
Identification of Altered Evoked and Non-Evoked Responses in a Heterologous Mouse Model of Endometriosis-Associated Pain

 , , ,
, , ,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
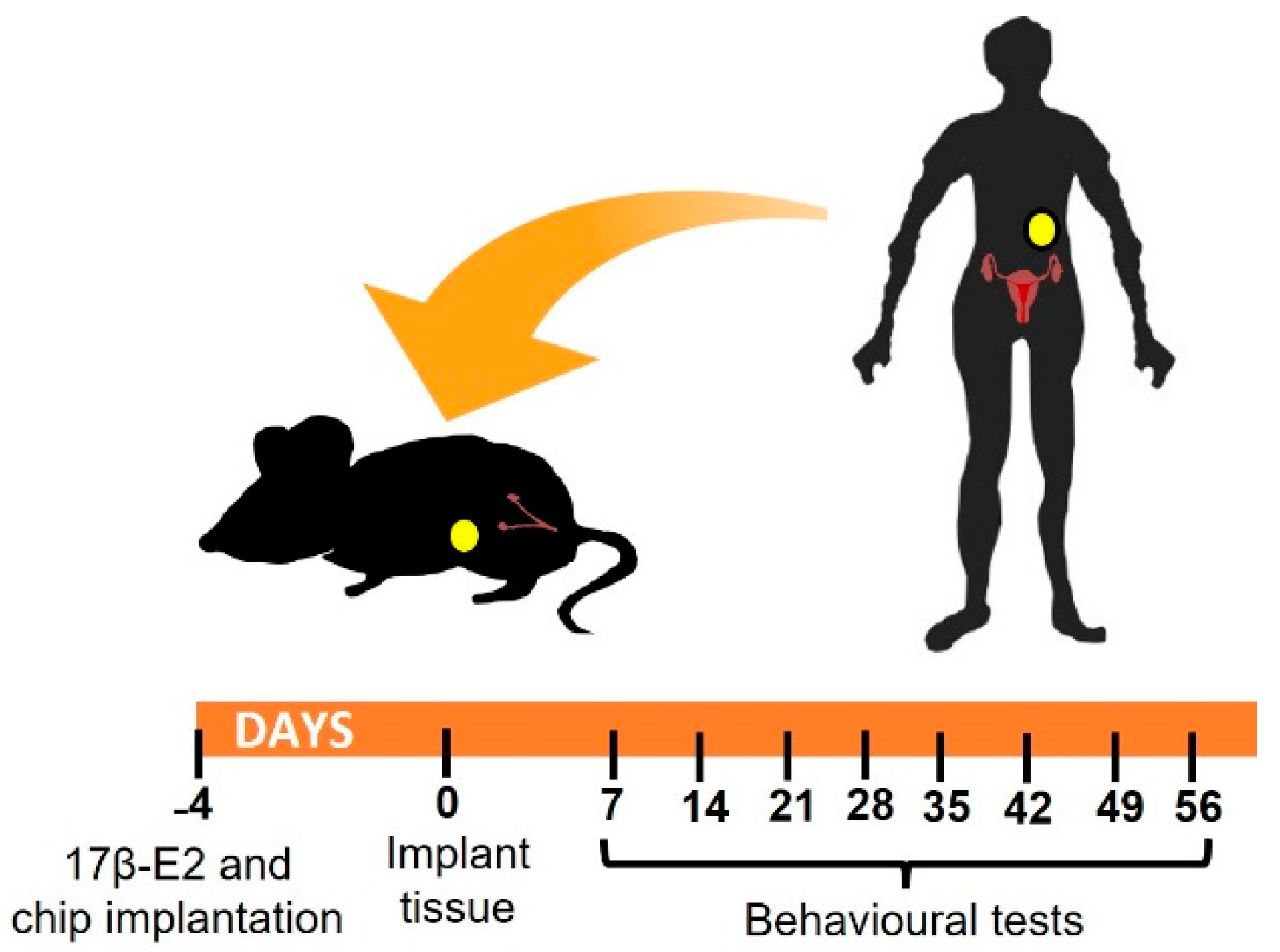
2.1. Experimental Design
2.2. Heterologous Mouse Model of Endometriosis
2.2.1. Preparation of Endometriosis Recipient Mice: Oestradiol Pellet and Chip Implantation
2.2.2. Obtention and Preprocessing of Human Endometrial Biopsies
2.2.3. Endometrial Fragment Implantation
2.3. Behavioral Tests
2.3.1. Mechanical Hypersensitivity
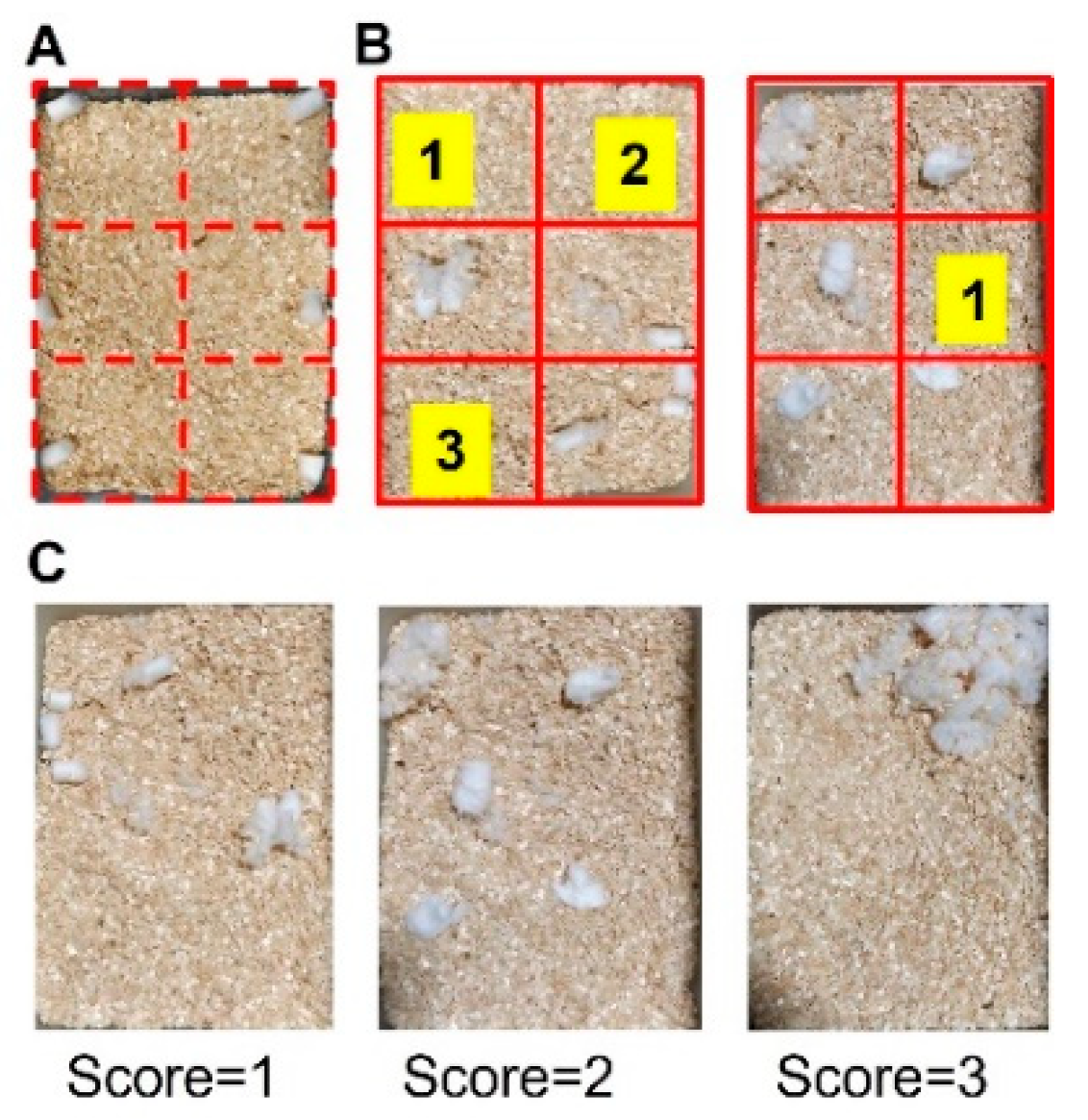
2.3.2. Nesting
2.3.3. Home Cage Activity (HCA)
2.4. Histology and Immunofluorescence
2.5. Statistical Analysis
3. Results
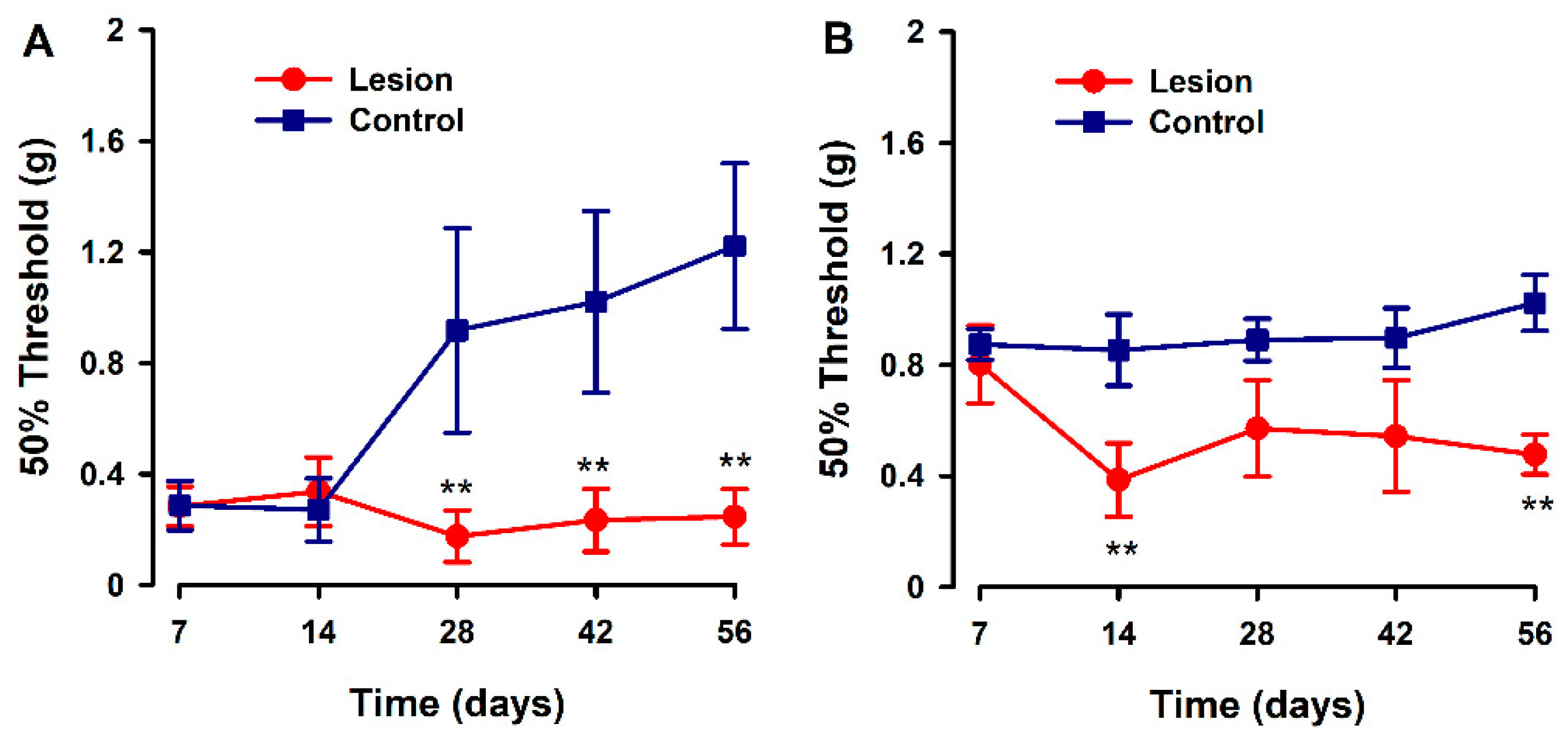
3.1. Mice with Human Endometriotic Lesions Present with Decreased Abdominal and Hind Paw Thresholds in Von Frey Tests
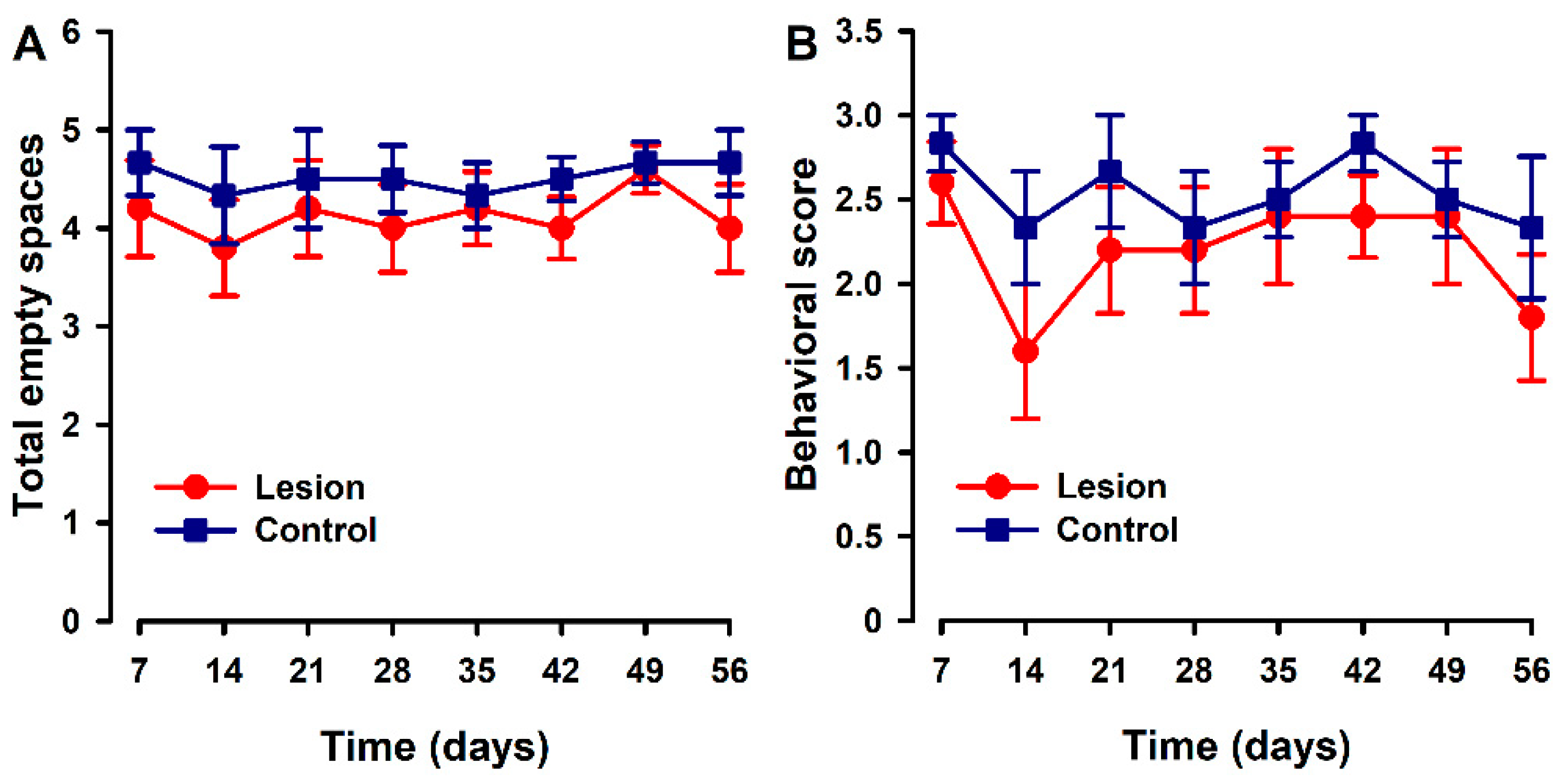
3.2. Nesting Behaviors Remained Unchanged in the Heterologous Model
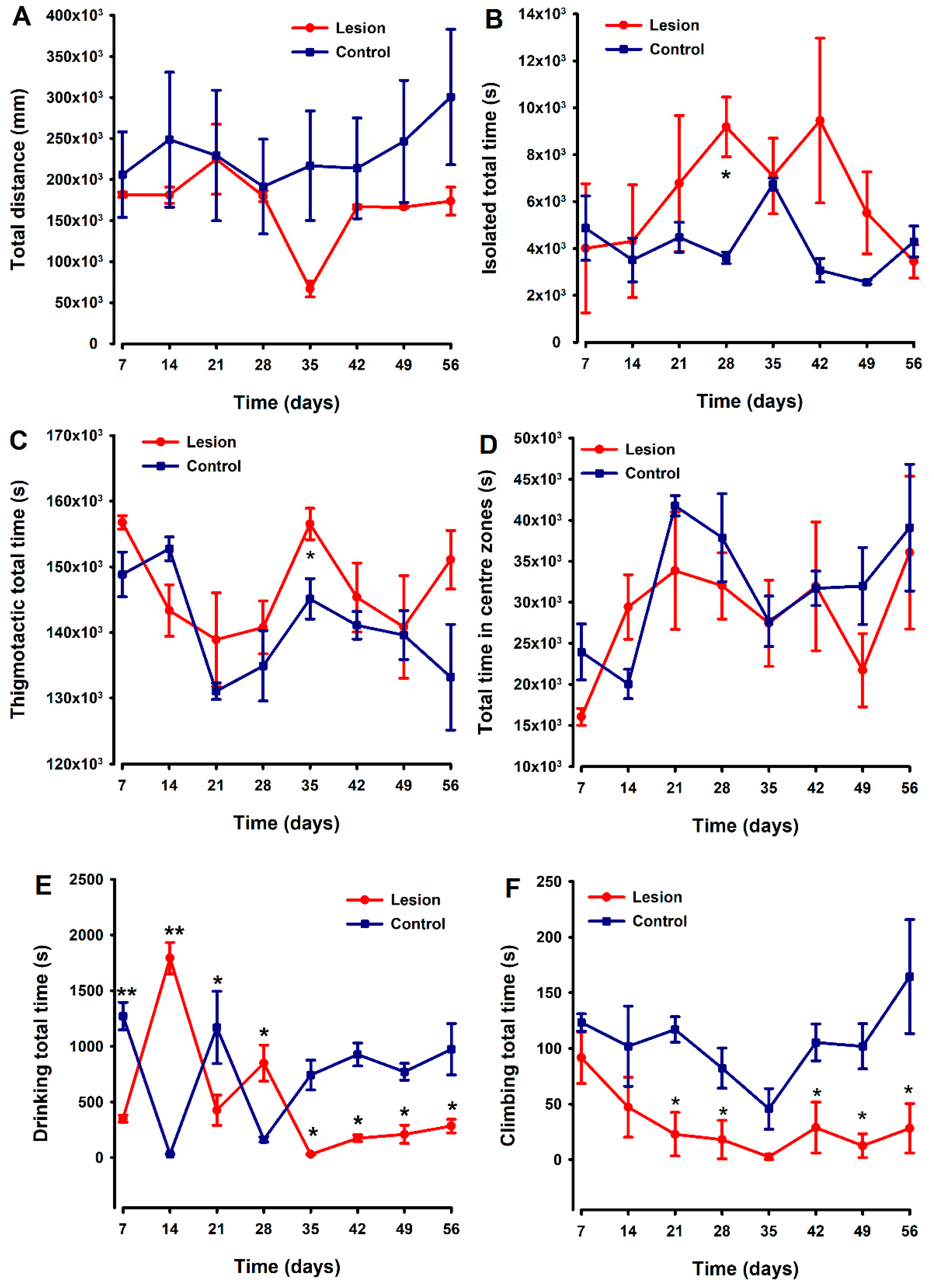
3.3. Mice with Human Endometriotic Lesions Present with Decreased Climbing and Drinking during Home Cage Analysis (HCA)
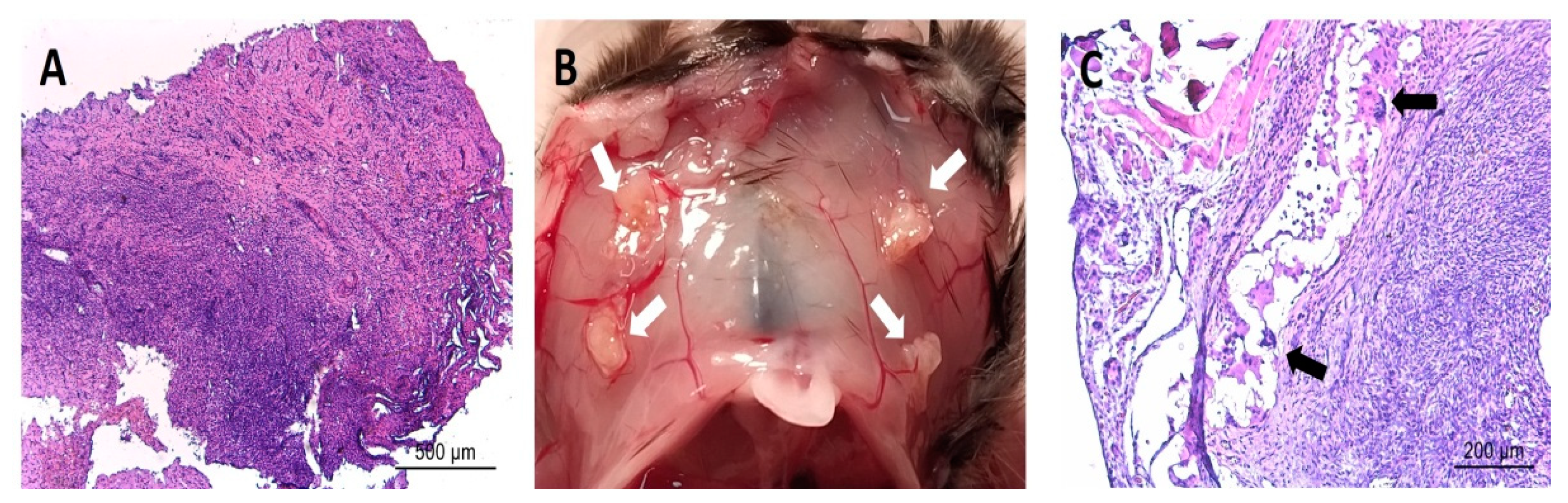
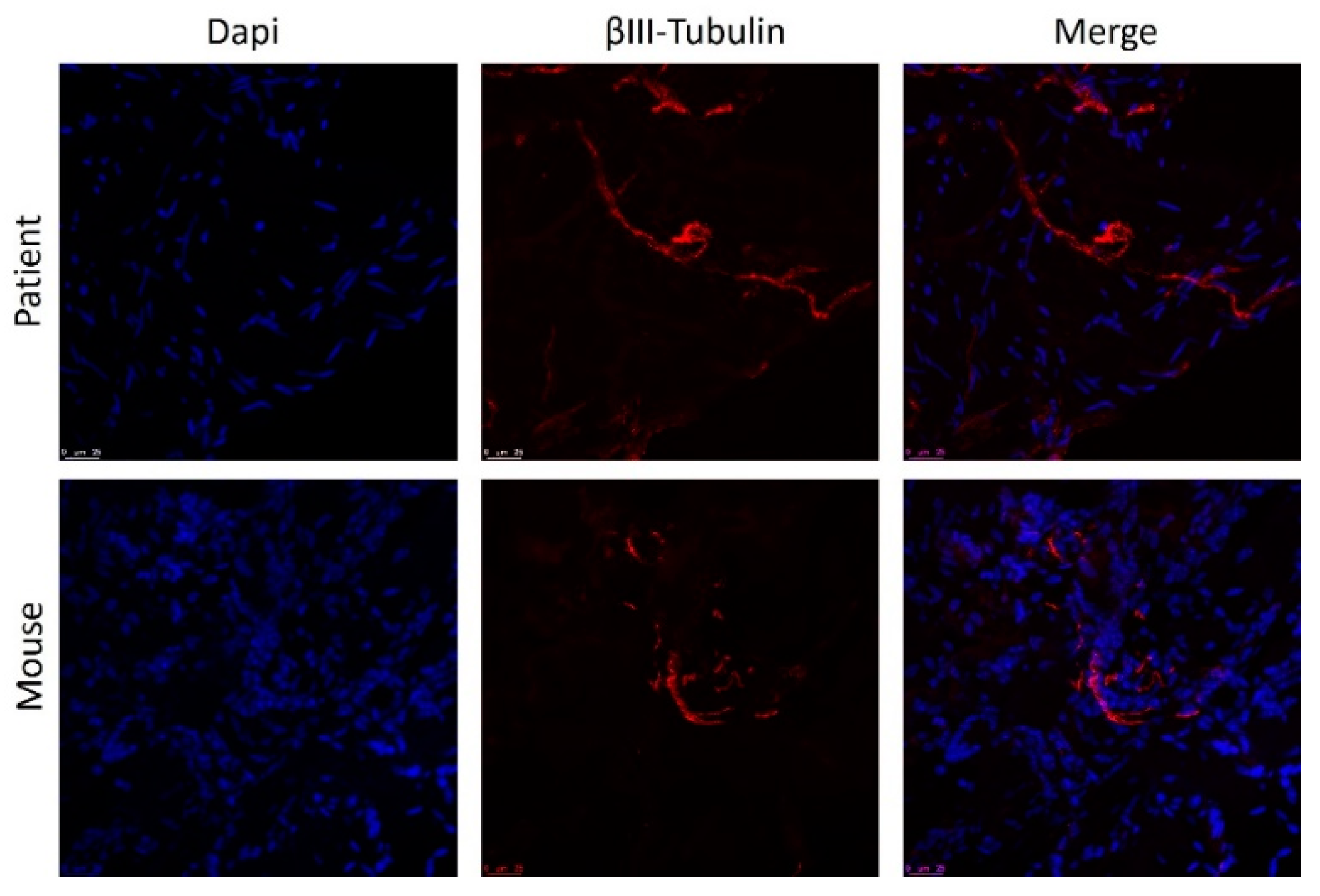
3.4. Post-Implantation Lesions Preserve the Histological Architecture and Nerve Fiber Density of Human Lesions at Time of Surgery
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zondervan, K.T.; Becker, C.M.; Missmer, S.A. Endometriosis. N. Engl. J. Med. 2020, 382, 1244–1256. [Google Scholar] [CrossRef] [PubMed]
- Giudice, L.C. Endometriosis. N. Engl. J. Med. 2010, 362, 2389–2398. [Google Scholar] [CrossRef] [PubMed]
- Shafrir, A.L.; Farland, L.V.; Shah, D.K.; Harris, H.R.; Kvaskoff, M.; Zondervan, K.; Missmer, S.A. Risk for and consequences of endometriosis: A critical epidemiologic review. Best Pract. Res. Clin. Obstet. Gynaecol. 2018, 51, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Saunders, P.T.K.; Horne, A.W. Endometriosis: Etiology, pathobiology, and therapeutic prospects. Cell 2021, 184, 2807–2824. [Google Scholar] [CrossRef] [PubMed]
