

Mathematical Model Predicting the Kinetics of Intracellular LCMV Replication

 , ,
, ,  , , and
, , and 
Abstract
:1. Introduction
1.1. Molecular and Genome Structure of LCMV
1.2. Intracellular Replication of LCMV
2. Experimental Data and Methods
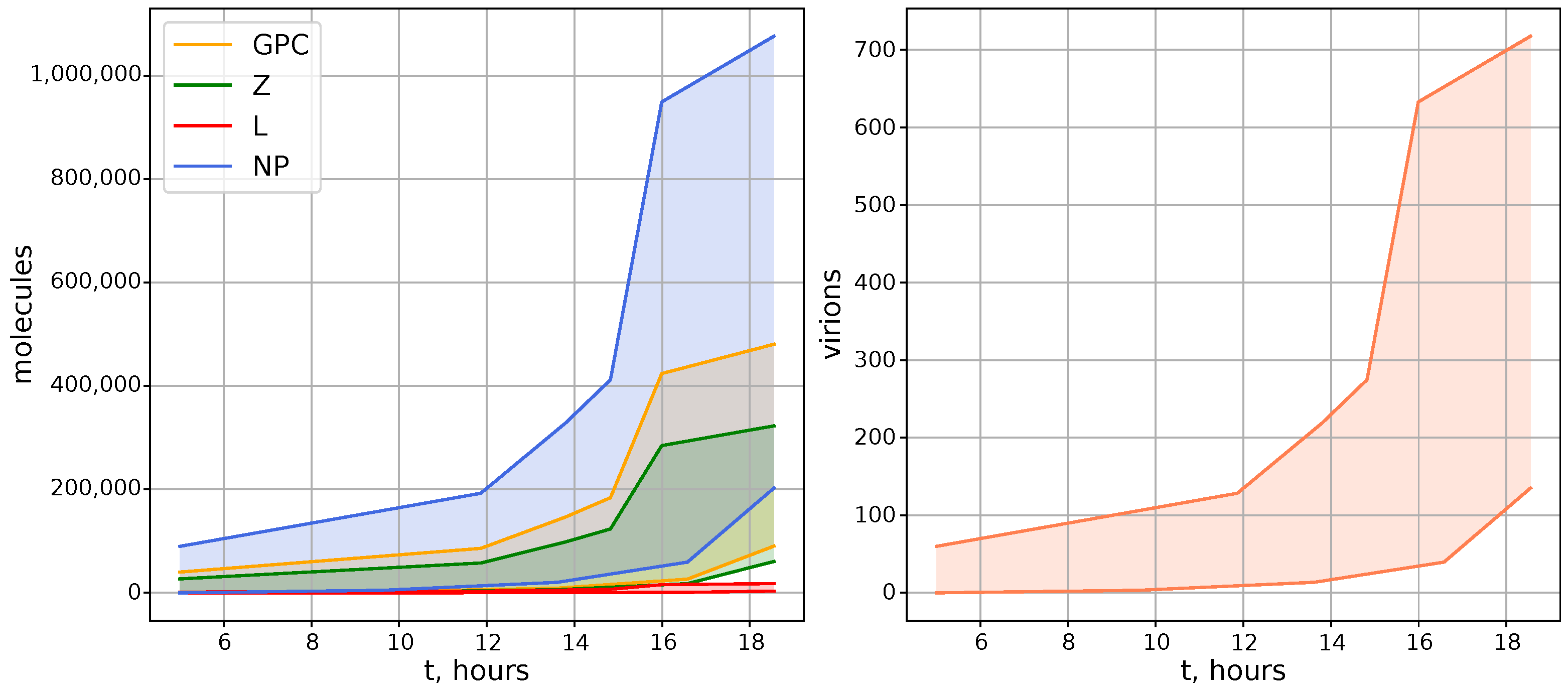
2.1. Kinetics of Virion Components in the Cell
- At all time points, the number of S RNA was significantly larger than L RNA. During the first hours of infection, S RNA was observed in large numbers, whereas L RNA abundance was characterized by undetectable numbers;
- NP mRNA reaches peak concentration in the early hours of cell infection;
- Z mRNA and Z protein concentrations increase at the end of the virus life cycle.
- At the end of the life cycle, the expected number of LCMV virions is estimated to be about several hundreds;
- The ratio of protein components should be as follows: NP > GPC > Z > L;
- The LCMV life cycle lasts from 20 to 40 h.
