Modeling and Controlling Epidemic Outbreaks: The Role of Population Size, Model Heterogeneity and Fast Response in the Case of Measles

Abstract
:1. Introduction
2. Materials and Methods
2.1. Model
2.2. Simulations
3. Results
3.1. Theoretical Results
3.1.1. Convergence as Population Size Increased
3.1.2. Outbreaks Are Longer with Two Subgroups
3.1.3. Heterogeneity in Infection Risk
3.2. Numerical Results
3.2.1. The Impact of Population Size
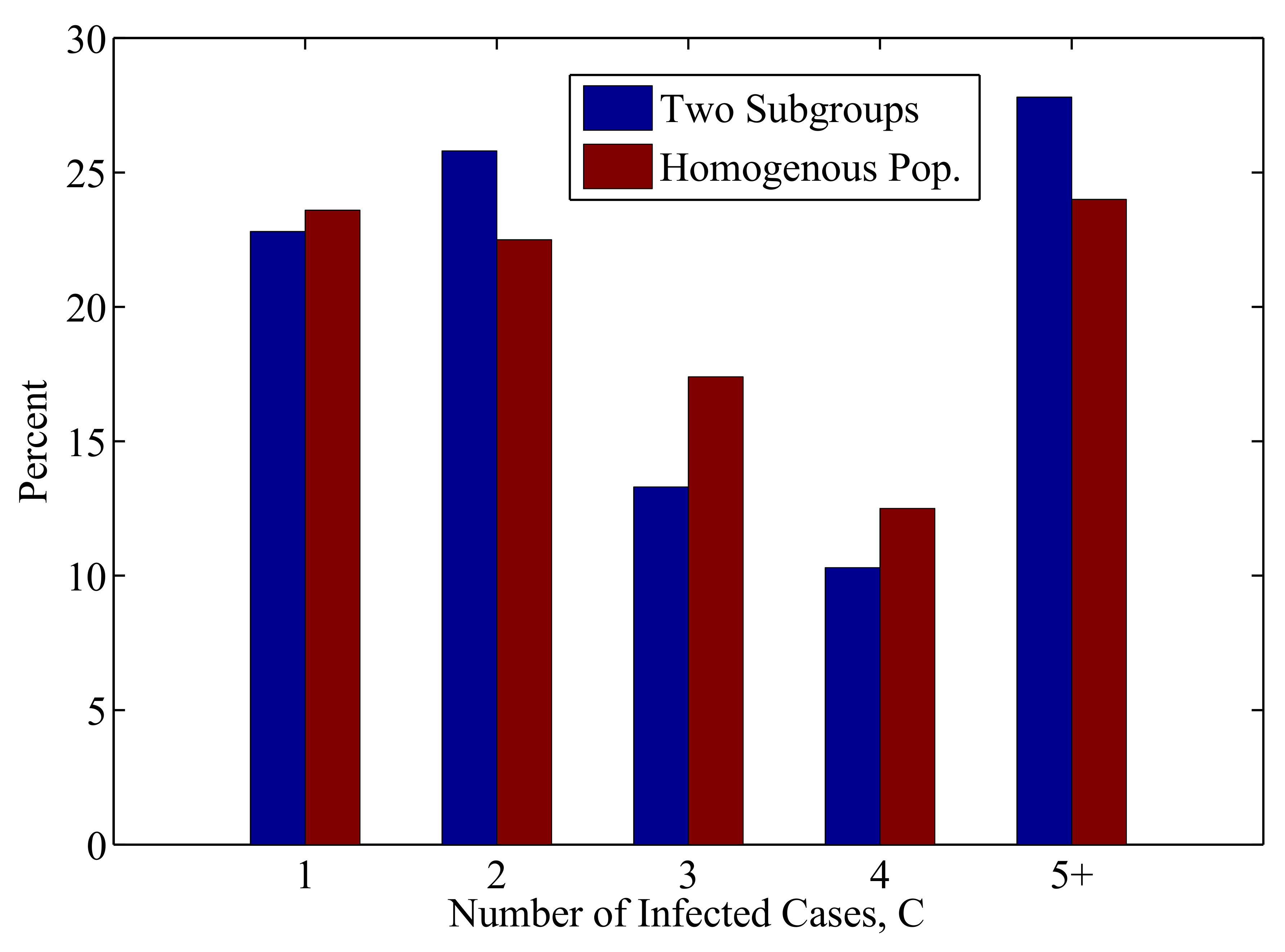