- Johnson, N.P.; Hummelshoj, L. Consensus on current management of endometriosis. Hum. Reprod. 2013, 28, 1552–1568. [Google Scholar] [CrossRef] [PubMed]
- Simoens, S.; Dunselman, G.; Dirksen, C.; Hummelshoj, L.; Bokor, A.; Brandes, I.; Brodszky, V.; Canis, M.; Colombo, G.L.; DeLeire, T.; et al. The burden of endometriosis: Costs and quality of life of women with endometriosis and treated in referral centres. Hum. Reprod. 2012, 27, 1292–1299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falcone, T.; Flyckt-Rebecca, R. Clinical management of endometriosis. Obstet. Gynecol. 2018, 131, 557–571. [Google Scholar] [CrossRef] [Green Version]
- Mirkin, D.; Murphy-Barron, C.; Iwasaki, K. Actuarial analysis of private payer administrative claims data for women with endometriosis. J. Manag. Care Pharm. 2007, 13, 262–272. [Google Scholar] [CrossRef]
- Ahn, S.H.; Singh, V.; Tayade, C. Biomarkers in endometriosis: Challenges and opportunities. Fertil. Steril. 2017, 107, 523–532. [Google Scholar] [CrossRef] [Green Version]
- Simoens, S.; Hummelshoj, L.; Dunselman, G.; Brandes, I.; Dirksen, C.; D’Hooghe, T. Endometriosis cost assessment (the EndoCost Study): A cost-of-illness study protocol. Gynecol. Obstet. Investig. 2011, 71, 170–176. [Google Scholar] [CrossRef]
- Falcone, T.; Lebovic, D.I. Clinical management of endometriosis. Obstet. Gynecol. 2011, 118, 691–705. [Google Scholar] [CrossRef] [PubMed]
- Soliman, A.M.; Surrey, E.S.; Bonafede, M.; Nelson, J.K.; Vora, J.B.; Agarwal, S.K. Health care utilization and costs associated with endometriosis among women with Medicaid insurance. J. Manag. Care Spec. Pharm. 2019, 25, 566–572. [Google Scholar] [CrossRef] [PubMed]
