

Therapeutic Effects of Omentin-1 on Pulmonary Fibrosis by Attenuating Fibroblast Activation via AMP-Activated Protein Kinase Pathway
 ,
,
Abstract
:1. Background
2. Methods
2.1. Experimental Animals and Treatment
2.2. Histopathology
2.3. Fibroblasts Isolation and Culture
2.4. RNA Isolation and Quantitative Polymerase Chain Reaction (qPCR)
2.5. Western Blotting
2.6. Hydroxyproline Assay
2.7. Immunohistochemistry
2.8. Enzyme-Linked Immunosorbent Assay
2.9. Immunofluorescence Staining
2.10. Flow Cytometry for Detecting of α-SMA Level
2.11. Statistical Analysis
3. Results
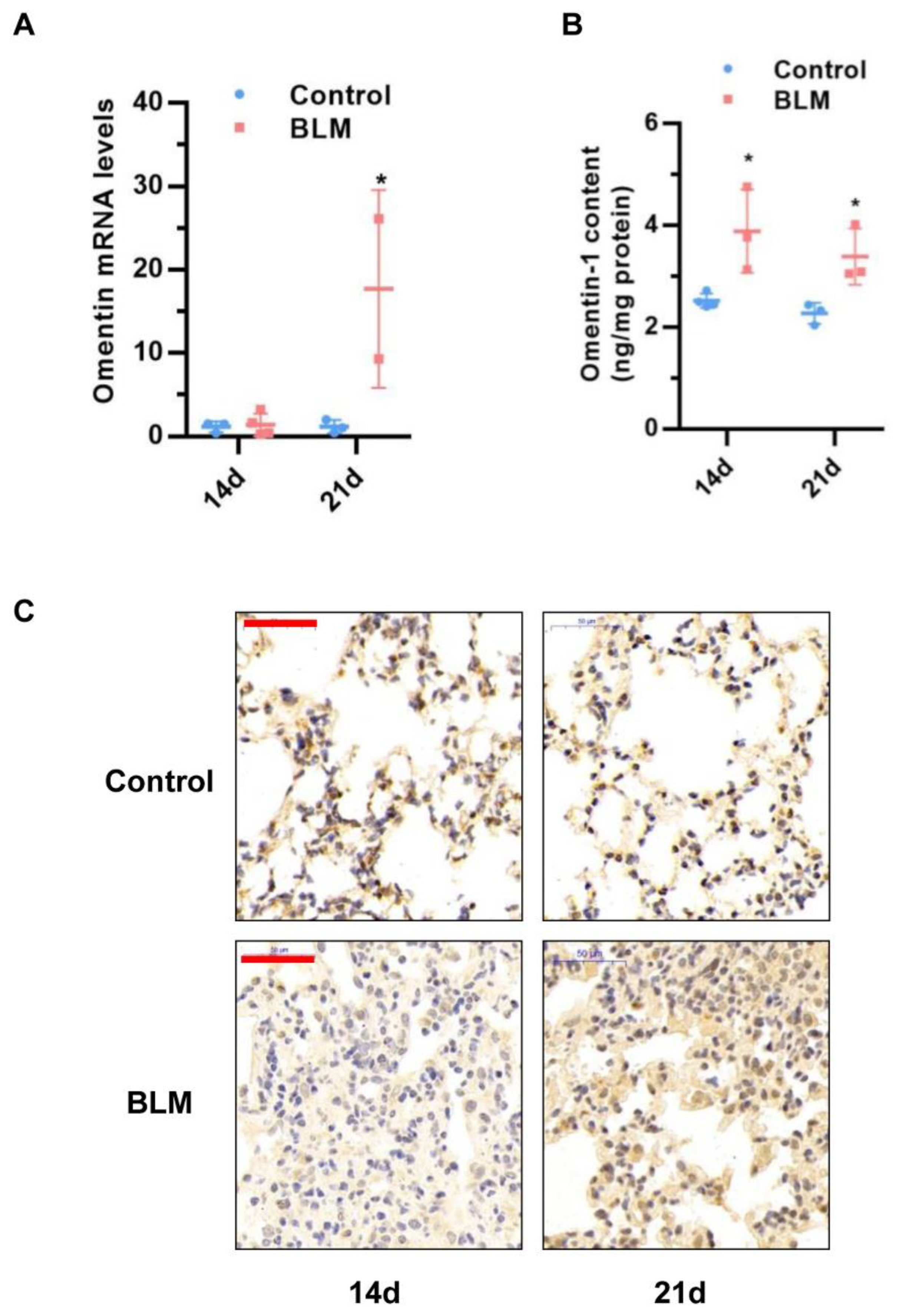
3.1. Omentin-1 Expression Increases during the Late Stages of Lung Fibrosis in Experimental Models
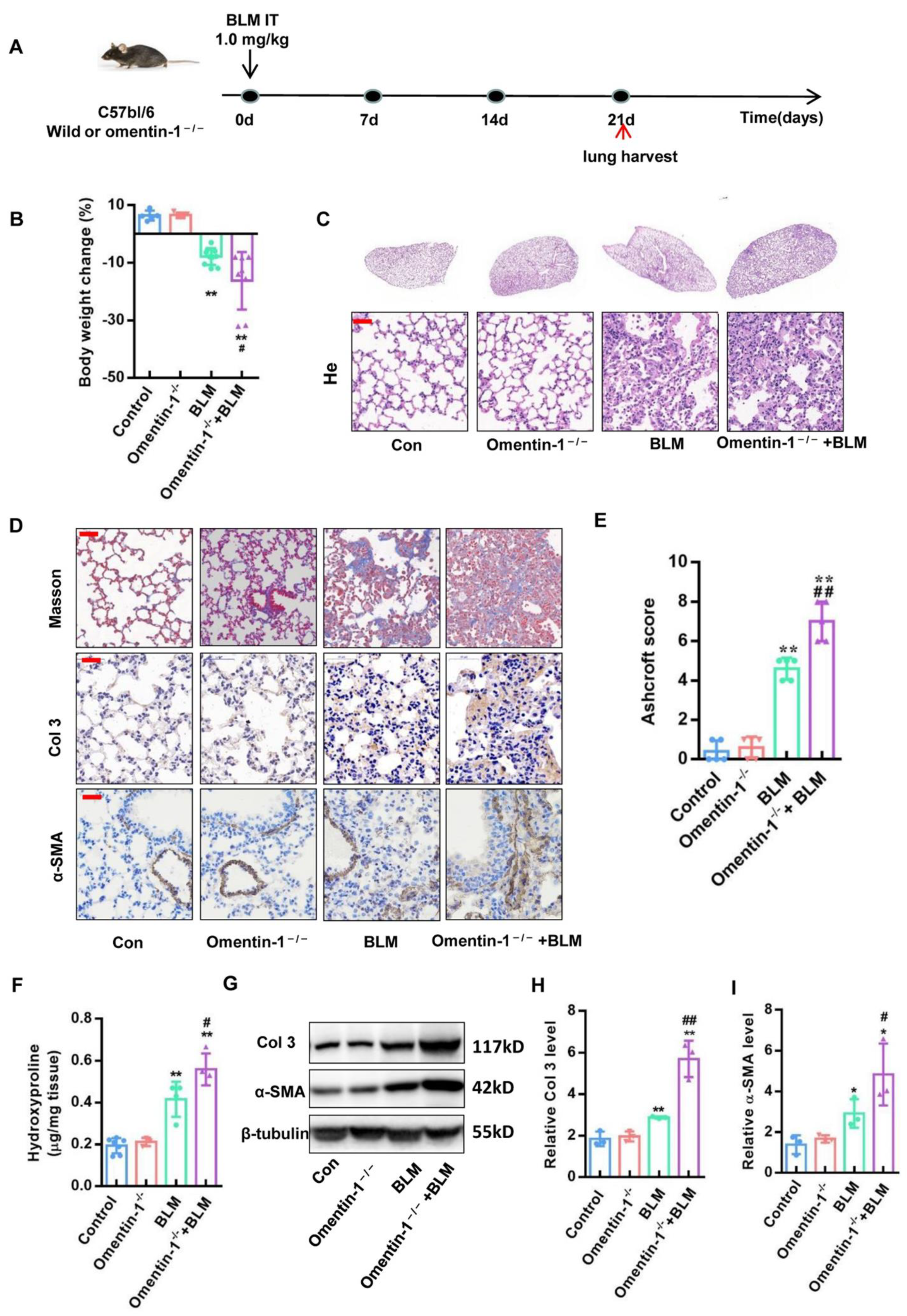
3.2. Omentin-1-Deficient Mice Show Exaggerated Lung Fibrotic Responses
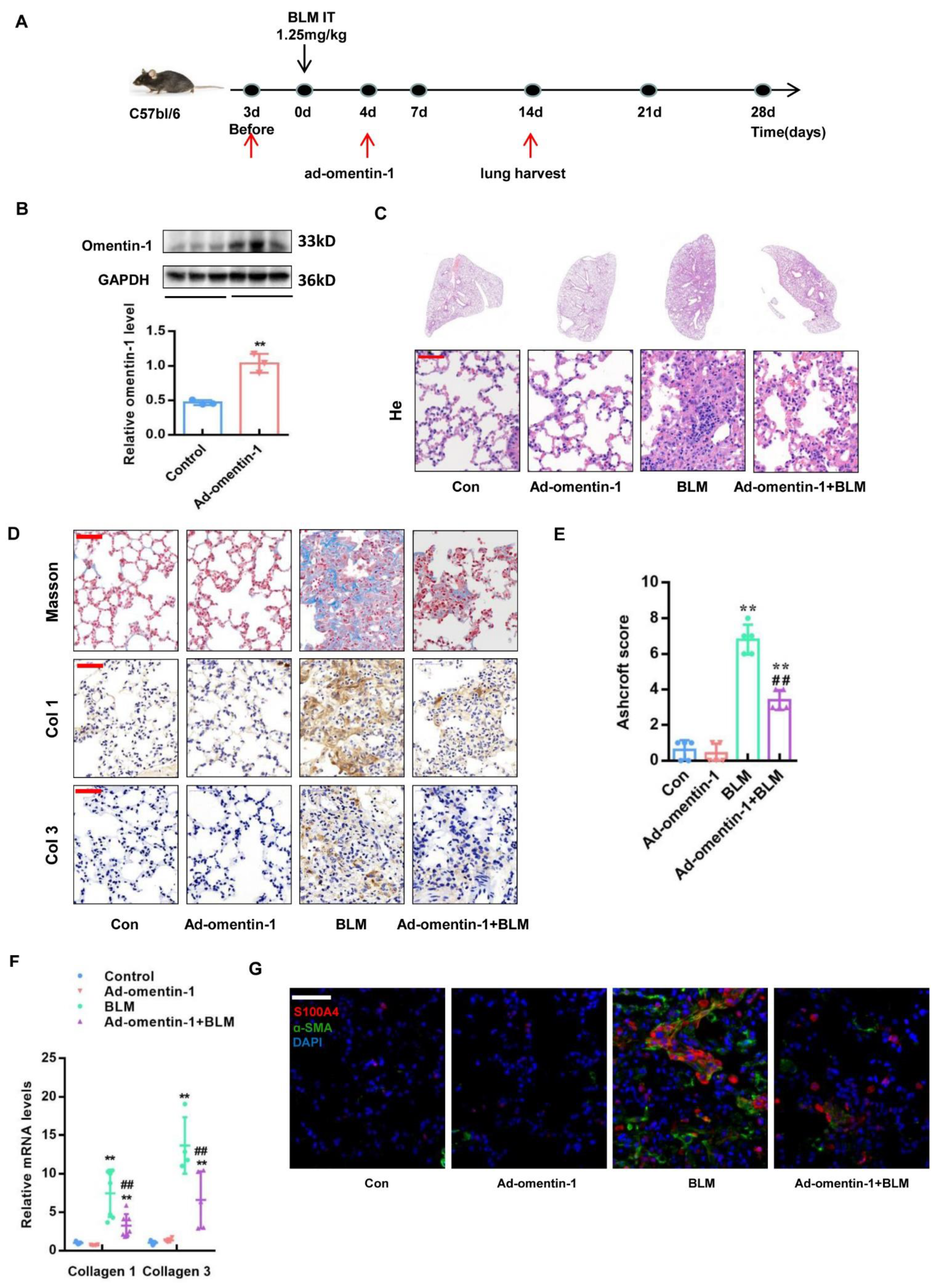
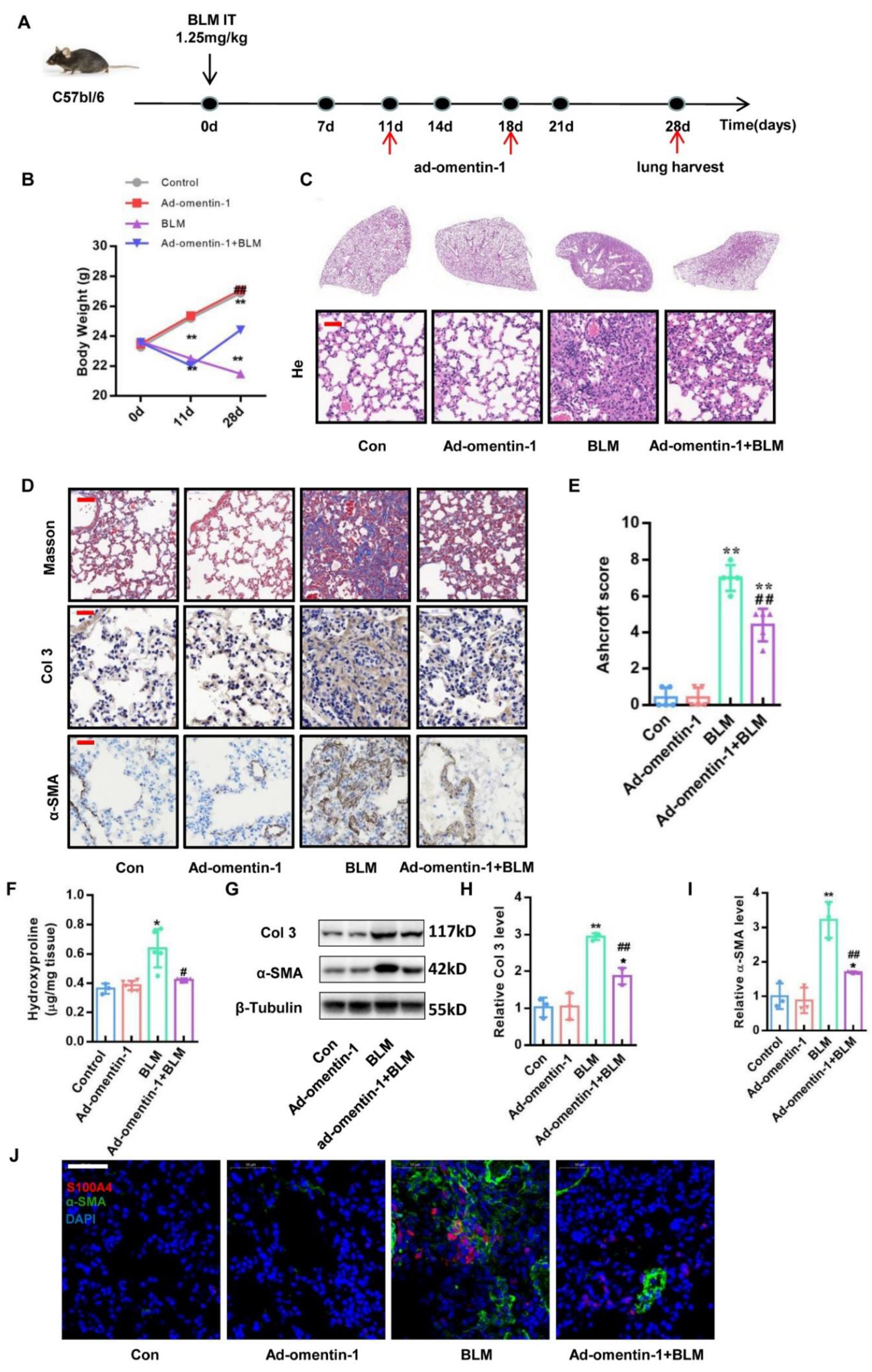
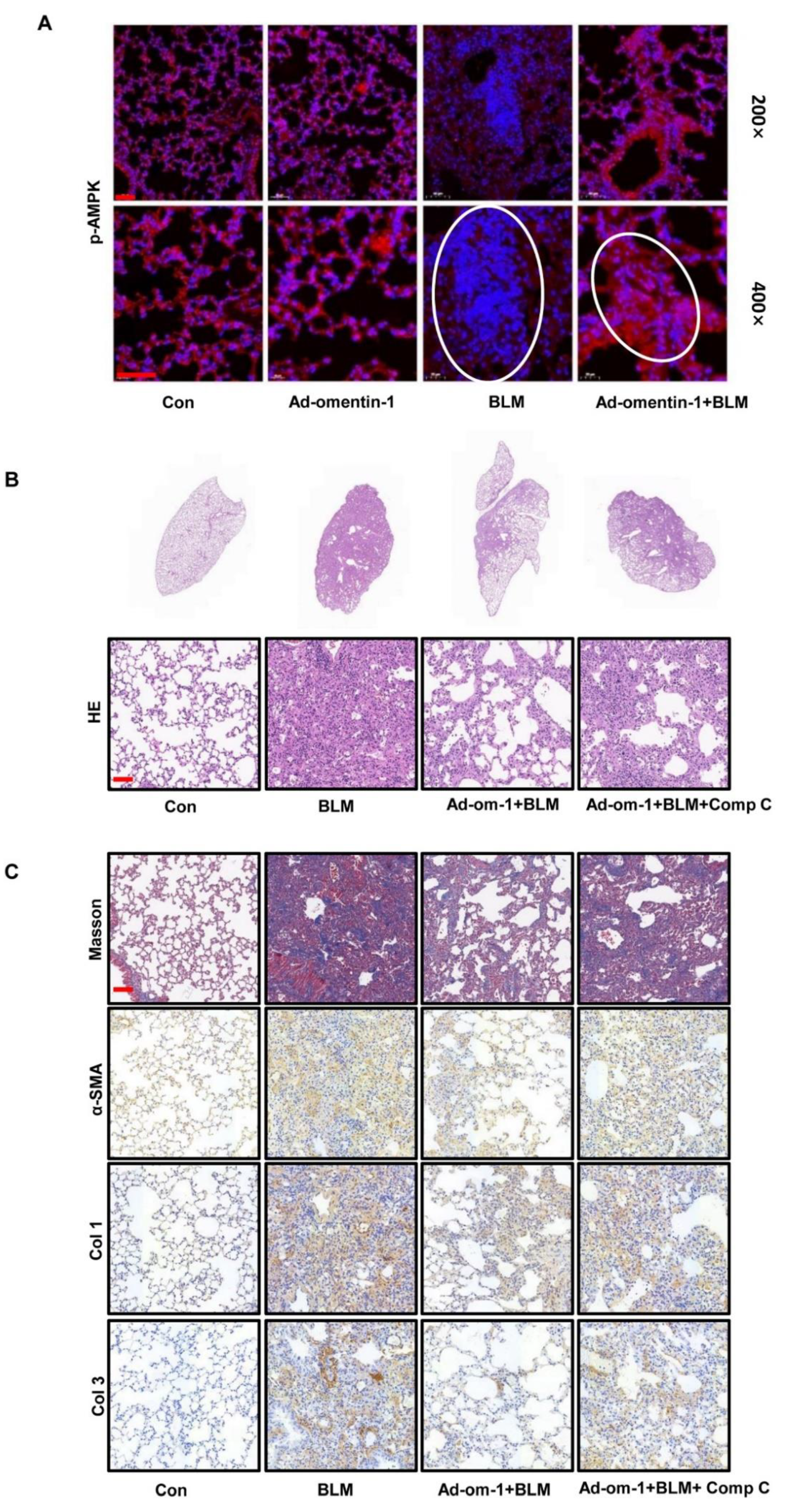
3.3. Omentin-1 Overexpression Attenuates BLM-Induced Lung Fibrosis with Preventative and Therapeutic Treatment
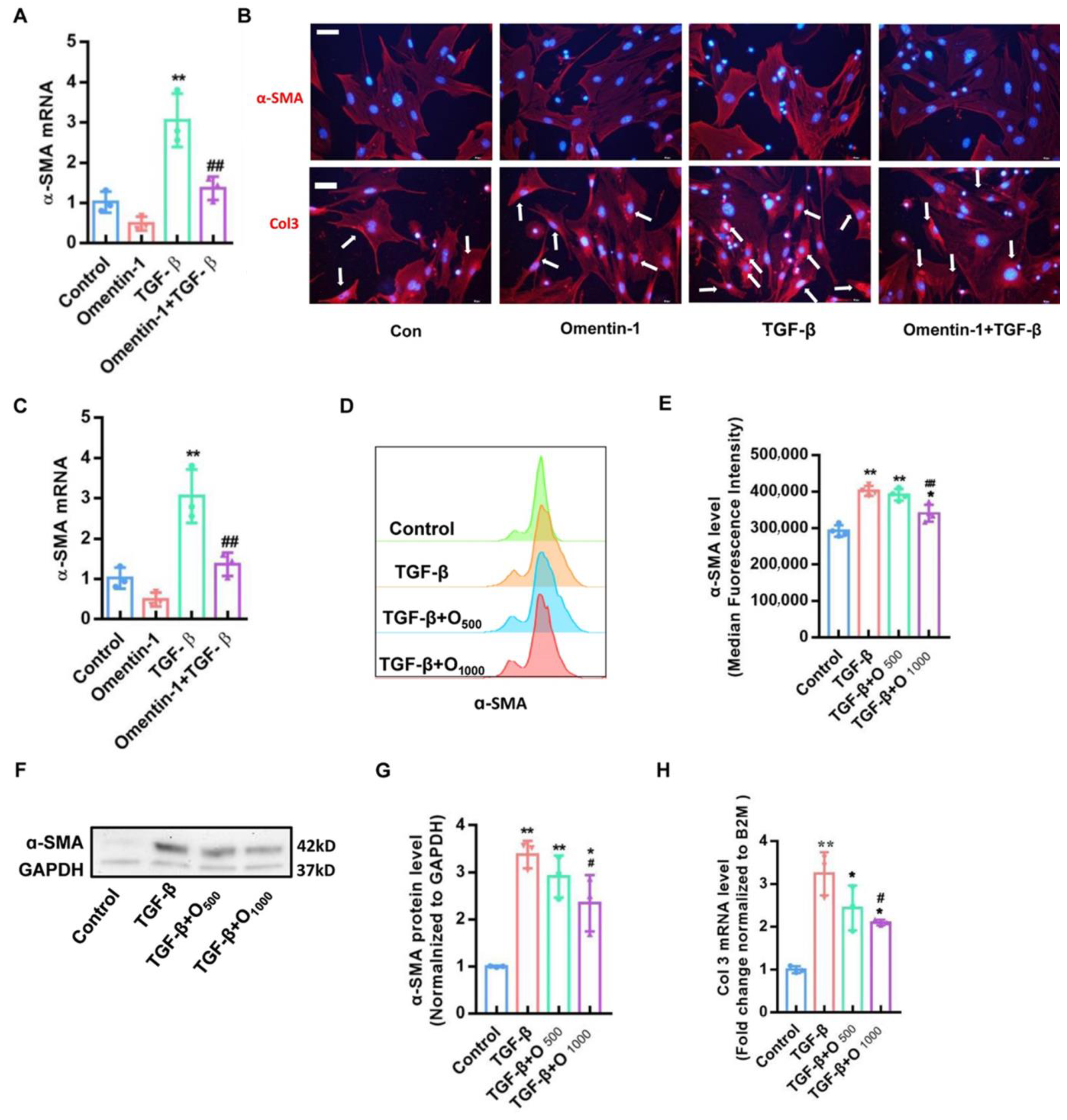
3.4. Recombinant Omentin-1 Attenuates TGF-β-Induced Myofibroblast Differentiation In Vitro
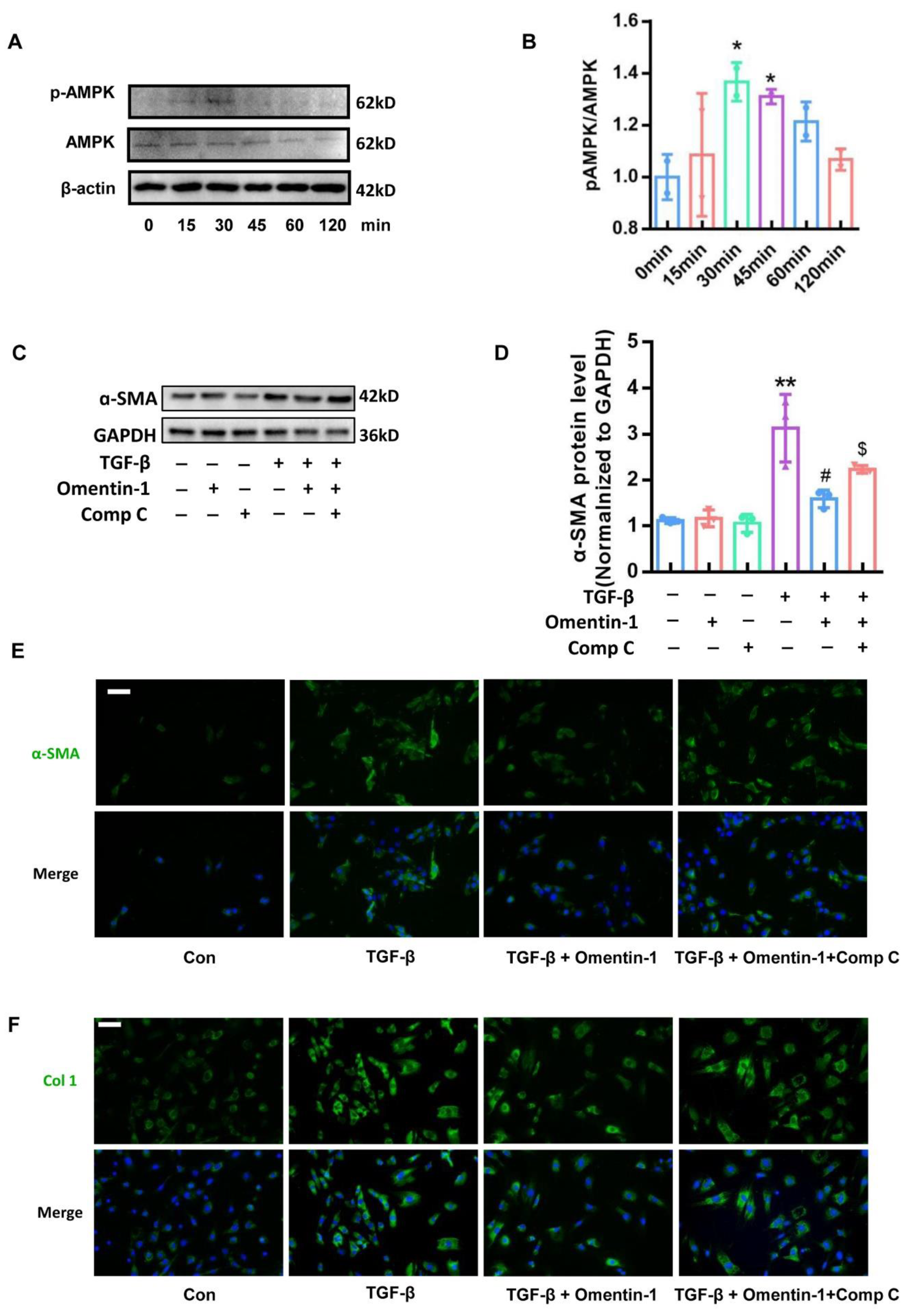
3.5. Omentin-1 Attenuates Lung Fibrosis via Activation of AMPK Pathway
3.6. Omentin-1 Attenuates TGF-β-Induced Fibroblast Activation via AMPK Pathway
4. Discussion
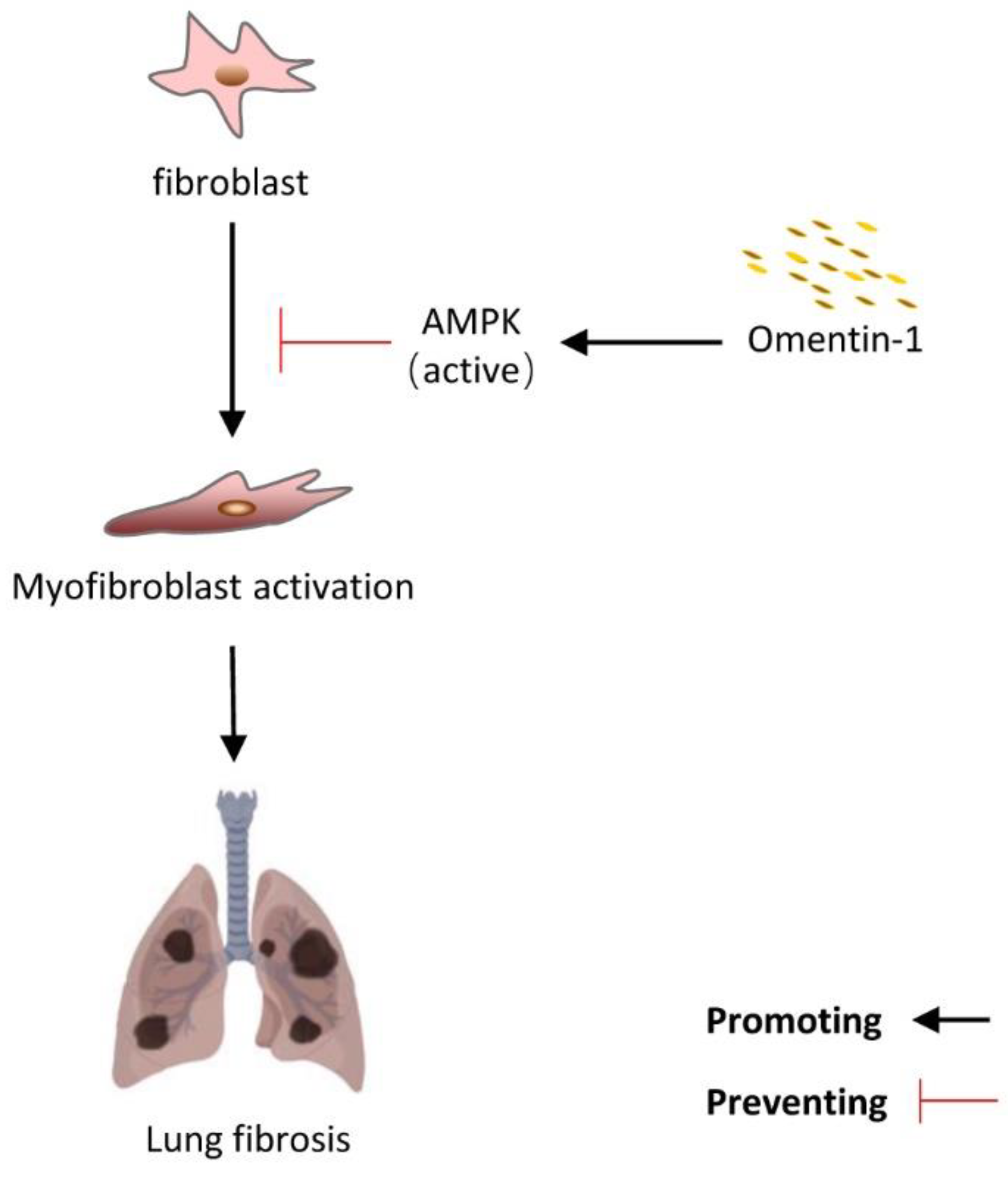
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ad-omentin-1 | Adenovirus omentin-1 |
| AMPK | AMP-activated protein kinase |
| ATP | Adenosine Triphosphate |
| α-SMA | α -smooth muscle actin |
| BLM | Bleomycin |
| B2M | β2 microglobulin |
| CT | computed tomography |
| Comp C | Compound C |
| ECM | extracellular matrix |
| GAPDH | Glyceraldehyde-3-phosphate dehydrogenase |
| H&E | Hematoxylin and eosin |
| HYP | hydroxyproline |
| IPF | idiopathic pulmonary fibrosis |
