Drift of Scroll Waves in a Mathematical Model of a Heterogeneous Human Heart Left Ventricle

 , , , and
, , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
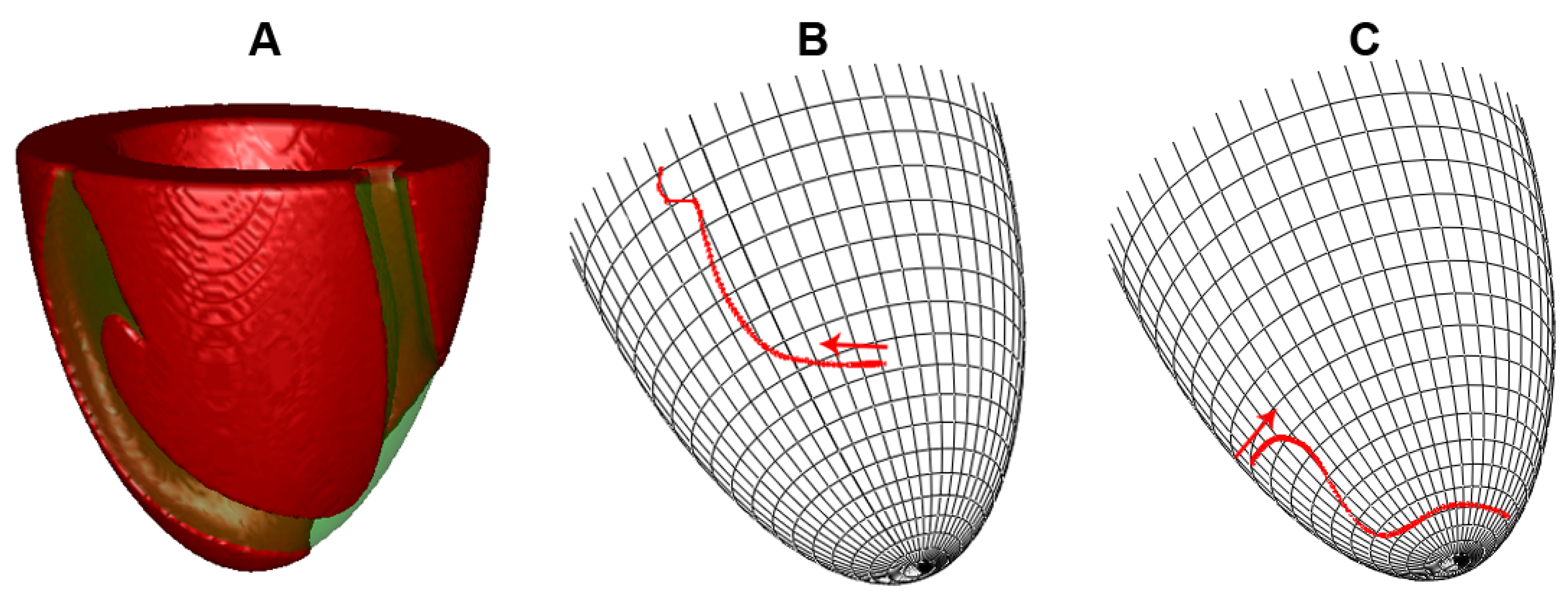
2. Materials and Methods
2.1. Baseline Homogeneous Model. Numerical Approach and Software