- Gellersen, B.; Brosens, J.J. Cyclic decidualization of the human endometrium in reproductive health and failure. Endocr. Rev. 2014, 35, 851–905. [Google Scholar] [CrossRef]
- Greaves, E.; Critchley, H.O.D.; Horne, A.W.; Saunders, P.T.K. Relevant human tissue resources and laboratory models for use in endometriosis research. Acta Obstet. Gynecol. Scand. 2017, 96, 644–658. [Google Scholar] [CrossRef] [PubMed]
- Nunez-Badinez, P.; De Leo, B.; Laux-Biehlmann, A.; Hoffmann, A.; Zollner, T.M.; Saunders, P.T.K.; Simitsidellis, I.; Charrua, A.; Cruz, F.; Gomez, R.; et al. Preclinical models of endometriosis and interstitial cystitis/bladder pain syndrome: An Innovative Medicines Initiative-PainCare initiative to improve their value for translational research in pelvic pain. Pain 2021, 162, 2349–2365. [Google Scholar] [CrossRef]
- Du Sert, P.N.; Rice, A.S. Improving the translation of analgesic drugs to the clinic: Animal models of neuropathic pain. Br. J. Pharmacol. 2014, 171, 2951–2963. [Google Scholar] [CrossRef] [Green Version]
- Tappe-Theodor, A.; King, T.; Morgan, M.M. Pros and cons of clinically relevant methods to assess pain in rodents. Neurosci. Biobehav. Rev. 2019, 100, 335–343. [Google Scholar] [CrossRef]
- Tappe-Theodor, A.; Kuner, R. Studying ongoing and spontaneous pain in rodents—Challenges and opportunities. Eur. J. Neurosci. 2014, 39, 1881–1890. [Google Scholar] [CrossRef]
- Wodarski, R.; Delaney, A.; Ultenius, C.; Morland, R.; Andrews, N.; Baastrup, C.; Bryden, L.A.; Caspani, O.; Christoph, T.; Gardiner, N.J.; et al. Cross-centre replication of suppressed burrowing behaviour as an ethologically relevant pain outcome measure in the rat: A prospective multicentre study. Pain 2016, 157, 2350–2365. [Google Scholar] [CrossRef] [Green Version]
- Alvarez, P.; Chen, X.; Hendrich, J.; Irwin, J.C.; Green, P.G.; Giudice, L.C.; Levine, J.D. Ectopic uterine tissue as a chronic pain generator. Neuroscience 2012, 225, 269–282. [Google Scholar] [CrossRef] [Green Version]
