
Pathophysiology of Perinatal Asphyxia in Humans and Animal Models

 ,
,  , and
, and
Abstract
:1. Introduction
2. Defining Birth Asphyxia
3. Circulatory Changes during Labor and Neonatal Transition
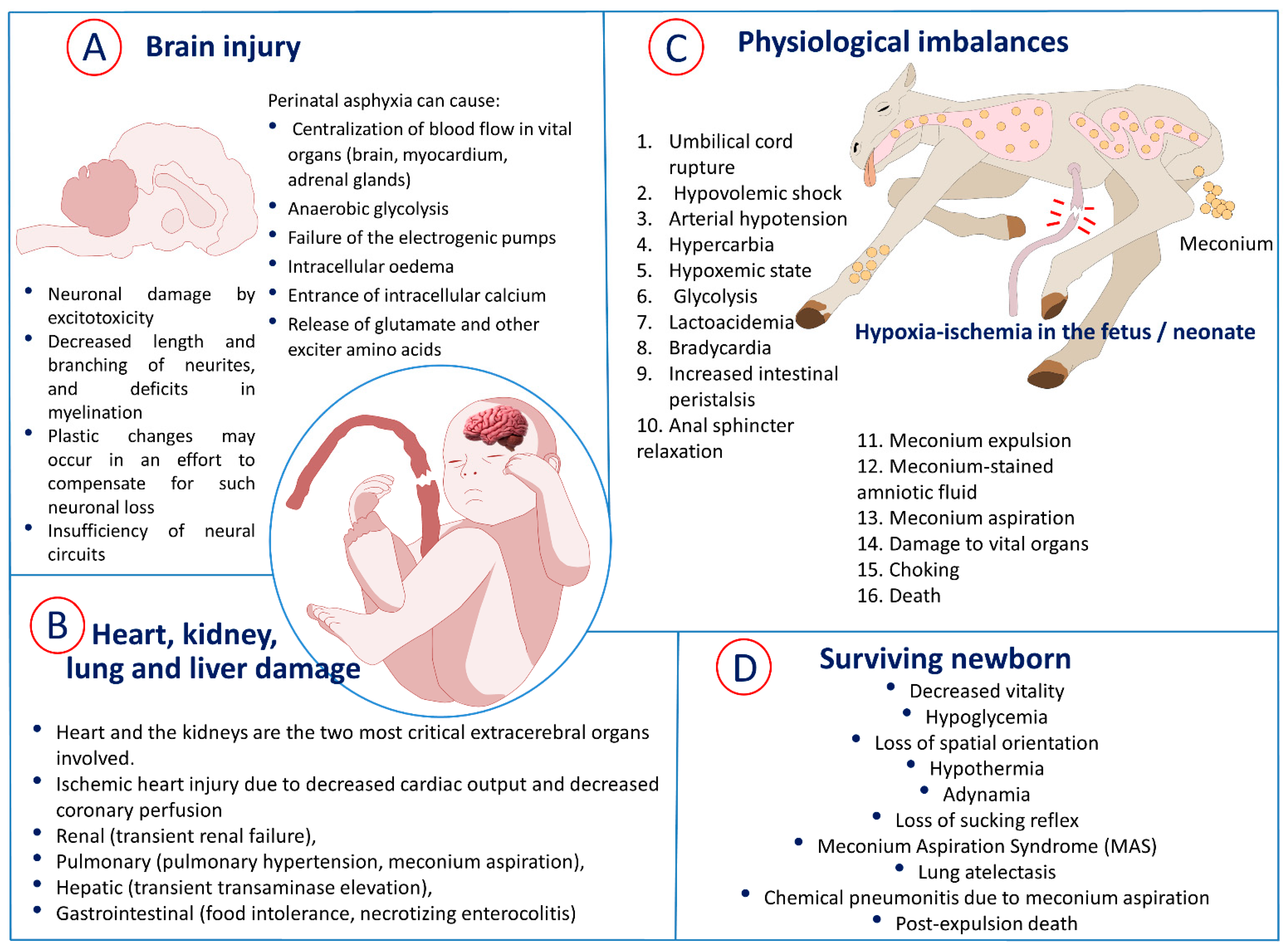
4. Pathophysiology of Birth Asphyxia
4.1. Physiological Changes during Birth Asphyxia
4.2. Mechanisms of Neuronal Injury in Perinatal Asphyxia
5. Cardiovascular Alterations and Multiorgan Dysfunction
5.1. Cardiovascular Response
5.2. Renal, Hepatic, Pulmonary, and Gastrointestinal Injury
6. Meconium Aspiration Syndrome
7. Criteria for Diagnosis of Hypoxia–Ischemia
- a.
- Apgar score of <5 at 5 and 10 min;
- b.
- Need for mechanical ventilation or resuscitation at 10 min;
- c.
- Acidemia documented in fetal umbilical artery (pH < 7.0 or base deficit ≥ 12 mmol/L);
- d.
- Multisystem organ failure;
- e.
- Evidence of moderate or severe encephalopathy staging, often supported by neuroimaging with evidence of acute brain injury consistent with hypoxia–ischemia.
Clinical Assessment
- Reduced responsiveness with hypotonia or incomplete reflexes (including weak suck) or clinical seizures.
- At least three signs from the following categories:
- (a)
- Reduced responsiveness;
- (b)
- Reduced activity;
- (c)
- Abnormal posture;
- (d)
- Abnormal tone;