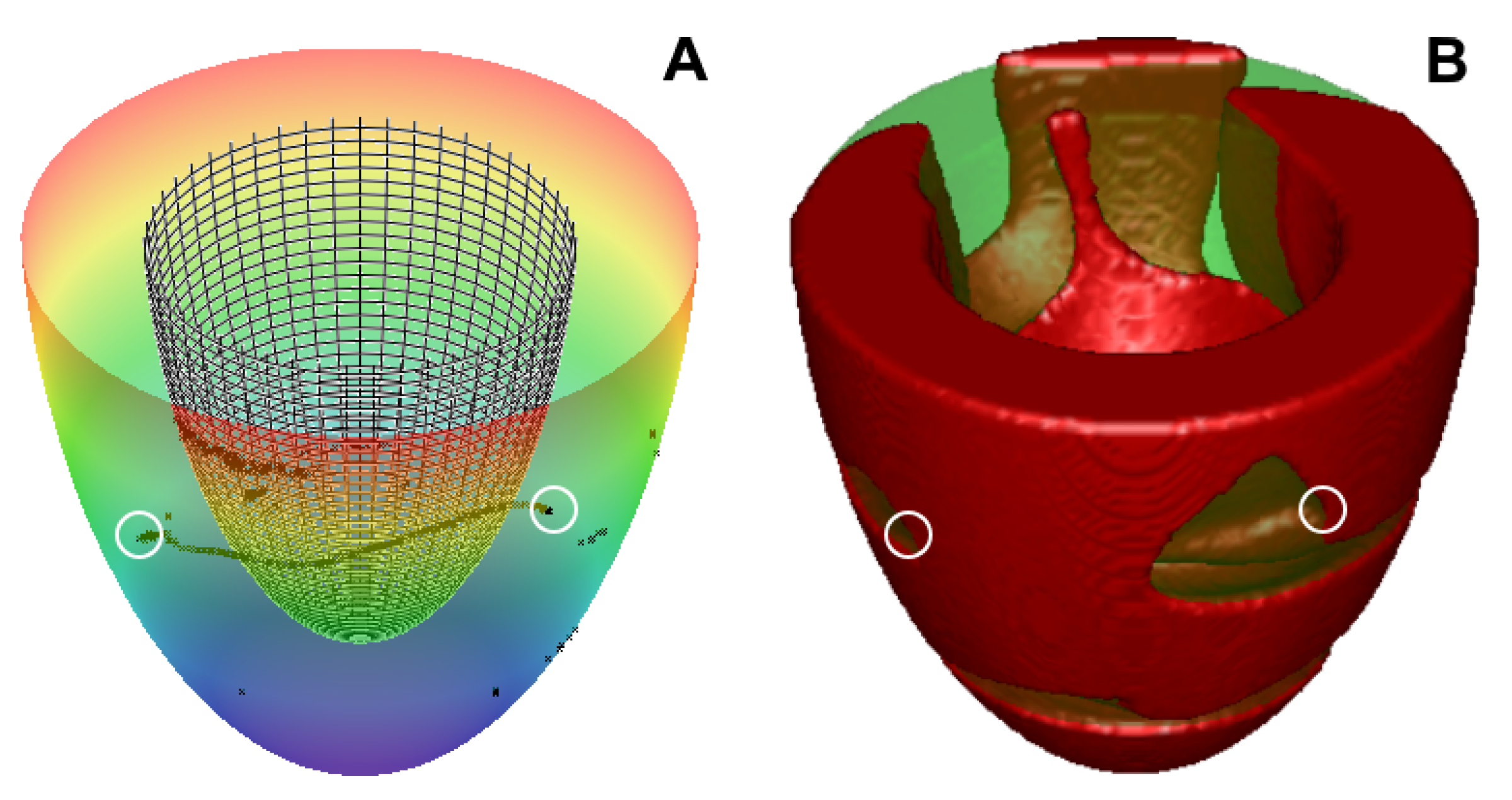
2.2. Heterogeneity Representation
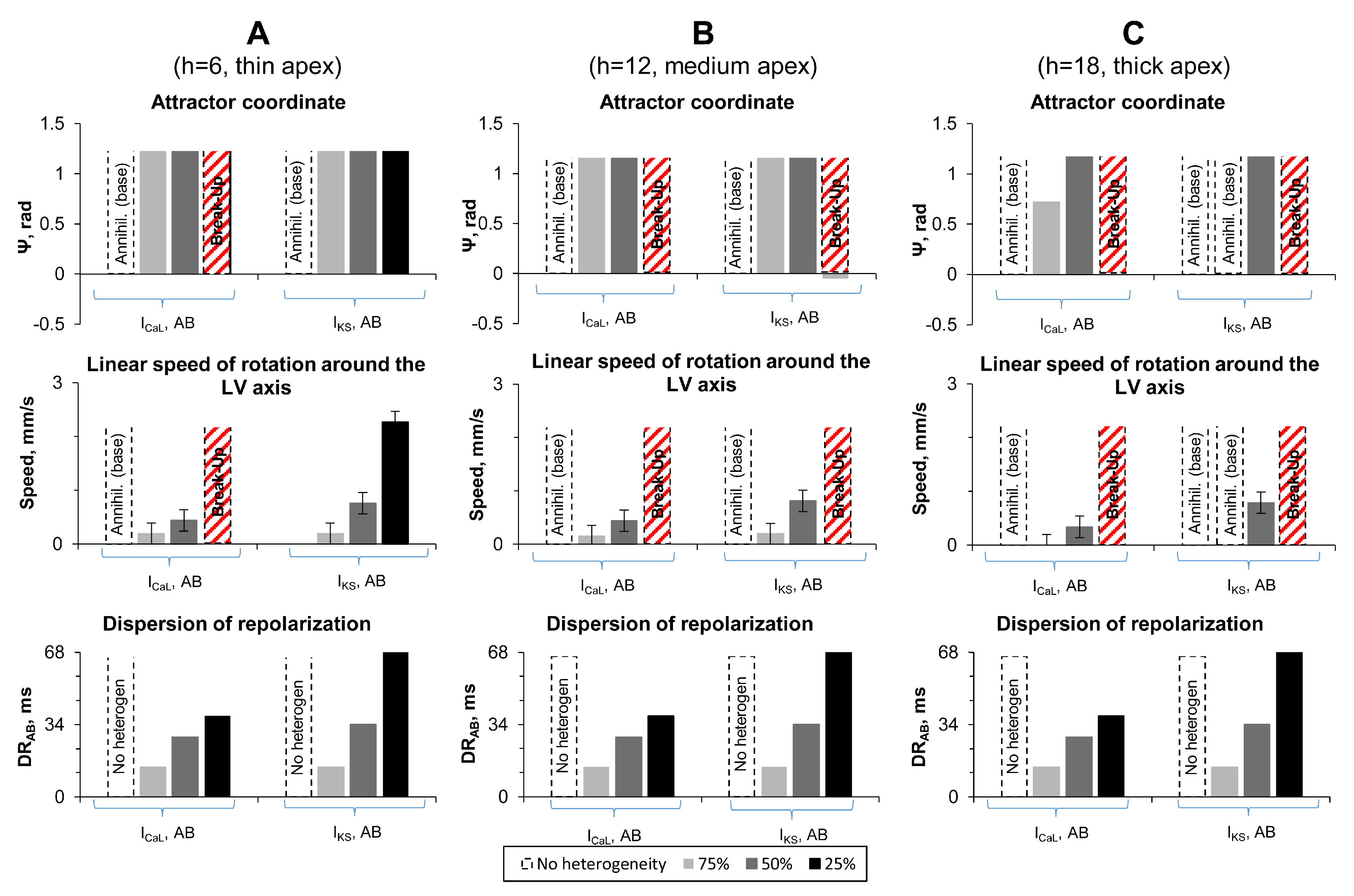
- Apicobasal heterogeneity caused by decrease of ICaL current with . We reduced ICaL current at the LV base to 75%, 50%, and 25% of its original value, which resulted in the gradients of 14, 28, and 38 ms. We denote these cases as ICaL-75, ICaL-50 and ICaL-25.
- Apicobasal heterogeneity caused by decrease of IKs current with . We reduced IKs at the apex to 75%, 50%, and 25% of its original value, which resulted in the APD gradients of 14, 34, and 68 ms. We denote these cases as IKs-75, IKs-50, and IKs-25.
