
The Ricker Competition Model of Two Species: Dynamic Modes and Phase Multistability


Abstract
:1. Introduction
2. Model (1) Study for Local Stability
- A trivial fixed point that corresponds to the extinction of both populations:
- 2.
- Two semitrivial solutions that correspond to the extinction of one of two species:
- 3.
- A nontrivial fixed point corresponding to the sustainable existence of both species in the community
- (Model (4) has four fixed points);
- (The system degenerates and has a non-simple nontrivial solution [73]);
- (The nontrivial fixed point is negative, i.e., species coexistence is not possible). It corresponds to a situation where interspecific competition between species is greater than their self-limitation, i.e., (αδ–βγ < 0). In [34,48,62,68], stable coexistence of two competing populations is shown to be impossible with .
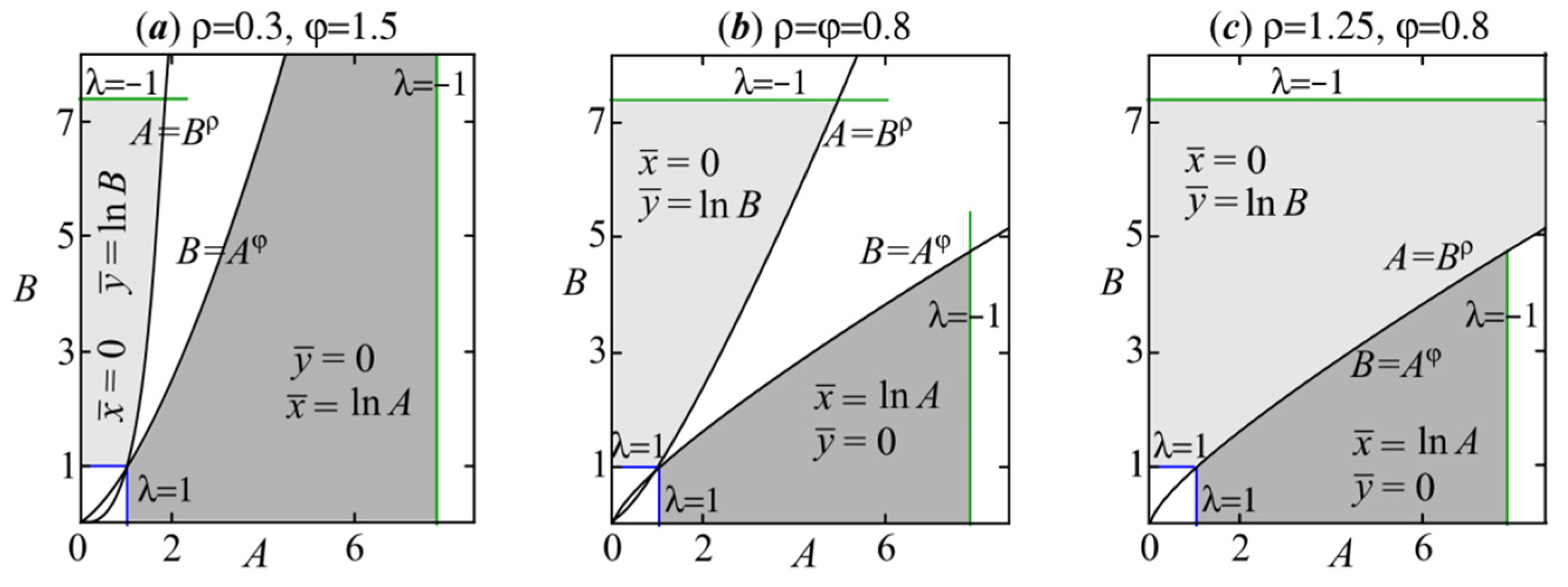
2.1. The Stability Area of Trivial Solution (5)
2.2. The Stability Areas of Semitrivial Solutions
2.3. The Stability Area of Nontrivial Solution with φρ < 1
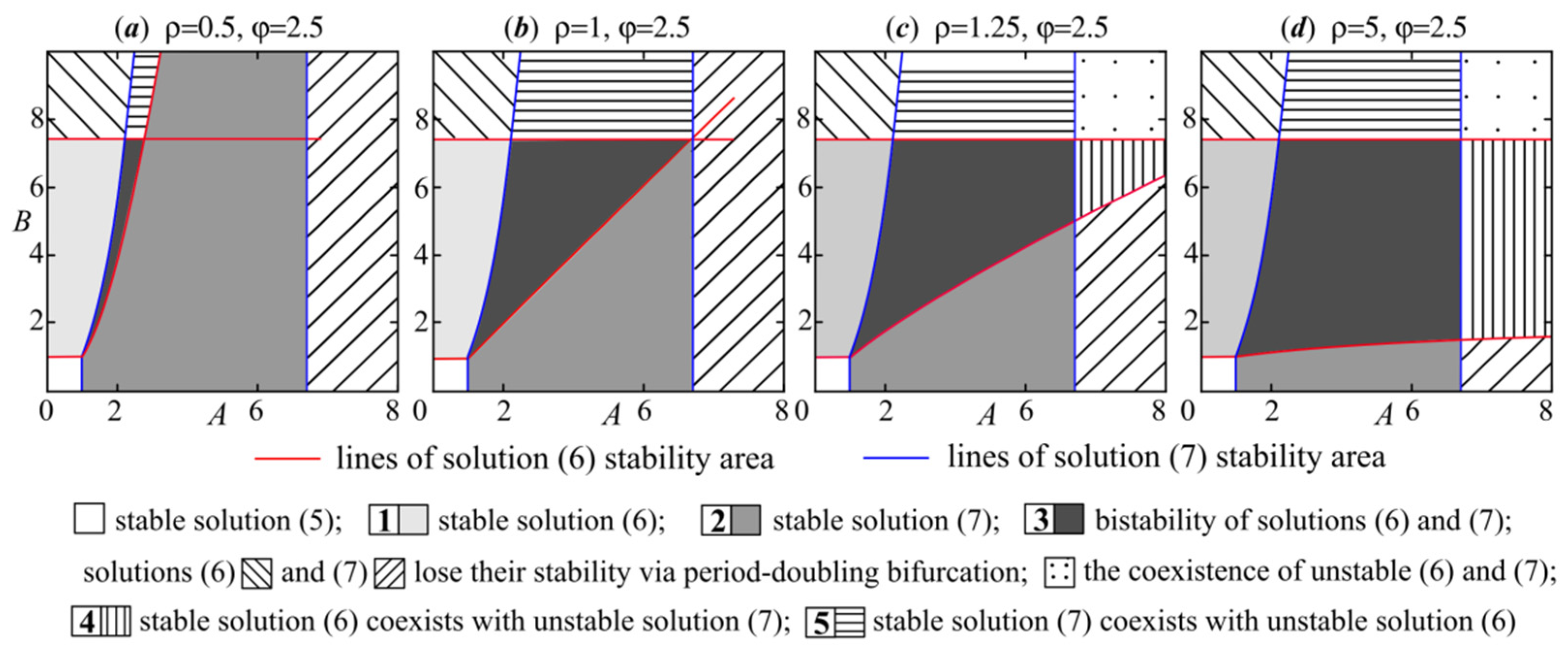
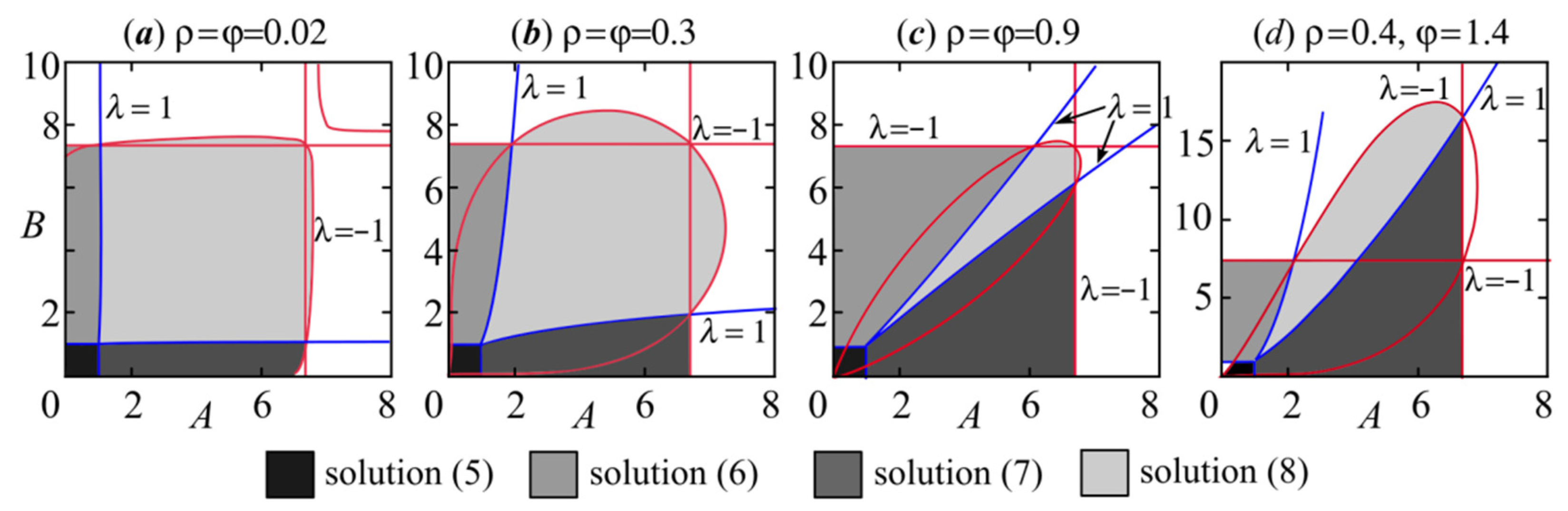
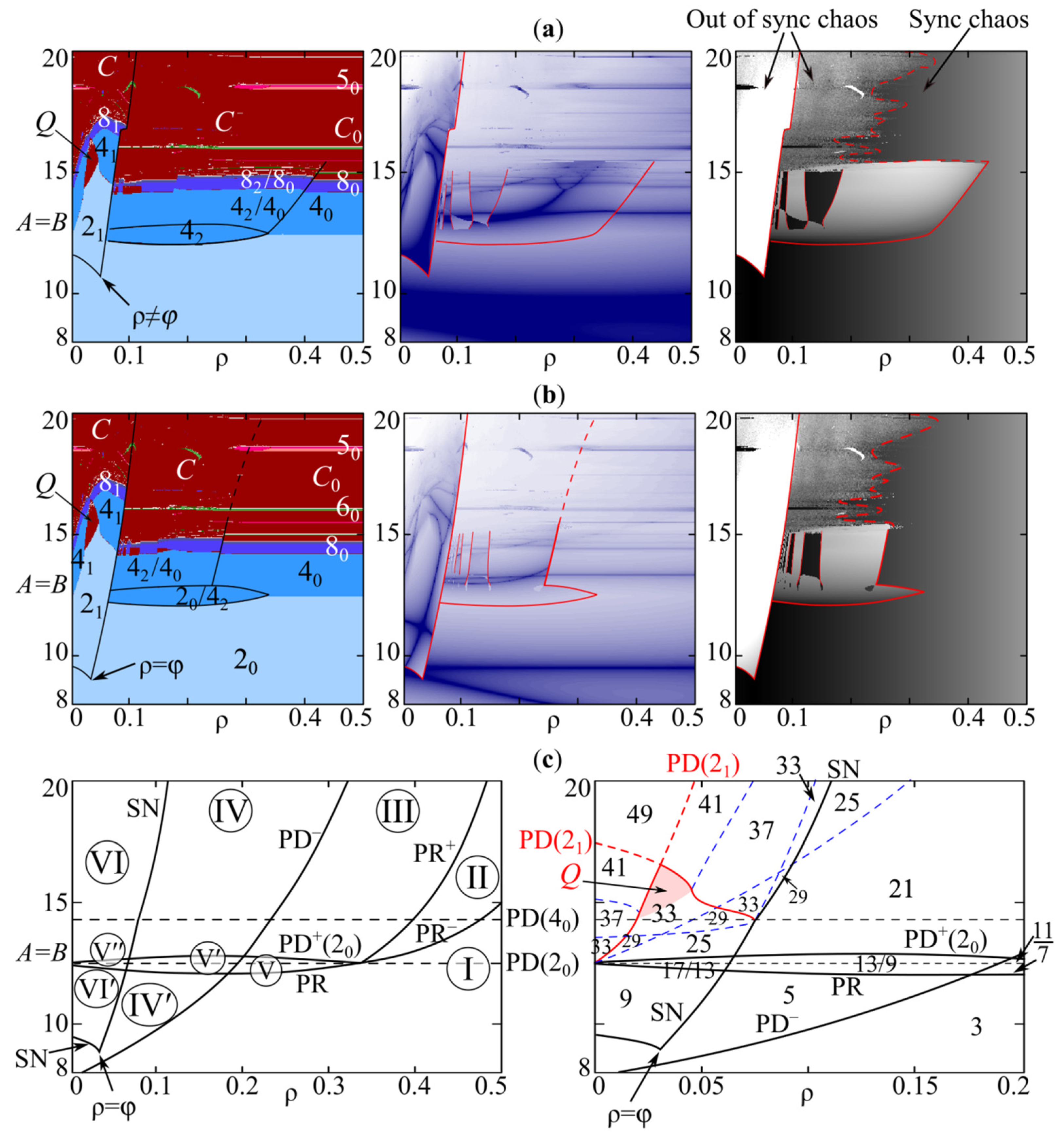
3. Model (4) Dynamics Modes
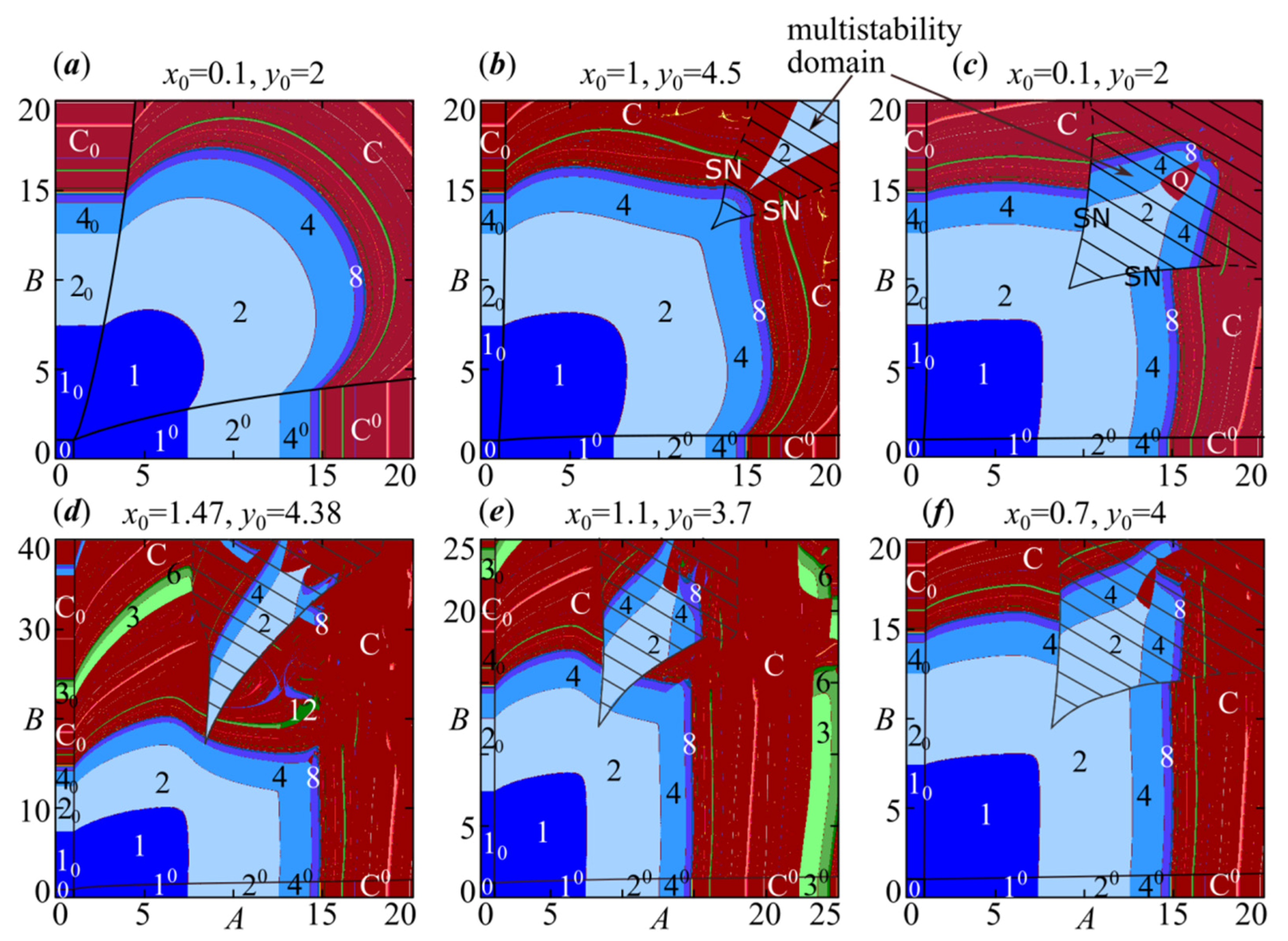
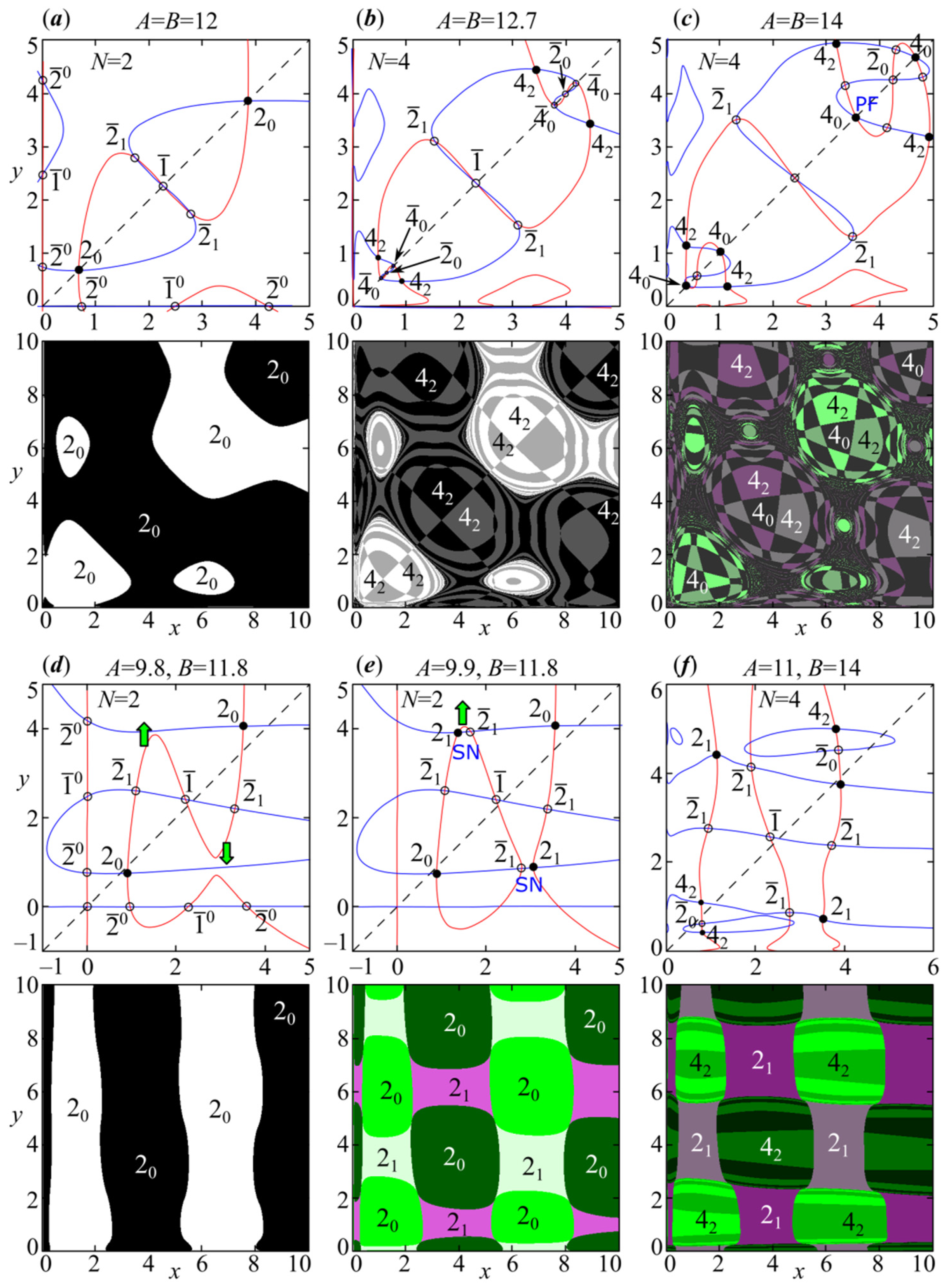
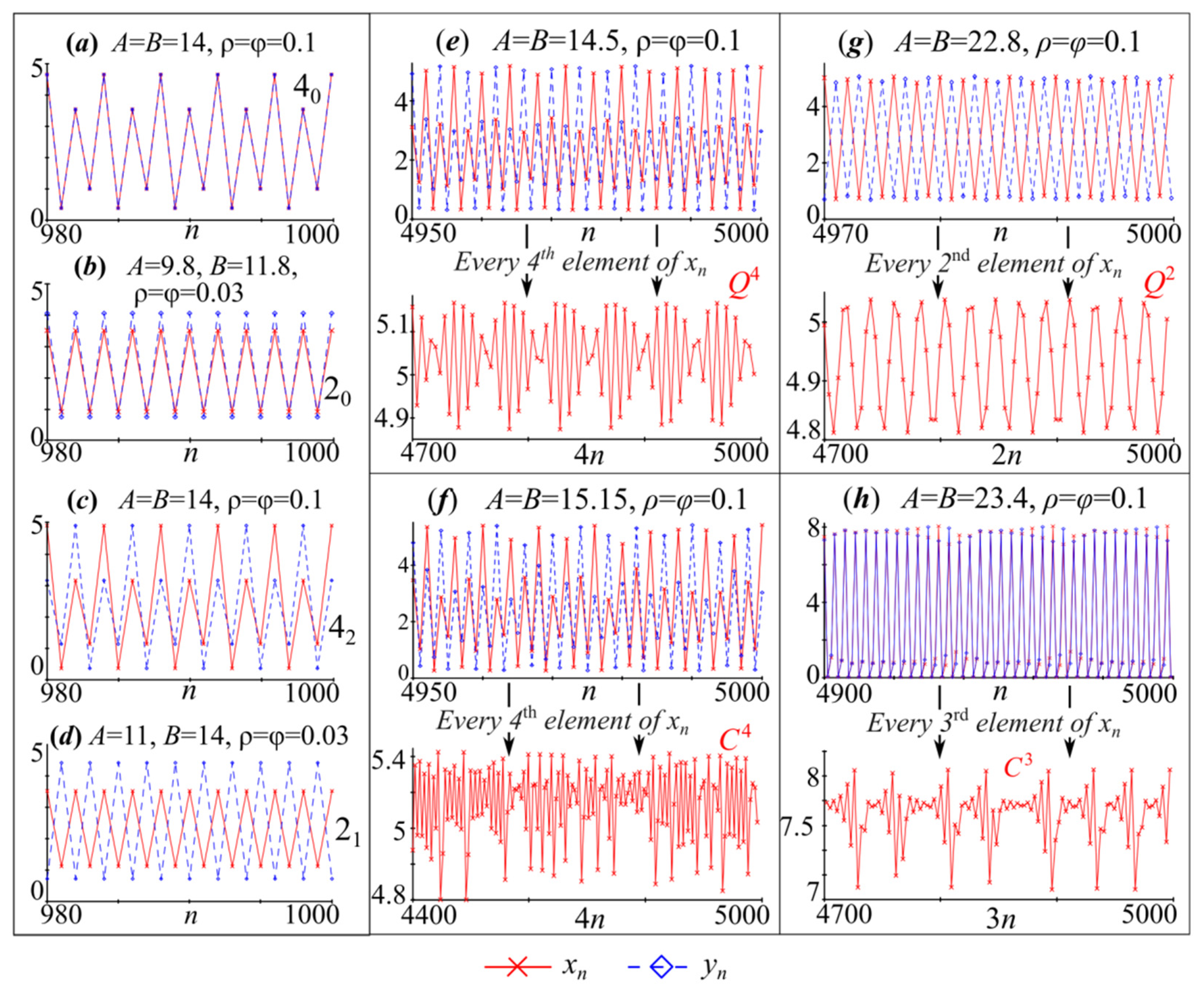
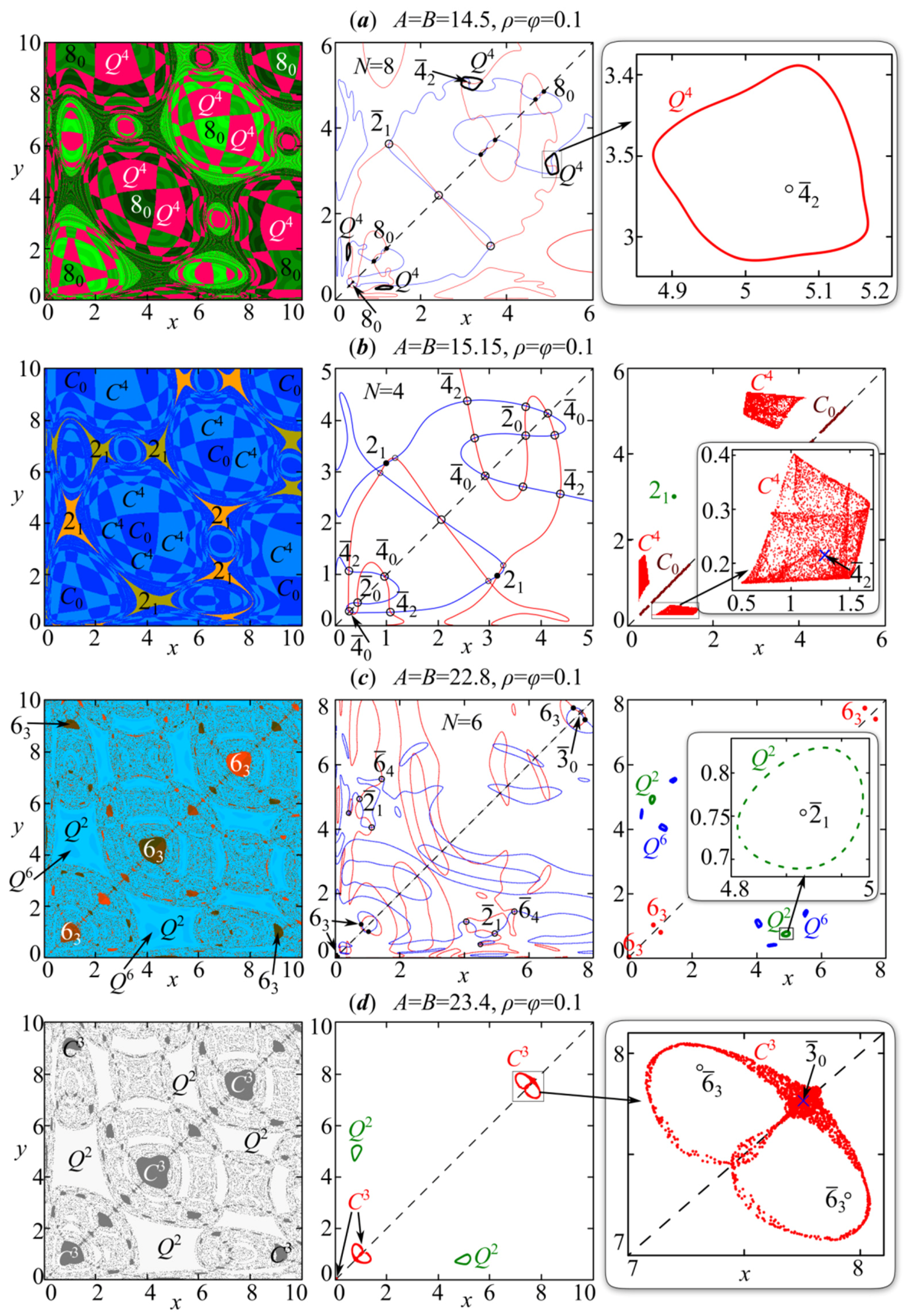
4. Periodic Fixed Points of Model (4): Phase Multistability
4.1. Symmetric Case with and , When Both Species Have the Same Growth Rates and Competition Parameters
4.2. Non-Symmetric Case with or , When Both Species Have Different Growth Rates or Competition Parameters
5. Discussion and Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Crombie, A.C. Interspecific competition. J. Anim. Ecol. 1947, 16, 44–73. [Google Scholar] [CrossRef]
- Gilad, O. Competition and Competition Models. Encyclopedia of Ecology; Jørgensen, S.E., Fath, B.D., Eds.; Academic Press: Cambridge, MA, USA, 2008; pp. 707–712. [Google Scholar] [CrossRef]
- Grant, P.R. Interspecific competition among rodents. Annu. Rev. Ecol. Syst. 1972, 3, 79–106. [Google Scholar] [CrossRef]
- Eccard, J.A.; Ylönen, H. Interspecific competition in small rodents: From populations to individuals. Evol. Ecol. 2003, 17, 423–440. [Google Scholar] [CrossRef]