| SARS-CoV-2 | Severe acute respiratory syndrome coronavirus 2 |
| TGF-β1 | transforming growth factor beta 1 |
| omentin-1−/− | omentin-1-deficient |
| p-AMPK | phosphorylation of AMPK |
| qPCR | quantitative Polymerase Chain Reaction |
| WB | Western blotting |
| WT | Wild Type |
References
- Lederer, D.J.; Martinez, F.J. Idiopathic Pulmonary Fibrosis. N. Engl. J. Med. 2018, 378, 1811–1823. [Google Scholar] [CrossRef] [PubMed]
- Distler, O.; Highland, K.B.; Gahlemann, M.; Azuma, A.; Fischer, A.; Mayes, M.D.; Raghu, G.; Sauter, W.; Girard, M.; Alves, M.; et al. Nintedanib for Systemic Sclerosis–Associated Interstitial Lung Disease. N. Engl. J. Med. 2019, 380, 2518–2528. [Google Scholar] [CrossRef] [PubMed]
- King, T.E., Jr.; Bradford, W.Z.; Castro-Bernardini, S.; Fagan, E.A.; Glaspole, I.; Glassberg, M.K.; Gorina, E.; Hopkins, P.M.; Kardatzke, D.; Lancaster, L.; et al. A Phase 3 Trial of Pirfenidone in Patients with Idiopathic Pulmonary Fibrosis. New Engl. J. Med. 2014, 370, 2083–2092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kropski, J.A.; Blackwell, T.S. Progress in Understanding and Treating Idiopathic Pulmonary Fibrosis. Annu. Rev. Med. 2019, 70, 211–224. [Google Scholar] [CrossRef]
- Hinz, B.; Lagares, D. Evasion of apoptosis by myofibroblasts: A hallmark of fibrotic diseases. Nat. Rev. Rheumatol. 2019, 16, 11–31. [Google Scholar] [CrossRef] [PubMed]
- Klingberg, F.; Hinz, B.; White, E.S. The myofibroblast matrix: Implications for tissue repair and fibrosis. J. Pathol. 2012, 229, 298–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jun, J.-I.; Lau, L.F. Resolution of organ fibrosis. J. Clin. Investig. 2018, 128, 97–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schäffler, A.; Neumeier, M.; Herfarth, H.; Fürst, A.; Schölmerich, J.; Büchler, C. Genomic structure of human omentin, a new adipocytokine expressed in omental adipose tissue. Biochim. Biophys. Acta (BBA)-Gene Struct. Expr. 2005, 1732, 96–102. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhang, B.; Hao, C.; Huang, X.; Li, X.; Huang, Y.; Luo, Z. Omentin-A Novel Adipokine in Respiratory Diseases. Int. J. Mol. Sci. 2017, 19, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kazama, K.; Okada, M.; Yamawaki, H. Adipocytokine, omentin inhibits doxorubicin-induced H9c2 cardiomyoblasts apoptosis through the inhibition of mitochondrial reactive oxygen species. Biochem. Biophys. Res. Commun. 2015, 457, 602–607. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Huang, D.; Liu, X.; Wang, Y.; Liu, J.; Liu, F.; Yu, B. Omentin-1 effects on mesenchymal stem cells: Proliferation, apoptosis, and angiogenesis in vitro. Stem Cell Res. Ther. 2017, 8, 224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, K.; Watanabe, R.; Konii, H.; Shirai, R.; Sato, K.; Matsuyama, T.-A.; Ishibashi-Ueda, H.; Koba, S.; Kobayashi, Y.; Hirano, T.; et al. Counteractive effects of omentin-1 against atherogenesis. Cardiovasc. Res. 2016, 110, 118–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, D.; Tang, X.; He, J.; Wang, D.; Zhao, Y.; Deng, W.; Deng, X.; Zhou, G.; Xia, J.; Zhong, X.; et al. Omentin protects against LPS-induced ARDS through suppressing pulmonary inflammation and promoting endothelial barrier via an Akt/eNOS-dependent mechanism. Cell Death Dis. 2016, 7, e2360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, T.; Watanabe-Kominato, K.; Takahashi, Y.; Kojima, M.; Watanabe, R. Adipose Tissue-Derived Omentin-1 Function and Regulation. Compr. Pshysiol. 2017, 7, 765–781. [Google Scholar] [CrossRef]
- Herzig, S.; Shaw, R.J. AMPK: Guardian of metabolism and mitochondrial homeostasis. Nat. Rev. Mol. Cell Biol. 2018, 19, 121–135. [Google Scholar] [CrossRef] [Green Version]