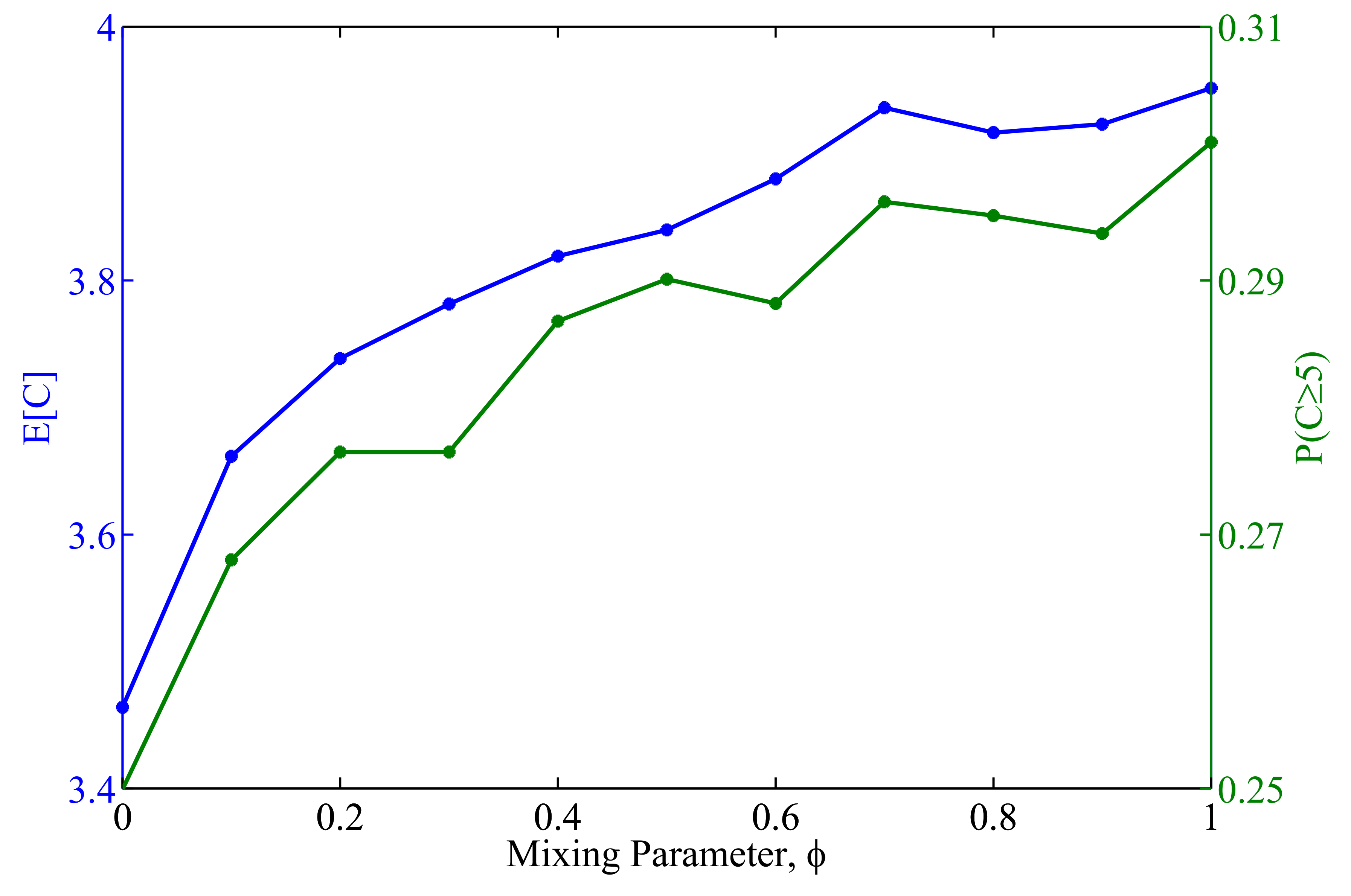
3.2.2. The Impact of Heterogeneity
3.2.3. The Impact of Public Health Interventions
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- DeRoo, S.S.; Pudalov, N.J.; Fu, L.Y. Planning for a COVID-19 Vaccination Program. JAMA 2020, 323, 2458–2459. [Google Scholar] [CrossRef]
- Measles Origin Finding Could Inform COVID-19 Research. Available online: https://medicalxpress.com/news/2020-06-measles-covid-.html (accessed on 18 October 2020).
