. (2021). Climate Change, Ecosystem Processes and Biological Diversity Responses in High Elevation Communities. Climate, 9(5), 87. https://doi.org/10.3390/cli9050087