- Hawley, S.A.; Davison, M.; Woods, A.; Davies, S.P.; Beri, R.K.; Carling, D.; Hardie, D.G. Characterization of the AMP-activated Protein Kinase Kinase from Rat Liver and Identification of Threonine 172 as the Major Site at Which It Phosphorylates AMP-activated Protein Kinase. J. Biol. Chem. 1996, 271, 27879–27887. [Google Scholar] [CrossRef] [Green Version]
- Rangarajan, S.; Bone, N.B.; Zmijewska, A.A.; Jiang, S.; Park, D.W.; Bernard, K.; Locy, M.L.; Ravi, S.; Deshane, J.; Mannon, R.B.; et al. Author Correction: Metformin reverses established lung fibrosis in a bleomycin model. Nat. Med. 2018, 24, 1627. [Google Scholar] [CrossRef] [Green Version]
- Gu, X.; Han, Y.-Y.; Yang, C.-Y.; Ji, H.-M.; Lan, Y.-J.; Bi, Y.-Q.; Zheng, C.; Qu, J.; Cheng, M.-H.; Gao, J. Activated AMPK by metformin protects against fibroblast proliferation during pulmonary fibrosis by suppressing FOXM1. Pharmacol. Res. 2021, 173, 105844. [Google Scholar] [CrossRef]
- Willette, R.N.; Mangrolia, P.; Pondell, S.M.; Lee, C.Y.W.; Yoo, S.; Rudoltz, M.S.; Cowen, B.R.; Welsch, D.J. Modulation of Oxidative Phosphorylation with IM156 Attenuates Mitochondrial Metabolic Reprogramming and Inhibits Pulmonary Fibrosis. J. Pharmacol. Exp. Ther. 2021, 379, 290–300. [Google Scholar] [CrossRef]
- Li, R.; Xu, G.-H.; Cao, J.; Liu, B.; Xie, H.-F.; Ishii, Y.; Zhang, C.-F. Alpha-Mangostin Ameliorates Bleomycin-Induced Pulmonary Fibrosis in Mice Partly Through Activating Adenosine 5′-Monophosphate-Activated Protein Kinase. Front. Pharmacol. 2019, 10, 1305. [Google Scholar] [CrossRef]
- Yang, J.-Y.; Tao, L.-J.; Liu, B.; You, X.-Y.; Zhang, C.-F.; Xie, H.-F.; Li, R. Wedelolactone Attenuates Pulmonary Fibrosis Partly Through Activating AMPK and Regulating Raf-MAPKs Signaling Pathway. Front. Pharmacol. 2019, 10, 151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Zhang, Y.; Tian, F.; Wang, Z.; Song, H.; Chen, H.; Wu, B. Omentin-1 promotes mitochondrial biogenesis via PGC1α-AMPK pathway in chondrocytes. Arch. Physiol. Biochem. 