- Sidiq, K.R.; Sabir, D.K.; Ali, S.M.; Kodzius, R. Does early childhood vaccination protect against COVID-19? Front. Mol. Biosci. 2020, 7, 120. [Google Scholar] [CrossRef]
- Franklin, R.; Young, A.; Neumann, B.; Fernandez, R.; Joannides, A.; Reyahi, A.; Modis, Y. Homologous protein domains in SARS-CoV-2 and measles, mumps and rubella viruses: Preliminary evidence that MMR vaccine might provide protection against COVID-19. medRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Deshpande, S.; Balji, S. MMR Vaccine and Covid-19: A Myth or a Low Risk-High Reward Preventive Measure? Indian Pediatrics 2020, 57, 773. [Google Scholar] [CrossRef]
- Shanker, V. Measles immunization: Worth considering containment Strategy for SARS-CoV-2 global outbreak. Indian Pediatrics 2020, 57, 380. [Google Scholar] [CrossRef]
- Fidel, P.L.; Noverr, M.C. Could an unrelated live attenuated vaccine serve as a preventive measure to dampen septic inflammation associated with COVID-19 Infection? mBio 2020, 11, e00907-20. [Google Scholar] [CrossRef]
- Sajuni, S. Vaksinasi Measles, Mumps, dan Rubella (MMR) Sebagai Prophylaxis Terhadap COVID-19. Keluwih J. Kesehat. Kedokt. 2020, 1, 25–28. [Google Scholar]
- Saad, M.E.; Elsalamony, R.A. Measles vaccines may provide partial protection against COVID-19. Int. J. Cancer Biomed. Res. 2020, 5, 14–19. [Google Scholar] [CrossRef] [Green Version]
- Salman, S.; Salem, M.L. Routine childhood immunization may protect against COVID-19. Med. Hypotheses 2020, 140, 109689. [Google Scholar] [CrossRef]
- Larenas-Linnemann, D.E.; Rodrigues-Monroy, F. Thirty-six COVID-19 cases preventively vaccinated with mumps-measles-rubella vaccine: All mild course. Allergy 2020. [Google Scholar] [CrossRef]
- Zhu, H.; Li, Y.; Jin, X.; Hunag, J.; Liu, X.; Tan, J. Transmission dynamics and control methodology of COVID-19: A modeling study Transmission dynamics and control methodology of COVID-19: A modeling study. Appl. Math. Model. 2020. [Google Scholar] [CrossRef]
- Chen, X.; Yu, B. First two months of the 2019 Coronavirus Disease (COVID-19) epidemic in China: Realtime surveillance and evaluation with a second derivative model. Glob. Health Res. Policy 2020, 5, 7. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Progress towards regional measles elimination—worldwide, 2000–2017 Wkly. Epidemiol. Rec. 2018, 93, 649–660. [Google Scholar]
- WHO. Measles, rubella and CRS: Disease description, epidemiology and diagnosis. In Surveillance Guidelines for Measles, Rubella and Congenital Rubella Syndrome in the WHO European Region; World Health Organization: Geneva, Switzerland, 2012. Available online: https://www.ncbi.nlm.nih.gov/books/NBK143257/ (accessed on 17 October 2020).