- (e)
- Incomplete reflexes;
- (f)
- Abnormal pupil response, heart rate, or respiration.
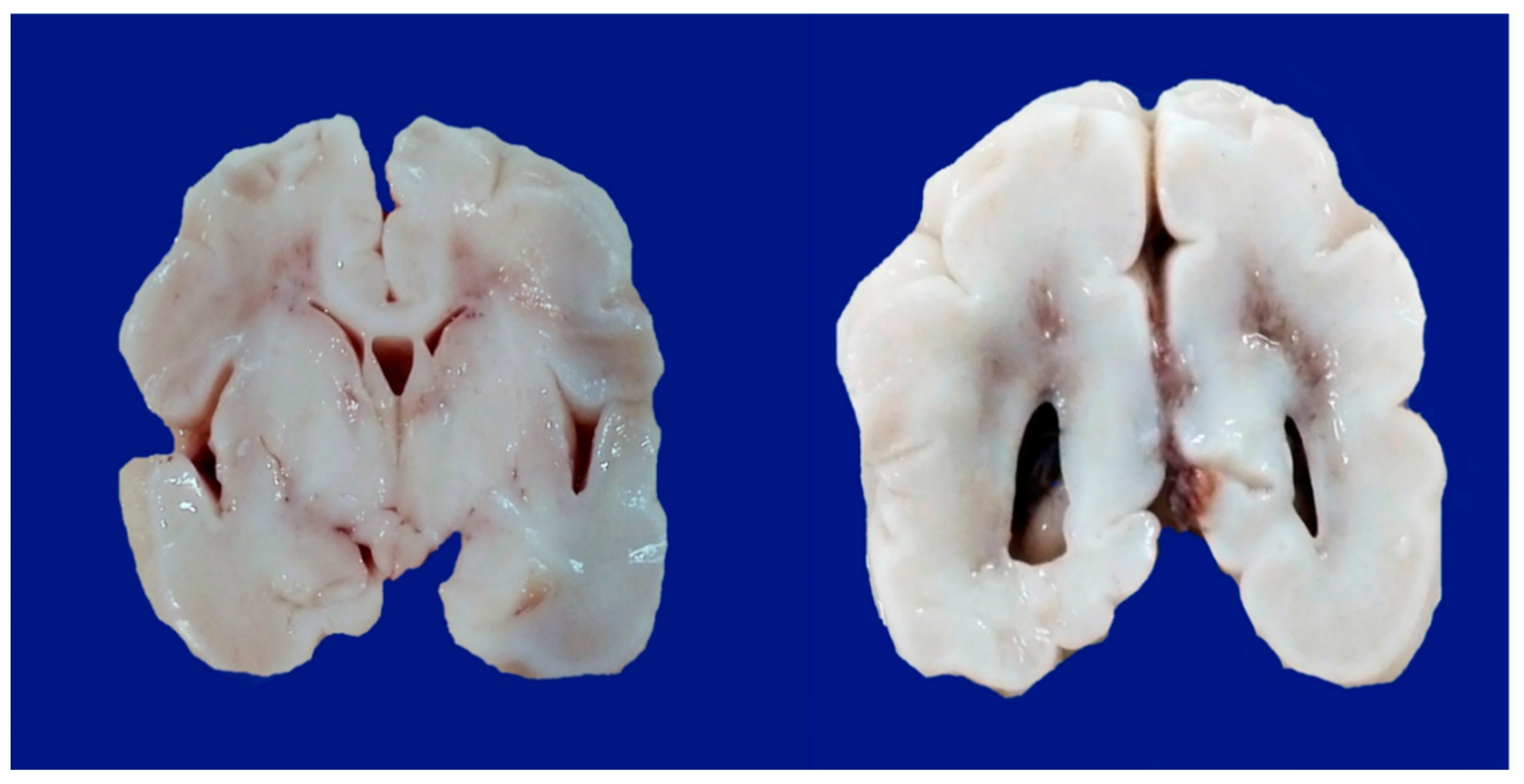
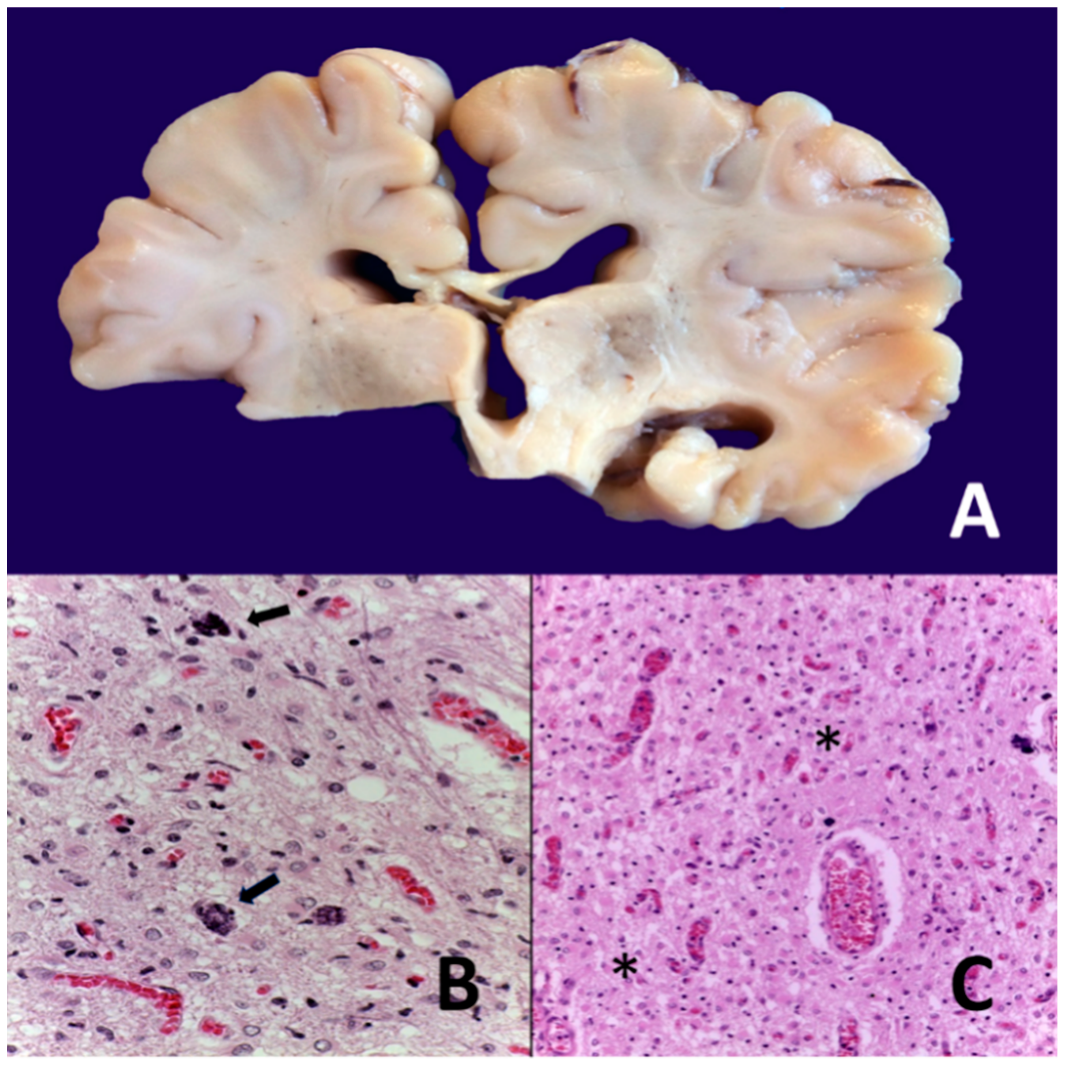
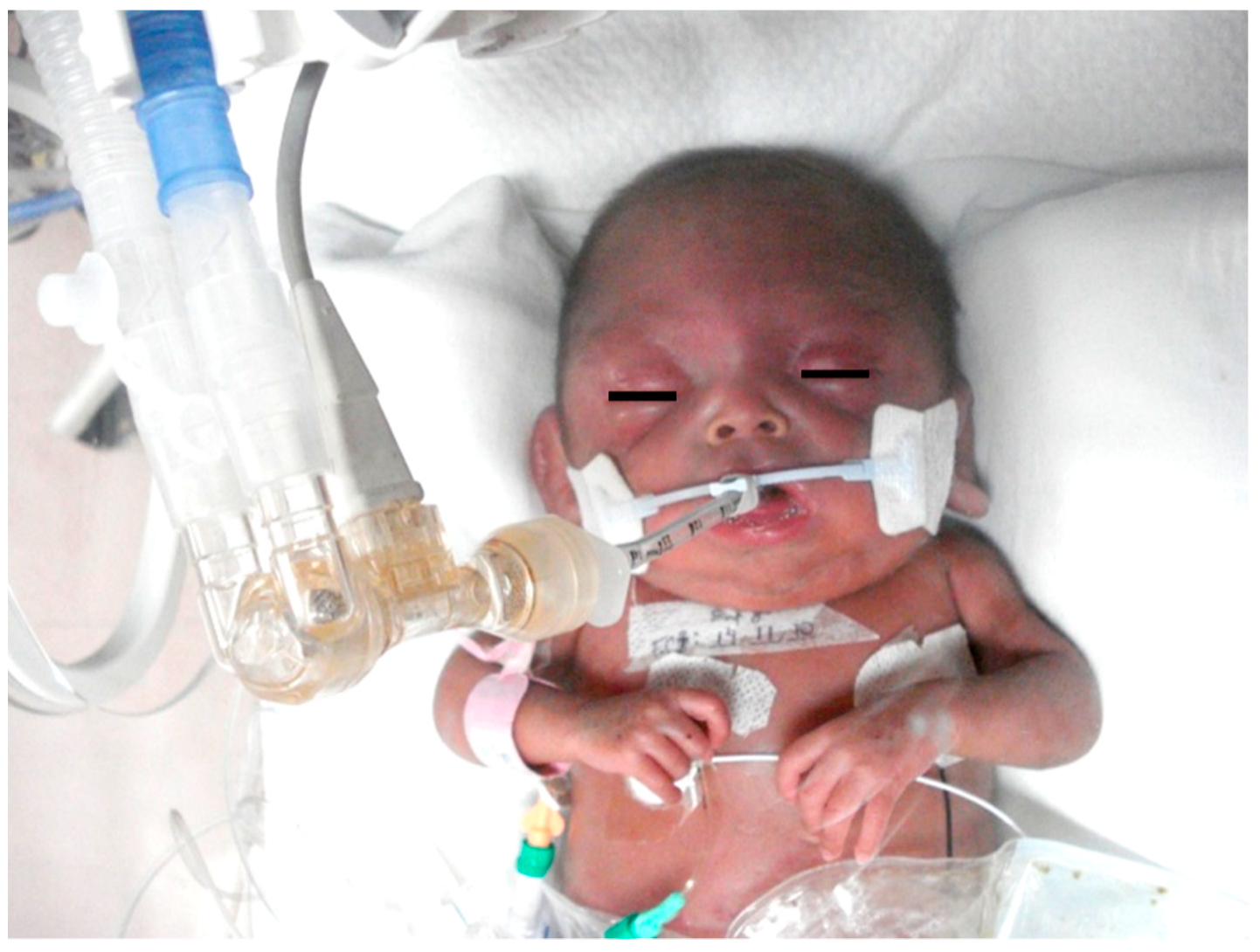
8. Neonatal Hypoxic–Ischemic Encephalopathy: Clinical Aspects
- -
- Parasagittal injury of the cerebral cortex, in subcortical white matter in the lateral convection of the superior-medial orientation, in the posterior–anterior direction.
- -
- Periventricular leukomalacia with necrosis in the subcortical white matter of the hemisphere, including descending motor fibers, optical radiations, and association fibers.
- -
- Focal and multifocal necrotic ischemia in the cerebral cortex and subcortical necrosis in white matter, mainly unilateral with a vascular distribution.
- -
8.1. Inflammatory Biomarkers of Birth Asphyxia
8.2. Placental Inflammatory Biomarkers
8.3. Serum Brain Biomarkers
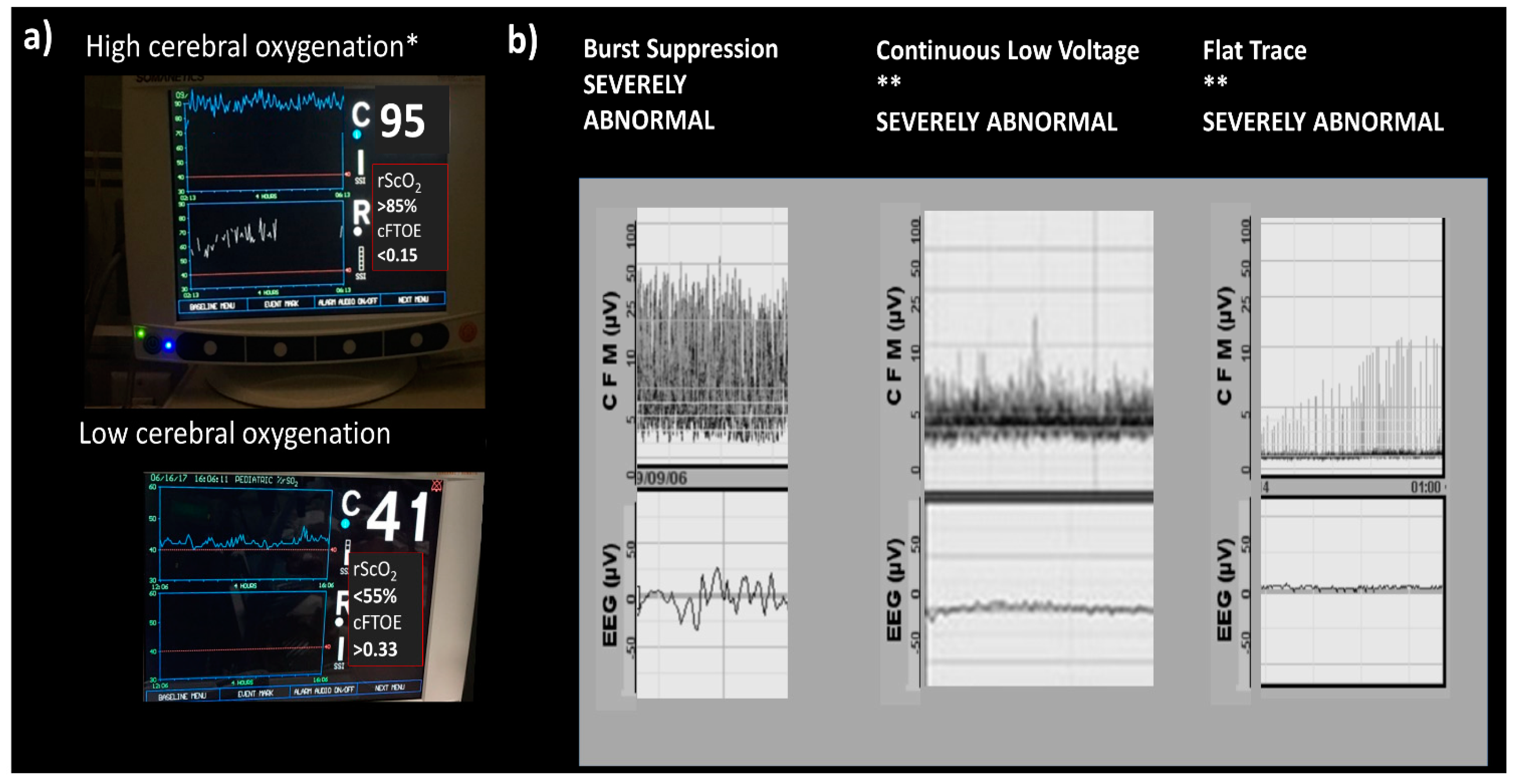
8.4. Electrophysiology
8.5. Near-Infrared Spectroscopy
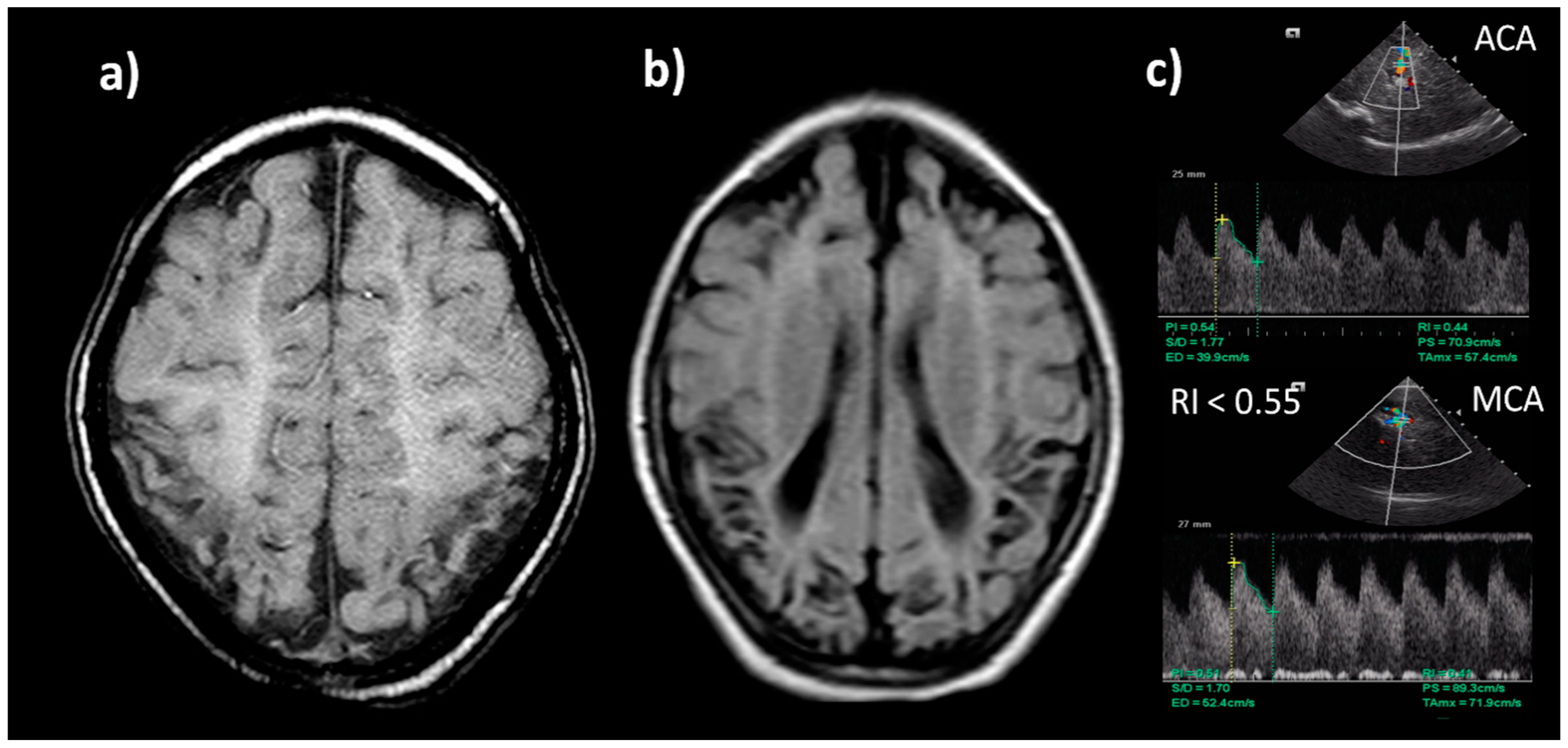
8.6. Neuroimaging
8.7. Hemodynamic Management
- Low systemic blood flow with normal oxygenation: with echocardiographic findings consistent of left ventricle (LV)/right ventricle (RV) dysfunction in which management must include positive inotropes.
