
Towards Understanding the Lymph Node Response to Skin Infection with Saprophytic Staphylococcus epidermidis


Abstract
:1. Introduction
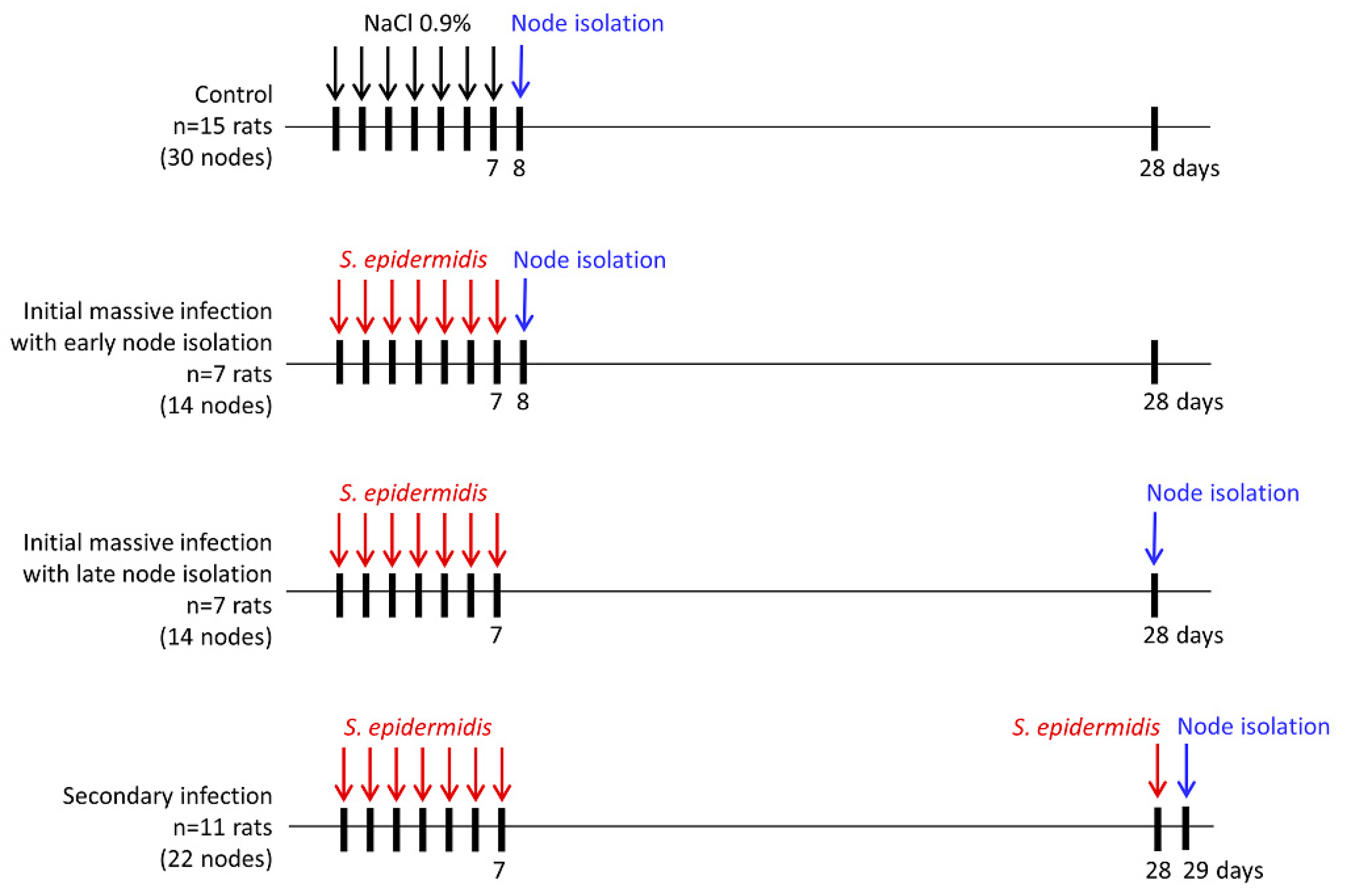
2. Materials and Methods
2.1. Animals
2.2. Bacterial Infection
2.3. Cells Isolation
2.4. Flow Cytometry
2.5. Immunohistochemistry and Quantitative Analysis of Cell Phenotypes
2.6. Statistical Analysis
3. Results
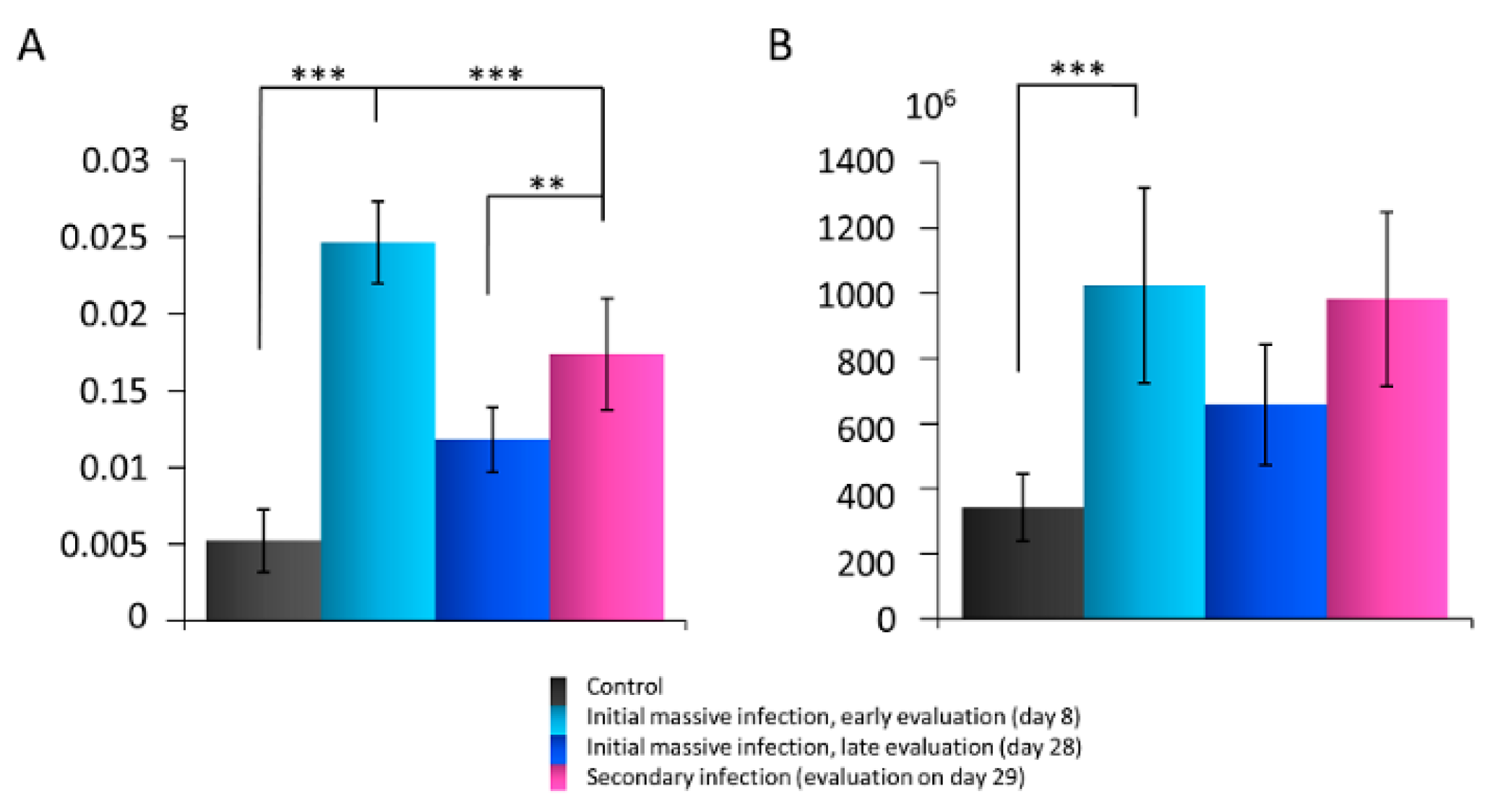
3.1. The Lymph Node Mass and Cell Number after Skin Infection with S. epidermidis
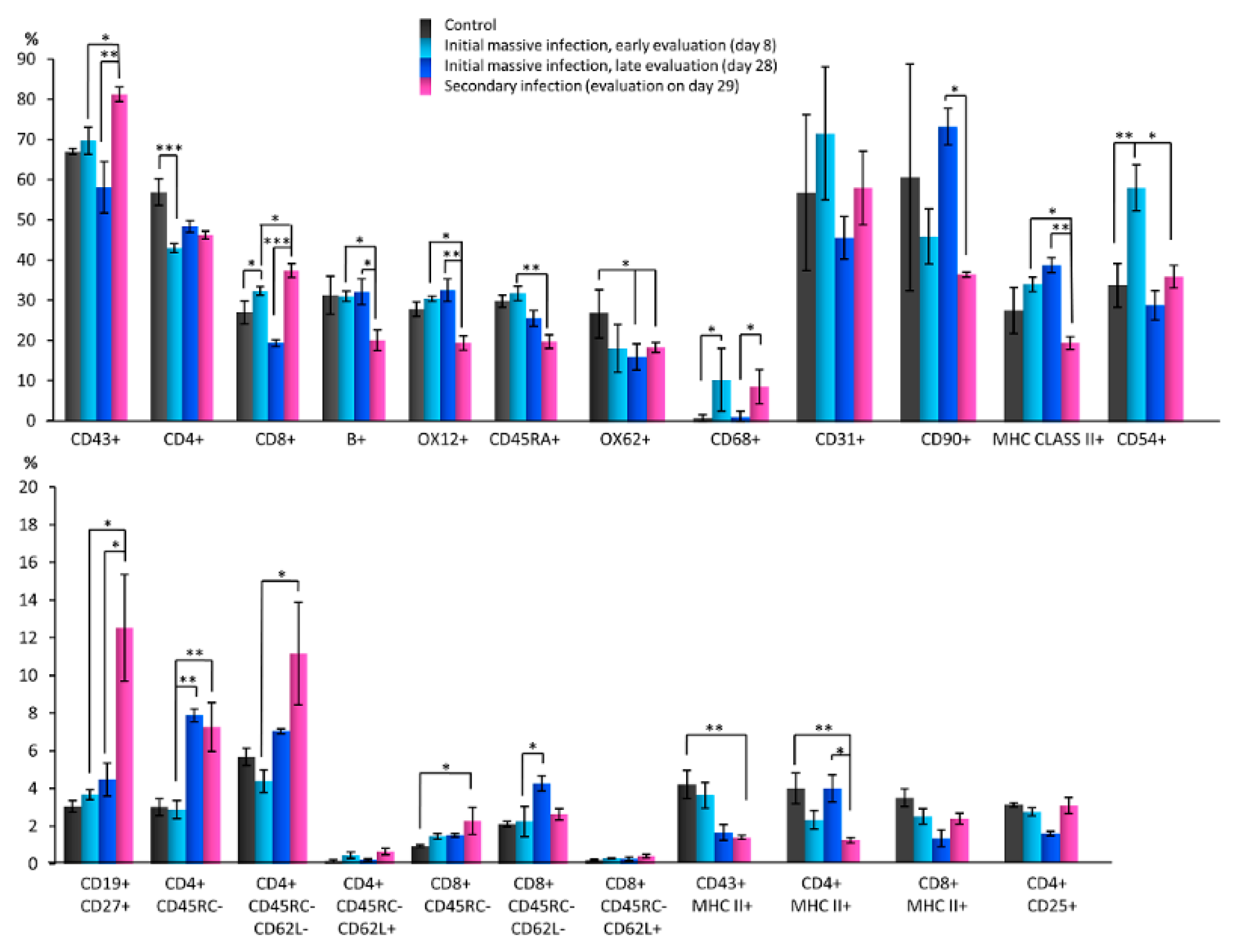
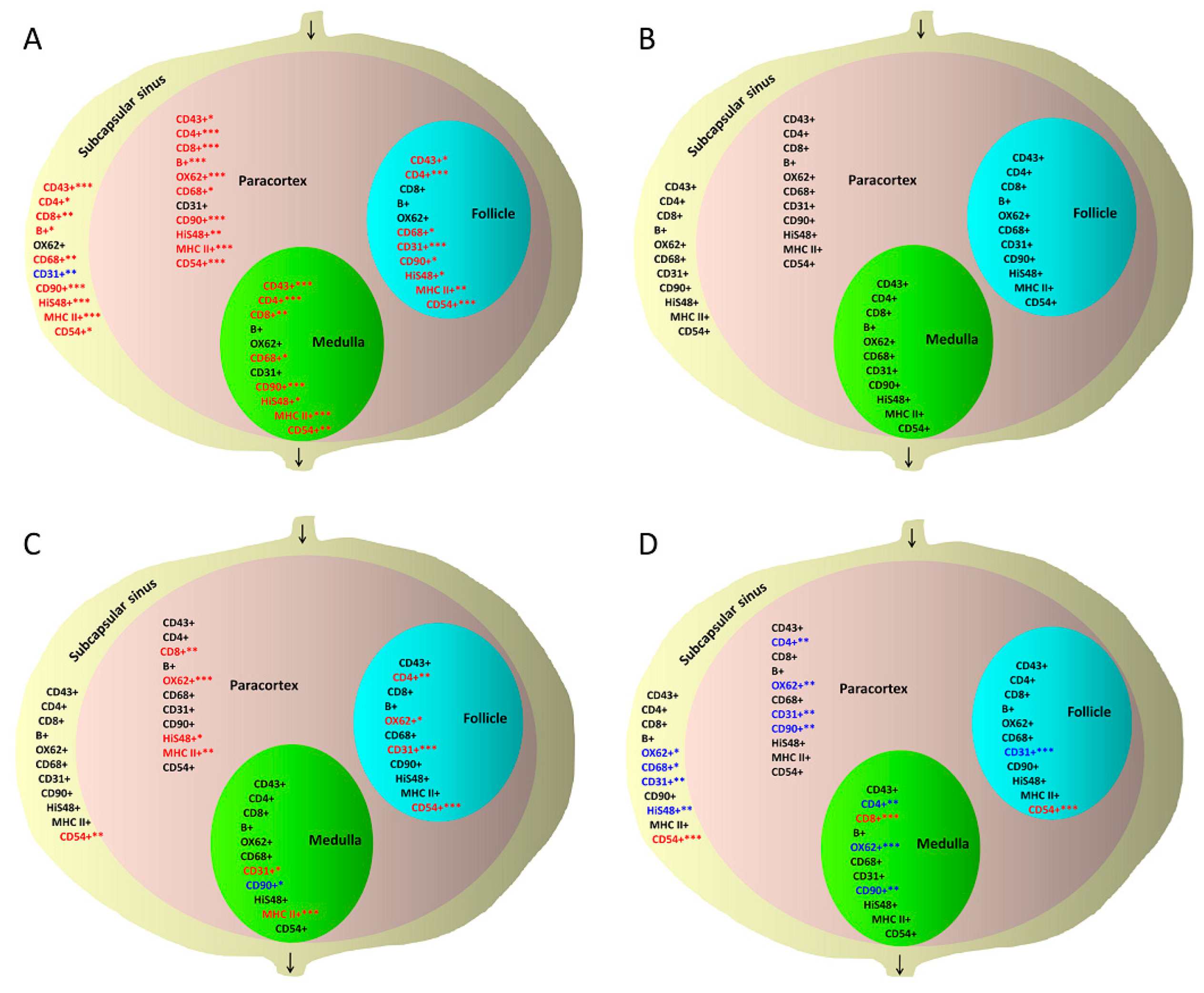
3.2. Quantitative Changes in Cell Subpopulations in Lymph Node Zones after S. epidermidis Infection