2020, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Li, X.; Zhang, J.; Zhang, Y. Omentin-1: Protective impact on ischemic stroke via ameliorating atherosclerosis. Clin. Chim. Acta 2021, 517, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Li, F.-X.; Lin, X.; Zhong, J.-Y.; Wu, F.; Shan, S.-K.; Tan, C.-M.; Yuan, L.-Q.; Liao, X.-B. Adipose tissue-derived omentin-1 attenuates arterial calcification via AMPK/Akt signaling pathway. Aging 2019, 11, 8760–8776. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Fang, S.; Liu, X.; Li, J.; Wang, X.; Cui, J.; Chen, T.; Li, Z.; Yang, F.; Tian, J.; et al. Omentin-1 protects against high glucose-induced endothelial dysfunction via the AMPK/PPARδ signaling pathway. Biochem. Pharmacol. 2020, 174, 113830. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Hao, C.; Li, C.; Huang, X.; Li, X.; Tang, Y.; Huang, Y.; Tang, S.; Liu, W.; Feng, D.; et al. Omentin-1 protects against bleomycin-induced acute lung injury. Mol. Immunol. 2018, 103, 96–105. [Google Scholar] [CrossRef]
- Liu, W.; Wan, J.; Han, J.-Z.; Li, C.; Feng, D.-D.; Yue, S.-J.; Huang, Y.-H.; Chen, Y.; Cheng, Q.-M.; Li, Y.; et al. Antiflammin-1 attenuates bleomycin-induced pulmonary fibrosis in mice. Respir. Res. 2013, 14, 101. [Google Scholar] [CrossRef] [Green Version]
- Wynn, T.A.; Ramalingam, T.R. Mechanisms of fibrosis: Therapeutic translation for fibrotic disease. Nat. Med. 2012, 18, 1028–1040. [Google Scholar] [CrossRef] [Green Version]
- Rao, S.-S.; Hu, Y.; Xie, P.-L.; Cao, J.; Wang, Z.-X.; Liu, J.-H.; Yin, H.; Huang, J.; Tan, Y.-J.; Luo, J.; et al. Omentin-1 prevents inflammation-induced osteoporosis by downregulating the pro-inflammatory cytokines. Bone Res. 2018, 6, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Zhang, B.; Li, C.; Huang, X.; Cheng, H.; Bao, X.; Zhao, F.; Cheng, Q.; Yue, S.; Han, J.; et al. Megakaryocytes participate in the occurrence of bleomycin-induced pulmonary fibrosis. Cell Death Dis. 2019, 10, 1–14. [Google Scholar] [CrossRef]