- Low systemic blood flow with impaired oxygenation. In this scenario, the echocardiographic finding can show:
- (a)
- Persistent pulmonary hypertension, where management should include pulmonary vasodilation, subsequentially augmenting systemic blood flow after pulmonary venous return improves;
- (b)
- LV dysfunction with PPHN, where management must include positive inotropy and maintenance of right-to-left ductal shunt to support systolic blood flow;
- (c)
- RV dysfunction with PPHN, where management must include positive inotropy, reduced RV afterload (pulmonary vasodilation and consideration of prostaglandin E1 if the ductus arteriosus is restrictive) and maintenance of adequate RV preload.
9. Targets for Neuroprotection
Potential Interventions for Birth Asphyxia: A Window for Reducing Further Brain Injury
10. Neurodevelopment of Babies with Asphyxia
11. Animal Models of Perinatal Asphyxia
12. Scientific Findings of Perinatal Asphyxia in Animal Models: Advantages and Limitations
13. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- WHO. Newborns: Reducing Mortality. Available online: https://www.who.int/news-room/fact-sheets/detail/newborns-reducing-mortality (accessed on 11 November 2021).
- Bhutta, Z.A.; Das, J.K.; Bahl, R.; Lawn, J.E.; Salam, R.A.; Paul, V.K.; Sankar, M.J.; Blencowe, H.; Rizvi, A.; Chou, V.B.; et al. Can available interventions end preventable deaths in mothers, newborn babies, and stillbirths, and at what cost? Lancet 2014, 384, 347–370. [Google Scholar] [CrossRef]
- Lawn, J.E.; Cousens, S.; Zupan, J.; Lancet Neonatal Survival Steering Team. 4 million neonatal deaths: When? Where? Why? Lancet 2005, 365, 891–900. [Google Scholar] [CrossRef]
- Liu, L.; Johnson, H.L.; Cousens, S.; Perin, J.; Scott, S.; Lawn, J.E.; Rudan, I.; Campbell, H.; Cibulskis, R.; Li, M.; et al. Global, regional, and national causes of child mortality: An updated systematic analysis for 2010 with time trends since 2000. Lancet 2012, 379, 2151–2161. [Google Scholar] [CrossRef]
- Perez, A.; Ritter, S.; Brotschi, B.; Werner, H.; Caflisch, J.; Martin, E.; Latal, B. Long-Term Neurodevelopmental Outcome with Hypoxic-Ischemic Encephalopathy. J. Pediatr. 2013, 163, 454–459.e1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Concepcion, K.; Meng, X.; Zhang, L. Brain-immune interactions in perinatal hypoxic-ischemic brain injury. Prog. Neurobiol. 2017, 159, 50–68. [Google Scholar] [CrossRef] [PubMed]
- Younge, N.; Goldstein, R.F.; Bann, C.M.; Hintz, S.R.; Patel, R.M.; Smith, P.B.; Bell, E.F.; Rysavy, M.A.; Duncan, A.F.; Vohr, B.R.; et al. Survival and Neurodevelopmental Outcomes among Periviable Infants. N. Engl. J. Med. 2017, 376, 617–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- National Institute of Health. Late Hypothermia for Hypoxic-Ischemic Encephalopathy. Available online: https://clinicaltrials.gov/ct2/show/NCT00614744 (accessed on 20 July 2021).
- National Institute of Health. Preemie Hypothermia for Neonatal Encephalopathy. Available online: https://clinicaltrials.gov/ct2/show/NCT01793129 (accessed on 11 August 2021).
- National Institute of Health. Erythropoietin for Hypoxic Ischemic Encephalopathy in Newborns (PAEAN). Available online: https://clinicaltrials.gov/ct2/show/NCT03079167 (accessed on 11 November 2021).
- Lincetto, O. Birth Asphyxia Summary of the Previous Meeting and Protocol Overview; World Health Organization: Gevena, Switzerland, 2007; pp. 1–34. [Google Scholar]
- Kamath-Rayne, B.D.; Hobe, A. Birth asphyxia. In Clinics in Perinatology; Elsevier: Philadelphia, PA, USA, 2016; p. 621. [Google Scholar]
- American Academy of Pediatrics; Committee on Fetus and Newborn; American College of Obstetricians and Gynecologists; Committee on Obstetric Practice. The Apgar score. Adv. Neonatal Care Off. J. Natl. Assoc. Neonatal Nurses 2006, 6, 220–223. [Google Scholar] [CrossRef] [PubMed]
- Rainaldi, M.A.; Perlman, J.M. Pathophysiology of Birth Asphyxia. Clin. Perinatol. 2016, 43, 409–422. [Google Scholar] [CrossRef]
- Pacora, P.; Romero, R.; Jaiman, S.; Erez, O.; Bhatti, G.; Panaitescu, B.; Benshalom-Tirosh, N.; Jung, E.J.; Hsu, C.-D.; Hassan, S.S.; et al. Mechanisms of death in structurally normal stillbirths. J. Perinat. Med. 2019, 47, 222–240. [Google Scholar] [CrossRef]
- Yli, B.M.; Kjellmer, I. Pathophysiology of fetal oxygenation and cell damage during labor. Best Pract. Res. Clin. Obstet. Gynaecol. 2016, 30, 9–21. [Google Scholar] [CrossRef]
- Richardson, B.S. Fetal Adaptive Responses to Asphyxia. Clin. Perinatol. 1989, 16, 595–611. [Google Scholar] [CrossRef]
- Jensen, A.; Roman, C.; Rudolph, A.M. Effects of reducing uterine blood flow on fetal blood flow distribution and oxygen delivery. J. Dev. Physiol. 1991, 15, 309–323. [Google Scholar] [PubMed]
- Low, J.A. Determining the contribution of asphyxia to brain damage in the neonate. J. Obstet. Gynaecol. Res. 2004, 30, 276–286. [Google Scholar] [CrossRef] [PubMed]
- Britton, J.R. The transition to extrauterine life and disorders of transition. Clin. Perinatol. 1998, 25, 271–294. [Google Scholar] [CrossRef]
- Morton, S.U.; Brodsky, D. Fetal Physiology and the Transition to Extrauterine Life. Clin. Perinatol. 2016, 43, 395–407. [Google Scholar] [CrossRef] [Green Version]
- Noori, S.; Wlodaver, A.; Gottipati, V.; McCoy, M.; Schultz, D.; Escobedo, M. Transitional Changes in Cardiac and Cerebral Hemodynamics in Term Neonates at Birth. J. Pediatrics 2012, 160, 943–948. [Google Scholar] [CrossRef]
- Wyss, M.T.; Jolivet, R.; Buck, A.; Magistretti, P.J.; Weber, B. In Vivo Evidence for Lactate as a Neuronal Energy Source. J. Neurosci. 2011, 31, 7477–7485. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Wang, X.-M. Expression Changes in Lactate and Glucose Metabolism and Associated Transporters in Basal Ganglia following Hypoxic-Ischemic Reperfusion Injury in Piglets. Am. J. Neuroradiol. 2018, 39, 569–576. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Engidawork, E.; Loidl, F.; Dell’Anna, E.; Goiny, M.; Lubec, G.; Andersson, K.; Herrera-Marschitz, M. Short- and long-term effects of perinatal asphyxia on monoamine, amino acid and glycolysis product levels measured in the basal ganglia of the rat. Brain Res. Dev. Brain Res. 1997, 104, 19–30. [Google Scholar] [CrossRef]
- Herrera, M.I.; Otero-Losada, M.; Udovin, L.D.; Kusnier, C.; Kölliker-Frers, R.; De Souza, W.; Capani, F. Could perinatal asphyxia induce a synaptopathy? New highlights from an experimental model. Neural Plast. 2017, 2017, 3436943. [Google Scholar] [CrossRef]
- Larroque, B.; Ancel, P.-Y.; Marret, S.; Marchand, L.; André, M.; Arnaud, C.; Pierrat, V.; Rozé, J.-C.; Messer, J.; Thiriez, G.; et al. Neurodevelopmental disabilities and special care of 5-year-old children born before 33 weeks of gestation (the EPIPAGE study): A longitudinal cohort study. Lancet 2008, 371, 813–820. [Google Scholar] [CrossRef]
- Laptook, A.R. Birth Asphyxia and Hypoxic-Ischemic Brain Injury in the Preterm Infant. Clin. Perinatol. 2016, 43, 529–545. [Google Scholar] [CrossRef] [PubMed]
- Antonucci, R.; Porcella, A.P.M. Perinatal asphyxia in the term newborn. J. Pediatr. Neonatal Individ. Med. 2014, 3, e030269. [Google Scholar]
- Summanen, M.; Bäck, S.; Voipio, J.; Kaila, K. Surge of Peripheral Arginine Vasopressin in a Rat Model of Birth Asphyxia. Front. Cell. Neurosci. 2018, 12, 2. [Google Scholar] [CrossRef] [PubMed]
- VanWoudenberg, C.D.; Wills, C.A.; Rubarth, L.B. Newborn Transition to Extrauterine Life. Neonatal Netw. 2012, 31, 317–322. [Google Scholar] [CrossRef] [PubMed]
- Evers, K.S.; Wellmann, S. Arginine vasopressin and copeptin in perinatology. Front. Pediatr. 2016, 4, 2–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dilenge, M.E.; Majnemer, A.; Shevell, M.I. Long-term developmental outcome of asphyxiated term neonates. J. Child Neurol. 2001, 16, 781–792. [Google Scholar] [CrossRef]
- Milsom, I.; Ladfors, L.; Thiringer, K.; Niklasson, A.; Odeback, A.; Thornberg, E. Influence of maternal, obstetric and fetal risk factors on the prevalence of birth asphyxia at term in a Swedish urban population. Acta Obstet. Gynecol. Scand. 2002, 81, 909–917. [Google Scholar] [CrossRef]
- Giussani, D.A. The fetal brain sparing response to hypoxia: Physiological mechanisms. J. Physiol. 2016, 594, 1215–1230. [Google Scholar] [CrossRef] [Green Version]
- Polglase, G.R.; Ong, T.; Hillman, N.H. Cardiovascular Alterations and Multiorgan Dysfunction After Birth Asphyxia. Clin. Perinatol. 2016, 43, 469–483. [Google Scholar] [CrossRef] [Green Version]
- Fattuoni, C.; Palmas, F.; Noto, A.; Fanos, V.; Barberini, L. Perinatal asphyxia: A review from a metabolomics perspective. Molecules 2015, 20, 7000–7016. [Google Scholar] [CrossRef] [PubMed]
- Denihan, N.M.; Boylan, G.B.; Murray, D.M. Metabolomic profiling in perinatal asphyxia: A promising new field. BioMed Res. Int. 2015, 2015, 254076. [Google Scholar] [CrossRef] [PubMed]
- Mota-Rojas, D.; Martínez-Burnes, J.; Trujillo, M.E.; López, A.; Rosales, A.M.; Ramírez, R.; Orozco, H.; Merino, A.; Alonso-Spilsbury, M. Uterine and fetal asphyxia monitoring in parturient sows treated with oxytocin. Anim. Reprod. Sci. 2005, 86, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Mota-Rojas, D.; Rosales, A.M.; Trujillo, M.E.; Orozco, H.; Ramírez, R.; Alonso-Spilsbury, M. The effects of vetrabutin chlorhydrate and oxytocin on stillbirth rate and asphyxia in swine. Theriogenology 2005, 64, 1889–1897. [Google Scholar] [CrossRef] [PubMed]
- Mota-Rojas, D.; Nava-Ocampo, A.A.; Trujillo, M.E.; Velázquez-Armenta, Y.; Ramírez-Necoechea, R.; Martínez-Burnes, J.; Alonso-Spilsbury, M. Dose minimization study of oxytocin in early labor in sows: Uterine activity and fetal outcome. Reprod. Toxicol. 2005, 20, 255–259. [Google Scholar] [CrossRef] [PubMed]