2.2. Z Protein-Mediated Inhibition Kinetics
2.3. Basic Computational Tools
2.4. Stochastic Modelling Algorithm
2.5. Sensitivity Analysis
3. Mathematical Model
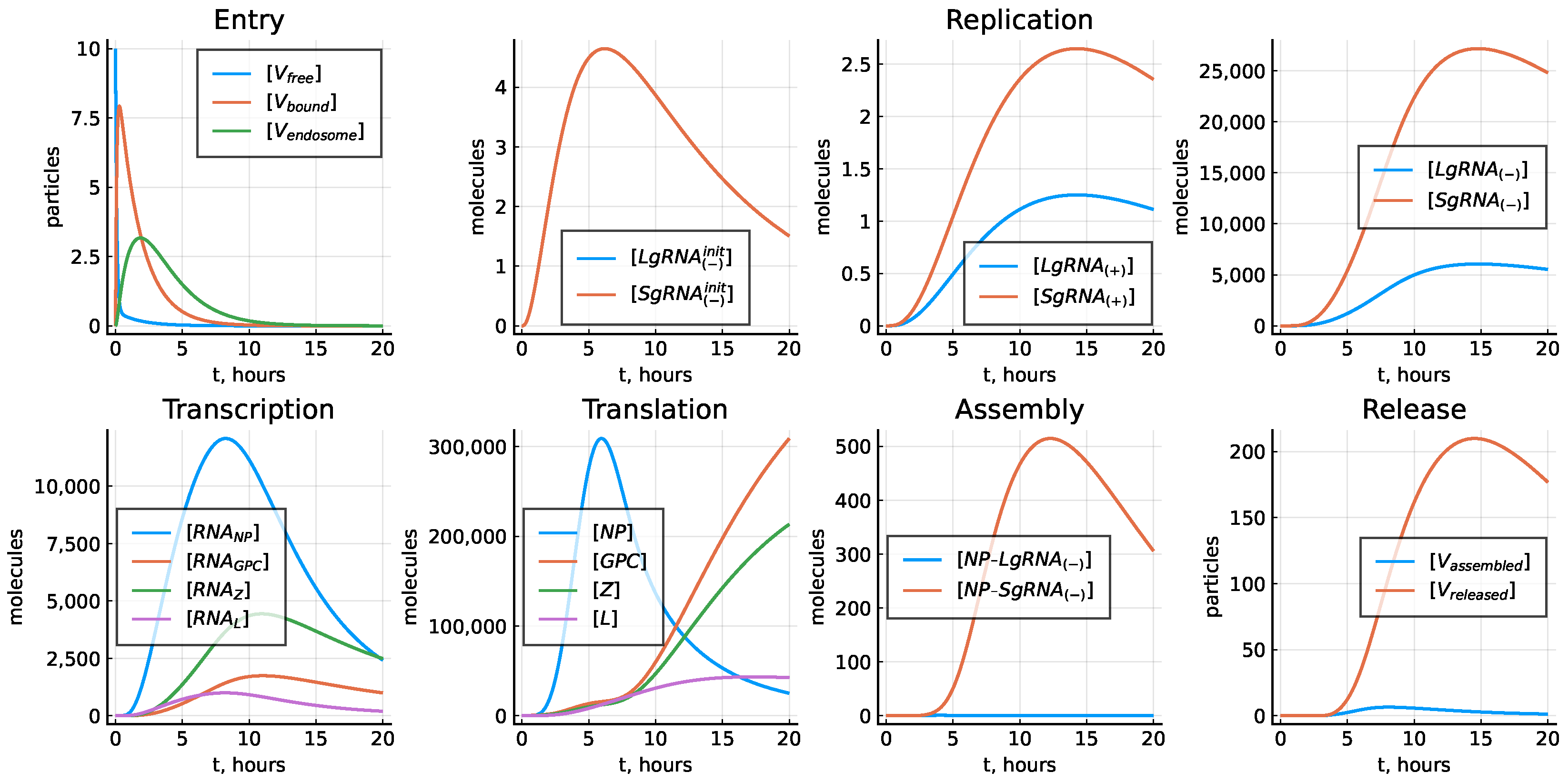
3.1. Deterministic Model of Intracellular LCMV Replication

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Free virions outside the cell membrane | |
| Virions bound to -DG receptor | |
| Virions in endosomes | |
| L negative genomic RNAs, released from virions, that infect the cell | |
| S negative genomic RNAs, released from virions, that infect the cell | |
| L positive genomic RNAs | |
| S positive genomic RNAs | |
| L negative genomic RNAs | |
| S negative genomic RNAs | |
| NP RNAs | |
| GPC RNAs | |
| Z RNAs | |
| L RNAs | |
| NP proteins | |
| Z proteins | |
| L proteins | |
| GPC proteins | |
| coated with NP | |
| coated with NP | |
| Assembled virions in endosomes | |
| Virions released via exocytosis |
3.2. Cell Entry
3.3. Replication of Genomic RNAs
3.4. Transcription
3.5. Translation
3.6. Assembly and Release of Virions
3.7. Calibration of LCMV Replication Model
| Rate of virion binding to -DG receptor, | 10 | [48] | |
| Clearance rate of extracellular virions, | 0.1 | [48] | |
| Dissociation rate constant of bound virions, | 0.51 | [48] | |
| Fusion rate constant, | 0.52 | [48] | |
| Uncoating rate constant, | 0.49 | [48] | |
| Degradation rate of virions in endosomes, | 0.05 | [48] | |
| Degradation rate of negative sense L RNAs in cell, | 0.1 | [48] | |
| Degradation rate of negative sense S RNAs in cell, | 0.1 | [48] | |
| Degradation rate of positive sense L RNAs in cell, | 0.1 | [48] | |
| Degradation rate of positive sense S RNAs in cell, | 0.1 | [48] | |
| Replication rate of positive sense RNAs, | 340 | [48] | |
| Replication rate of negative sense RNAs, | [48] | ||
| Rate of the nucleocapsid formation | 0.3 | [48] | |
| Threshold number of NP proteins at which nucleocapsid formation slows down, molecules | [48] | ||
| Threshold number of Z proteins at which assembly slows down, molecules | 450 | [40] | |
| Threshold number of L proteins at which assembly slows down, molecules | 25 | [40] | |
| Threshold number of GPC proteins at which assembly slows down, molecules | 670 | [40] | |
| Threshold number of RNA enhancing RNA transcription and replication, molecules | 20 | [40] | |
| Threshold number of Z molecules for half-maximal inhibition of L, molecules | [20] | ||