3. Results
3.1. Apicobasal Heterogeneity
3.2. Transmural Heterogeneity
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Winfree, A.T.; Strogatz, S.H. Organizing centres for three-dimensional chemical waves. Nature 1984, 311, 611–615. [Google Scholar] [CrossRef] [PubMed]
- Mehra, R. Global public health problem of sudden cardiac death. J. Electrocardiol. 2007, 40, S118–S122. [Google Scholar] [CrossRef] [PubMed]
- Gray, R.A.; Jalife, J.; Panfilov, A.; Baxter, W.T.; Cabo, C.; Davidenko, J.M.; Pertsov, A.M. Nonstationary Vortexlike Reentrant Activity as a Mechanism of Polymorphic Ventricular Tachycardia in the Isolated Rabbit Heart. Circulation 1995, 91, 2454–2469. [Google Scholar] [CrossRef] [PubMed]
- Katz, A.M. Physiology of the Heart; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2010. [Google Scholar]
- Panfilov, A.; Rudenko, A.; Krinsky, V. Scroll rings in three-dimensional active medium with two component diffusion. Biofizika 1986, 31, 850–854. [Google Scholar]
- Panfilov, A.; Aliev, R.; Mushinsky, A. An integral invariant for scroll rings in a reaction-diffusion system. Phys. D Nonlinear Phenom. 1989, 36, 181–188. [Google Scholar] [CrossRef]
- Pravdin, S.; Dierckx, H.; Markhasin, V.S.; Panfilov, A.V. Drift of Scroll Wave Filaments in an Anisotropic Model of the Left Ventricle of the Human Heart. BioMed Res. Int. 2015, 2015. [Google Scholar] [CrossRef] [Green Version]
- Rogers, J.M.; McCulloch, A.D. Nonuniform muscle fiber orientation causes spiral wave drift in a finite element model of cardiac action potential propagation. J. Cardiovasc. Electrophysiol. 1994, 5, 496–509. [Google Scholar] [CrossRef]
- Dierckx, H.; Bernus, O.; Verschelde, H. A geometric theory for scroll wave filaments in anisotropic excitable media. Phys. D Nonlinear Phenom. 2009, 238, 941–950. [Google Scholar] [CrossRef]
- Nabauer, M.; Beuckelmann, D.J.; Uberfuhr, P.; Steinbeck, G. Regional Differences in Current Density and Rate-Dependent Properties of the Transient Outward Current in Subepicardial and Subendocardial Myocytes of Human Left Ventricle. Circulation 1996, 93, 168–177. [Google Scholar] [CrossRef]
- Glukhov, A.V.; Fedorov, V.V.; Lou, Q.; Ravikumar, V.K.; Kalish, P.W.; Schuessler, R.B.; Moazami, N.; Efimov, I.R. Transmural Dispersion of Repolarization in Failing and Nonfailing Human Ventricle. Circ. Res. 2010, 106, 981–991. [Google Scholar] [CrossRef] [Green Version]
- Opthof, T.; Janse, M.J.; Meijborg, V.M.; Cinca, J.; Rosen, M.R.; Coronel, R. Dispersion in ventricular repolarization in the human, canine and porcine heart. Prog. Biophys. Mol. Biol. 2016, 120, 222–235. [Google Scholar] [CrossRef] [PubMed]
- Franz, M.R.; Bargheer, K.; Rafflenbeul, W.; Haverich, A.; Lichtlen, P.R. Monophasic action potential mapping in human subjects with normal electrocardiograms: Direct evidence for the genesis of the T wave. Circulation 1987, 75, 379–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janse, M.J.; Coronel, R.; Opthof, T.; Sosunov, E.A.; Anyukhovsky, E.P.; Rosen, M.R. Repolarization gradients in the intact heart: Transmural or apico-basal? Prog. Biophys. Mol. Biol. 2012, 109, 6–15. [Google Scholar] [CrossRef]
- Boukens, B.J.; Sulkin, M.S.; Gloschat, C.R.; Ng, F.S.; Vigmond, E.J.; Efimov, I.R. Transmural APD gradient synchronizes repolarization in the human left ventricular wall. Cardiovasc. Res. 2015, 108, 188–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szentadrassy, N.; Banyasz, T.; Biro, T.; Szabo, G.; Toth, B.I.; Magyar, J.; Lazar, J.; Varro, A.; Kovacs, L.; Nanasi, P.P. Apico-basal inhomogeneity in distribution of ion channels in canine and human ventricular myocardium. Cardiovasc. Res. 2005, 65, 851–860. [Google Scholar] [CrossRef] [Green Version]