- Mota-Rojas, D.; Martínez-Burnes, J.; Napolitano, F.; Domínguez-Muñoz, M.; Guerrero-Legarreta, I.; Mora-Medina, P.; Ramírez-Necoechea, R.; Lezama-García, K.; González-Lozano, M.; Mota-Rojas, D.; et al. Dystocia: Factors affecting parturition in domestic animals. CAB Rev. 2020, 15, 1–16. [Google Scholar] [CrossRef]
- Alonso-Spilsbury, M.; Mota-Rojas, D.; Villanueva-García, D.; Martínez-Burnes, J.; Orozco, H.; Ramírez-Necoechea, R.; Mayagoitia, A.L.; Trujillo, M.E. Perinatal asphyxia pathophysiology in pig and human: A review. Anim. Reprod. Sci. 2005, 90, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Mota-Rojas, D.; Martinez-Burnes, J.; Alonso-Spilsbury, M.L.; Lopez, A.; Ramirez-Necoechea, R.; Trujillo-Ortega, M.E.; Medina-Hernandez, F.J.; de la Cruz, N.I.; Albores-Torres, V.; Loredo-Osti, J. Meconium staining of the skin and meconium aspiration in porcine intrapartum stillbirths. Livest. Sci. 2006, 102, 155–162. [Google Scholar] [CrossRef]
- Mota, D.; Orozco, H.; Villanueva, D.; Bonilla, H.; Suárez, X.; Hernandez, R.; Roldan, P.; Trujillo, M. Fetal and neonatal energy metabolism in pigs and humans: A review. Vet. Med. 2011, 56, 215–225. [Google Scholar] [CrossRef] [Green Version]
- Mota-Rojas, D.; Velarde, A.; Maris-Huertas, S.; Cajiao, M.N. Animal Welfare, a Global Vision in Ibero-America, 3rd ed.; Mota-Rojas, D., Velarde, A., Maris-Huertas, S., Cajiao, M.N., Eds.; Elsevier: Barcelona, Spain, 2016; ISBN 978-84-9113-026-0. [Google Scholar]
- Mota-Rojas, D.; López, A.; Martínez-Burnes, J.; Muns, R.; Villanueva-García, D.; Mora-Medina, P.; González-Lozano, M.; Olmos-Hernández, A.; Ramírez-Necoechea, R. Is vitality assessment important in neonatal animals? CAB Rev. 2018, 13, 1–13. [Google Scholar] [CrossRef]
- Martínez-Burnes, J.; Mota-Rojas, D.; Villanueva-García, D.; Ibarra-Rios, D.; Lezama-García, K.; Barrios-García, H.; Lopez, A. Meconium aspiration syndrome in mammals. CAB Rev. 2019, 14, 1–11. [Google Scholar] [CrossRef]
- González-Lozano, M.; Mota-Rojas, D.; Orihuela, A.; Martínez-Burnes, J.; Di Francia, A.; Braghieri, A.; Berdugo-Gutiérrez, J.; Mora-Medina, P.; Ramírez-Necoechea, R.; Napolitano, F. Review: Behavioral, physiological, and reproductive performance of buffalo cows during eutocic and dystocic parturitions. Appl. Anim. Sci. 2020, 36, 407–422. [Google Scholar] [CrossRef]
- Placha, K.; Luptakova, D.; Baciak, L.; Ujhazy, E.; Juranek, I. Neonatal brain injury as a consequence of insufficient cerebral oxygenation. Neuro Endocrinol. 2016, 37, 79–96. [Google Scholar]
- Mota-Rojas, D.; Martinez-Burnes, J.; Villanueva-Garcia, D.; Roldan-Santiago, P.; Trujillo-Ortega, M.E.; Orozco-Gregorio, H.; Bonilla-Jaime, H.; Lopez-Mayagoitia, A. Animal welfare in the newborn piglet: A review. Vet. Med. 2012, 57, 338–349. [Google Scholar] [CrossRef] [Green Version]
- Riljak, V.; Kraf, J.; Daryanani, A.; Jiruška, P.; Otáhal, J. Pathophysiology of perinatal hypoxic-ischemic encephalopathy—Biomarkers, animal models and treatment perspectives. Physiol. Res. 2016, 65, S533–S545. [Google Scholar] [CrossRef]
- Engelhardt, B.; Carare, R.O.; Bechmann, I.; Flügel, A.; Laman, J.D.; Weller, R.O. Vascular, glial, and lymphatic immune gateways of the central nervous system. Acta Neuropathol. 2016, 132, 317–338. [Google Scholar] [CrossRef] [Green Version]
- Van Bel, F.V.; Groenendaal, F. Drugs for neuroprotection after birth asphyxia: Pharmacologic adjuncts to hypothermia. Semin. Perinatol. 2016, 40, 152–159. [Google Scholar]
- Hassell, K.J.; Ezzati, M.; Alonso-Alconada, D.; Hausenloy, D.J.; Robertson, N.J. New horizons for newborn brain protection: Enhancing endogenous neuroprotection. Arch. Dis. Child. Fetal Neonatal Ed. 2015, 100, F541–F551. [Google Scholar] [CrossRef]
- Bhatti, A.; Kumar, P. Systemic effects of perinatal asphyxia. Indian J. Pediatrics 2014, 81, 231–233. [Google Scholar] [CrossRef]
- Jensen, A.; Garnier, Y.; Berger, R. Dynamics of fetal circulatory responses to hypoxia and asphyxia. Eur. J. Obstet. Gynecol. Reprod. Biol. 1999, 84, 155–172. [Google Scholar] [CrossRef]
- Pertierra Cortada, À.; Figueras Aloy, J.; Sebastiani, G.; Rovira Girabal, N.; Krauel Vidal, X. Asfixia perinatal: Relación entre afectación cardiovascular, neurológica y multisistémica. Acta Pediatr. Esp. 2008, 66, 494–501. [Google Scholar]
- Hankins, G.D.V.; Koen, S.; Gei, A.F.; Lopez, S.M.; Van Hook, J.W.; Anderson, G.D. Neonatal organ system injury in acute birth asphyxia sufficient to result in neonatal encephalopathy. Obstet. Gynecol. 2002, 99, 688–691. [Google Scholar] [PubMed]
- Kara, T.; Narkiewicz, K.; Somers, V.K. Chemoreflexes—Physiology and clinical implications. Acta Physiol. Scand. 2003, 177, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Giussani, D.A.; Spencer, J.A.; Moore, P.J.; Bennet, L.; Hanson, M.A. Afferent and efferent components of the cardiovascular reflex responses to acute hypoxia in term fetal sheep. J. Physiol. 1993, 461, 431–449. [Google Scholar] [CrossRef]
- Giussani, D.A.; Spencer, J.A.; Hanson, M.A. Fetal cardiovascular reflex responses to hypoxaemia. Fetal Matern. Med. Rev. 1994, 6, 17–37. [Google Scholar] [CrossRef]
- Cullen, P.; Salgado, E. Conceptos básicos para el manejo de la asfixia perinatal y la encefalopatía hipóxico-isquémica en el neonato. Rev. Mex. Pediatr. 2009, 76, 174–180. [Google Scholar]
- Jacobs, S.E.; Berg, M.; Hunt, R.; Tarnow-Mordi, W.O.; Inder, T.E.; Davis, P.G. Cooling for newborns with hypoxic ischemic encephalopathy. Cochrane Database Syst. Rev. 2013, 2013, CD003311. [Google Scholar]
- Sehgal, A.; Wong, F.; Mehta, S. Reduced cardiac output and its correlation with coronary blood flow and troponin in asphyxiated infants treated with therapeutic hypothermia. Eur. J. Pediatr. 2012, 171, 1511–1517. [Google Scholar] [CrossRef]
- LaRosa, D.A.; Ellery, S.J.; Walker, D.W.; Dickinson, H. Understanding the Full Spectrum of organ injury following intrapartum asphyxia. Front. Pediatr. 2017, 5, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dattilo, G.; Tulino, V.; Tulino, D.; Lamari, A.; Falanga, G.; Marte, F.; Patanè, S. Perinatal asphyxia and cardiac abnormalities. Int. J. Cardiol. 2011, 147, e39–e40. [Google Scholar] [CrossRef]
- Perlman, J.M. Acute Systemic Organ Injury in Term Infants After Asphyxia. Arch. Pediatr. Adolesc. Med. 1989, 143, 617. [Google Scholar] [CrossRef] [PubMed]
- Gunn, A.J.; Bennet, L. Fetal Hypoxia Insults and Patterns of Brain Injury: Insights from Animal Models. Clin. Perinatol. 2009, 36, 579–593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikeda, T.; Murata, Y.; Quilligan, E.J.; Parer, J.T.; Murayama, T.; Koono, M. Histologic and biochemical study of the brain, heart, kidney, and liver in asphyxia caused by occlusion of the umbilical cord in near-term fetal lambs. Am. J. Obstet. Gynecol. 2000, 182, 449–457. [Google Scholar] [CrossRef]
- Ikeda, T.; Choi, B.H.; Yee, S.; Murata, Y.; Quilligan, E.J. Oxidative stress, brain white matter damage and intrauterine asphyxia in fetal lambs. Int. J. Dev. Neurosci. 1999, 17, 1–14. [Google Scholar] [CrossRef]
- Shankaran, S.; Woldt, E.; Koepke, T.; Bedard, M.P.; Nandyal, R. Acute neonatal morbidity and long-term central nervous system sequelae of perinatal asphyxia in term infants. Early Hum. Dev. 1991, 25, 135–148. [Google Scholar] [CrossRef]
- Aggarwal, A.; Kumar, P.; Chowdhary, G.; Majumdar, S.; Narang, A. Evaluation of Renal Functions in Asphyxiated Newborns. J. Trop. Pediatr. 2005, 51, 295–299. [Google Scholar] [CrossRef] [Green Version]
- Saikumar, P.; Venkatachalam, M.A. Role of apoptosis in hypoxic/ischemic damage in the kidney. Semin. Nephrol. 2003, 23, 511–521. [Google Scholar] [CrossRef]
- Askenazi, D.J.; Feig, D.I.; Graham, N.M.; Hui-Stickle, S.; Goldstein, S.L. 3–5 year longitudinal follow-up of pediatric patients after acute renal failure. Kidney Int. 2006, 69, 184–189. [Google Scholar] [CrossRef] [Green Version]
- Jetton, J.G.; Askenazi, D.J. Acute Kidney Injury in the Neonate. Clin. Perinatol. 2014, 41, 487–502. [Google Scholar] [CrossRef]
- Beath, S.V. Hepatic function and physiology in the newborn. Semin. Neonatol. 2003, 8, 337–346. [Google Scholar] [CrossRef]
- Islam, M.T.; Islam, M.N.; Mollah, A.H.; Hoque, M.A.; Hossain, M.A.; Nazir, F.; Ahsan, M.M. Status of liver enzymes in babies with perinatal asphyxia. Mymensingh Med. J. MMJ 2011, 20, 446–449. [Google Scholar] [PubMed]
- Gluckman, P.D.; Wyatt, J.S.; Azzopardi, D.; Ballard, R.; Edwards, A.D.; Ferriero, D.M.; Polin, R.A.; Robertson, C.M.; Thoresen, M.; Whitelaw, A.; et al. Selective head cooling with mild systemic hypothermia after neonatal encephalopathy: Multicentre randomised trial. Lancet 2005, 365, 663–670. [Google Scholar] [CrossRef]
- Forman, K.R.; Diab, Y.; Wong, E.C.C.; Baumgart, S.; Luban, N.L.C.; Massaro, A.N. Coagulopathy in newborns with hypoxic ischemic encephalopathy (HIE) treated with therapeutic hypothermia: A retrospective case-control study. BMC Pediatr. 2014, 14, 277. [Google Scholar] [CrossRef] [Green Version]
- Paprocka, J.; Kijonka, M.; Rzepka, B.; Sokół, M. Melatonin in Hypoxic-Ischemic Brain Injury in Term and Preterm Babies. Int. J. Endocrinol. 2019, 2019, 9626715. [Google Scholar] [CrossRef] [PubMed]
- American College of Obstetricians and Gynecologists Committee on Obstetric Practice. Inappropriate use of the terms fetal distress and birth asphyxia. Obstet. Gynecol. 2005, 106, 1469–1470. [Google Scholar] [CrossRef] [PubMed]
- Committee on Obstetric Practice, ACOG; American Academy of Pediatrics; Committee on Fetus and Newborn, ACOG. ACOG Committee Opinion No. 333: The Apgar Score. Obstet. Gynecol. 2006, 107, 1209. [Google Scholar] [CrossRef] [PubMed]
- Rahman, S.; Unsworth, J.; Vause, S. Meconium in labor. Obstet. Gynaecol. Reprod. Med. 2013, 23, 247–252. [Google Scholar] [CrossRef]