| Transcription rate of RNAs, copies | adjusted | ||
| Transcription rate of NP RNAs, copies | adjusted | ||
| Degradation rate of NP RNA in cell, | 0.31 | [48] | |
| Degradation rate of Z RNA in cell, | 0.3 | [48] | |
| Degradation rate of L RNA in cell, | 0.32 | [48] | |
| Degradation rate of GPC RNA in cell, | 0.29 | [48] | |
| Rate of virion assembly, | 1 | [48] | |
| Translation rate, nt/mRNA | [48] | ||
| Translation rate of Z, nt/mRNA | 2250 | [48] | |
| Degradation rate of NP protein in cell, | 0.021 | [48] | |
| Degradation rate of Z protein in cell, | 0.03 | [48] | |
| Degradation rate of L protein in cell, | 0.04 | [48] | |
| Degradation rate of GPC protein in cell, | 0.022 | [48] | |
| Degradation rate of ribonucleoprotein of , | 0.2 | [48] | |
| Degradation rate of ribonucleoprotein of , | 0.2 | [48] | |
| Rate of virion release via exocytosis, | 7 | [48] | |
| Assembled virion degradation rate, | 0.07 | [48] | |
| Number of NP protein per virion, molecules | 1500 | [40] | |
| Number of Z protein per virion, molecules | 450 | [40] | |
| Number of L protein per virion, molecules | 25 | [40] | |
| Number of GPC protein per virion, molecules | 670 | [40] | |
| Length of RNA genome coding NP protein, nt | 1674 | [50] | |
| Length of RNA genome coding Z protein, nt | 270 | [50] | |
| Length of RNA genome coding L protein, nt | 6630 | [50] | |
| Length of RNA genome coding GPC protein, nt | 1494 | [50] | |
| Length of S segment of genomic RNA, nt | 3400 | [50] | |
| Length of L segment of genomic RNA, nt | 7200 | [50] | |
| Number of Z proteins, initially released to the cell from , molecules | 450 | [40] | |
| Number of L proteins, initially released to the cell from , molecules | 25 | [40] | |
| Number of GPC proteins, initially released to the cell from , molecules | 670 | [40] |
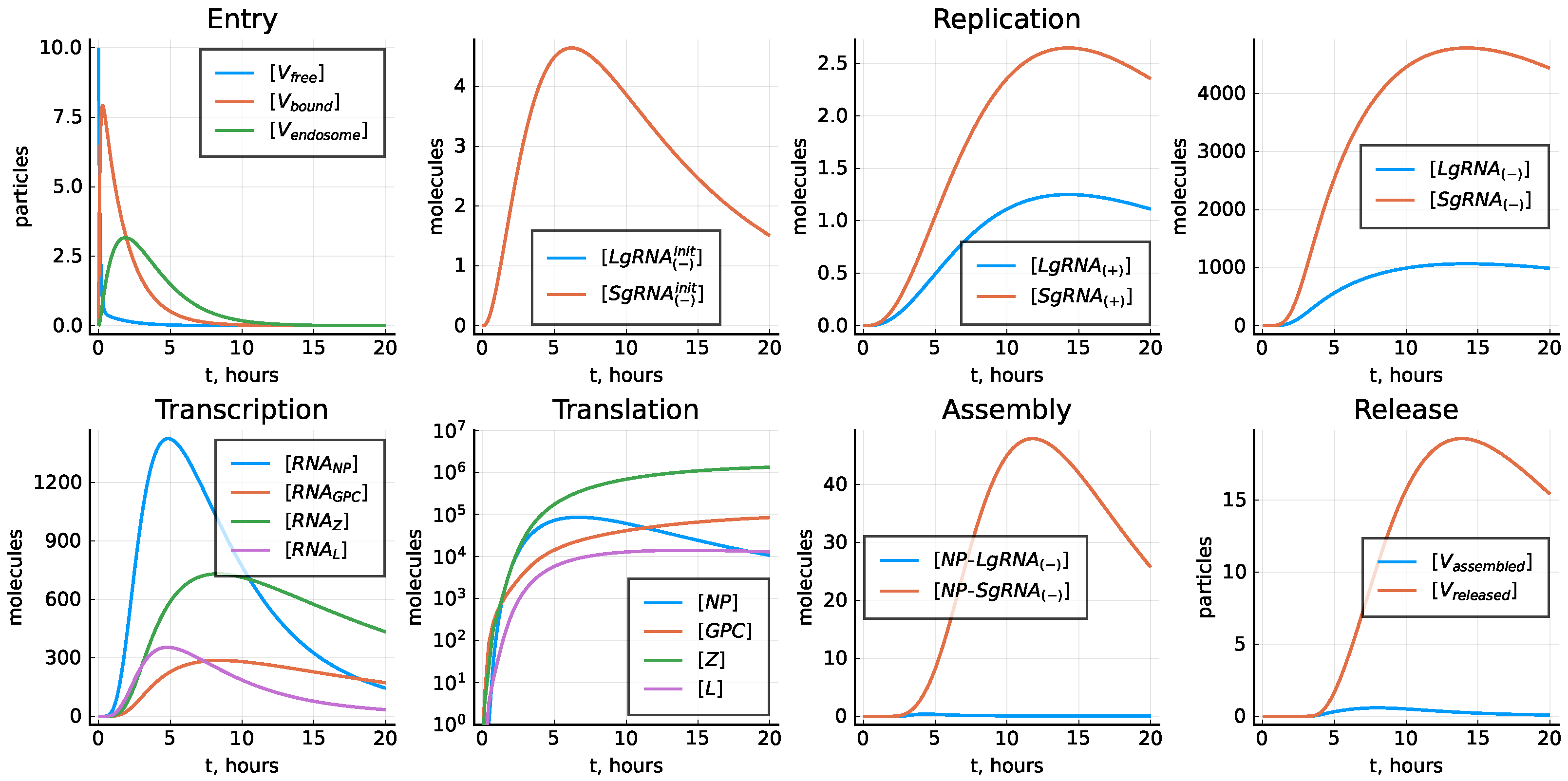
3.8. Stochastic Model
4. Results
4.1. Implications of the Model Calibration Uncertainty
4.2. Sensitivity Analysis of the Deterministic Model
- Translation (negative effect for Z, positive for the rest);
- Transcription (positive effect for NP, negative for the rest);
- Degradation of NP mRNA (negative effect);
- Degradation of free virions (negative effect);
- Fusion with endosomal membrane (positive effect);
- Unpacking (positive effect);
- Replication (positive effect for (+)RNA replication, negative effect for (-)RNA).