- Tejada, M.Á.; Santos-Llamas, A.I.; Fernández-Ramírez, M.J.; Tarín, J.J.; Cano, A.; Gómez, R. A reassessment of the therapeutic potential of a dopamine receptor 2 agonist (D2-AG) in endometriosis by comparison against a standardized antiangiogenic treatment. Biomedicines 2021, 9, 269. [Google Scholar] [CrossRef] [PubMed]
- Martinez, J.; Bisbal, V.; Marin, N.; Cano, A.; Gómez, R. Noninvasive monitoring of lesion size in a heterologous mouse model of endometriosis. J. Vis. Exp. 2019, 26, 144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaplan, S.R.; Bach, F.W.; Pogrel, J.W.; Chung, J.M.; Yaksh, T.L. Quantitative assessment of tactile allodynia in the rat paw. J. Neurosci. Methods 1994, 53, 55–63. [Google Scholar] [CrossRef]
- Negus, S.S.; Neddenriep, B.; Altarifi, A.A.; Carroll, F.I.; Leitl, M.D.; Miller, L.L. Effects of ketoprofen, morphine, and kappa opioids on pain-related depression of nesting in mice. Pain 2015, 156, 1153–1160. [Google Scholar] [CrossRef] [Green Version]
- Deacon, R.M. Assessing nest building in mice. Nat. Protoc. 2006, 1, 1117–1119. [Google Scholar] [CrossRef]
- Bains, R.S.; Cater, H.L.; Sillito, R.R.; Chartsias, A.; Sneddon, D.; Concas, D.; Keskivali-Bond, P.; Lukins, T.C.; Wells, S.; Acevedo Arozena, A.; et al. Analysis of individual mouse activity in group housed animals of different inbred strains using a novel automated home cage analysis system. Front. Behav. Neurosci. 2016, 10, 106. [Google Scholar] [CrossRef] [Green Version]
- Greaves, E.; Collins, F.; Esnal-Zufiaurre, A.; Giakoumelou, S.; Horne, A.W.; Saunders, P.T.K. Estrogen receptor (ER) agonists differentially regulate neuroangiogenesis in peritoneal endometriosis via the repellent factor SLIT3. Endocrinology 2014, 155, 4015–4026. [Google Scholar] [CrossRef]
