A Generalized Framework for Analyzing Taxonomic, Phylogenetic, and Functional Community Structure Based on Presence–Absence Data



Abstract
:1. Introduction
2. Preliminaries
3. The SDR Simplex and Its Generalization
3.1. Fractions of the Generalized Simplex
3.2. A Measure of Redundancy
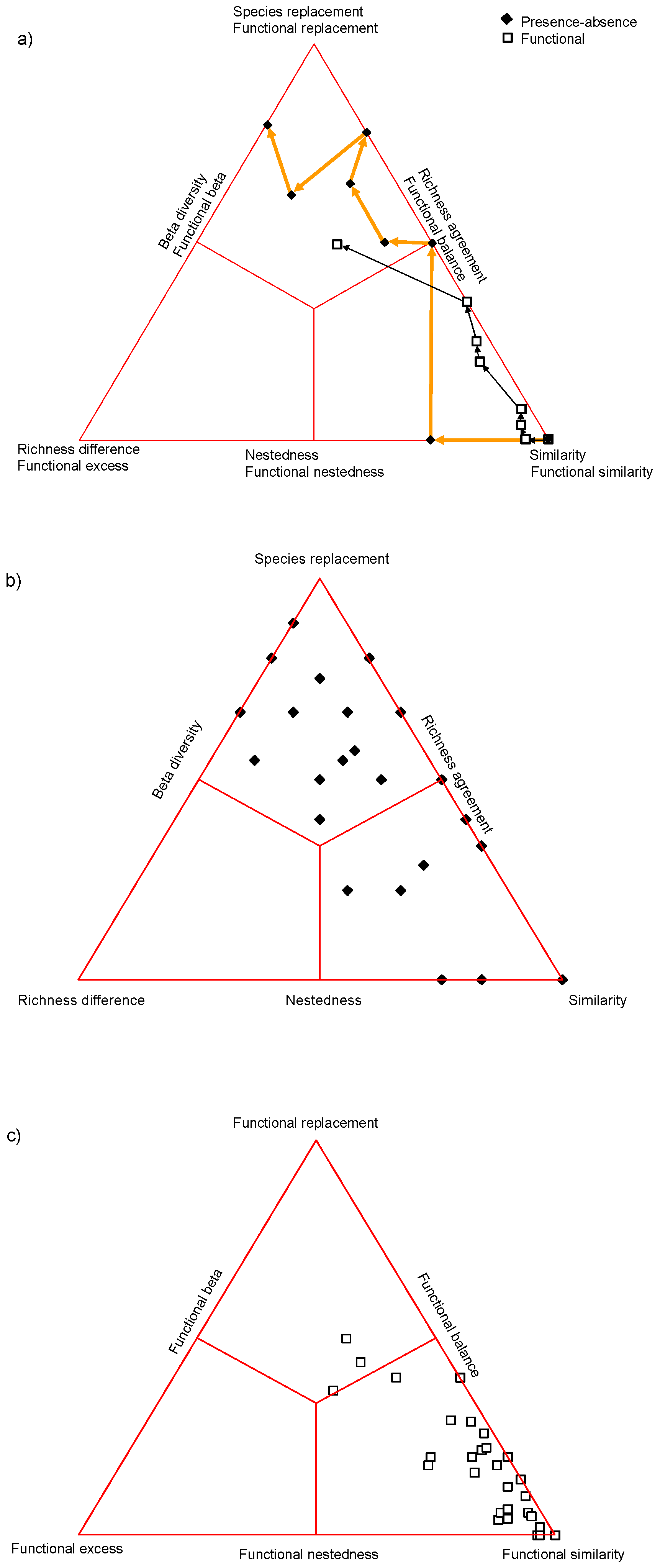
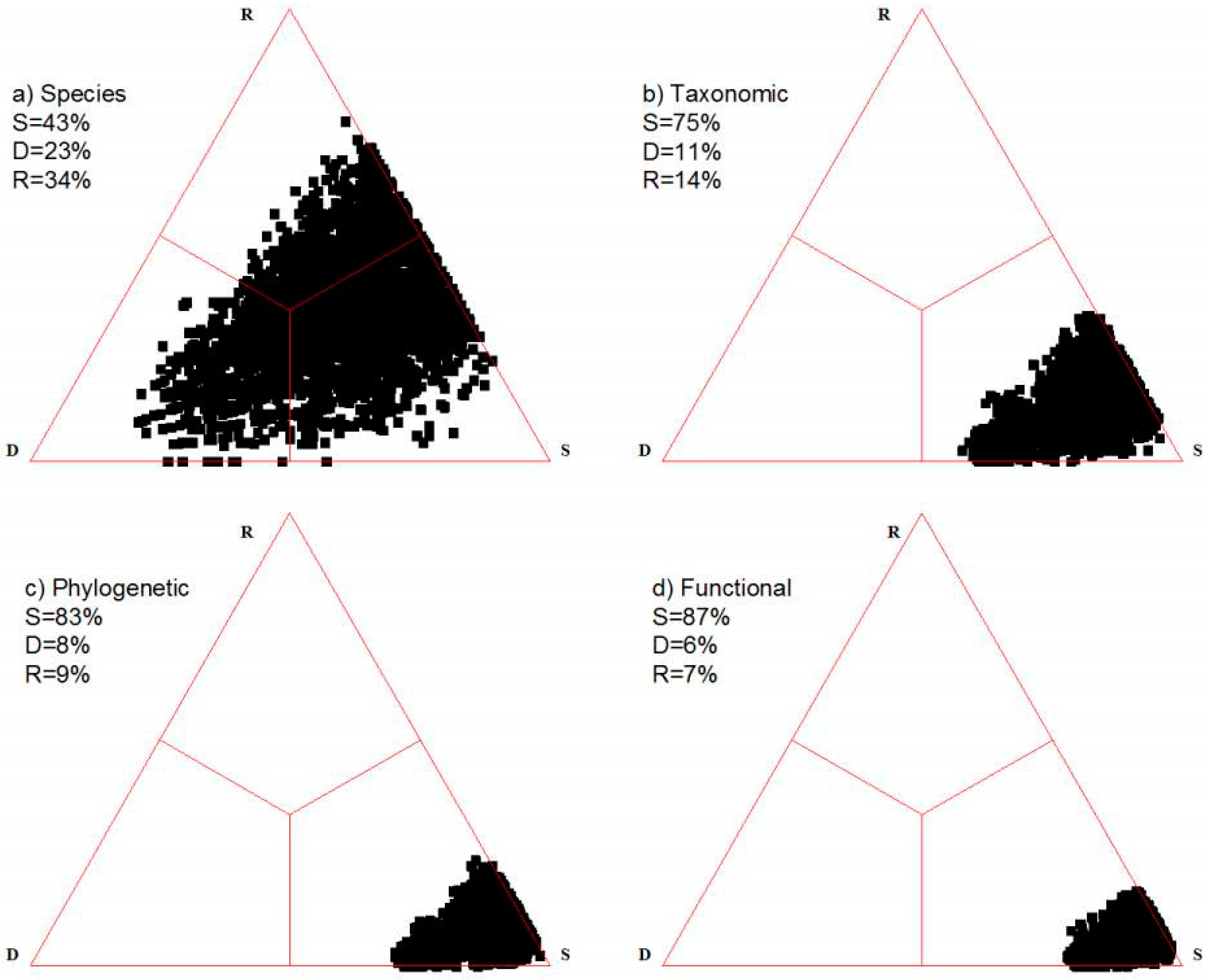
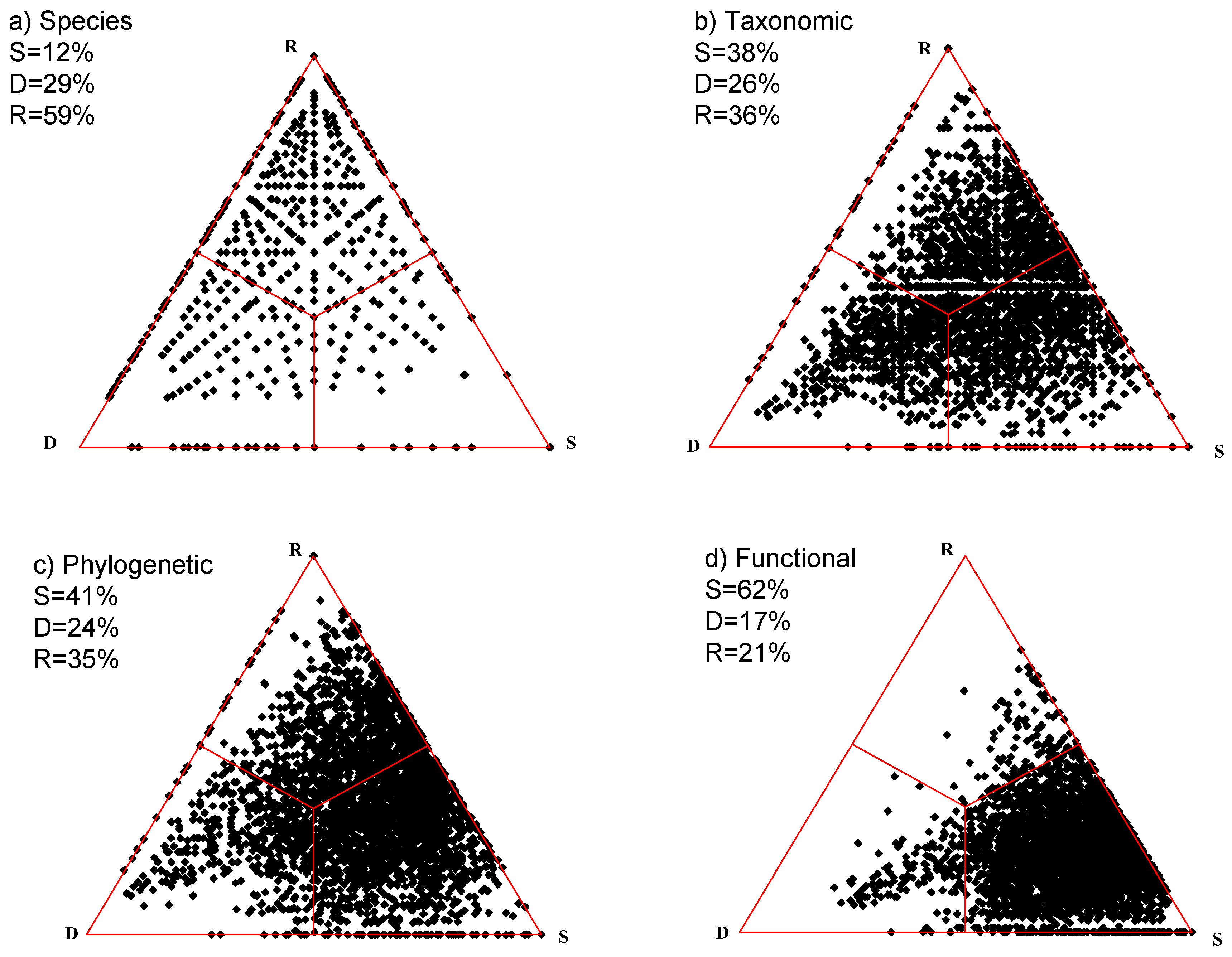
4. Illustrative Examples
4.1. Artificial Data
4.2. Actual Examples
4.2.1. Grassland
4.2.2. Primary Succession in the Alpine Zone
4.2.3. Coastal Marsh Vegetation
5. Discussion
- In fact this order cannot be evaluated whenever the measures applied to each level have different theoretical background. For example, Weinstein et al. [16] calculated taxon-based beta using the Sorensen index, and phylogenetic beta using the PhyloSor measure, so far so good, but functional beta was the mean nearest taxon distance calculated after PCA of the trait matrix. Tucker et al. [41] used the total branch length of the cladogram to measure phylogenetic beta and the convex hull for the functional beta in a simulation experiment. Most published research, like these, fails to satisfy this methodological consistency criterion and, therefore, their results cannot be compared with ours. Moreover, when the authors did care attention deliberately to commensurability [15], the results were in matrix form and remained unavailable for comparison.
- Functional diversity was the lowest (alpine meadow) when the number of functional variables was the lowest (3), raising the possibility that there is some direct relationship between these two.
- The choice of functional variables is subject to arbitrary decisions, while taxonomy and phylogeny do not depend on the authors’ wish. There is always the question if we indeed use the most meaningful set of functional characters in a given study.
- Taxonomy and phylogeny are constrained by their own hierarchy, neighbors or close relatives can only replace each other, whereas in terms of functionality two phylogenetically remote taxa may agree just as well, i.e., convergence increases the probability that one absent species may be replaced by another with similar functional traits.