- Dhondt, A.A. Interspecific Competition in Birds, Vol. 2; Oxford University Press: New York, NY, USA, 2012; p. 296. [Google Scholar] [CrossRef]
- Mainwaring, M.C.; Hartley, I.R. Blue Tits. Encyclopedia of Animal Behavior, 2nd ed.; Choe, J.C., Ed.; Academic Press: Cambridge, MA, USA, 2019; pp. 11–22. [Google Scholar] [CrossRef]
- Goldberg, D.E.; Barton, A.M. Patterns and consequences of interspecific competition in natural communities: A review of field experiments with plants. Am. Nat. 1992, 139, 771–801. [Google Scholar] [CrossRef]
- Hunter, J.; Caro, T. Interspecific competition and predation in American carnivore families. Ethol. Ecol. Evol. 2008, 20, 295–324. [Google Scholar] [CrossRef]
- Caro, T.M.; Stoner, C.J. The potential for interspecific competition among African carnivores. Biol. Conserv. 2003, 110, 67–75. [Google Scholar] [CrossRef]
- Ayala, F.J.; Gilpin, M.E.; Ehrenfeld, J.G. Competition between species: Theoretical models and experimental tests. Theor. Popul. Biol. 1973, 4, 331–356. [Google Scholar] [CrossRef] [Green Version]
- Funakoshi, S. Intraspecific and interspecific competition for larval nests of the caddisflies Stenopsyche marmorata and Stenopsyche sauteri. Entomol. Sci. 2005, 8, 339–345. [Google Scholar] [CrossRef]
- Sommer, U. The role of competition for resources in phytoplankton succession. In Plankton Ecology; Sommer, U., Ed.; Springer: Berlin/Heidelberg, Germany, 1989; pp. 57–106. [Google Scholar] [CrossRef]
- Litchman, E. Resource Competition and the Ecological Success of Phytoplankton. Evol. Prim. Prod. Sea 2007, 351–375. [Google Scholar] [CrossRef]
- Zhao, J.; Ramin, M.; Cheng, V.; Arhonditsis, G.B. Competition patterns among phytoplankton functional groups: How useful are the complex mathematical models? Acta Oecol. 2008, 33, 324–344. [Google Scholar] [CrossRef]
- Miller, R.S. Pattern and process in competition. Adv. Ecol. Res. 1967, 4, 1–74. [Google Scholar] [CrossRef]
- Schoener, T.W. Field experiments on interspecific competition. Am. Nat. 1983, 122, 240–285. [Google Scholar] [CrossRef]
- Saenz, R.; Stein, J.; Yakubu, A.; Jones, L.; Tisch, N. Intraspecific Competition in the Population of Danaus plexippus (L.); Cornell University: Ithaca, NY, USA, 2002. [Google Scholar]
- Schreier, B.M.; Harcourt, A.H.; Coppeto, S.A.; Somi, M.F. Interspecific competition and niche separation in primates: A global analysis. Biotropica 2009, 41, 283–291. [Google Scholar] [CrossRef]