- Greaves, E.; Horne, A.W.; Jerina, H.; Mikolajczak, M.; Hilferty, L.; Mitchell, R.; Fleetwood-Walker, S.M.; Saunders, P.T. EP2 receptor antagonism reduces peripheral and central hyperalgesia in a preclinical mouse model of endometriosis. Sci. Rep. 2017, 7, 44169. [Google Scholar] [CrossRef] [Green Version]
- Eggermont, J.; Donnez, J.; Casanas-Roux, F.; Scholtes, H.; Van Langendonckt, A. Time course of pelvic endometriotic lesion revascularization in a nude mouse model. Fertil. Steril. 2005, 84, 492–499. [Google Scholar] [CrossRef]
- Asante, A.; Taylor, R.N. Endometriosis: The role of neuroangiogenesis. Annu. Rev. Physiol. 2011, 73, 163–182. [Google Scholar] [CrossRef] [Green Version]
- Woolf, C.J. Central sensitization: Implications for the diagnosis and treatment of pain. Pain 2011, 152, S2–S15. [Google Scholar] [CrossRef] [PubMed]
- Ge, P.; Ren, J.; Harrington, A.M.; Grundy, L.; Castro, J.; Brierley, S.M.; Hannig, G. Linaclotide treatment reduces endometriosis-associated vaginal hyperalgesia and mechanical allodynia through viscerovisceral cross-talk. Pain 2019, 160, 2566–2579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lian, Y.L.; Cheng, M.J.; Zhang, X.X.; Wang, L. Elevated expression of transient receptor potential vanilloid type 1 in dorsal root ganglia of rats with endometriosis. Mol. Med. Rep. 2017, 16, 1920–1926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fattori, V.; Franklin, N.S.; Gonzalez-Cano, R.; Peterse, D.; Ghalali, A.; Madrian, E.; Verri, W.A., Jr.; Andrews, N.; Woolf, C.J.; Rogers, M.S. Nonsurgical mouse model of endometriosis-associated pain that responds to clinically active drugs. Pain 2020, 161, 1321–1331. [Google Scholar] [CrossRef] [PubMed]
- Bilbao, A.; Leixner, S.; Wei, S.; Cantacorps, L.; Valverde, O.; Spanagel, R. Reduced sensitivity to ethanol and excessive drinking in a mouse model of neuropathic pain. Addict. Biol. 2019, 24, 1008–1018. [Google Scholar] [CrossRef] [PubMed]