- Miller, D.L. Frequency of complications of measles, 1963. Br. Med. J. 1964, 2, 75–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gindler, J.; Tinker, S.; Markowitz, L.; Atkinson, W.; Dales, L.; Papania, M.J. Acute measles mortality in the United States, 1987–2002. J. Infect. Dis. 2004, 189 (Suppl. 1), 69–77. [Google Scholar] [CrossRef] [Green Version]
- Blumberg, S.; Enaroria, W.T.A.; Lyoyd-Smith, J.O.; Lietman, T.M.; Porco, T.C. Identifying post-elimination trends for the introduction and transmissibility of measles in the United States. Am. J. Epidemiol. 2014, 79, 1375–1382. [Google Scholar] [CrossRef]
- Patel, M.; Lee, A.D.; Redd, S.B.; Clemmons, N.S.; McNall, R.J.; Cohn, A.C.; Gastañaduy, P.A. Increase in measles cases—United States, January 1–26 April 2019. Morb. Mortal. Wkly. Rep. 2019, 68, 402–404. [Google Scholar] [CrossRef]
- Graham, M.K.; Winter, A.K.; Ferrari, M.; Grenfell, B.; Moss, W.J.; Azman, A.W.C.; Metcalf, J.E.; Lessler, J. Measles and the canonical path to elimination. Science 2019, 364, 584–587. [Google Scholar] [CrossRef] [Green Version]
- Health Protection Agency. Laboratory confirmed cases of measles, mumps and rubella in England and Wales: Update to end June 2012. Health Protection Report. Available online: http://www.hpa.org.uk/hpr/archives/2012/hpr3412.pdf (accessed on 24 August 2012).
- Fiebelkorn, A.P.; Redd, S.B.; Gallagher, K.; Rota, P.A.; Rota, J.; Bellini, W.; Seward, J. Measles in the United States during the post-elimination era. J. Infect. Dis. 2010, 202, 1520–1528. [Google Scholar] [CrossRef]
- The Lancet Child Adolescent Health. Vaccine hesitancy: A generation at risk. Lancet Child. Adolesc. Health 2019, 3, 281. [Google Scholar] [CrossRef]
- Arede, M.; Bravo-Araya, M.; Milie Bouchard, É.; Singh Gill, G.; Plajer, V.; Shehraj, A.; Shuaib, Y.A. Combating Vaccine Hesitancy: Teaching the Next Generation to Navigate Through the Post Truth Era. Front. Public Health 2019, 14, 381–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, S.; Dubey, V. Addressing vaccine hesitancy. Clinical guidance for primary care physicians working with parents. Can. Fam. Physician 2019, 65, 175–181. [Google Scholar] [PubMed]
- Marcus, B. A Nursing Approach to the Largest Measles Outbreak in Recent U.S. History: Lessons Learned Battling Homegrown Vaccine Hesitancy. Online J. Issues Nurs. 2020, 25. [Google Scholar] [CrossRef]
- Banerjee, E.; Griffith, J.; Kenyon, C.; Christianson, B.; Strain, A.; Martin, K.; McMahon, M.; Bagstad, E.; Hardy, K.; Grilli, G.; et al. Containing a measles outbreak in Minnesota, 2017: Methods and challenges. Perspect. Public Health 2020, 140, 162–171. [Google Scholar] [CrossRef] [PubMed]
- Gastanaduy, P.A.; Banerjee, E.; DeBolt, C.; Bravo-Alcantara, P.; Samad, S.A.; Pastor, D.; Rota, P.A.; Patel, M.; Crowcroft, N.S.; Durrheim, D.N. Public health responses during measles outbreaks in elimination settings: Strategies and challenges. Hum. Vaccines Immunother. 2018, 14, 2222–2238. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. CDC Measles—United States, 24 August 2012. Morb. Mortal. Wkly. Rep. 2012, 61, 647–652. [Google Scholar]
- Gastañaduy, P.A.; Redd, S.B.; Fiebelkorn, A.P.; Rota, J.S.; Rota, P.A.; Bellini, W.J.; Seward, J.F.; Wallace, G.S. CDC Measles—United States, 1 January–23 May 2014. Morb. Mortal. Wkly. Rep. 2014, 63, 496–499. [Google Scholar]
- CDC Measles Cases. Available online: http://www.cdc.gov/measles/cases-outbreaks.html (accessed on 12 August 2020).