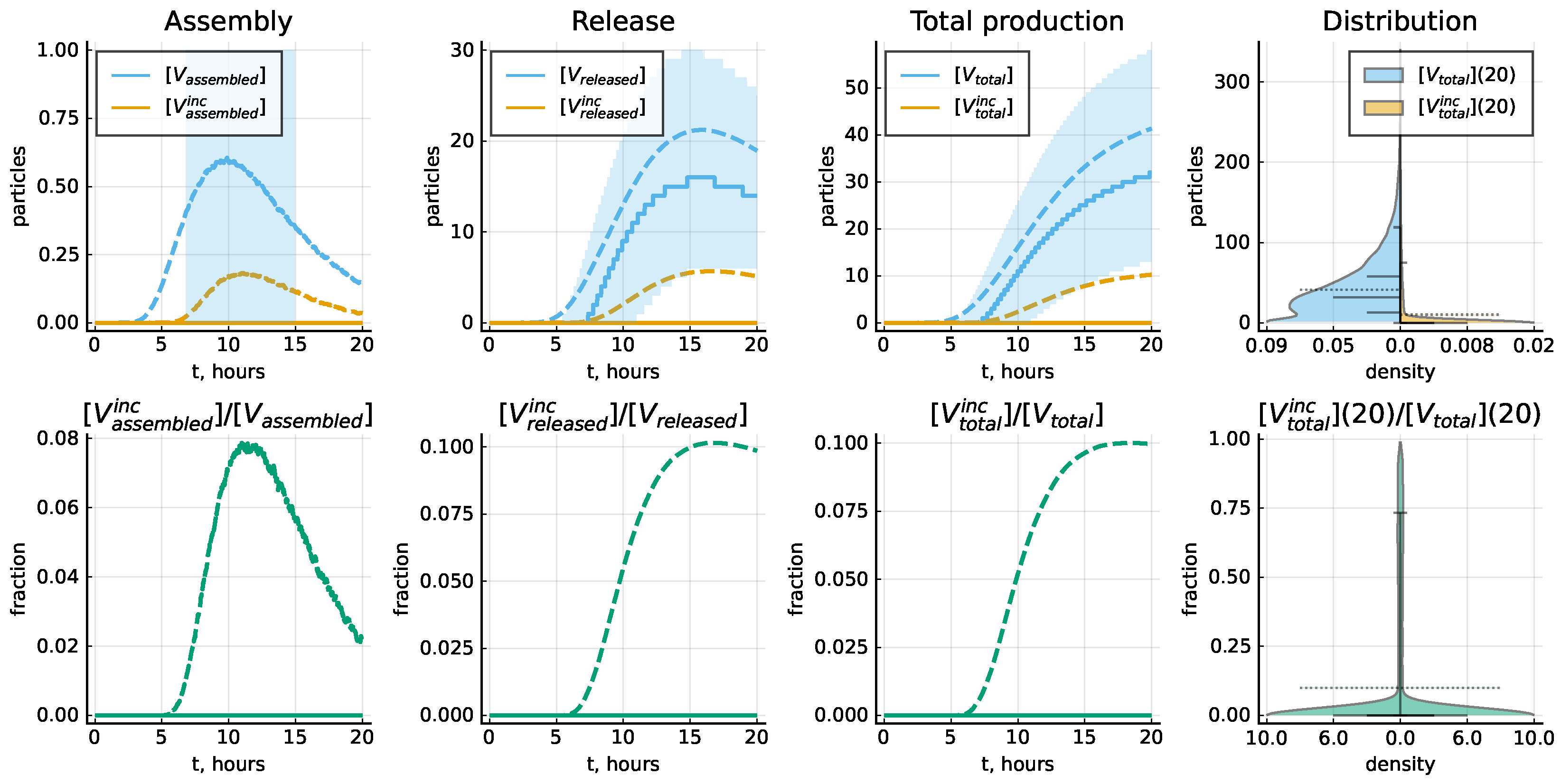
4.3. Kinetic Variability of the LCMV Life Cycle
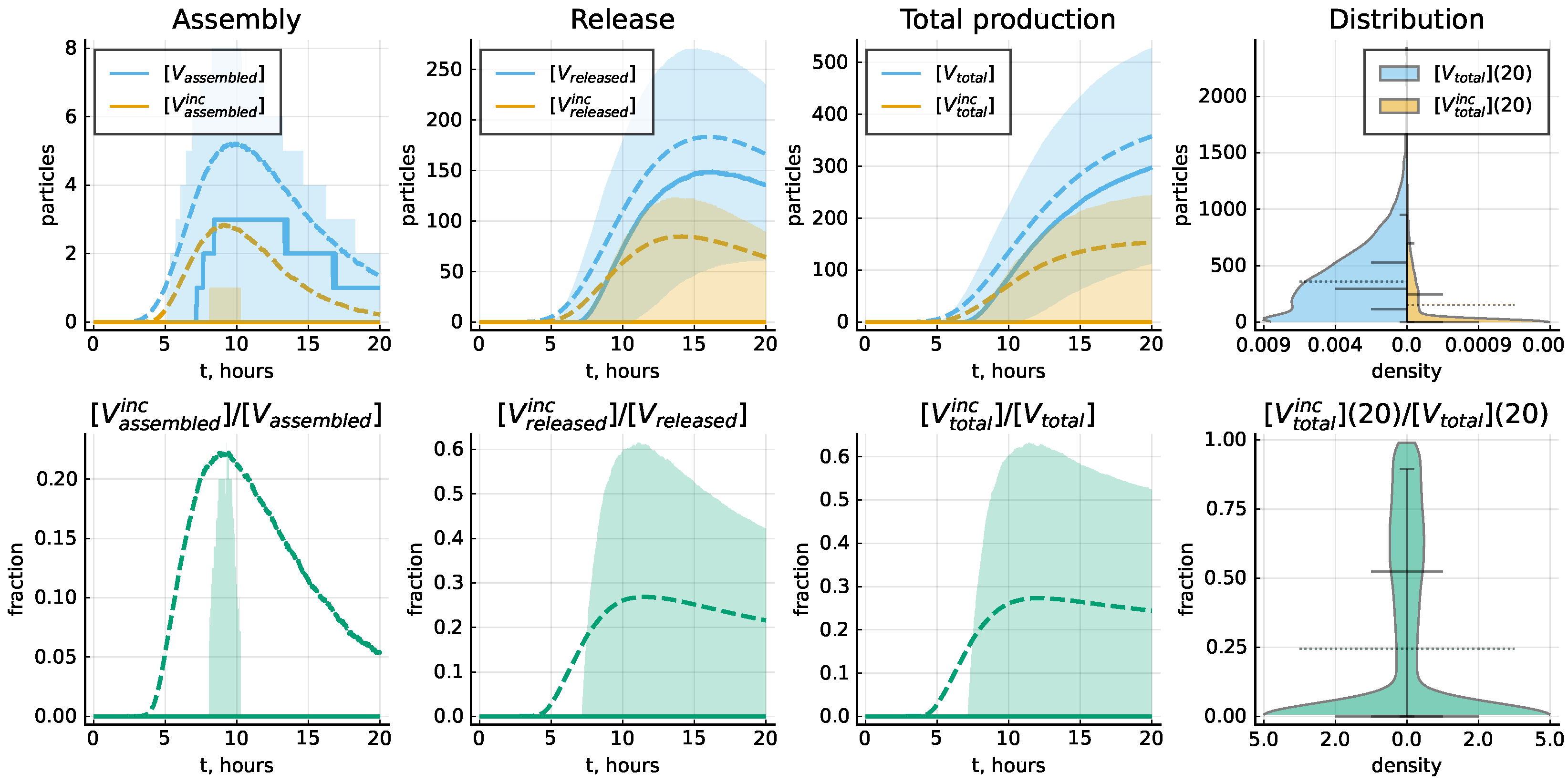
4.4. Probability of Productive Infection
4.5. Protein-Deficient Virions
5. Discussion
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| LCMV | Lymphocytic choriomeningitis virus |
| RING | Really interesting gene |
| ESCRT | Endosomal sorting complexes required for transport |
| -DG | -Dystroglycan |
| RdRp | RNA-dependent RNA polymerase |
| RNP | Ribonucleoprotein |
| SSP | Stable signal peptide |
| SSA | Stochastic simulation algorithm |
| ODE | Ordinary differential equation |
| MC | Markov chain |
References
- Poon, M.M.L.; Farber, D.L. The Whole Body as the System in Systems Immunology. iScience 2020, 23, 101509. [Google Scholar] [CrossRef] [PubMed]
- Germain, R.N.; Radtke, A.J.; Thakur, N.; Schrom, E.C.; Hor, J.L.; Ichise, H.; Arroyo-Mejias, A.J.; Chu, C.J.; Grant, S. Understanding immunity in a tissue-centric context: Combining novel imaging methods and mathematics to extract new insights into function and dysfunction. Immunol. Rev. 2022, 306, 8–24. [Google Scholar] [CrossRef] [PubMed]
- Hor, J.L.; Germain, R.N. Intravital and high-content multiplex imaging of the immune system. Trends Cell Biol. 2022, 32, 406–420. [Google Scholar] [CrossRef]
- Cardozo-Ojeda, E.F.; Perelson, A.S. Modeling HIV-1 Within-Host Dynamics After Passive Infusion of the Broadly Neutralizing Antibody VRC01. Front. Immunol. 2021, 12, 710012. [Google Scholar] [CrossRef]
- Sanche, S.; Cassidy, T.; Chu, P.; Perelson, A.S.; Ribeiro, R.M.; Ke, R. A simple model of COVID-19 explains disease severity and the effect of treatments. Sci. Rep. 2022, 12, 14210. [Google Scholar] [CrossRef] [PubMed]
- Nikas, A.; Ahmed, H.; Moore, M.R.; Zarnitsyna, V.I.; Antia, R. When does humoral memory enhance infection? PLoS Comput Biol. 2023, 19, e1011377. [Google Scholar] [CrossRef] [PubMed]
- Vilibic-Cavlek, T.; Savic, V.; Ferenc, T.; Mrzljak, A.; Barbic, L.; Bogdanic, M.; Stevanovic, V.; Tabain, I.; Ferencak, I.; Zidovec-Lepej, S. Lymphocytic Choriomeningitis—Emerging Trends of a Neglected Virus: A Narrative Review. Trop. Med. Infect. Dis. 2021, 6, 88. [Google Scholar] [CrossRef] [PubMed]
- Bocharov, G.; Argilaguet, J.; Meyerhans, A. Understanding Experimental LCMV Infection of Mice: The Role of Mathematical Models. J. Immunol. Res. 2015, 2015, 739706. [Google Scholar] [CrossRef]
- Argilaguet, J.; Pedragosa, M.; Esteve-Codina, A.; Riera, G.; Vidal, E.; Peligero-Cruz, C.; Casella, V.; Andreu, D.; Kaisho, T.; Bocharov, G.; et al. Systems analysis reveals complex biological processes during virus infection fate decisions. Genome Res. 2019, 29, 907–919. [Google Scholar] [CrossRef]
- Pedragosa, M.; Riera, G.; Casella, V.; Esteve-Codina, A.; Steuerman, Y.; Seth, C.; Bocharov, G.; Heath, S.; Gat-Viks, I.; Argilaguet, J.; et al. Linking Cell Dynamics With Gene Coexpression Networks to Characterize Key Events in Chronic Virus Infections. Front. Immunol. 2019, 10, 1002. [Google Scholar] [CrossRef]