- Falk, S.; Gallego-Pedersen, S.; Petersen, N.C. Grid-climbing behaviour as a pain measure for cancer-induced bone pain and neuropathic pain. In Vivo 2017, 31, 619–623. [Google Scholar] [CrossRef] [Green Version]
- McAllister, S.L.; Dmitrieva, N.; Berkley, K.J. Sprouted innervation into uterine transplants contributes to the development of hyperalgesia in a rat model of endometriosis. PLoS ONE 2012, 7, e31758. [Google Scholar] [CrossRef]
- Anaf, V.; Chapron, C.; El Nakadi, I.; De Moor, V.; Simonart, T.; Noël, J.C. Pain, mast cells, and nerves in peritoneal, ovarian, and deep infiltrating endometriosis. Fertil. Steril. 2006, 86, 1336–1343. [Google Scholar] [CrossRef]
- Raimondo, D.; Mastronardi, M.; Mabrouk, M.; Cafagna, G.; Salucci, P.; Arena, A.; Iodice, R.; Borghese, G.; Casadio, P.; Del Forno, S.; et al. Rectosigmoid endometriosis vascular patterns at intraoperative indocyanine green angiography and their correlation with clinicopathological data. Surg. Innov. 2020, 27, 474–480. [Google Scholar] [CrossRef]
- McKinnon, B.; Bersinger, N.A.; Wotzkow, C.; Mueller, M.D. Endometriosis-associated nerve fibers, peritoneal fluid cytokine concentrations, and pain in endometriotic lesions from different locations. Fertil. Steril. 2012, 97, 373–380. [Google Scholar] [CrossRef]
- Hogg, C.; Panir, K.; Dhami, P.; Rosser, M.; Mack, M.; Soong, D.; Pollard, J.W.; Jenkins, S.J.; Horne, A.W.; Greaves, E. Macrophages inhibit and enhance endometriosis depending on their origin. Proc. Natl. Acad. Sci. USA 2021, 118, e2013776118. [Google Scholar] [CrossRef] [PubMed]
- Dorning, A.; Dhami, P.; Panir, K.; Hogg, C.; Park, E.; Ferguson, G.D.; Hargrove, D.; Karras, J.; Horne, A.W.; Greaves, E. Bioluminescent imaging in induced mouse models of endometriosis reveals differences in four model variations. Dis. Models Mech. 2021, 14, dmm049070. [Google Scholar] [CrossRef] [PubMed]