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Park, Y.J.; Kim, C.W.; Lee, H.K. Interactions between host immunity and skin-colonizing Staphylococci: No two siblings are alike. Int. J. Mol. Sci. 2019, 20, 718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olszewski, W.L.; Jamal, S.; Manokaran, G.; Pani, S.; Kumaraswami, V.; Kubicka, U.; Lukomska, B.; Tripathi, F.M.; Swoboda, E.; Meisel-Mikolajczyk, F.; et al. Bacteriological studies of blood, tissue fluid, lymph and lymph nodes in patients with acute dermatolymphangioadenitis (DLA) in course of ‘filarial’ lymphedema. Acta Trop. 1999, 73, 217–224. [Google Scholar] [CrossRef] [Green Version]
- Cogen, A.L.; Yamasaki, K.; Sanchez, K.M.; Dorschner, R.A.; Lai, Y.; MacLeod, D.T.; Torpey, J.W.; Otto, M.; Nizet, V.; Kim, J.E.; et al. Selective antimicrobial action is provided by phenol-soluble modulins derived from Staphylococcus epidermidis, a normal resident of the skin. J. Investig. Dermatol. 2010, 130, 192–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; MacLeod, D.T.; Di Nardo, A. Commensal bacteria lipoteichoic acid increases skin mast cell antimicrobial activity against vaccinia viruses. J. Immunol. 2012, 189, 1551–1558. [Google Scholar] [CrossRef] [Green Version]
- Lai, Y.; Di Nardo, A.; Nakatsuji, T.; Leichtle, A.; Yang, Y.; Cogen, A.L.; Wu, Z.R.; Hooper, L.V.; Schmidt, R.R.; von Aulock, S.; et al. Commensal bacteria regulate Toll-like receptor 3-dependent inflammation after skin injury. Nat. Med. 2009, 15, 1377–1382. [Google Scholar] [CrossRef]