- Bocharov, G.A. Modelling the dynamics of LCMV infection in mice: Conventional and exhaustive CTL responses. J. Theor. Biol. 1998, 192, 283–308. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, M.; Haenni, A.L. Expression strategies of ambisense viruses. Virus Res. 2003, 93, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Salvato, M.S.; Shimomaye, E.M. The completed sequence of lymphocytic choriomeningitis virus reveals a unique RNA structure and a gene for a zinc finger protein. Virology 1989, 173, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Southern, P.J.; Singh, M.K.; Riviere, Y.; Jacoby, D.R.; Buchmeier, M.J.; Oldstone, M.B. Molecular characterization of the genomic S RNA segment from lymphocytic choriomeningitis virus. Virology 1987, 157, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Meyer, B.J.; De La Torre, J.C.; Southern, P.J. Arenaviruses: Genomic RNAs, Transcription, and Replication. Curr. Top. Microbiol. Immunol. 2002, 262, 139–157. [Google Scholar] [CrossRef] [PubMed]
- Salvato, M.S.; Clegg, J.C.S.; Buchmeier, M.J.; Charrel, R.N.; Gonzales, J.P.; Lukashevich, I.S.; Peters, C.J.; Rico-Hesse, R.; Romanowski, V. Family Arenaviridae. In Virus Taxonomy: Ninth Report of the International Committee on Taxonomy of Viruses; King, A.M.Q., Adams, M.J., Carstens, E.B., Lefkowitz, E.F., Eds.; Elsevier: San Diego, CA, USA, 2012; pp. 715–723. [Google Scholar]
- Pinschewer, D.D.; Perez, M.; de la Torre, J.C. Dual role of the lymphocytic choriomeningitis virus intergenic region in transcription termination and virus propagation. J. Virol. 2005, 79, 4519–4526. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.J.; Novella, I.S.; Teng, M.N.; Oldstone, M.B.; de la Torre, J.C. NP and L proteins of lymphocytic choriomeningitis virus (LCMV) are sufficient for efficient transcription and replication of LCMV genomic RNA analogs. J. Virol. 2000, 74, 3470–3477. [Google Scholar] [CrossRef] [PubMed]
- Cornu, T.I.; de la Torre, J.C. RING finger Z protein of lymphocytic choriomeningitis virus (LCMV) inhibits transcription and RNA replication of an LCMV S-segment minigenome. J. Virol. 2001, 75, 9415–9426. [Google Scholar] [CrossRef]
- Kranzusch, P.J.; Whelan, S.P. Arenavirus Z protein controls viral RNA synthesis by locking a polymerase–promoter complex. Proc. Natl. Acad. Sci. USA 2011, 108, 19743–19748. [Google Scholar] [CrossRef]
- Perez, M.; Craven, R.C.; de la Torre, J.C. The small RING finger protein Z drives arenavirus budding: Implications for antiviral strategies. Proc. Natl. Acad. Sci. USA 2003, 100, 12978–12983. [Google Scholar] [CrossRef]
- Fehling, S.K.; Lennartz, F.; Strecker, T. Multifunctional nature of the arenavirus RING finger protein Z. Viruses 2012, 4, 2973–3011. [Google Scholar] [CrossRef] [PubMed]
- Borden, K.L.; Campbell Dwyer, E.J.; Salvato, M.S. An arenavirus RING (zinc-binding) protein binds the oncoprotein promyelocyte leukemia protein (PML) and relocates PML nuclear bodies to the cytoplasm. J. Virol. 1998, 72, 758–766. [Google Scholar] [CrossRef] [PubMed]