- In theory, all the differential species in every comparison can be completely replaced regarding functionality, while this is impossible taxonomically and phylogenetically.
- Functional diversity as we measure here apparently correlates with environmental heterogeneity. Here it is the lowest in the alpine meadow vegetation (uniform habitat) followed by the grasslands also with fairly uniform habitat which differ only in exposition, and then comes the marshland which had the highest habitat heterogeneity;
- Thus, theoretically it may happen that environmental heterogeneity exceeds phylogenetic heterogeneity, i.e. when fairly related species were forced to adapt to extremely different environmental conditions and in such cases the beta redundancy hypothesis may not be true. In other words, the above order may be constrained by environmental heterogeneity and if we collect an extremely diverse sample (in which sites have nothing to do with each other).
- Our results do not support the view that phylogenetic beta often serves as a good surrogate to functional beta at the local scale; there was quite a big difference between them in homogeneous (alpine meadow) and heterogeneous (coastal marsh) environment as well. We agree with Losos [42] and Swenson et al. [43] who reached similar conclusions regarding the predictability of functional dissimilarity by phylogenetic dissimilarity.
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 1 | 1 | 1 | 1 | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| 1 | 1 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| 0 | 0 | 1 | 1 | 0 | 1 | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| 0 | 0 | 0 | 1 | 0 | 0 | 1 | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 0 |
| 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 0 | 0 | 1 | 0 | 0 |
| 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 0 | 0 |
| 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 1 | 1 | 0 |
| 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 |
| 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 |
| 0 | 0 | 0.1 | 0.2 | 0.2 | 0.3 | 0.4 | 0.4 | 0.5 | 0.4 | 0.6 | 0.7 | 1 | 0.9 | 0.8 |
| 0 | 0 | 0.1 | 0.2 | 0.2 | 0.3 | 0.4 | 0.4 | 0.5 | 0.4 | 0.6 | 0.7 | 1 | 0.9 | 0.8 |
| 0.1 | 0.1 | 0 | 0.1 | 0.3 | 0.4 | 0.3 | 0.3 | 0.4 | 0.5 | 0.7 | 0.8 | 1 | 0.8 | 0.9 |
| 0.2 | 0.2 | 0.1 | 0 | 0.4 | 0.5 | 0.2 | 0.4 | 0.3 | 0.6 | 0.8 | 0.9 | 0.9 | 0.9 | 1 |
| 0.2 | 0.2 | 0.3 | 0.4 | 0 | 0.1 | 0.4 | 0.4 | 0.5 | 0.4 | 0.4 | 0.7 | 1 | 0.9 | 0.8 |
| 0.3 | 0.3 | 0.4 | 0.5 | 0.1 | 0 | 0.3 | 0.3 | 0.4 | 0.3 | 0.3 | 0.6 | 1 | 0.8 | 0.7 |
| 0.4 | 0.4 | 0.3 | 0.2 | 0.4 | 0.3 | 0 | 0.2 | 0.1 | 0.4 | 0.6 | 0.7 | 0.7 | 0.7 | 0.8 |
| 0.4 | 0.4 | 0.3 | 0.4 | 0.4 | 0.3 | 0.2 | 0 | 0.1 | 0.2 | 0.4 | 0.5 | 0.7 | 0.5 | 0.6 |
| 0.5 | 0.5 | 0.4 | 0.3 | 0.5 | 0.4 | 0.1 | 0.1 | 0 | 0.3 | 0.5 | 0.6 | 0.6 | 0.6 | 0.7 |
| 0.4 | 0.4 | 0.5 | 0.6 | 0.4 | 0.3 | 0.4 | 0.2 | 0.3 | 0 | 0.2 | 0.3 | 0.7 | 0.5 | 0.4 |
| 0.6 | 0.6 | 0.7 | 0.8 | 0.4 | 0.3 | 0.6 | 0.4 | 0.5 | 0.2 | 0 | 0.3 | 0.7 | 0.5 | 0.4 |
| 0.7 | 0.7 | 0.8 | 0.9 | 0.7 | 0.6 | 0.7 | 0.5 | 0.6 | 0.3 | 0.3 | 0 | 0.4 | 0.2 | 0.1 |
| 1 | 1 | 1 | 0.9 | 1 | 1 | 0.7 | 0.7 | 0.6 | 0.7 | 0.7 | 0.4 | 0 | 0.2 | 0.3 |
| 0.9 | 0.9 | 0.8 | 0.9 | 0.9 | 0.8 | 0.7 | 0.5 | 0.6 | 0.5 | 0.5 | 0.2 | 0.2 | 0 | 0.1 |
| 0.8 | 0.8 | 0.9 | 1 | 0.8 | 0.7 | 0.8 | 0.6 | 0.7 | 0.4 | 0.4 | 0.1 | 0.3 | 0.1 | 0 |
References
- Orlóci, L. Multivariate Analysis in Vegetation Research, 2nd ed.; Junk: The Hague, The Netherlands, 1978. [Google Scholar]