- Glass, K.; Kappey, J.; Grenfell, B.T. The effect of heterogeneity in measles vaccination on population immunity. Epidemiol. Infect. 2004, 132, 675–683. [Google Scholar] [CrossRef]
- Barlett, M.S. Measles periodicity and community Size. J. R. Stat. Soc. A 1957, 120, 48–70. [Google Scholar] [CrossRef]
- Barlett, M.S. The critical community size for measles in the United States. J. R. Stat. Soc. A 1960, 123, 37–44. [Google Scholar] [CrossRef]
- Black, F. Measles endemicity in insular populations: Critical community size and its evolutionary implication. J. Theor. Biol. 1966, 11, 207–211. [Google Scholar] [CrossRef]
- Keeling, M.J.; Grenfell, B.T. Disease extinction and community size: Modeling the persistence of measles. Science 1997, 275, 65–67. [Google Scholar] [CrossRef]
- Anderson, R.M.; May, R.M. Infectious Diseases in Humans. Dynamics and Control; Oxford University Press: Oxford, UK, 1992. [Google Scholar]
- Measles History. Available online: https://www.cdc.gov/measles/about/history.html (accessed on 17 October 2020).
- Murray, G.D.; Cliff, A.D. A stochastic model for measles epidemics in a multi-region setting. Trans. Inst. Br. Geogr. 1977, 2, 158–174. [Google Scholar] [CrossRef]
- Bolker, B.; Grenfell, B.T. Space, persistence and dynamics of measles epidemics. Phil. Trans. R. Soc. Lond. 1995, B251, 75–81. [Google Scholar]
- Hethcote, H.W.; Van Ark, J.W. Epidemiological models for heterogeneous populations: Proportionate mixing, parameter estimation and immunization programs. Math. Biosci. 1987, 75, 205–227. [Google Scholar] [CrossRef]
- Castillo-Chavez, C.; Hethcote, H.W.; Andreasen, V.; Levin, S.A.; Liu, W.M. Epidemiological models with age structure, proportionate mixing, and cross-immunity. J. Math. Biol. 1989, 27, 233–258. [Google Scholar] [CrossRef] [PubMed]
- Hethcote, H.W. Modeling heterogeneous mixing in infectious disease dynamics. In Models for Infectious Human Diseases: Their Structure and Relation to Data; Isham, V., Medley, G., Eds.; Cambridge University Press: Cambridge, UK, 1996; pp. 215–238. [Google Scholar]
- Llyod, A.L.; May, R.M. Spatial heterogeneity in epidemic models. J. Theor. Biol. 1996, 179, 1–11. [Google Scholar] [CrossRef]
- Sattenspiel, L.; Dietz, K. A structured epidemic model incorporating geographic mobility among regions. Math. Biosci. 1995, 128, 71–91. [Google Scholar] [CrossRef]
- Allen, L.J.S.; Jones, M.A.; Martin, C.F. A discrete-time model with vaccination for a measles epidemic. Math. Biosci. 1991, 105, 111. [Google Scholar] [CrossRef]
- Bonačić Marinović, A.A.; Swaan, C.; Wichmann, O.; Steenbergen, J.V.; Kretzschmar, M. Effectiveness and timing of vaccination during school measles outbreak. Emerg. Infect. Dis. 2009, 18, 1405–1413. [Google Scholar] [CrossRef]
- Ejima, K.; Omori, R.; Aihara, K.; Nishiura, H. Real-time investigation of measles epidemics with estimate of vaccine efficacy. Int. J. Biol. Sci. 2012, 8, 620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babad, H.R.; Nokes, D.J.; Gay, N.J.; Miller, E.; Morgan-Capner, P.; Anderson, R.M. Predicting the impact of measles vaccination in England and Wales: Model validation and analysis of policy options. Epidemiol. Infect. 1995, 114, 319–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lessler, J.; Metcalf, C.J.E.; Grais, R.F.; Luquero, F.J.; Cummings, D.A.; Grenfell, B.T. Measuring the Performance of Vaccination Programs Using Cross-Sectional Surveys: A Likelihood Framework and Retrospective Analysis. PLoS Med. 2011, 8, e1001110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Public Health Agency of Canada. Measles. Available online: http://www.phac-aspc.gc.ca/im/vpd-mev/measles-rougeole-eng.php (accessed on 12 July 2020).