- Vain, N.E.; Batton, D.G. Meconium “aspiration” (or respiratory distress associated with meconium-stained amniotic fluid?). Semin. Fetal Neonatal Med. 2017, 22, 214–219. [Google Scholar] [CrossRef]
- Chettri, S.; Bhat, B.V.; Adhisivam, B. Current Concepts in the Management of Meconium Aspiration Syndrome. Indian J. Pediatr. 2016, 83, 1125–1130. [Google Scholar] [CrossRef]
- Yurdakök, M. Meconium aspiration syndrome: Do we know? Turk. J. Pediatr. 2011, 53, 121–129. [Google Scholar]
- Lee, J.; Romero, R.; Lee, K.A.; Kim, E.N.; Korzeniewski, S.J.; Chaemsaithong, P.; Yoon, B.H. Meconium aspiration syndrome: A role for fetal systemic inflammation. Am. J. Obstet. Gynecol. 2016, 214, 366.e1–366.e9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendoza, M.M.; Martinez-Burnes, J.; Calderon, R.; Hayen, S.; Mota-Rojas, D.; Medellin, J.A. Meconium staining of foals at birth. In Animal Perinatology: Clinical and Experimental Approaches; Mota-Rojas, D., Nava-Ocampo, A.A., Villanueva-Garcia, D.A.-S.M., Eds.; BM Editores: Ciudad de México, Mexico, 2008; pp. 351–362. [Google Scholar]
- Villanueva-García, D. Hipertensión arterial pulmonar. In Neonatología, Esencia, Arte y Praxis; Murguía, T., Villanueva, D.L.G., Eds.; Mc Graw Hill: Ciudad de México, Mexico, 2011; pp. 88–94. [Google Scholar]
- Ibarra-Ríos, D.; Villanueva-García, D. Síndrome de Aspiración de Meconio. Programa de Actualización Continua. In PAC® Neonatología-4; Libro 2; Insuficiencia Respiratoria Neonatal, Ed.; Intersistemas Editores: Ciudad de México, Mexico, 2016; pp. 59–67. ISBN 978-607-443-552-8. [Google Scholar]
- Mota-Rojas, D.; Villanueva-García, D.; Hernández, R.; Martínez-Rodríguez, R.; Mora-Medina, P.; Gonzalez, B.; Sánchez, M. Assessment of the vitality of the newborn: An overview. Sci. Res. Essays 2012, 7, 712–718. [Google Scholar] [CrossRef]
- Dawes, G.S. The natural history of asphyxia and resuscitation at birth. In Fetal and Neonatal Physiology: A Comparative Study of the Changes at Birth; Yearbook Medical Publishers: Chicago, IL, USA, 1968. [Google Scholar]
- Dawes, G.S.; Jacobson, H.N.; Mott, J.C.; Shelley, H.J.; Stafford, A. The treatment of asphyxiated, mature fetal lambs and rhesus monkeys with intravenous glucose and sodium carbonate. J. Physiol. 1963, 169, 167–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobs, S.E.; Morley, C.J.; Inder, T.E.; Stewart, M.J.; Smith, K.R.; McNamara, P.J.; Wright, I.M.R.; Kirpalani, H.M.; Darlow, B.A.; Doyle, L.W. Whole-body hypothermia for term and near-term newborns with hypoxic-ischemic encephalopathy: A randomized controlled trial. Arch. Pediatr. Adolesc. Med. 2011, 165, 692–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarnat, H.B. Neonatal Encephalopathy Following Fetal Distress. Arch. Neurol. 1976, 33, 696–705. [Google Scholar] [CrossRef] [PubMed]
- Thompson, C.; Puterman, A.; Linley, L.; Hann, F.; Elst, C.; Molteno, C.; Malan, A. The value of a scoring system for hypoxic ischemic encephalopathy in predicting neurodevelopmental outcome. Acta Paediatr. 1997, 86, 757–761. [Google Scholar] [CrossRef] [PubMed]
- Shankaran, S.; Laptook, A.R.; Ehrenkranz, R.A.; Tyson, J.E.; McDonald, S.A.; Donovan, E.F.; Fanaroff, A.A.; Poole, W.K.; Wright, L.L.; Higgins, R.D.; et al. Whole-Body Hypothermia for Neonates with Hypoxic-Ischemic Encephalopathy. N. Engl. J. Med. 2005, 353, 1574–1584. [Google Scholar] [CrossRef]
- Ducsay, C.A.; Goyal, R.; Pearce, W.J.; Wilson, S.; Hu, X.Q.; Zhang, L. Gestational hypoxia and developmental plasticity. Physiol. Rev. 2018, 98, 1241–1334. [Google Scholar] [CrossRef]
- Dos Riesgo, R.S.; Becker, M.M.; Ranzan, J.; Winckler, M.I.B.; Ohlweiler, L. Avances en el abordaje de la hipoxia neonatal. Rev. Neurol. 2013, 57, S17–S21. [Google Scholar] [CrossRef]
- Negro, S.; Benders, M.J.N.L.; Tataranno, M.L.; Coviello, C.; de Vries, L.S.; van Bel, F.; Groenendaal, F.; Longini, M.; Proietti, F.; Belvisi, E.; et al. Early prediction of hypoxic-ischemic brain injury by a new panel of biomarkers in a population of term newborns. Oxidative Med. Cell. Longev. 2018, 2018, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Volpe, J.J.; Inder, T.; Darras, B.; Vries, L.; Plessis, A.; Neil, J.; Perlman, J. Volpe´s Neurology of the Newborn. Philadelphia; Saunders: Philadelphia, PA, USA, 2001; pp. 1–1224. [Google Scholar]
- Chaiworapongsa, T.; Romero, R.; Erez, O.; Tarca, A.L.; Conde-Agudelo, A.; Chaemsaithong, P.; Kim, C.J.; Kim, Y.M.; Kim, J.-S.; Yoon, B.H.; et al. The prediction of fetal death with a simple maternal blood test at 24–28 weeks: A role for angiogenic index-1 (PlGF/sVEGFR-1 ratio). Am. J. Obstet. Gynecol. 2017, 217, 682.e1–682.e13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thoresen, M.; Liu, X.; Jary, S.; Brown, E.; Sabir, H.; Stone, J.; Cowan, F.; Karlsson, M. Lactate dehydrogenase in hypothermia-treated newborn infants with hypoxic-ischemic encephalopathy. Acta Paediatr. 2012, 101, 1038–1044. [Google Scholar] [CrossRef] [PubMed]
- Ramaswamy, V.; Horton, J.; Vandermeer, B.; Buscemi, N.; Miller, S.; Yager, J. Systematic review of biomarkers of brain injury in term neonatal encephalopathy. Pediatr. Neurol. 2009, 40, 215–226. [Google Scholar] [CrossRef] [PubMed]
- Chalak, L.F. Inflammatory Biomarkers of Birth Asphyxia. Clin. Perinatol. 2016, 43, 501–510. [Google Scholar] [CrossRef]
- Mir, I.N.; Johnson-Welch, S.F.; Nelson, D.B.; Brown, L.S.; Rosenfeld, C.R.; Chalak, L.F. Placental pathology is associated with severity of neonatal encephalopathy and adverse developmental outcomes following hypothermia. Am. J. Obstet. Gynecol. 2015, 213, 849.e1–849.e7. [Google Scholar] [CrossRef]
- Chalak, L.F.; Sánchez, P.J.; Adams-Huet, B.; Laptook, A.R.; Heyne, R.J.; Rosenfeld, C.R. Biomarkers for severity of neonatal hypoxic-ischemic encephalopathy and outcomes in newborns receiving hypothermia therapy. J. Pediatr. 2014, 164, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Massaro, A.N.; Chang, T.; Kadom, N.; Tsuchida, T.; Scafidi, J.; Glass, P.; McCarter, R.; Baumgart, S.; Vezina, G.; Nelson, K.B. Biomarkers of brain injury in neonatal encephalopathy treated with hypothermia. J. Pediatr. 2012, 161, 434–440. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.-W.; Li, S.; Dai, S.-S. Neutrophils in traumatic brain injury (TBI): Friend or foe? J. Neuroinflamm. 2018, 15, 146. [Google Scholar] [CrossRef] [Green Version]
- Sorokina, E.G.; Semenova, Z.B.; Reutov, V.P.; Arsenieva, E.N.; Karaseva, O.V.; Fisenko, A.P.; Roshal, L.M.; Pinelis, V.G. Brain Biomarkers in children after mild and severe traumatic brain injury. In Intracranial Pressure and Neuromonitoring XVII; Depreitere, B., Meyfroidt, G., Güiza, F., Eds.; Springer Nature Switzerland AG: Cham, Switzerland, 2021; pp. 103–107. [Google Scholar]
- Li, Y.; Dammer, E.B.; Zhang-Brotzge, X.; Chen, S.; Duong, D.M.; Seyfried, N.T.; Kuan, C.-Y.; Sun, Y.-Y. Osteopontin is a blood biomarker for microglial activation and brain injury in experimental hypoxic-ischemic encephalopathy. Eneuro 2017, 4. [Google Scholar] [CrossRef] [Green Version]
- Graham, E.M.; Everett, A.D.; Delpech, J.C.; Northington, F.J. Blood biomarkers for evaluation of perinatal encephalopathy: State of the art. Curr. Opin. Pediatr. 2018, 30, 199–203. [Google Scholar] [CrossRef] [Green Version]
- Spitzmiller, R.E.; Phillips, T.; Meinzen-Derr, J.; Hoath, S.B. Amplitude-integrated EEG is useful in predicting neurodevelopmental outcome in full-term infants with hypoxic-ischemic encephalopathy: A meta-analysis. J. Child Neurol. 2007, 22, 1069–1078. [Google Scholar] [CrossRef] [PubMed]
- Lemmers, P.M.A.; Zwanenburg, R.J.; Benders, M.J.N.L.; de Vries, L.S.; Groenendaal, F.; van Bel, F.; Toet, M.C. Cerebral oxygenation and brain activity after perinatal asphyxia: Does hypothermia change their prognostic value? Pediatr. Res. 2013, 74, 180–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Archer, L.N.; Levene, M.; Evans, D. Cerebral artery doppler ultrasonography for prediction of outcome after perinatal asphyxia. Lancet 1986, 328, 1116–1118. [Google Scholar] [CrossRef]
- Bednarek, N.; Mathur, A.; Inder, T.; Wilkinson, J.; Neil, J.; Shimony, J. Impact of therapeutic hypothermia on MRI diffusion changes in neonatal encephalopathy. Neurology 2012, 78, 1420–1427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trivedi, S.B.; Vesoulis, Z.A.; Rao, R.; Liao, S.M.; Shimony, J.S.; McKinstry, R.C.; Mathur, A.M. A validated clinical MRI injury scoring system in neonatal hypoxic-ischemic encephalopathy. Pediatr. Radiol. 2017, 47, 1491–1499. [Google Scholar] [CrossRef] [PubMed]
- Groenendaal, F.; De Vries, L.S. Fifty years of brain imaging in neonatal encephalopathy following perinatal asphyxia. Pediatr. Res. 2017, 81, 150–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giesinger, R.E.; Bailey, L.J.; Deshpande, P.; McNamara, P.J. Hypoxic-Ischemic Encephalopathy and Therapeutic Hypothermia: The Hemodynamic Perspective. J. Pediatr. 2017, 180, 22–30.e2. [Google Scholar] [CrossRef]
- Pelletier, J.-S.; LaBossiere, J.; Dicken, B.; Gill, R.S.; Sergi, C.; Tahbaz, N.; Bigam, D.; Cheung, P.-Y. Low-Dose vasopressin improves cardiac function in newborn piglets with acute hypoxia-reoxygenation. Shock 2013, 40, 320–326. [Google Scholar] [CrossRef]
- Bischoff, A.R.; Habib, S.; McNamara, P.J.; Giesinger, R.E. Hemodynamic response to milrinone for refractory hypoxemia during therapeutic hypothermia for neonatal hypoxic ischemic encephalopathy. J. Perinatol. 2021, 41, 2345–2354. [Google Scholar] [CrossRef]
- Hochwald, O.; Jabr, M.; Osiovich, H.; Miller, S.P.; McNamara, P.J.; Lavoie, P.M. Preferential cephalic redistribution of left ventricular cardiac output during therapeutic hypothermia for perinatal hypoxic-ischemic encephalopathy. J. Pediatr. 2014, 164, 999–1004.e1. [Google Scholar] [CrossRef] [Green Version]