- Solovyova, O.; Katsnelson, L.; Konovalov, P.; Kursanov, A.; Vikulova, N.; Kohl, P.; Markhasin, V. The cardiac muscle duplex as a method to study myocardial heterogeneity. Prog. Biophys. Mol. Biol. 2014, 115, 115–128. [Google Scholar] [CrossRef] [Green Version]
- Antzelevitch, C. Heterogeneity and cardiac arrhythmias: An overview. Heart Rhythm 2007, 4, 964–972. [Google Scholar] [CrossRef] [Green Version]
- Keldermann, R.H.; ten Tusscher, K.H.; Nash, M.P.; Hren, R.; Taggart, P.; Panfilov, A.V. Effect of heterogeneous APD restitution on VF organization in a model of the human ventricles. Am. J. Physiol.-Heart Circ. Physiol. 2008, 294, H764–H774. [Google Scholar] [CrossRef] [Green Version]
- Rudenko, A.; Panfilov, A. Drift and interaction of vortices in two-dimensional heterogeneous active medium. Stud. Biophys. 1983, 98, 183–188. [Google Scholar]
- Ten Tusscher, K.; Panfilov, A.V. Reentry in heterogeneous cardiac tissue described by the Luo–Rudy ventricular action potential model. Am. J. Physiol.-Heart Circ. Physiol. 2003, 284, H542–H548. [Google Scholar] [CrossRef]
- Panfilov, A.V.; Rudenko, A.N.; Pertsov, A.M. Twisted scroll waves in three- dimensional active media. Dokl. Akad. Nauk SSSR 1984, 279, 1000–1002. [Google Scholar]
- Panfilov, A.V.; Keener, J.P. Twisted scroll waves in heterogeneous excitable media. Int. J. Bifurc. Chaos 1993, 3, 445–450. [Google Scholar] [CrossRef]
- Pravdin, S.F.; Berdyshev, V.I.; Panfilov, A.V.; Katsnelson, L.B.; Solovyova, O.; Markhasin, V.S. Mathematical model of the anatomy and fibre orientation field of the left ventricle of the heart. Biomed. Eng. Online 2013, 12, 54. [Google Scholar] [CrossRef] [Green Version]
- Pravdin, S.F.; Dierckx, H.; Katsnelson, L.B.; Solovyova, O.; Markhasin, V.S.; Panfilov, A.V. Electrical wave propagation in an anisotropic model of the left ventricle based on analytical description of cardiac architecture. PLoS ONE 2014, 9. [Google Scholar] [CrossRef]
- Konovalov, P.V.; Pravdin, S.F.; Solovyova, O.E.; Panfilov, A.V. Scroll Wave dynamics in a model of the heterogeneous heart. JETP Lett. 2016, 104, 821. [Google Scholar] [CrossRef]
- Aliev, R.R.; Panfilov, A.V. A simple two-variable model of cardiac excitation. Chaos Solitons Fractals 1996, 7, 293–301. [Google Scholar] [CrossRef]
- Panfilov, A. Three-dimensional organization of electrical turbulence in the heart. Phys. Rev. E 1999, 59, R6251–R6254. [Google Scholar] [CrossRef] [Green Version]
- Pravdin, S.F.; Dierckx, H.; Panfilov, A.V. Drift of scroll waves in a generic axisymmetric model of the cardiac left ventricle. Chaos Solitons Fractals 2019, 120, 222–233. [Google Scholar] [CrossRef]
- Streeter, D., Jr. The Heart. In Handbook of Physiology; Section 2; American Physiological Society: Bethesda, MD, USA, 1979; Chapter Gross Morphology and Fiber Geometry of the Heart; Volume I, pp. 61–112. [Google Scholar]
- Panfilov, A.; Holden, A. Computer simulation of re-entry sources in myocardium in two and three dimensions. J. Theor. Biol. 1993, 161, 271–285. [Google Scholar] [CrossRef]
- Ten Tusscher, K.; Panfilov, A.V. Alternans and spiral breakup in a human ventricular tissue model. Am. J. Physiol.-Heart Circ. Physiol. 2006, 291, H1088–H1100. [Google Scholar] [CrossRef]