- Borden, K.L.; CampbellDwyer, E.J.; Carlile, G.W.; Djavani, M.; Salvato, M.S. Two RING finger proteins, the oncoprotein PML and the arenavirus Z protein, colocalize with the nuclear fraction of the ribosomal P proteins. J. Virol. 1998, 72, 3819–3826. [Google Scholar] [CrossRef] [PubMed]
- Campbell, D.E.J.; Lai, H.; MacDonald, R.C.; Salvato, M.S.; Borden, K.L. The lymphocytic choriomeningitis virus RING protein Z associates with eukaryotic initiation factor 4E and selectively represses translation in a RING-dependent manner. J. Virol. 2000, 74, 3293–3300. [Google Scholar] [CrossRef]
- Pinschewer, D.D.; Perez, M.; de la Torre, J.C. Role of the virus nucleoprotein in the regulation of lymphocytic choriomeningitis virus transcription and RNA replication. J. Virol. 2003, 77, 3882–3887. [Google Scholar] [CrossRef] [PubMed]
- Bederka, L.H.; Bonhomme, C.J.; Ling, E.L.; Buchmeier, M.J. Arenavirus stable signal peptide is the keystone subunit for glycoprotein complex organization. MBio 2014, 5, e02063-14. [Google Scholar] [CrossRef] [PubMed]
- Wright, K.E.; Spiro, R.C.; Burns, J.W.; Buchmeier, M.J. Post-translational processing of the glycoproteins of lymphocytic choriomeningitis virus. Virology 1990, 177, 175–183. [Google Scholar] [CrossRef]
- Cao, W.; Henry, M.D.; Borrow, P.; Yamada, H.; Elder, J.H.; Ravkov, E.V.; Nichol, S.T.; Compans, R.W.; Campbell, K.P.; Oldstone, M.B.A. Identification of α-dystroglycan as a receptor for lymphocytic choriomeningitis virus and Lassa fever virus. Science 1998, 282, 2079–2081. [Google Scholar] [CrossRef]
- Sevilla, N.; Kunz, S.; Holz, A.; Lewicki, H.; Homann, D.; Yamada, H.; Campbell, K.P.; de la Torre, J.C.; Oldstone, M.B.A. Immunosuppression and resultant viral persistence by specific viral targeting of dendritic cells. J. Exp. Med. 2000, 192, 1249–1260. [Google Scholar] [CrossRef]
- Kunz, S.; Sevilla, N.; McGavern, D.B.; Campbell, K.P.; Oldstone, M.B.A. Molecular analysis of the interaction of LCMV with its cellular receptor α-dystroglycan. J. Cell Biol. 2001, 155, 301–310. [Google Scholar] [CrossRef]
- Oldstone, M.B.; Campbell, K.P. Decoding arenavirus pathogenesis: Essential roles for alpha-dystroglycan-virus interactions and the immune response. Virology 2011, 411, 170–179. [Google Scholar] [CrossRef]
- Quirin, K.; Eschli, B.; Scheu, I.; Poort, L.; Kartenbeck, J.; Helenius, A. Lymphocytic choriomeningitis virus uses a novel endocytic pathway for infectious entry via late endosomes. Virology 2008, 378, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Rojek, J.M.; Sanchez, A.B.; Nguyen, N.T.; de la Torre, J.C.; Kunz, S. Different mechanisms of cell entry by human-pathogenic Old World and New World arenaviruses. J. Virol. 2008, 82, 7677–7687. [Google Scholar] [CrossRef] [PubMed]
- Rojek, J.M.; Perez, M.; Kunz, S. Cellular entry of lymphocytic choriomeningitis virus. J. Virol. 2008, 82, 1505–1517. [Google Scholar] [CrossRef] [PubMed]
- Lehmann-Grube, F.; Slenczka, W. Über die Vermehrung von LCM-Virus (Stamm WE3) in Zell-kulturen. Zentralblatt Bakteriol. Infekt. 1967, 206, 525. [Google Scholar]