- Perez, L.; Dragicevic, S. An Agent-based Approach for Modeling Dynamics of Contagious Disease Spread. Int. J. Health Geogr. 2009, 8, 50. [Google Scholar] [CrossRef] [Green Version]
- Glasser, J.W.; Feng, Z.; Omer, S.B.; Smith, P.J.; Rodewald, L.E. The effect of heterogeneity in uptake of the measles, mumps, and rubella vaccine on the potential for outbreaks of measles: A modelling study. Lancet Infect. Dis. 2016, 16, 599–605. [Google Scholar] [CrossRef] [Green Version]
- Wallinga, J.; Heijne, J.C.M.; Kretzschmar, M. A Measles Epidemic Threshold in a Highly Vaccinated Population. PLoS Med. 2005, 2, e316. [Google Scholar] [CrossRef] [PubMed]
- Kutty, P.; Rota, J.; Bellini, W. Manual for the Surveillance of Vaccine-Preventable Diseases, 6th ed.; Centers for Disease Control and Prevention: Atlanta, GA, USA, 2013; Chapter 7 Measles. Available online: http://www.cdc.gov/vaccines/pubs/surv-manual/chpt07-measles.html (accessed on 12 April 2020).
- Marx, G.E.; Chase, J.; Jasperse, J.; Stinson, K.; McDonald, C.E.; Runfola, J.K.; Jaskunas, J.; Hite, D.; Barnes, M.; Askenazi, M.; et al. Public Health Economic Burden Associated with Two Single Measles Case Investigations—Colorado, 2016–2017. Morb. Mortal. Wkly. Rep. 2017, 66, 1272–1275. [Google Scholar] [CrossRef] [Green Version]
- National Center for Education. Available online: https://nces.ed.gov/programs/digest/d19/tables/dt19_214.40.asp (accessed on 17 October 2020).
- Malone, K.M.; Hinman, A.R. Vaccination mandates: The public health imperative and individual rights. In Law in Public Health Practice, 2nd ed.; Goodman, R.A., Hoofman, R.E., Lopez, W., Matthews, G.W., Rothstein, M.A., Foster, K.L., Eds.; Oxford University Press: Oxford, NY, USA, 2007; pp. 338–360. [Google Scholar]
- Woudenberg, T.; Woonink, F.; Kerkhof, J.; Cox, K.; Ruijs, W.L.M.; van Binnendijk, R.; de Melker, H.; Hahné, S.J.M.; Wallinga, J. The tip of the iceberg: Incompleteness of measles reporting during a large outbreak in The Netherlands in 2013–2014. Epidemiol. Infect. 2018, 147, e23. [Google Scholar] [CrossRef] [Green Version]
- Parent du Châtelet, I.; Waku-Kouomou, D.; Freymuth, F.; Maine, C.; Lévy-Bruhl, D. La rougeole en France en 2008: Bilan de la déclaration obligatoire. [Measles in France in 2008: Results of the mandatory declaration]. French. Institut de Veille Sanitaire. Bull. Epidemiol. Heb. 2009, 39–40, 415–419. Available online: http://www.invs.sante.fr/beh/2009/39_40/beh_39_40_2009.pdf (accessed on 16 October 2020).
- CDC. Measles, Mumps, and Rubella (MMR) Vaccination Coverage Among Children 19–35 Months by State, HHS Region, and the United States, National Immunization Survey-Child (NIS-Child), 1995 through 2017. Available online: https://www.cdc.gov/vaccines/imz-managers/coverage/childvaxview/data-reports/mmr/trend/index.html (accessed on 16 October 2020).