- Kazbanov, I.V.; Clayton, R.H.; Nash, M.P.; Bradley, C.P.; Paterson, D.J.; Hayward, M.P.; Taggart, P.; Panfilov, A.V. Effect of Global Cardiac Ischemia on Human Ventricular Fibrillation: Insights from a Multi-scale Mechanistic Model of the Human Heart. PLoS Comput. Biol. 2014, 10, e1003891. [Google Scholar] [CrossRef] [PubMed]
- Rush, S.; Larsen, H. A practical algorithm for solving dynamic membrane equations. IEEE Trans. Biomed. Eng. 1978, 4, 389–392. [Google Scholar] [CrossRef] [PubMed]
- Pereon, Y.; Demolombe, S.; Baro, I.; Drouin, E.; Charpentier, F.; Escande, D. Differential expression of KvLQT1 isoforms across the human ventricular wall. Am. J. Physiol.-Heart Circ. Physiol. 2000, 278, H1908–H1915. [Google Scholar] [CrossRef] [Green Version]
- Ten Tusscher, K.; Noble, D.; Noble, P.J.; Panfilov, A.V. A model for human ventricular tissue. Am. J. Physiol.-Heart Circ. Physiol. 2004, 286, H1573–H1589. [Google Scholar] [CrossRef] [PubMed]
- Krinskii, V. Spread of excitation in an inhomogeneous medium (state similar to cardiac fibrillation). Biophysics-USSR 1966, 11, 676–683. [Google Scholar]
- Henze, C.; Lugosi, E.; Winfree, A. Helical organizing centers in excitable media. Can. J. Phys. 1990, 68, 683–710. [Google Scholar] [CrossRef]
- Papadimitriou, F. Geo-mathematical modeling of spatial-ecological complex systems: An evaluation. Geogr. Environ. Sustain. 2010, 3, 67–80. [Google Scholar] [CrossRef]
- Biktashev, V.; Holden, A.; Zhang, H. Tension of organizing filaments of scroll waves. Philos. Trans. R. Soc. London. Ser. A Phys. Eng. Sci. 1994, 347, 611–630. [Google Scholar]
- Panfilov, A.V.; Hogeweg, P. Mechanisms of cardiac fibrillation. Science 1995, 270, 1223–1224. [Google Scholar]
- Verschelde, H.; Dierckx, H.; Bernus, O. Covariant stringlike dynamics of scroll wave filaments in anisotropic cardiac tissue. Phys. Rev. Lett. 2007, 99, 168104. [Google Scholar] [CrossRef]
- Cuculich, P.S.; Schill, M.R.; Kashani, R.; Mutic, S.; Lang, A.; Cooper, D.; Faddis, M.; Gleva, M.; Noheria, A.; Smith, T.W.; et al. Noninvasive cardiac radiation for ablation of ventricular tachycardia. N. Engl. J. Med. 2017, 377, 2325–2336. [Google Scholar] [CrossRef] [PubMed]
- Arevalo, H.J.; Vadakkumpadan, F.; Guallar, E.; Jebb, A.; Malamas, P.; Wu, K.C.; Trayanova, N.A. Arrhythmia risk stratification of patients after myocardial infarction using personalized heart models. Nat. Commun. 2016, 7, 11437. [Google Scholar] [CrossRef] [PubMed]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pravdin, S.; Konovalov, P.; Dierckx, H.; Solovyova, O.; Panfilov, A.V. Drift of Scroll Waves in a Mathematical Model of a Heterogeneous Human Heart Left Ventricle. Mathematics 2020, 8, 776. https://doi.org/10.3390/math8050776
Pravdin S, Konovalov P, Dierckx H, Solovyova O, Panfilov AV. Drift of Scroll Waves in a Mathematical Model of a Heterogeneous Human Heart Left Ventricle. Mathematics. 2020; 8(5):776. https://doi.org/10.3390/math8050776
Chicago/Turabian StylePravdin, Sergey, Pavel Konovalov, Hans Dierckx, Olga Solovyova, and Alexander V. Panfilov. 2020. "Drift of Scroll Waves in a Mathematical Model of a Heterogeneous Human Heart Left Ventricle" Mathematics 8, no. 5: 776. https://doi.org/10.3390/math8050776
APA StylePravdin, S., Konovalov, P., Dierckx, H., Solovyova, O., & Panfilov, A. V. (2020). Drift of Scroll Waves in a Mathematical Model of a Heterogeneous Human Heart Left Ventricle. Mathematics, 8(5), 776. https://doi.org/10.3390/math8050776