- Legendre, P.; Legendre, L. Numerical Ecology, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Magurran, A.E. Measuring Biological Diversity; Blackwell Science: Oxford, UK, 2004. [Google Scholar]
- Baselga, A. Multiple site dissimilarity quantifies compositional heterogeneity among several sites, while average pairwise dissimilarity may be misleading. Ecography 2013, 36, 124–128. [Google Scholar] [CrossRef]
- Baselga, A. Partitioning the turnover and nestedness components of beta diversity. Glob. Ecol. Biogeogr. 2010, 19, 134–143. [Google Scholar] [CrossRef]
- Podani, J.; Schmera, D. A new conceptual and methodological framework for exploring and explaining pattern in presence–absence data. Oikos 2011, 120, 1625–1638. [Google Scholar] [CrossRef]
- Carvalho, J.C.; Cardoso, P.; Gomes, P. Determining the relative roles of species replacement and species richness differences in generating beta-diversity patterns. Glob. Ecol. Biogeogr. 2012, 21, 760–771. [Google Scholar] [CrossRef]
- Legendre, P. Interpreting the replacement and richness difference components of beta diversity. Glob. Ecol. Biogeogr. 2014, 23, 1324–1334. [Google Scholar] [CrossRef] [Green Version]
- Jaccard, P. Distribution de la flore alpine dans le bassin des Dranses et dans quelques régions voisines. Bulletin. Société Vaudoise Sci. Naturelles 1901, 37, 241–272. [Google Scholar]
- Warwick, R.M.; Clarke, K.R. New “biodiversity” measures reveal a decrease in taxonomic distinctness with increasing stress. Mar. Ecol. Prog. Ser. 1995, 129, 301–305. [Google Scholar] [CrossRef]
- Warwick, R.M.; Clarke, K.R. Taxonomic distinctness and environmental assessment. J. Appl. Ecol. 1998, 35, 532–543. [Google Scholar] [CrossRef] [Green Version]
- Pavoine, S.; Baguette, M.; Stevens, V.M.; Leibold, M.; Turlure, C.; Bonsall, M.B. Life history traits, but not phylogeny, drive compositional patterns in a butterfly metacommunity. Ecology 2014, 95, 3304–3313. [Google Scholar] [CrossRef] [Green Version]
- Gerhold, P.; Cahill, J.; Winter, M.; Bartish, I.V.; Prinzing, A. Phylogenetic patterns are not proxies of community assembly mechanisms (they are far better). Funct. Ecol. 2015, 29, 600–614. [Google Scholar] [CrossRef] [Green Version]
- Cardoso, P.; Rigal, F.; Carvalho, J.C.; Fortelius, M.; Borges, P.A.V.; Podani, J.; Schmera, D. Partitioning taxon, phylogenetic and functional beta diversity into replacement and richness difference components. J. Biogeogr. 2014, 41, 749–761. [Google Scholar] [CrossRef]
- Corbelli, J.M.; Zurita, G.A.; Filloy, J.; Galvis, J.P.; Vespa, N.I.; Bellocq, I. Integrating taxonomic, functional and phylogenetic beta diversities: Interactive effects with the biome and land use across taxa. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [PubMed]
- Weinstein, B.G.; Tinoco, B.; Parra, J.L.; Brown, L.M.; McGuire, J.A.; Stiles, F.G.; Graham, C.H. Taxonomic, phylogenetic, and trait beta diversity in South American hummingbirds. Am. Nat. 2014, 184, 211–224. [Google Scholar] [CrossRef] [PubMed]
- Ricotta, C.; Podani, J.; Pavoine, S. A family of functional dissimilarity measures for presence and absence data. Ecol. Evol. 2016, 6, 5383–5389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izsak, C.; Price, A.R.G. Measuring β-diversity using a taxonomic similarity index, and its relation to spatial scale. Mar. Ecol. Prog. Ser. 2001, 215, 69–77. [Google Scholar] [CrossRef]