- Salinas-Ramos, V.B.; Ancillotto, L.; Bosso, L.; Sánchez-Cordero, V.; Russo, D. Interspecific competition in bats: State of knowledge and research challenges. Mammal Rev. 2020, 50, 68–81. [Google Scholar] [CrossRef]
- Nicholson, A.J. An outline of the dynamics of animal populations. Aust. J. Zool. 1954, 2, 9–65. [Google Scholar] [CrossRef]
- Varley, G.C.; Gradwell, G.R.; Hassell, M.P. Insect Population Ecology: An Analytical Approach; University of California Press: Berkeley, CA, USA; Los Angeles, CA, USA, 1974. [Google Scholar]
- Hassell, M.P. Density-dependence in single-species populations. J. Anim. Ecol. 1975, 44, 283–295. [Google Scholar] [CrossRef]
- Jones, F.G.W.; Perry, J.N. Modelling populations of cyst-nematodes (Nematoda: Heteroderidae). J. Appl. Ecol. 1978, 15, 349–371. [Google Scholar] [CrossRef]
- Waltman, P. Competition Models in POPULATION Biology; Society for Industrial and Applied Mathematics: Philadelphia, PA, USA, 1983. [Google Scholar]
- Hofbauer, J.; Hutson, V.; Jansen, W. Coexistence for systems governed by difference equations of Lotka-Volterra type. J. Math. Biol. 1987, 25, 553–570. [Google Scholar] [CrossRef]
- Krawcewicz, W.; Rogers, T.D. Perfect harmony: The discrete dynamics of cooperation. J. Math. Biol. 1990, 28, 383–410. [Google Scholar] [CrossRef]
- Huisman, J.; Weissing, F.J. Fundamental unpredictability in multispecies competition. Am. Nat. 2001, 157, 488–494. [Google Scholar] [CrossRef]
- Nishimura, K.; Kishida, O. Coupling of two competitive systems via density dependent migration. Ecol. Res. 2001, 16, 359–368. [Google Scholar] [CrossRef]
- Brännström, Å.; Sumpter, D.J.T. The role of competition and clustering in population dynamics. Proc. R. Soc. B Biol. Sci. 2005, 272, 2065–2072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cushing, J.M.; LeVarge, S. Some discrete competition models and the principle of competitive exclusion. Differ. Equ. Discret. Dyn. Syst. 2005, 10, 283–301. [Google Scholar] [CrossRef] [Green Version]
- Moll, J.D.; Brown, J.S. Competition and coexistence with multiple life-history stages. Am. Nat. 2008, 171, 839–843. [Google Scholar] [CrossRef]
- Łomnicki, A. Scramble and contest competition, unequal resource allocation, and resource monopolization as determinants of population dynamics. Evol. Ecol. Res. 2009, 11, 371–380. [Google Scholar]