- Shahzeidi, S.; Mulier, B.; de Crombrugghe, B.; Jeffery, P.K.; McAnulty, R.J.; Laurent, G.J. Enhanced type III collagen gene expression during bleomycin induced lung fibrosis. Thorax 1993, 48, 622–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hecker, L.; Jagirdar, R.; Jin, T.; Thannickal, V.J. Reversible differentiation of myofibroblasts by MyoD. Exp. Cell Res. 2011, 317, 1914–1921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, K.; Logsdon, N.J.; Shin, Y.-J.; Palumbo, S.; Knox, A.; Irish, J.D.; Rounseville, S.P.; Rummel, S.R.; Mohamed, M.; Ahmad, K.; et al. Impaired Myofibroblast Dedifferentiation Contributes to Nonresolving Fibrosis in Aging. Am. J. Respir. Cell Mol. Biol. 2020, 62, 633–644. [Google Scholar] [CrossRef] [PubMed]
- Piera-Velazquez, S.; Jimenez, S. Endothelial to Mesenchymal Transition: Role in Physiology and in the Pathogenesis of Human Diseases. Physiol. Rev. 2019, 99, 1281–1324. [Google Scholar] [CrossRef]
- Xia, H.; Gilbertsen, A.; Herrera, J.; Racila, E.; Smith, K.; Peterson, M.; Griffin, T.; Benyumov, A.; Yang, L.; Bitterman, P.; et al. Calcium-binding protein S100A4 confers mesenchymal progenitor cell fibrogenicity in idiopathic pulmonary fibrosis. J. Clin. Investig. 2017, 127, 2586–2597. [Google Scholar] [CrossRef] [Green Version]
- Lagares, D.; Ghassemi-Kakroodi, P.; Tremblay, C.; Santos, A.; Probst, C.K.; Franklin, A.; Dos Santos, D.M.; Grasberger, P.; Ahluwalia, N.; Montesi, S.B.; et al. ADAM10-mediated ephrin-B2 shedding promotes myofibroblast activation and organ fibrosis. Nat. Med. 2017, 23, 1405–1415. [Google Scholar] [CrossRef]
- Moore, B.B.; Hogaboam, C.M. Murine models of pulmonary fibrosis. Am. J. Physiol. Cell. Mol. Physiol. 2008, 294, L152–L160. [Google Scholar] [CrossRef] [Green Version]