- Sullivan, B.M.; Emonet, S.F.; Welch, M.J.; Lee, A.M.; Campbell, K.P.; de la Torre, J.C.; Oldstone, M.B. Point mutation in the glycoprotein of lymphocytic choriomeningitis virus is necessary for receptor binding, dendritic cell infection, and long-term persistence. Proc. Natl. Acad. Sci. USA 2011, 108, 2969–2974. [Google Scholar] [CrossRef]
- Wen, Y.; Xu, H.; Wan, W.; Shang, W.; Jin, R.; Zhou, F.; Mei, H.; Wang, J.; Xiao, G.; Chen, H.; et al. Visualizing lymphocytic choriomeningitis virus infection in cells and living mice. iScience 2022, 25, 105090. [Google Scholar] [CrossRef] [PubMed]
- Strecker, T.; Eichler, R.; Meulen, J.T.; Weissenhorn, W.; Dieter Klenk, H.; Garten, W.; Lenz, O. Lassa virus Z protein is a matrix protein sufficient for the release of virus-like particles. J. Virol. 2003, 77, 10700–10705. [Google Scholar] [CrossRef] [PubMed]
- Salvato, M.S.; Schweighofer, K.J.; Burns, J.; Shimomaye, E.M. Biochemical and immunological evidence that the 11 kDa zinc-binding protein of lymphocytic choriomeningitis virus is a structural component of the virus. Virus Res. 1992, 22, 185–198. [Google Scholar] [CrossRef]
- Kang, H.; Cong, J.; Wang, C.; Ji, W.; Xin, Y.; Qian, Y.; Li, X.; Chen, Y.; Rao, Z. Structural basis for recognition and regulation of arenavirus polymerase L by Z protein. Nat. Commun. 2021, 12, 4134. [Google Scholar] [CrossRef]
- Gillespie, D.T. A general method for numerically simulating the stochastic time evolution of coupled chemical reactions. J. Comput. Phys. 1976, 22, 403–434. [Google Scholar] [CrossRef]
- Gillespie, D.T. Exact stochastic simulation of coupled chemical reactions. J. Phys. Chem. 1977, 81, 2340–2361. [Google Scholar] [CrossRef]
- Marchetti, L.; Priami, C.; Thanh, V.H. Simulation Algorithms for Computational Systems Biology; An EATCS Series; Texts in Theoretical Computer Science; Springer International Publishing: Cham, Switzerland, 2017; ISBN 9783319631110. [Google Scholar]
- Sazonov, I.; Grebennikov, D.; Meyerhans, A.; Bocharov, G. Markov Chain-Based Stochastic Modelling of HIV-1 Life Cycle in a CD4 T Cell. Mathematics 2021, 9, 2025. [Google Scholar] [CrossRef]
- Thanh, V.H.; Zunino, R.; Priami, C. Efficient Constant-Time Complexity Algorithm for Stochastic Simulation of Large Reaction Networks. IEEE/ACM Trans. Comput. Biol. Bioinf. 2017, 14, 657–667. [Google Scholar] [CrossRef] [PubMed]
- Shcherbatova, O.; Grebennikov, D.; Sazonov, I.; Meyerhans, A.; Bocharov, G. Modeling of the HIV-1 life cycle in productively infected cells to predict novel therapeutic targets. Pathogens 2020, 9, 255. [Google Scholar] [CrossRef] [PubMed]
- Grebennikov, D.; Kholodareva, E.; Sazonov, I.; Karsonova, A.; Meyerhans, A.; Bocharov, G. Intracellular life cycle kinetics of SARS-CoV-2 predicted using mathematical modelling. Viruses 2021, 13, 1735. [Google Scholar] [CrossRef] [PubMed]