- Ackleh, A.S.; Chiquet, R.A. Competitive exclusion in a discrete juvenile–adult model with continuous and seasonal reproduction. J. Differ. Equ. Appl. 2011, 17, 955–975. [Google Scholar] [CrossRef]
- Luis, R.; Elaydi, S.; Oliveira, H. Stability of a Ricker-type competition model and the competitive exclusion principle. J. Biol. Dyn. 2011, 5, 636–660. [Google Scholar] [CrossRef]
- Kang, Y.; Smith, H. Global dynamics of a discrete two-species Lottery-Ricker competition model. J. Biol. Dyn. 2012, 6, 358–376. [Google Scholar] [CrossRef] [Green Version]
- Nedorezov, L.V. The Lotka–Volterra model of competition between two species and Gause’s experiments: Is there any correspondence? Biophysics 2015, 60, 862–863. [Google Scholar] [CrossRef]
- Luís, R.; Rodrigues, E. Local stability in 3D discrete dynamical systems: Application to a Ricker competition model. Discret. Dyn. Nat. Soc. 2017, 2017, 6186354. [Google Scholar] [CrossRef] [Green Version]
- Razzhevaikin, V.N. The multicomponent Gause principle in models of biological communities. Biol. Bull. Rev. 2018, 8, 421–430. [Google Scholar] [CrossRef]
- Dallas, T.; Melbourne, B.; Hastings, A. Community context and dispersal stochasticity drive variation in spatial spread. J. Anim. Ecol. 2020, 89, 2657–2664. [Google Scholar] [CrossRef] [PubMed]
- Mokni, K.; Elaydi, S.; Ch-Chaoui, M.; Eladdadi, A. Discrete evolutionary population models: A new approach. J. Biol. Dyn. 2020, 14, 454–478. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Salceanu, P.L.; Wang, H. Dispersal-driven coexistence in a multiple-patch competition model for zebra and quagga mussels. J. Differ. Equ. Appl. 2022, 28, 183–197. [Google Scholar] [CrossRef]
- Li, G.; Yao, Y. Two-species competition model with chemotaxis: Well-posedness, stability and dynamics. Nonlinearity 2022, 35, 135. [Google Scholar] [CrossRef]
- Mathew, S.M.; Dilip, D.S. Dynamics of interspecific k species competition model. J. Interdiscip. Math. 2022. [Google Scholar] [CrossRef]
- Ricker, W.E. Stock and recruitment. J. Fish. Board Can. 1954, 5, 559–623. [Google Scholar] [CrossRef]
- Shapiro, A.P. On the Question of Cycles in Return Sequences. In Management and Information: Vol. 3; Far Eastern Scientific Center of the Academy of Sciences of the USSR: Vladivostok, Russia, 1972; pp. 96–118. (In Russian) [Google Scholar]