- Sakhuja, P.; More, K.; Ting, J.Y.; Sheth, J.; Lapointe, A.; Jain, A.; McNamara, P.J.; Moore, A.M. Gastrointestinal hemodynamic changes during therapeutic hypothermia and after rewarming in neonatal hypoxic-Ischemic encephalopathy. Pediatr. Neonatol. 2019, 60, 669–675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giesinger, R.E.; El Shahed, A.I.; Castaldo, M.P.; Breatnach, C.R.; Chau, V.; Whyte, H.E.; El-Khuffash, A.F.; Mertens, L.; McNamara, P.J. Impaired right ventricular performance is associated with adverse outcome after hypoxic ischemic encephalopathy. Am. J. Respir. Crit. Care Med. 2019, 200, 1294–1305. [Google Scholar] [CrossRef] [PubMed]
- Thayyil, S.; Pant, S.; Montaldo, P.; Shukla, D.; Oliveira, V.; Ivain, P.; Bassett, P.; Swamy, R.; Mendoza, J.; Moreno-Morales, M.; et al. Hypothermia for moderate or severe neonatal encephalopathy in low-income and middle-income countries (HE-LIX): A randomised controlled trial in India, Sri Lanka, and Bangladesh. Lancet Glob. Health 2021, 9, e1273–e1285. [Google Scholar] [CrossRef]
- Montaldo, P.; Cunnington, A.; Oliveira, V.; Swamy, R.; Bandya, P.; Pant, S.; Lally, P.J.; Ivain, P.; Mendoza, J.; Atreja, G.; et al. Transcriptomic profile of adverse neurodevelopmental outcomes after neonatal encephalopathy. Sci. Rep. 2020, 10, 13100. [Google Scholar] [CrossRef]
- Tetorou, K.; Sisa, C.; Iqbal, A.; Dhillon, K.; Hristova, M. Current therapies for neonatal hypoxic-ischemic and infection-sensitised hypoxic-ischemic brain damage. Front. Synaptic Neurosci. 2021, 13, 1–30. [Google Scholar] [CrossRef]
- Trescher, W.H.; Ishiwa, S.; Johnston, M.V. Brief post-hypoxic-ischemic hypothermia markedly delays neonatal brain injury. Brain Dev. 1997, 19, 326–338. [Google Scholar] [CrossRef]
- Gunn, A.J.; Gunn, T.R.; De Haan, H.H.; Williams, C.E.; Gluckman, P.D. Dramatic neuronal rescue with prolonged selective head cooling after ischemia in fetal lambs. J. Clin. Investig. 1997, 99, 248–256. [Google Scholar] [CrossRef]
- Bingham, A.L.A. Hypothermia for Neonatal Hypoxic-Ischemic Encephalopathy: Different Cooling Regimens and Infants not Included in Prior Trials. In Neonatology Questions and Controversies; Perlman, J.C.M., Ed.; Elsevier: Philadelphia, PA, USA, 2019; pp. 77–91. [Google Scholar]
- Shankaran, S.; Laptook, A.R.; Pappas, A.; McDonald, S.A.; Das, A.; Tyson, J.E.; Poindexter, B.B.; Schibler, K.; Bell, E.F.; Heyne, R.J.; et al. Effect of Depth and Duration of Cooling on Deaths in the NICU Among Neonates With Hypoxic Ischemic Encephalopathy. JAMA 2014, 312, 2629. [Google Scholar] [CrossRef] [Green Version]
- Gunn, A.J.; Gunn, T.R.; Gunning, M.I.; Williams, C.E.; Gluckman, P.D. Neuroprotection with prolonged head cooling started before postischemic seizures in fetal sheep. Pediatrics 1998, 102, 1098–1106. [Google Scholar] [CrossRef]
- Laptook, A.; Tyson, J.; Shankaran, S.; McDonald, S.; Ehrenkranz, R.; Fanaroff, A.; Donovan, E.; Goldberg, R.; O’Shea, T.M.; Higgins, R.D.; et al. Elevated temperature after hypoxic-ischemic encephalopathy: Risk factor for adverse outcomes. Pediatrics 2008, 122, 491–499. [Google Scholar] [CrossRef] [Green Version]
- Kasdorf, E.; Perlman, J. General supportive management of the term Infant with neonatal encephalopathy following intrapartum hypoxia-ischemia. In Neonatology Questions and Controversies; Perlman, J.C.M., Ed.; Elsevier: Philadelphia, PA, USA, 2019; pp. 77–91. [Google Scholar]
- Azzopardi, D.; Robertson, N.J.; Bainbridge, A.; Cady, E.; Charles-Edwards, G.; Deierl, A.; Fagiolo, G.; Franks, N.P.; Griffiths, J.; Hajnal, J.; et al. Moderate hypothermia within 6 h of birth plus inhaled xenon versus moderate hypothermia alone after birth asphyxia (TOBY-Xe): A proof-of-concept, open-label, randomised controlled trial. Lancet Neurol. 2016, 15, 145–153. [Google Scholar] [CrossRef] [Green Version]
- Kelen, D.; Robertson, N.J. Experimental treatments for hypoxic ischemic encephalopathy. Early Hum. Dev. 2010, 86, 369–377. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, F.F.; Abel, R.; Almli, C.R.; Mu, D.; Wendland, M.; Ferriero, D.M. Erythropoietin sustains cognitive function and brain volume after neonatal stroke. Dev. Neurosci. 2009, 31, 403–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aydin, A.; Genç, K.; Akhisaroglu, M.; Yorukoglu, K.; Gokmen, N.; Gonullu, E. Erythropoietin exerts neuroprotective effect in neonatal rat model of hypoxic-ischemic brain injury. Brain Dev. 2003, 25, 494–498. [Google Scholar] [CrossRef]
- Kellert, B.A.; McPherson, R.J.; Juul, S.E. A comparison of high-dose recombinant erythropoietin treatment regimens in brain-injured neonatal rats. Pediatr. Res. 2007, 61, 451–455. [Google Scholar] [CrossRef] [Green Version]
- Razak, A.; Hussain, A. Erythropoietin in perinatal hypoxic-ischemic encephalopathy: A systematic review and meta-analysis. J. Perinat. Med. 2019, 47, 478–489. [Google Scholar] [CrossRef]
- Victor, S.; Rocha-Ferreira, E.; Rahim, A.; Hagberg, H.; Edwards, D. New possibilities for neuroprotection in neonatal hypoxic-ischemic encephalopathy. Eur. J. Pediatr. 2021, 1–13. [Google Scholar] [CrossRef]
- Groenendaal, F. Time to start hypothermia after perinatal asphyxia: Does it matter? BMJ Paediatr. Open 2019, 14, e000494. [Google Scholar] [CrossRef] [PubMed]
- Popescu, M.R.; Panaitescu, A.M.; Pavel, B.; Zagrean, L.; Peltecu, G.; Zagrean, A.-M. Getting an early start in understanding perinatal asphyxia impact on the cardiovascular system. Front. Pediatr. 2020, 8, 1–18. [Google Scholar] [CrossRef]
- Shah, P.; Anvekar, A.; McMichael, J.; Rao, S. Outcomes of infants with Apgar score of zero at 10 min: The West Australian experience. Arch. Dis. Child. Fetal Neonatal Ed. 2015, 100, F492–F494. [Google Scholar] [CrossRef]
- Milner, K.M.; Neal, E.F.G.; Roberts, G.; Steer, A.C.; Duke, T. Long-term neurodevelopmental outcome in high-risk newborns in resource-limited settings: A systematic review of the literature. Paediatr. Int. Child Health 2015, 35, 227–242. [Google Scholar] [CrossRef] [PubMed]
- Martinez, C.; Carneiro, L.; Vernier, L.; Cesa, C.; Guardiola, A.; Vidor, D. Language in children with Neonatal Hypoxic-Ischemic Encephalopathy. Int. Arch. Otorhinolaryngol. 2014, 18, 255–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fisher, P.G. In search of biomarkers for HIE. J. Pediatr. 2018, 194, 3. [Google Scholar] [CrossRef] [PubMed]
- Mota-Rojas, D.; Fierro, R.; Roldan-Santiago, P.; Orozco-Gregorio, H.; González-Lozano, M.; Bonilla, H.; Martínez-Rodríguez, R.; García-Herrera, R.; Mora-Medina, P.; Flores-Peinado, S.; et al. Outcomes of gestation length in relation to farrowing performance in sows and daily weight gain and metabolic profiles in piglets. Anim. Prod. Sci. 2015, 55, 93–100. [Google Scholar] [CrossRef]
- Mota-Rojas, D.; Olmos-Hernández, A.; Verduzco-Mendoza, A.; Lecona-Butrón, H.; Martínez-Burnes, J.; Mora-Medina, P.; Gómez-Prado, J.; Orihuela, A. Infrared thermal imaging associated with pain in laboratory animals. Exp. Anim. 2021, 70, 1–12. [Google Scholar] [CrossRef]
- Martínez-Burnes, J.; Muns, R.; Barrios-García, H.; Villanueva-García, D.; Domínguez-Oliva, A.; Mota-Rojas, D. Parturition in mammals: Animal models, pain and distress. Animals 2021, 11, 2960. [Google Scholar] [CrossRef]
- Villanueva-García, D.; Mota-Rojas, D.; Miranda-Cortés, A.; Ibarra-Ríos, D.; Casas-Alvarado, A.; Mora-Medina, P.; Martínez-Burnes, J.; Olmos-Hernández, A.; Hernández-avalos, I. Caffeine: Cardiorespiratory effects and tissue protection in animal models. Exp. Anim. 2021, 70, 20–0185. [Google Scholar] [CrossRef]
- Villanueva-García, D.; Mota-Rojas, D.; Miranda-Cortés, A.; Mora-Medina, P.; Hernández-Avalos, I.; Casas-Alvarado, A.; Olmos-Hernández, A.; Martínez-Burnes, J. Neurobehavioral and neuroprotector effects of caffeine in animal models. J. Anim. Behav. Biometeorol. 2020, 8, 298–307. [Google Scholar] [CrossRef]
- Redaelli, V.; Papa, S.; Marsella, G.; Grignaschi, G.; Bosi, A.; Ludwig, N.; Luzi, F.; Vismara, I.; Rimondo, S.; Veglianese, P.; et al. A refinement approach in a mouse model of rehabilitation research. Analgesia strategy, reduction approach and infrared thermography in spinal cord injury. PLoS ONE 2019, 14, e0224337. [Google Scholar] [CrossRef]
- Casas-Alvarado, A.; Mota-Rojas, D.; Hernández-Ávalos, I.; Mora-Medina, P.; Olmos-Hernández, A.; Verduzco-Mendoza, A.; Reyes-Sotelo, B.; Martínez-Burnes, J. Advances in infrared thermography: Surgical aspects, vascular changes, and pain monitoring in veterinary medicine. J. Therm. Biol. 2020, 92, 102664. [Google Scholar] [CrossRef]
- Yáñez, A.; Mota-Rojas, D.; Ramírez-Necoechea, R.; Castillo-Rivera, M.; Roldán, P.; Mora-Medina, P.; González-Lozano, M. Application of infrared thermography to assess the effect of different types of environmental enrichment on the ocular, auricular pavilion and nose area temperatures of weaned piglets. Comput. Electron. Agric. 2019, 156, 33–42. [Google Scholar] [CrossRef]
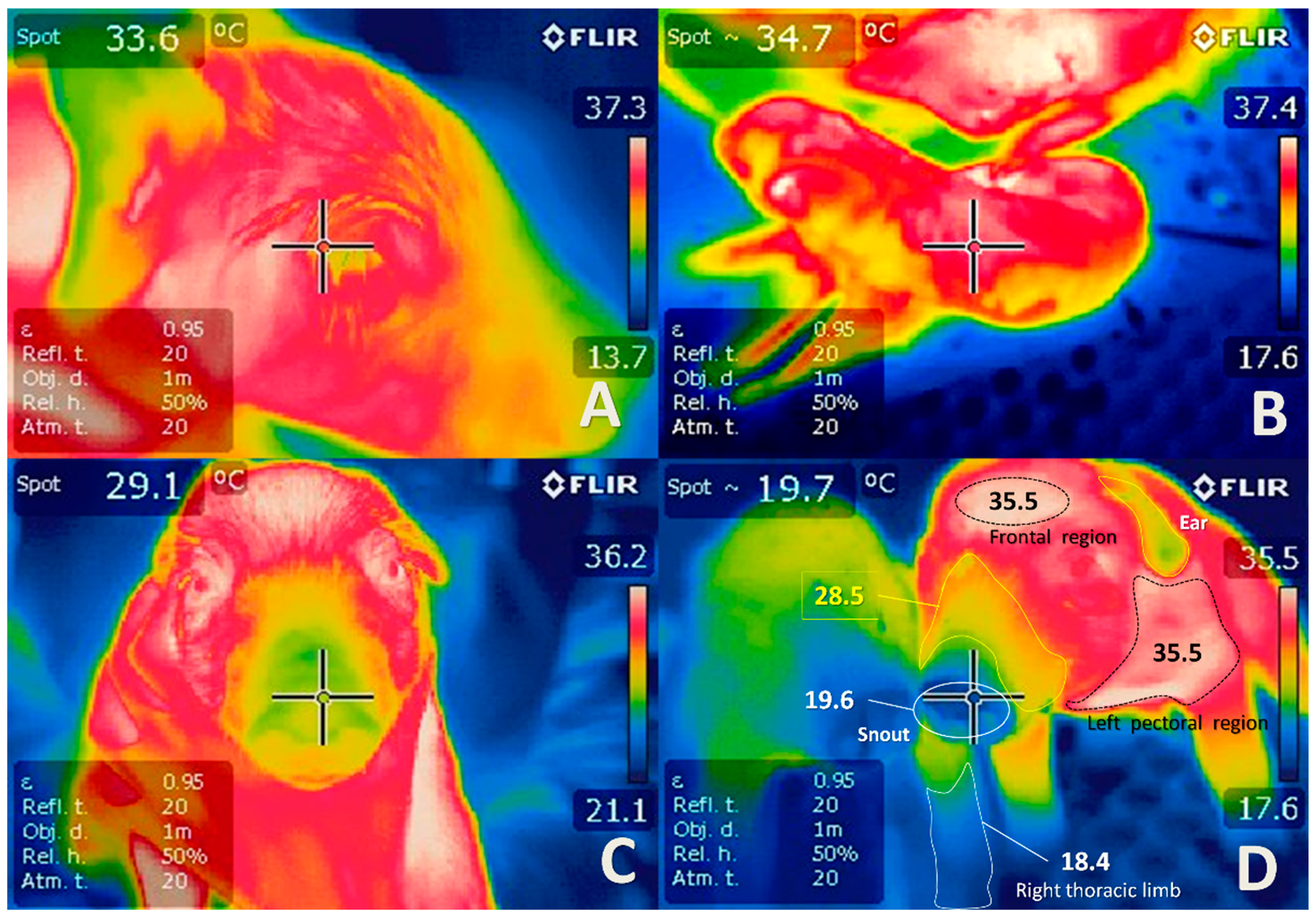
- Mota-Rojas, D.; Wang, D.; Titto, C.G.; Gómez-Prado, J.; Carvajal-de la Fuente, V.; Ghezzi, M.; Boscato-Funes, L.; Barrios-García, H.; Torres-Bernal, F.; Casas-Alvarado, A.; et al. Pathophysiology of fever and application of infrared thermography (IRT) in the detection of sick domestic animals: Recent advances. Animals 2021, 11, 2316. [Google Scholar] [CrossRef] [PubMed]
- Mota-Rojas, D.; Titto, C.G.; Orihuela, A.; Martínez-Burnes, J.; Gómez-Prado, J.; Torres-Bernal, F.; Flores-Padilla, K.; Carvajal-de la Fuente, V.; Wang, D. Physiological and Behavioral Mechanisms of Thermoregulation in Mammals. Animals 2021, 11, 1733. [Google Scholar] [CrossRef] [PubMed]
- Misbe, E.N.J.; Richards, T.L.; McPherson, R.J.; Burbacher, T.M.; Juul, S.E. Perinatal asphyxia in a nonhuman primate model. Dev. Neurosci. 2011, 33, 210–221. [Google Scholar] [CrossRef] [Green Version]