- Azimi, P.; Keshavarz, Z.; Cedeno Laurent, J.G.; Allen, J.G. Estimating the nationwide transmission risk of measles in US schools and impacts of vaccination and supplemental infection control strategies. BMC Infect. Dis. 2020, 20, 497. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population, n |
| Fraction unvaccinated, v |
| Susceptibles, S(t) |
| Individuals in day k of the incubation period, Ek(t) |
| Individuals in day k of the infectious period, Ik(t) |
| Recovered individuals, R(t) |
| Total number of infectious individuals, T(t) |
| New infections, N(t) |
| Probability a susceptible individual is infected, p(t) |
| Basic reproduction number, |
| Number of reported individuals, Z(t) |
| Probability an infectious case is reported each day, q |
| Lag between the first reported case in same subgroup and stopping, ε |
| Stopping time, τ |
| Mixing parameter, |
| Lag between the first reported case in the other subgroup and stopping, |
| Parameter | Value | Source |
|---|---|---|
| Population, n (people) | 1000 | [57] |
| Basic reproduction number, | 18 | [37] |
| Fraction unvaccinated, v (%) | 10% | [58] |
| Incubation period (days) | 10 | [51] |
| Infectious period (days) | 8 | [52] |
| Mixing parameter, (%) | 0.5 | |
| Probability of the case being reported each contagious day, q (%) | 0.1 | [54,59,60] |
| Lag between the first reported case in the subgroup stopping, ε (days) | 3 | [56] |
| Lag between the first reported case in the other subgroup and stopping, σ (days) | 5 |
| Two Community Groups | One Community | |||||
|---|---|---|---|---|---|---|
| E[C] | E[C] | |||||
| Daily reporting probability, q | 0.05 | 5.9 | 0.465 | 0.650 | 5.2 | 0.403 |
| 0.10 | 3.9 | 0.287 | 0.554 | 3.4 | 0.257 | |
| 0.15 | 3.2 | 0.199 | 0.533 | 2.8 | 0.159 | |
| 0.20 | 2.7 | 0.116 | 0.499 | 2.6 | 0.108 | |
| Lag between first case reported and simulation stopped,ε | 0 | 3.1 | 0.212 | 0.558 | 2.8 | 0.185 |
| 1 | 3.4 | 0.247 | 0.531 | 3.0 | 0.192 | |
| 2 | 3.7 | 0.262 | 0.567 | 3.4 | 0.239 | |
| 3 | 3.7 | 0.251 | 0.551 | 3.5 | 0.242 | |
| E[C] | ||||
|---|---|---|---|---|
| Fraction vaccinated, %, (1-v) | 80 | 8.3 | 0.621 | 0.823 |
| 85 | 5.9 | 0.464 | 0.717 | |
| 90 | 4.0 | 0.308 | 0.579 | |
| 95 | 2.1 | 0.076 | 0.306 | |
| Mixing parameter, | 0 | 3.4 | 0.237 | 0.000 |
| 0.25 | 3.8 | 0.285 | 0.451 | |
| 0.5 | 3.9 | 0.297 | 0.570 | |
| 0.75 | 3.9 | 0.279 | 0.659 | |
| 1 | 4.1 | 0.315 | 0.677 | |
| Daily reporting probability, q | 0.05 | 5.9 | 0.460 | 0.639 |
| 0.1 | 3.8 | 0.276 | 0.552 | |
| 0.15 | 3.1 | 0.200 | 0.525 | |
| 0.2 | 2.8 | 0.128 | 0.492 | |
| Lag between first case reported and simulation stopped,ε | 0 | 3.1 | 0.211 | 0.537 |
| 1 | 3.6 | 0.261 | 0.572 | |
| 2 | 3.8 | 0.286 | 0.556 | |
| 3 | 3.9 | 0.307 | 0.572 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yagci Sokat, K.; Armbruster, B. Modeling and Controlling Epidemic Outbreaks: The Role of Population Size, Model Heterogeneity and Fast Response in the Case of Measles. Mathematics 2020, 8, 1892. https://doi.org/10.3390/math8111892
Yagci Sokat K, Armbruster B. Modeling and Controlling Epidemic Outbreaks: The Role of Population Size, Model Heterogeneity and Fast Response in the Case of Measles. Mathematics. 2020; 8(11):1892. https://doi.org/10.3390/math8111892
Chicago/Turabian StyleYagci Sokat, Kezban, and Benjamin Armbruster. 2020. "Modeling and Controlling Epidemic Outbreaks: The Role of Population Size, Model Heterogeneity and Fast Response in the Case of Measles" Mathematics 8, no. 11: 1892. https://doi.org/10.3390/math8111892
APA StyleYagci Sokat, K., & Armbruster, B. (2020). Modeling and Controlling Epidemic Outbreaks: The Role of Population Size, Model Heterogeneity and Fast Response in the Case of Measles. Mathematics, 8(11), 1892. https://doi.org/10.3390/math8111892