- May, R.M. Biological populations with non-overlapping generations: Stable points, stable cycles, and chaos. Science 1974, 186, 645–647. [Google Scholar] [CrossRef] [Green Version]
- May, R.M. Biological population obeying difference equations: Stable points, stable cycles, and chaos. J. Theor. Biol. 1975, 51, 511–524. [Google Scholar] [CrossRef]
- Shapiro, A.P.; Luppov, S.P. Recurrent Equations in the Theory of Population Biology; Nauka: Moscow, Russia, 1983; 134p. (In Russian) [Google Scholar]
- Frisman, E.Y.; Zhdanova, O.L.; Kulakov, M.P.; Neverova, G.P.; Revutskaya, O.L. Mathematical Modeling of Population Dynamics Based on Recurrent Equations: Results and Prospects. Part I. Biol. Bull. 2021, 48, 1–15. [Google Scholar] [CrossRef]
- Oppo, G.-L.; Kapral, R. Discrete models for the formation and evolution of spatial structure in dissipative systems. Phys. Rev. A 1986, 33, 4219. [Google Scholar] [CrossRef] [PubMed]
- Kuznetsov, A.P.; Kuznetsov, S.P. Critical dynamics of lattices of coupled mappings at the threshold of chaos. Izv. Vyssh. Uchebn. Zaved. Radiofiz. 1991, 34, 1079–1115. [Google Scholar]
- Gyllenberg, M.; Hanski, I. Single-species metapopulation dynamics: A structured model. Theor. Popul. Biol. 1992, 42, 35–61. [Google Scholar] [CrossRef]
- Gyllenberg, M.; Söderbacka, G.; Ericson, S. Does migration stabilize local population dynamics? Analysis of a discrete matapopulation model. Math. Biosci. 1993, 118, 25–49. [Google Scholar] [CrossRef]
- Hanski, I.; Gyllenberg, M. Two general metapopulation models and the core-satellite species hypothesis. Am. Nat. 1993, 142, 17–41. [Google Scholar] [CrossRef]
- Bezruchko, B.P.; Prokhorov, M.D.; Seleznev, Y.P. Oscillation types, multistability, and basins of attractors in symmetrically coupled period-doubling systems. Chaos Solitons Fractals 2003, 15, 695–711. [Google Scholar] [CrossRef]
- Astakhov, V.V.; Shabunin, A.V.; Stal’makhov, P.A. Bifurcation mechanisms of destruction of antiphase synchronization of chaos in coupled systems with discrete time. Izv. Vyssh. Uchebn. Zaved. Prikl. Nelineinaya Din. 2006, 14, 100–111. (In Russian) [Google Scholar]