- González-Cano, R.; Montilla-García, Á.; Ruiz-Cantero, M.C.; Bravo-Caparrós, I.; Tejada, M.Á.; Nieto, F.R.; Cobos, E.J. The search for translational pain outcomes to refine analgesic development: Where did we come from and where are we going? Neurosci. Biobehav. Rev. 2020, 113, 238–261. [Google Scholar] [CrossRef] [PubMed]
- Hobson, L.; Bains, R.S.; Greenaway, S.; Wells, S.; Nolan, P.M. Phenotyping in mice using continuous home cage monitoring and ultrasonic vocalization recordings. Curr. Protoc. Mouse Biol. 2020, 10, e80. [Google Scholar] [CrossRef]
- Berkley, K.J.; Dmitrieva, N.; Curtis, K.S.; Papka, R.E. Innervation of ectopic endometrium in a rat model of endometriosis. Proc. Natl. Acad. Sci. USA 2004, 101, 11094–11098. [Google Scholar] [CrossRef] [Green Version]
- Greaves, E.; Cousins, F.L.; Murray, A.; Esnal-Zufiaurre, A.; Fassbender, A.; Horne, A.W.; Saunders, P.T.K. A novel mouse model of endometriosis mimics human phenotype and reveals insights into the inflammatory contribution of shed endometrium. Am. J. Pathol. 2014, 184, 1930–1939. [Google Scholar] [CrossRef] [Green Version]
- Greaves, E.; Rosser, M.; Saunders, P.T.K. Endometriosis-associated pain—Do preclinical rodent models provide a good platform for translation? Adv. Anat. Embryol. Cell Biol. 2020, 232, 25–55. [Google Scholar] [CrossRef]
- Colette, S.; Defrère, S.; Lousse, J.C.; Van Langendonckt, A.; Loumaye, E.; Donnez, J. Evaluation of estrogen treatment in an immunodeficient mouse endometriosis model. Gynecol. Obstet. Invest. 2009, 68, 262–268. [Google Scholar] [CrossRef]
- Persoons, E.; De Clercq, K.; Van den Eynde, C.; Pinto, S.J.P.C.; Luyten, K.; Van Bree, R.; Tomassetti, C.; Voets, T.; Vriens, J. Mimicking sampson’s retrograde menstrual theory in rats: A new rat model for ongoing endometriosis-associated pain. Int. J. Mol. Sci. 2020, 21, 2326. [Google Scholar] [CrossRef] [Green Version]
- Castro, J.; Maddern, J.; Grundy, L.; Manavis Harrington, A.M.; Schober, G.; Brierley, S.M. A mouse model of endometriosis that displays vaginal, colon, cutaneous, and bladder sensory comorbidities. FASEB J. 2021, 35, e21430. [Google Scholar] [CrossRef]