- Carvalho, J.C.; Cardoso, P.; Borges, P.A.V.; Schmera, D.; Podani, J. Measuring fractions of beta diversity and their relationships to nestedness: A theoretical and empirical comparison of novel approaches. Oikos 2013, 122, 825–834. [Google Scholar] [CrossRef] [Green Version]
- Podani, J. Extending Gower’s general coefficient of similarity to ordinal characters. Taxon 1999, 48, 331–340. [Google Scholar] [CrossRef]
- Podani, J.; Schmera, D. On dendrogram-based measures of functional diversity. Oikos 2006, 115, 179–185. [Google Scholar] [CrossRef] [Green Version]
- Pavoine, S.; Vallet, J.; Dufour, A.-B.; Gacher, S.; Daniel, H. On the challenge of treating various types of variables: Application for improving the measurement of functional diversity. Oikos 2009, 118, 391–402. [Google Scholar] [CrossRef]
- Pavoine, S.; Marcon, E.; Ricotta, C. “Equivalent numbers” for species, phylogenetic, or functional diversity in a nested hierarchy of multiple scales. Meth. Ecol. Evol. 2016, 7, 1152–1163. [Google Scholar] [CrossRef]
- Chase, M.W.; Reveal, J.L. A phylogenetic classification of the land plants to accompany APG III. Bot. J. Linn. Soc. 2009, 161, 122–127. [Google Scholar] [CrossRef] [Green Version]
- Angiosperm Phylogeny Group. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. Soc. 2016, 181, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Podani, J.; Miklós, I. Resemblance coefficients and the horseshoe effect in principal coordinates analysis. Ecology 2002, 83, 3331–3343. [Google Scholar] [CrossRef]
- Phylotools: Phylogenetic Tools for Eco-Phylogenetics; R package Version 0.1.2. Available online: https://cran.r-project.org/web/packages/phylotools/index.html (accessed on 12 November 2018).
- Paradis, E.; Claude, J.; Strimmer, K. APE: Analyses of phylogenetics and evolution in R language. Bioinformatics 2004, 20, 289–290. [Google Scholar] [CrossRef] [PubMed]
- Caccianiga, M.; Luzzaro, A.; Pierce, S.; Ceriani, R.M.; Cerabolini, B. The functional basis of a primary succession resolved by CSR classification. Oikos 2006, 112, 10–20. [Google Scholar] [CrossRef]
- Ricotta, C.; Luzzaro, A.; Pierce, S.; Ceriani, R.M.; Cerabolini, B. Measuring the functional redundancy of biological communities: A quantitative guide. Meth. Ecol. Evol. 2016, 7, 1386–1395. [Google Scholar] [CrossRef]
- Ricotta, C.; Bacaro, G.; Caccianiga, M.; Cerabolini, B.E.L.; Moretti, M. A classical measure of phylogenetic dissimilarity and its relationship with beta diversity. Basic Appl. Ecol. 2015, 16, 10–18. [Google Scholar] [CrossRef]
- Durka, W.; Michalski, S.G. Daphne: A dated phylogeny of a large European flora for phylogenetically informed ecological analyses. Ecology 2012, 93, 2297. [Google Scholar] [CrossRef]
- Pavoine, S.; Vela, E.; Gacher, S.; de Bélair, G.; Bonsall, M.B. Linking patterns in phylogeny, traits, abiotic variables and space: A novel approach to linking environmental filtering and plant community assembly. J. Ecol. 2011, 99, 165–175. [Google Scholar] [CrossRef]
- De Bélair, G. Biogéographie et Aménagement: La Plaine de La Mafragh (Annaba, Algérie). Ph.D. Thesis, Université Paul Valéry, Montpellier, France, 1981. [Google Scholar]
- Gachet, S.; Véla, E.; Tatoni, T. BASECO: A floristic and ecological database of Mediterranean French flora. Biodivers. Conserv. 2005, 14, 1023–1034. [Google Scholar] [CrossRef]
- Webb, C.O.; Ackerly, D.D.; Kembel, S.W. Phylocom: Software for the analysis of phylogenetic community structure and character evolution. Bioinformatics 2008, 24, 2098–2100. [Google Scholar] [CrossRef] [PubMed]