- Heldt, F.S.; Kupke, S.Y.; Dorl, S.; Reichl, U.; Frensing, T. Single-cell analysis and stochastic modelling unveil large cell-to-cell variability in influenza A virus infection. Nat. Commun. 2015, 6, 8938. [Google Scholar] [CrossRef]
- UniProt Consortium. UniProt: A worldwide hub of protein knowledge. Nucleic Acids Res. 2019, 47, D506–D515. [Google Scholar] [CrossRef]
- Heldt, F.S.; Frensing, T.; Reichl, U. Modeling the intracellular dynamics of influenza virus replication to understand the control of viral RNA synthesis. J. Virol. 2012, 86, 7806–7817. [Google Scholar] [CrossRef]
- Sazonov, I.; Grebennikov, D.; Meyerhans, A.; Bocharov, G. Sensitivity of SARS-CoV-2 Life Cycle to IFN Effects and ACE2 Binding Unveiled with a Stochastic Model. Viruses 2022, 14, 403. [Google Scholar] [CrossRef]
- Zhou, X.; Ramach, R.S.; Mann, M.; Popkin, D.L. Role of lymphocytic choriomeningitis virus (LCMV) in understanding viral immunology: Past, present and future. Viruses 2012, 4, 2650–2669. [Google Scholar] [CrossRef]
- Lee, A.M.; Cruite, J.; Welch, M.J.; Sullivan, B.; Oldstone, M.B. Pathogenesis of Lassa fever virus infection: I. Susceptibility of mice to recombinant Lassa Gp/LCMV chimeric virus. Virology 2013, 442, 114–121. [Google Scholar] [CrossRef]
- Mohammadi, P.; Desfarges, S.; Bartha, I.; Joos, B.; Zangger, N.; Muñoz, M.; Günthard, H.F.; Beerenwinkel, N.; Telenti, A.; Ciuffi, A. 24 hours in the life of HIV-1 in a T cell line. PLoS Pathog. 2013, 9, e1003161. [Google Scholar] [CrossRef]













| m | Elementary Reaction | Transition Intensity, |
|---|---|---|
| Cell entry | ||
| 1 | ||
| 2 | ||
| 3 | ||
| 4 | ||
| 5 | ||
| 6 | ||
| 7 | ||
| Replication | ||
| 8 | ||
| 9 | ||
| 10 | ||
| 11 | ||
| 12 | ||
| 13 | ||
| 14 | ||
| 15 | ||
| 16 | ||
| 17 | ||
| 18 | ||
| 19 | ||
| Transcription | ||
| 20 | ||
| 21 | ||
| 22 | ||
| 23 | ||
| 24 | ||
| 25 | ||
| 26 | ||
| 27 | ||
| Translation | ||
| 28 | ||
| 29 | ||
| 30 | ||
| 31 | ||
| 32 | ||
| 33 | ||
| 34 | ||
| 35 | ||
| Assembly and release | ||
| 36 | ||
| 37 | ||
| 38 | ||
| 39 | ||
| 40 | ||
| 41 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sergeeva, J.; Grebennikov, D.; Casella, V.; Cebollada Rica, P.; Meyerhans, A.; Bocharov, G. Mathematical Model Predicting the Kinetics of Intracellular LCMV Replication. Mathematics 2023, 11, 4454. https://doi.org/10.3390/math11214454
Sergeeva J, Grebennikov D, Casella V, Cebollada Rica P, Meyerhans A, Bocharov G. Mathematical Model Predicting the Kinetics of Intracellular LCMV Replication. Mathematics. 2023; 11(21):4454. https://doi.org/10.3390/math11214454
Chicago/Turabian StyleSergeeva, Julia, Dmitry Grebennikov, Valentina Casella, Paula Cebollada Rica, Andreas Meyerhans, and Gennady Bocharov. 2023. "Mathematical Model Predicting the Kinetics of Intracellular LCMV Replication" Mathematics 11, no. 21: 4454. https://doi.org/10.3390/math11214454
APA StyleSergeeva, J., Grebennikov, D., Casella, V., Cebollada Rica, P., Meyerhans, A., & Bocharov, G. (2023). Mathematical Model Predicting the Kinetics of Intracellular LCMV Replication. Mathematics, 11(21), 4454. https://doi.org/10.3390/math11214454