- Simitsidellis, I.; Gibson, D.A.; Saunders, P.T.K. Animal models of endometriosis: Replicating the aetiology and symptoms of the human disorder. Best Pract. Res. Clin. Endocrinol. Metab. 2018, 32, 257–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tejada, M.A.; Santos-Llamas, A.I.; Escriva, L.; Tarin, J.J.; Cano, A.; Fernández-Ramírez, M.J.; Nunez-Badinez, P.; De Leo, B.; Saunders, P.T.K.; Vidal, V.; et al. Identification of Altered Evoked and Non-Evoked Responses in a Heterologous Mouse Model of Endometriosis-Associated Pain. Biomedicines 2022, 10, 501. https://doi.org/10.3390/biomedicines10020501
Tejada MA, Santos-Llamas AI, Escriva L, Tarin JJ, Cano A, Fernández-Ramírez MJ, Nunez-Badinez P, De Leo B, Saunders PTK, Vidal V, et al. Identification of Altered Evoked and Non-Evoked Responses in a Heterologous Mouse Model of Endometriosis-Associated Pain. Biomedicines. 2022; 10(2):501. https://doi.org/10.3390/biomedicines10020501
Chicago/Turabian StyleTejada, Miguel A., Ana I. Santos-Llamas, Lesley Escriva, Juan J. Tarin, Antonio Cano, Maria J. Fernández-Ramírez, Paulina Nunez-Badinez, Bianca De Leo, Philippa T. K. Saunders, Victor Vidal, and et al. 2022. "Identification of Altered Evoked and Non-Evoked Responses in a Heterologous Mouse Model of Endometriosis-Associated Pain" Biomedicines 10, no. 2: 501. https://doi.org/10.3390/biomedicines10020501
APA StyleTejada, M. A., Santos-Llamas, A. I., Escriva, L., Tarin, J. J., Cano, A., Fernández-Ramírez, M. J., Nunez-Badinez, P., De Leo, B., Saunders, P. T. K., Vidal, V., Barthas, F., Vincent, K., Sweeney, P. J., Sillito, R. R., Armstrong, J. D., Nagel, J., & Gomez, R. (2022). Identification of Altered Evoked and Non-Evoked Responses in a Heterologous Mouse Model of Endometriosis-Associated Pain. Biomedicines, 10(2), 501. https://doi.org/10.3390/biomedicines10020501