- Kulakov, M.P.; Frisman, E.Y. Synchronization of 2-cycles in a system of symmetrically connected populations in which the stock-recruitment is described by the Ricker function. Izv. Vyssh. Uchebn. Zaved. Prikl. Nelineinaya Din. 2010, 18, 25–41. (In Russian) [Google Scholar]
- Frisman, E.Y.; Zhdanova, O.L.; Kulakov, M.P.; Neverova, G.P.; Revutskaya, O.L. Mathematical Modeling of Population Dynamics Based on Recurrent Equations: Results and Prospects. Part II. Biol. Bull. 2021, 48, 239–250. [Google Scholar] [CrossRef]
- Shapiro, A.P. Some mathematical models of food competition between two fish populations and the Gause law. Probl. Cybern. 1972, 25, 161–166. (In Russian) [Google Scholar]
- Shapiro, A.P. Discrete model of competition between two populations. DAN USSR 1974, 218, 699–701. (In Russian) [Google Scholar]
- Liu, P.; Elaydi, S.N. Discrete competitive and cooperative models of Lotka–Volterra type. J. Comput. Anal. Appl. 2001, 3, 53–73. [Google Scholar] [CrossRef]
- Gromova, N.P. Equilibrium and oscillatory limiting regimes in models of two competing populations with discrete time. Math. Res. Popul. Ecol. 1988, 9, 107–116. (In Russian) [Google Scholar]
- Cabral Balreira, E.; Elaydi, S.; Luis, R. Local stability implies global stability for the planar Ricker competition model. Discret. Contin. Dyn. Syst.-B 2014, 19, 323–351. [Google Scholar] [CrossRef]
- Hassell, M.P.; Comins, H.N. Discrete time models for two-species competition. Theor. Popul. Biol. 1976, 9, 202–221. [Google Scholar] [CrossRef]
- Cushing, J.M.; Levarge, S.; Chitnis, N.; Henson, S.M. Some discrete competition models and the competitive exclusion principle. J. Differ. Equ. Appl. 2004, 10, 1139–1151. [Google Scholar] [CrossRef] [Green Version]
- Cushing, J.M.; Henson, S.M.; Blackburn, C.C. Multiple mixed-type attractors in a competition model. J. Biol. Dyn. 2007, 1, 347–362. [Google Scholar] [CrossRef]
- Ackleh, A.S.; Salceanu, P.L. Competitive exclusion and coexistence in an n-species Ricker model. J. Biol. Dyn. 2015, 9, 321–331. [Google Scholar] [CrossRef] [Green Version]
- Skaletskaya, E.I.; Frisman, E.Y.; Shapiro, A.P. Discrete Models of Population Dynamics and Fishery Optimization; Nauka: Moscow, Russia, 1979; 165p. [Google Scholar]
- Jiang, H.; Rogers, T.D. The discrete dynamics of symmetric competition in the plane. J. Math. Biol. 1987, 25, 573–596. [Google Scholar] [CrossRef]