- Moeller, A.; Ask, K.; Warburton, D.; Gauldie, J.; Kolb, M. The bleomycin animal model: A useful tool to investigate treatment options for idiopathic pulmonary fibrosis? Int. J. Biochem. Cell Biol. 2008, 40, 362–382. [Google Scholar] [CrossRef] [Green Version]
- Carrión, M.; Frommer, K.W.; Pérez-García, S.; Müller-Ladner, U.; Gomariz, R.P.; Neumann, E. The Adipokine Network in Rheumatic Joint Diseases. Int. J. Mol. Sci. 2019, 20, 4091. [Google Scholar] [CrossRef] [Green Version]
- Luo, L.; Li, J.; Liu, H.; Jian, X.; Zou, Q.; Zhao, Q.; Le, Q.; Chen, H.; Gao, X.; He, C. Adiponectin Is Involved in Connective Tissue Growth Factor-Induced Proliferation, Migration and Overproduction of the Extracellular Matrix in Keloid Fibroblasts. Int. J. Mol. Sci. 2017, 18, 1044. [Google Scholar] [CrossRef]
- Yao, R.; Cao, Y.; He, Y.-R.; Lau, W.B.; Zeng, Z.; Liang, Z.-A. Adiponectin Attenuates Lung Fibroblasts Activation and Pulmonary Fibrosis Induced by Paraquat. PLoS ONE 2015, 10, e0125169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thakur, S.; Viswanadhapalli, S.; Kopp, J.B.; Shi, Q.; Barnes, J.L.; Block, K.; Gorin, Y.; Abboud, H.E. Activation of AMP-Activated Protein Kinase Prevents TGF-β1–Induced Epithelial-Mesenchymal Transition and Myofibroblast Activation. Am. J. Pathol. 2015, 185, 2168–2180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lagares, D.; Santos, A.; Grasberger, P.E.; Liu, F.; Probst, C.K.; Rahimi, R.A.; Sakai, N.; Kuehl, T.; Ryan, J.; Bhola, P.; et al. Targeted apoptosis of myofibroblasts with the BH3 mimetic ABT-263 reverses established fibrosis. Sci. Transl. Med. 2017, 9, eaal3765. [Google Scholar] [CrossRef] [Green Version]
- Kataoka, Y.; Shibata, R.; Ohashi, K.; Kambara, T.; Enomoto, T.; Uemura, Y.; Ogura, Y.; Yuasa, D.; Matsuo, K.; Nagata, T.; et al. Omentin Prevents Myocardial Ischemic Injury Through AMP-Activated Protein Kinase- and Akt-Dependent Mechanisms. J. Am. Coll. Cardiol. 2014, 63, 2722–2733. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer Sequence (5 to 3) | Reverse Primer Sequence (5 to 3) | Length (bp) |