- Hedges, S.B.; Kumar, S. (Eds.) The Timetree of Life; Oxford University Press: Oxford, UK, 2009. [Google Scholar]
- Webb, C.O.; Ackerly, D.D.; McPeek, M.A.; Donoghue, M.J. Phylogenies and Community Ecology. Annu. Rev. Ecol. Syst. 2002, 33, 475–505. [Google Scholar] [CrossRef]
- Villéger, S.S.; Grenouillet, G.; Brosse, S. Decomposing functional beta-diversity reveals that low functional beta-diversity is driven by low functional turnover in European fish assemblages. Glob. Ecol. Biogeogr. 2013, 22, 671–681. [Google Scholar] [CrossRef]
- Leprieur, F.; Albouy, C.; De Bortoli, J.; Cowman, P.F.; Bellwood, D.R.; Mouillot, D. Quantifying phylogenetic beta diversity: Distinguishing between ‘true’ turnover of lineages and phylogenetic diversity gradients. PLoS ONE 2012, 7. [Google Scholar] [CrossRef]
- Tucker, C.M.; Davies, T.J.; Cadotte, M.W.; Pearse, W.D. On the relationship between phylogenetic diversity and trait diversity. Ecology 2018, 99, 1473–1479. [Google Scholar] [CrossRef] [PubMed]
- Losos, J.B. Phylogenetic niche conservatism, phylogenetic signal and the relationship between phylogenetic relatedness and ecological similarity among species. Ecol. Lett. 2008, 11, 995–1003. [Google Scholar] [CrossRef] [PubMed]
- Swenson, N.G.; Erickson, D.; Mi, X.; Bourg, N.A.; Forero-Montana, J.; Ge, X.; Howe, R.; Lake, J.K.; Liu, X.; Ma, K.; et al. Phylogenetic and functional alpha and beta diversity in temperate and tropical tree communities. Ecology 2012, 93. [Google Scholar] [CrossRef] [Green Version]




| Component | Taxon | Taxonomic | Phylogenetic | Functional |
|---|---|---|---|---|
| dij = 1 | 0 ≤ dij ≤ 1 | |||
| S | similarity | t. similarity | ph. similarity | f. similarity |
| D | richness difference | t. excess | ph. excess | f. excess |
| R | replacement | t. replacement | ph. replacement | f. replacement (turnover) |
| β = R + D = JAC | dissimilarity (pairwise beta diversity, species turnover) | t. dissimilarity (=t. beta) | ph. dissimilarity (ph. beta) | f. dissimilarity (f. beta) |
| G = R + S | richness agreement | t. balance | ph. balance | f. balance |
| N = D + S (S > 0) | nestedness | t. nestedness | ph. nestedness | f. nestedness |
| Case study | Taxonomic (E′st) | Phylogenetic (E′sp) | Functional (E′sf) |
|---|---|---|---|
| Grassland | 0.27 | 0.35 | 0.38 |
| Alpine meadow | 0.29 | 0.41 | 0.57 |
| Salt marsh | 0.25 | 0.27 | 0.44 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Podani, J.; Pavoine, S.; Ricotta, C. A Generalized Framework for Analyzing Taxonomic, Phylogenetic, and Functional Community Structure Based on Presence–Absence Data. Mathematics 2018, 6, 250. https://doi.org/10.3390/math6110250
Podani J, Pavoine S, Ricotta C. A Generalized Framework for Analyzing Taxonomic, Phylogenetic, and Functional Community Structure Based on Presence–Absence Data. Mathematics. 2018; 6(11):250. https://doi.org/10.3390/math6110250
Chicago/Turabian StylePodani, János, Sandrine Pavoine, and Carlo Ricotta. 2018. "A Generalized Framework for Analyzing Taxonomic, Phylogenetic, and Functional Community Structure Based on Presence–Absence Data" Mathematics 6, no. 11: 250. https://doi.org/10.3390/math6110250
APA StylePodani, J., Pavoine, S., & Ricotta, C. (2018). A Generalized Framework for Analyzing Taxonomic, Phylogenetic, and Functional Community Structure Based on Presence–Absence Data. Mathematics, 6(11), 250. https://doi.org/10.3390/math6110250