- Smith, H.L. Planar competitive and cooperative difference equations. J. Differ. Equ. Appl. 1998, 3, 335–357. [Google Scholar] [CrossRef]
- Franke, J.E.; Yakubu, A.A. Exclusionary population dynamics in size-structured, discrete competitive systems. J. Differ. Equ. Appl. 1999, 5, 235–249. [Google Scholar] [CrossRef]
- Clark, D.; Kulenović, M.R.S.; Selgrade, J.F. Global asymptotic behavior of a two-dimensional difference equation modelling competition. Nonlinear Anal. Theory Methods Appl. 2003, 52, 1765–1776. [Google Scholar] [CrossRef]
- Arrowsmith, D.K.; Place, C.M. Ordinary Differential Equations: A Qualitative Approach with Applications; Chapman and Hall: London, UK, 1982; 252p. [Google Scholar]
- Elaydi, S. Discrete Chaos: With Applications in Science and Engineering, 2nd ed.; Chapman and Hall/CRC: New York, NY, USA, 2007; 440p. [Google Scholar] [CrossRef]
- Gause, G.F. Experimental demonstration of Volterra’s periodic oscillations in the numbers of animals. J. Exp. Biol. 1935, 12, 44–48. [Google Scholar] [CrossRef]
- Hutchinson, G.E. The paradox of the plankton. Am. Nat. 1961, 18695, 137–145. [Google Scholar] [CrossRef] [Green Version]
- Roy, S.; Chattopadhyay, J. Towards a resolution of ‘the paradox of the plankton’: A brief overview of the proposed mechanisms. Ecol. Complex. 2007, 4, 26–33. [Google Scholar] [CrossRef]
- Lynch, S. Dynamical Systems with Applications using Python; Springer International Publishing: Cham, Switzerland, 2018. [Google Scholar]
- Kuznetsov, Y.A. Elements of Applied Bifurcation Theory, 3rd ed.; Applied Mathematical Sciences; Springer: Berlin/Heidelberg, Germany, 2004; Volume 112. [Google Scholar]
- Kuznetsov, A.P.; Savin, A.V.; Sedova, Y.V.; Tyuryukin, L.V. Bifurcations of Maps; Publishing Center “Science”: Saratov, Russia, 2012. (In Russian) [Google Scholar]
- Lloyd, A.L. The coupled logistic map: A simple model for the effects of spatial heterogeneity on population dynamics. J. Theor. Biol. 1995, 173, 217–230. [Google Scholar] [CrossRef] [Green Version]
- Udwadia, F.E.; Raju, N. Dynamics of coupled nonlinear maps and its application to ecological modeling. Appl. Math. Comput. 1997, 82, 137–179. [Google Scholar] [CrossRef]
- Kulakov, M.P.; Aksenovich, T.I.; Frisman, E.Y. Approaches to the description of disturbances in the dynamics of dynamically related populations: Analysis of disturbance cycles. Reg. Probl. 2013, 16, 5–14. (In Russian) [Google Scholar]