|---|---|---|---|
| M-GAPDH | GAAGGTGGTGAAGCAGGCATCT | CGGCATCGAAGGTGGAAGAGTG | 116 |
| M-α-SMA | CTTCGCTGGTGATGATGCTC | GTTGGTGATGATGCCGTGTT | 175 |
| M-Omentin-1 | AGTGCAGCTGAAGAGAACCT | ACTTCCCACGCATGTTGTTC | 229 |
| M-Col1a1 | GAGCGGAGAGTACTGGATCG | GCTTCTTTTCCTTGGGGTTC | 158 |
| M-Col3 | GCACAGCAGTCCAACGTAGA | TCTCCAAATGGGATCTCTGG | 185 |
| H-B2M | TTTCATCCATCCGACATTGA | CCTCCATGATGCTGCTTACA | 228 |
| H-α-SMA | TTCAATGTCCCAGCCATGTA | GAAGGAATAGCCACGCTCAG | 222 |
| H-Col 1a1 | CCAAATCTGTCTCCCCAGAA | TCAAAAACGAAGGGGAGATG | 214 |
| H-Col3 | TACGGCAATCCTGAACTTCC | GTGTGTTTCGTGCAACCATC | 245 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, Y.; Zhang, Y.; Cheng, H.; Li, X.; Feng, D.; Yue, S.; Xu, J.; Xie, H.; Luo, Z. Therapeutic Effects of Omentin-1 on Pulmonary Fibrosis by Attenuating Fibroblast Activation via AMP-Activated Protein Kinase Pathway. Biomedicines 2022, 10, 2715. https://doi.org/10.3390/biomedicines10112715
Zhou Y, Zhang Y, Cheng H, Li X, Feng D, Yue S, Xu J, Xie H, Luo Z. Therapeutic Effects of Omentin-1 on Pulmonary Fibrosis by Attenuating Fibroblast Activation via AMP-Activated Protein Kinase Pathway. Biomedicines. 2022; 10(11):2715. https://doi.org/10.3390/biomedicines10112715
Chicago/Turabian StyleZhou, Yan, Yunna Zhang, Haipeng Cheng, Xiaohong Li, Dandan Feng, Shaojie Yue, Jianping Xu, Hui Xie, and Ziqiang Luo. 2022. "Therapeutic Effects of Omentin-1 on Pulmonary Fibrosis by Attenuating Fibroblast Activation via AMP-Activated Protein Kinase Pathway" Biomedicines 10, no. 11: 2715. https://doi.org/10.3390/biomedicines10112715
APA StyleZhou, Y., Zhang, Y., Cheng, H., Li, X., Feng, D., Yue, S., Xu, J., Xie, H., & Luo, Z. (2022). Therapeutic Effects of Omentin-1 on Pulmonary Fibrosis by Attenuating Fibroblast Activation via AMP-Activated Protein Kinase Pathway. Biomedicines, 10(11), 2715. https://doi.org/10.3390/biomedicines10112715