- Manueldas, S.; Benterud, T.; Rueegg, C.S.; Garberg, H.T.; Huun, M.U.; Pankratov, L.; Åsegg-Atneosen, M.; Solberg, R.; Escobar, J.; Saugstad, O.D.; et al. Temporal patterns of circulating cell-free DNA (cfDNA) in a newborn piglet model of perinatal asphyxia. PLoS ONE 2018, 13, e0206601. [Google Scholar] [CrossRef]
- Petersson, K.H.; Pinar, H.; Stopa, E.G.; Sadowska, G.B.; Hanumara, R.C.; Stonestreet, B.S. Effects of exogenous glucose on brain ischemia in ovine fetuses. Pediatr. Res. 2004, 56, 621–629. [Google Scholar] [CrossRef] [Green Version]
- McAdams, R.M.; McPherson, R.J.; Kapur, R.P.; Juul, S.E. Focal brain injury associated with a model of severe hypoxic-ischemic encephalopathy in nonhuman primates. Dev. Neurosci. 2017, 39, 107–123. [Google Scholar] [CrossRef] [Green Version]
- Whitelaw, A.; Thoresen, M. Animal research has been essential to saving babies’ lives. BMJ 2014, 348, g4174. [Google Scholar] [CrossRef]
- Raju, T.N. Some animal models for the study of perinatal asphyxia. Neonatology 1992, 62, 202–214. [Google Scholar] [CrossRef]
- Ball, R.H.; Espinoza, M.I.; Parer, J.T.; Alon, E.; Vertommen, J.; Johnson, J. Regional blood flow in asphyxiated fetuses with seizures. Am. J. Obstet. Gynecol. 1994, 170, 156–161. [Google Scholar] [CrossRef]
- Lear, C.A.; Davidson, J.O.; Mackay, G.R.; Drury, P.P.; Galinsky, R.; Quaedackers, J.S.; Gunn, A.J.; Bennet, L. Antenatal dexamethasone before asphyxia promotes cystic neural injury in preterm fetal sheep by inducing hyperglycemia. J. Cereb. Blood Flow Metab. 2018, 38, 706–718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castillo-Melendez, M.; Baburamani, A.A.; Cabalag, C.; Yawno, T.; Witjaksono, A.; Miller, S.L.; Walker, D.W. Experimental modelling of the consequences of brief late gestation asphyxia on newborn lamb behaviour and brain structure. PLoS ONE 2013, 8, e77377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nardiello, C.; Mižíková, I.; Morty, R.E. Looking ahead: Where to next for animal models of bronchopulmonary dysplasia? Cell Tissue Res. 2017, 367, 457–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solevåg, A.L.; Luong, D.; Lee, T.-F.; O’Reilly, M.; Cheung, P.-Y.; Schmölzer, G.M. Non-perfusing cardiac rhythms in as-phyxiated newborn piglets. PLoS ONE 2019, 14, e0214506. [Google Scholar]
- van Dijk, A.J.; van Loon, J.P.A.M.; Taverne, M.A.M.; Jonker, F.H. Umbilical cord clamping in term piglets: A useful model to study perinatal asphyxia? Theriogenology 2008, 70, 662–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, W.; Xu, X.; Lee, T.-F.; Schmölzer, G.; Cheung, P.-Y. The relationship between heart rate and left ventricular isovolumic relaxation during rormoxia and hypoxia-asphyxia in newborn piglets. Front. Physiol. 2019, 10, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fanos, V.; Noto, A.; Xanthos, T.; Lussu, M.; Murgia, F.; Barberini, L.; Finco, G.; D’Aloja, E.; Papalois, A.; Iacovidou, N.; et al. Metabolomics network characterization of resuscitation after normocapnic hypoxia in a newborn piglet model supports the hypothesis that room air is better. Biomed. Res. Int. 2014, 2014, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Solberg, R.; Enot, D.; Deigner, H.-P.; Koal, T.; Scholl-Bürgi, S.; Saugstad, O.D.; Keller, M. metabolomic analyses of plasma reveals new insights into asphyxia and resuscitation in pigs. PLoS ONE 2010, 5, e9606. [Google Scholar] [CrossRef] [PubMed]
- Ara, J.; Fekete, S.; Frank, M.; Golden, J.A.; Pleasure, D.; Valencia, I. Hypoxic-preconditioning induces neuroprotection against hypoxia–ischemia in newborn piglet brain. Neurobiol. Dis. 2011, 43, 473–485. [Google Scholar] [CrossRef]
- Villanueva-García, D.; Wang, K.T.; Nielsen, H.C.; Ramadurai, S.M. Expression of specific protein kinase c (PKC) isoforms and ligand-specific activation of pkca in late gestation fetal lung. Exp. Lung Res. 2007, 33, 185–196. [Google Scholar] [CrossRef]
- Boynton, B.R.; Villanueva, D.; Hammond, M.D.; Vreeland, P.N.; Buckley, B.; Frantz, I.D. Effect of mean airway pressure on gas exchange during high-frequency oscillatory ventilation. J. Appl. Physiol. 1991, 70, 701–707. [Google Scholar] [CrossRef] [PubMed]
- Salaets, T.; Gie, A.; Tack, B.; Deprest, J.; Toelen, J. modelling bronchopulmonary dysplasia in animals: Arguments for the preterm rabbit model. Curr. Pharm. Des. 2017, 23, 5887–5901. [Google Scholar] [CrossRef] [PubMed]
- Ujhazy, E.; Dubovicky, M.; Navarova, J.; Sedlackova, N.; Danihel, L.; Brucknerova, I.; Mach, M. Subchronic perinatal asphyxia in rats: Embryo–foetal assessment of a new model of oxidative stress during critical period of development. Food Chem. Toxicol. 2013, 61, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Vlassaks, E.; Mencarelli, C.; Nikiforou, M.; Strackx, E.; Ferraz, M.J.; Aerts, J.M.; De Baets, M.H.; Martinez-Martinez, P.; Gavilanes, A.W.D. Fetal asphyxia induces acute and persisting changes in the ceramide metabolism in rat brain. J. Lipid Res. 2013, 54, 1825–1833. [Google Scholar] [CrossRef] [Green Version]
- Galeano, P.; Blanco, E.; Logica Tornatore, T.M.A.; Romero, J.I.; Holubiec, M.I.; RodríGuez De Fonseca, F.; Capani, F. Life-long environmental enrichment counteracts spatial learning, reference and working memory deficits in middle-aged rats subjected to Perinatal asphyxia. Front. Behav. Neurosci. 2015, 8, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Caputa, M.; Rogalska, J.; Wentowska, K.; Nowakowska, A. Perinatal asphyxia, hyperthermia and hyperferremia as factors inducing behavioural disturbances in adulthood: A rat model. Behav. Brain Res. 2005, 163, 246–256. [Google Scholar] [CrossRef]
- Sánchez-Aparicio, P.; Mota-Rojas, D.; Nava-Ocampo, A.A.; Trujillo-Ortega, M.E.; Alfaro-Rodríguez, A.; Arch, E.; Alonso-Spilsbury, M. Effects of sildenafil on the fetal growth of guinea pigs and their ability to survive induced intrapar-tum asphyxia. Am. J. Obstet. Gynecol. 2008, 198, 127.e1–127.e6. [Google Scholar] [CrossRef]
- Coalson, J.J.; Kuehl, T.J.; Escobedo, M.B.; Leonard Hilliard, J.; Smith, F.; Meredith, K.; Null, D.M.; Walsh, W.; Johnson, D.; Robotham, J.L. A baboon model of bronchopulmonary dysplasia. Exp. Mol. Pathol. 1982, 37, 335–350. [Google Scholar] [CrossRef]
- Escobedo, M.B.; Leonard Hilliard, J.; Smith, F.; Meredith, K.; Walsh, W.; Johnson, D.; Coalson, J.J.; Kuehl, T.J.; Null, D.M.; Robotham, J.L. A baboon model of bronchopulmonary dysplasia. I. Clinical features. Exp. Mol. Pathol. 1982, 37, 323–334. [Google Scholar] [CrossRef]
- Yoder, B.A.; Coalson, J.J. Animal models of bronchopulmonary dysplasia. The preterm baboon models. Am. J. Physiol.-Lung Cell Mol. Physiol. 2014, 307, L970–L977. [Google Scholar] [CrossRef] [Green Version]
- Peeples, E.S.; Ezeokeke, C.K.; Juul, S.E.; Mourad, P.D. Evaluating a targeted bedside measure of cerebral perfusion in a nonhuman primate model of neonatal hypoxic-ischemic encephalopathy. J. Ultrasound Med. 2018, 37, 913–920. [Google Scholar] [CrossRef] [PubMed]
- Lewis, K.J.R.; Tibbitt, M.W.; Zhao, Y.; Branchfield, K.; Sun, X.; Balasubramaniam, V.; Anseth, K.S. In vitro model alveoli from photodegradable microsphere templates. Biomater. Sci. 2015, 3, 821–832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beckstrom, A.C.; Humston, E.M.; Snyder, L.R.; Synovec, R.E.; Juul, S.E. Application of comprehensive two-dimensional gas chromatography with time-of-flight mass spectrometry method to identify potential biomarkers of perinatal asphyxia in a non-human primate model. J. Chromatogr. A 2011, 1218, 1899–1906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juul, S.E.; Aylward, E.; Richards, T.; McPherson, R.J.; Kuratani, J.; Burbacher, T.M. Prenatal cord clamping in newborn Macaca nemestrina: A model of perinatal asphyxia. Dev. Neurosci. 2007, 29, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Dunn, W.B.; Wilson, I.D.; Nicholls, A.W.; Broadhurst, D. The importance of experimental design and QC samples in large-scale and MS-driven untargeted metabolomic studies of humans. Bioanalysis 2012, 4, 2249–2264. [Google Scholar] [CrossRef] [Green Version]
- Pomfy, M.; Franko, J. Validation of a four-vessel occlusion model for transient global cerebral ischemia in dogs. J. Hirnforsch. 1999, 39, 465–471. [Google Scholar] [PubMed]
- Uleanya, N.D.; Aniwada, E.C.; Ekwochi, U.; Uleanya, N.D. Short term outcome and predictors of survival among birth asphyxiated babies at a tertiary academic hospital in Enugu, South East, Nigeria. Afr. Health Sci. 2019, 19, 1554. [Google Scholar] [CrossRef] [Green Version]
- Ment, L.R.; Stewart, W.B.; Gore, J.C.; Duncan, C.C. Beagle puppy model of perinatal asphyxia: Alterations in cerebral blood flow and metabolism. Pediatr. Neurol. 1988, 4, 98–104. [Google Scholar] [CrossRef]
- Walsh, W.F.; Butler, D.; Schmidt, J.W. Report of a pilot study of Cooling four preterm infants 32–35 weeks gestation with HIE. J. Neonatal Perinatal Med. 2015, 8, 47–51. [Google Scholar] [CrossRef]
- Choudhary, M.; Sharma, D.; Dabi, D.; Lamba, M.; Pandita, A.; Shastri, S. Hepatic dysfunction in asphyxiated neonates: Prospective case-controlled study. Clin. Med. Insights Pediatr. 2015, 9, 1–6. [Google Scholar] [CrossRef]
- Liang, C.; Hong, Q.; Jiang, T.-T.; Gao, Y.; Yao, X.-F.; Luo, X.-X.; Zhuo, X.-H.; Shinn, J.B.; Jones, R.O.; Zhao, H.-B.; et al. The effects and outcomes of electrolyte disturbances and asphyxia on newborns hearing. Int. J. Pediatr. Otorhinolaryngol. 2013, 77, 1072–1076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aly, H.; Hassanein, S.; Nada, A.; Mohamed, M.H.; Atef, S.H.; Atiea, W. Vascular endothelial growth factor in neonates with perinatal asphyxia. Brain Dev. 2009, 31, 600–604. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species/ Models | Objective | Contribution | Authors |
|---|---|---|---|