- Kuryzhov, E.; Karatetskaia, E.; Mints, D. Lorenz- and Shilnikov-Shape Attractors in the Model of Two Coupled Parabola Maps. Rus. J. Nonlin. Dyn 2021, 17, 165–174. [Google Scholar] [CrossRef]
- Maistrenko, Y.L.; Maistrenko, V.L.; Popovich, A.; Mosekilde, E. Transverse instability and riddled basins in a system of two coupled logistic maps. Phys. Rev. E 1998, 57, 2713. [Google Scholar] [CrossRef] [Green Version]
- Medvinsky, A.B.; Adamovich, B.V.; Rusakov, A.V.; Tikhonov, D.A.; Nurieva, N.I.; Tereshko, V.M. Population dynamics: Mathematical modeling and reality. Biophysics 2019, 64, 956–977. [Google Scholar] [CrossRef]
- Neverova, G.P.; Kulakov, M.P.; Frisman, E.Y. Changes in population dynamics regimes as result of both multistability and climatic fluctuation. Nonlinear Dyn. 2019, 97, 107–122. [Google Scholar] [CrossRef]
- Neverova, G.P.; Zhdanova, O.L.; Ghosh, B.; Frisman, E.Y. Dynamics of a discrete-time stage-structured predator–prey system with Holling type II response function. Nonlinear Dyn. 2019, 98, 427–446. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Intervals for the Parameter Values | Development Scenario | |
|---|---|---|
| species y displaces species x | ||
| species x displaces species y | ||
| species x and y coexist | ||
| species y displaces species x | ||
| species x displaces species y | ||
| the displacement of a species by another one depends on the values of initial conditions | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kulakov, M.; Neverova, G.; Frisman, E. The Ricker Competition Model of Two Species: Dynamic Modes and Phase Multistability. Mathematics 2022, 10, 1076. https://doi.org/10.3390/math10071076
Kulakov M, Neverova G, Frisman E. The Ricker Competition Model of Two Species: Dynamic Modes and Phase Multistability. Mathematics. 2022; 10(7):1076. https://doi.org/10.3390/math10071076
Chicago/Turabian StyleKulakov, Matvey, Galina Neverova, and Efim Frisman. 2022. "The Ricker Competition Model of Two Species: Dynamic Modes and Phase Multistability" Mathematics 10, no. 7: 1076. https://doi.org/10.3390/math10071076
APA StyleKulakov, M., Neverova, G., & Frisman, E. (2022). The Ricker Competition Model of Two Species: Dynamic Modes and Phase Multistability. Mathematics, 10(7), 1076. https://doi.org/10.3390/math10071076