| Ovine (fetuses) | Effects of dexamethasone on brain injury due to asphyxia using one dose and a clinically relevant form of administration (12 mg of maternal IM) | It highlights the possible adverse neural effects of glucocorticoid treatment before perinatal asphyxia | [166] |
| Ovine (lambs) | 1. To determine the effects on the survival and the behavior of the lamb of a brief asphyxial attack induced by occlusion of the umbilical cord at 132 days of gestation 2. To report the type and distribution of brain injury present in the newborn after an asphyxia event at 132 days of gestation. | It was shown that brief fetal asphyxia in the uterus in late pregnancy, increase the probability of premature delivery, and the lambs have significant behavioral deficits after birth that appear to arise from the underlying neuropathology caused by asphyxia, and not from premature delivery per se. They identified specific areas of the brain vulnerable to hypoxic damage in late pregnancy. | [167] |
| Ovine (fetuses) | To determine the changes in the regional blood flow of the fetal sheep during severe asphyxia, and with neurological damage (presence of seizures) | The pattern of redistribution of the blood flow of the ovine fetus exposed to severe asphyxia is comparable to the response of the mild asphyxia, except that a significant increase in total cerebral blood flow does not occur, a relevant finding in the likely association with the development of long-term neurological damage | [165] |
| Ovine (fetuses) | To evaluate the consequences of acute hypoxia on arterial and central venous pressures, carotid and femoral blood flows and HR in intact and carotid denervated fetal sheep. | The initial cardiovascular responses to hypoxia in the near-term sheep fetus have a strong carotid chemoreflex component. Moreover, fetal survival during hypoxia is dependent on this chemoreflex and the release of catecholamines from the adrenal medulla. | [61] |
| Ovine (Fetuses) | To evaluate the role of oxidative stress in asphyxia induced perinatal brain injury in near-term fetal lambs subjected to umbilical cord occlusion | Authors suggest that the developing telencephalic white matter seems to be most vulnerable to the effects of intrauterine fetal asphyxia and that oxidative stress may be a significant contributing factor in the pathogenesis of perinatal HIE | [71] |
| Species/ Models | Objective | Contribution | Authors |
|---|---|---|---|
| Porcine (Piglets) | To examine the relationship between isovolumic relaxation time constant (IVR Tau), functional heart parameters. and heart rate (HR) during normoxia and hypoxia–asphyxia (HA) in newborn piglets. | It was demonstrated that HR and IVR Tau significantly accoupled in normoxia; however, they uncoupled during hypoxia–asphyxia (HA) in a piglet model of asphyxia. | [171] |
| Porcine (Piglets) | To establish methods for free DNA evaluation from circulant cells (cfDNA) and to investigate the temporary changes of cfDNA in blood for a clinically relevant piglet model of hypoxia–reoxygenation. | First methodological study for the extraction and evaluation of cfADN using a piglet model of hypoxia–reoxygenation. cfADN could be an early indicator of the damage caused by perinatal asphyxia. | [160] |
| Porcine (Piglets) | To investigate whether different metabolomic profiles are produced according to the oxygen administered during resuscitation. | The results indicated that the use of 21% oxygen seems to be better for resuscitation in piglets with normocapnic hypoxia. | [172] |
| Porcine (Piglets) | To evaluate the effects of asphyxia and resuscitation with different concentrations of oxygen on plasma metabolites in newborn piglets. | Identification of a set of markers with good correlation with the duration of hypoxia. Plasma metabolites indicated an earlier recovery of mitochondrial function when 21% oxygen is used for resuscitation compared to 100% oxygen. | [173] |
| Porcine (Piglets) | To develop an hypoxic-preconditioning (PC) model of ischemic tolerance in newborn piglets that imitates relevant clinical similarities to humans with birth asphyxia and to characterize some of the molecular mechanisms implicated in PC-induced neuroprotection in rodent models. | Results confirm, for the first time, the protective efficacy of PC against hypoxic–ischemic injury in a newborn piglet model, which reiterates many pathophysiological features of asphyxiated human neonates. PC-induced protection in neonatal piglets may involve upregulation of VEGF. | [174] |
| Species/ Models | Objective | Contribution | Authors |
|---|---|---|---|
| Murine (Rats) | To study the neuroprotector role of palmitoylethanolamide (PEA) on the hippocampus of a 30 day-old rat after perinatal asphyxia. | Treatment with PEA (10 mg/kg) during the first hour of life could attenuate the alterations induced by perinatal asphyxia in the CA1 hippocampus neurons. Hence, PEA represents a recognized protective agent for hippocampal disorders. | [26] |
| Murine (Fetal Rats) | To investigate the acute changes that occur in the sphingomyelin/ceramide pathway after sublethal fetal asphyxia injury. To identify relevant molecules for brain tolerance. | Acute and persistent prenatal and postnatal changes in the metabolism of ceramide were found in rat brain under asphyxia, leading to positive regulation of ceramide and an increase in apoptosis. | [179] |
| Murine (Rats) | To evaluate the kinetics of arginine–vasopressin (AVP)/copeptin release during asphyxia and validate the use of the current rodent model in preclinical work on asphyxia at birth | Demonstrated that the proposed rat model meets the standard acid–base criteria for the diagnosis of asphyxia at birth and identified the production of a massive wave of AVP. | [30] |
| Murine (Rats) | To assess whether the ifetime exposure to an enriched environment (EE) (18 months) could counteract the cognitive anomalies observed in middle-aged rats that suffered 19 minutes of asphyxia at birth. | Lifelong EE was able to counteract cognitive anomalies and improved the performance of spatial learning. Results support the relevance of EE across the lifespan to prevent cognitive deficits induced by perinatal asphyxia. | [180] |
| Murine (Rats) | To evaluate the effects of both the physiological body temperature (33 ◦C) and excessive body temperature (37 and 39 ◦C) in neonatal rats exposed to a severe anoxia and of post-anoxic chelation of iron in neonatal rats exposed to both critical anoxia and hyperthermia on stress responses of the animals at the age of 4 months. | Authors concluded that permanent post-anoxic behavioral disorders are caused by iron-dependent oxidative brain injury, which can be prevented by reducing neonatal body temperature. | [181] |
| Guinea Pigs | To determine whether sildenafil increased fetal weight and favored fetal tolerance to induced asphyxia at birth. | Low doses of sildenafil administered from day 35 to the end of pregnancy favored fetal tolerability of intrapartum-induced asphyxia. High doses of sildenafil increased fetal weight. | [182] |
| Species/ Models | Objective | Contribution | Authors |
|---|---|---|---|
| Primates (Macaca nemestrina) | To investigate the sse of metabolomic and analysis tools to detect potential biomarkers of perinatal asphyxia. To evaluate a model of asphyxia by clamping the umbilical cord and to evaluate the differences between pre- and post-asphyxia. | Through metabolomic analyses, a profile of metabolites was identified with a significant elevation in response to asphyxia at birth (succinic acid, lactate, glucose, malate, arachidonic acid, glutamate, and butanoic acid, among others). | [188] |
| Primates (Macaca nemestrina) | To evaluate the safety and efficacy of erythropoietin (EPO) plus hypothermia for the treatment of perinatal HIE in a non-human primate model. To characterize the acute and chronic consequences of perinatal asphyxia with diagnostic imaging tools to correlate brain injury and neurodevelopmental tests to evaluate early motor and cognitive outcomes. | Occlusion of the umbilical cord for between 15 and 18 minutes can induce severe asphyxia at birth. Asphyxiated neonates developed long-term physical and cognitive deficits. | [159] |
| Primates (Macaca nemestrina) | To establish a non-human primate model of perinatal asphyxia suitable for preclinical evaluation of neuroprotective treatment strategies in conditions resembling human neonatal emergencies and testing erythropoietin neuroprotective treatment. | The model demonstrated changes in magnetic resonance/spectroscopy images consistent with hypoxia, significant motor and behavioral anomalies, and evidence of brain gliosis and was found to be an appropriate model of moderate-to-severe perinatal hypoxic–ischemic injury | [189] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mota-Rojas, D.; Villanueva-García, D.; Solimano, A.; Muns, R.; Ibarra-Ríos, D.; Mota-Reyes, A. Pathophysiology of Perinatal Asphyxia in Humans and Animal Models. Biomedicines 2022, 10, 347. https://doi.org/10.3390/biomedicines10020347
Mota-Rojas D, Villanueva-García D, Solimano A, Muns R, Ibarra-Ríos D, Mota-Reyes A. Pathophysiology of Perinatal Asphyxia in Humans and Animal Models. Biomedicines. 2022; 10(2):347. https://doi.org/10.3390/biomedicines10020347
Chicago/Turabian StyleMota-Rojas, Daniel, Dina Villanueva-García, Alfonso Solimano, Ramon Muns, Daniel Ibarra-Ríos, and Andrea Mota-Reyes. 2022. "Pathophysiology of Perinatal Asphyxia in Humans and Animal Models" Biomedicines 10, no. 2: 347. https://doi.org/10.3390/biomedicines10020347
APA StyleMota-Rojas, D., Villanueva-García, D., Solimano, A., Muns, R., Ibarra-Ríos, D., & Mota-Reyes, A. (2022). Pathophysiology of Perinatal Asphyxia in Humans and Animal Models. Biomedicines, 10(2), 347. https://doi.org/10.3390/biomedicines10020347