- Gallo, R.L.; Nakatsuji, T. Microbial symbiosis with the innate immune defense system of the skin. J. Investig. Dermatol. 2011, 131, 1974–1980. [Google Scholar] [CrossRef] [Green Version]
- Grice, E.A.; Segre, J.A. The skin microbiome. Nat. Rev. Microbiol. 2011, 9, 244–253. [Google Scholar] [CrossRef]
- Swaney, M.H.; Kalan, L.R. Living in your skin: Microbes, molecules, and mechanisms. Infect. Immun. 2021, 89, e00695-20. [Google Scholar] [CrossRef]
- Scharschmidt, T.C. Establishing tolerance to commensal skin bacteria: Timing is everything. Dermatol. Clin. 2017, 35, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Scharschmidt, T.C.; Vasquez, K.S.; Truong, H.A.; Gearty, S.V.; Pauli, M.L.; Nosbaum, A.; Gratz, I.K.; Otto, M.; Moon, J.J.; Liese, J.; et al. A wave of regulatory T cells into neonatal skin mediates tolerance to commensal microbes. Immunity 2015, 43, 1011–1021. [Google Scholar] [CrossRef] [Green Version]
- Shen, W.; Li, W.; Hixon, J.A.; Bouladoux, N.; Belkaid, Y.; Dzutzev, A.; Durum, S.K. Adaptive immunity to murine skin commensals. Proc. Natl. Acad. Sci. USA 2014, 111, E2977–E2986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naik, S.; Bouladoux, N.; Linehan, J.L.; Han, S.J.; Harrison, O.J.; Wilhelm, C.; Conlan, S.; Himmelfarb, S.; Byrd, A.L.; Deming, C.; et al. Commensal-dendritic-cell interaction specifies a unique protective skin immune signature. Nature 2015, 520, 104–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linehan, J.L.; Harrison, O.J.; Han, S.J.; Byrd, A.L.; Vujkovic-Cvijin, I.; Villarino, A.V.; Sen, S.K.; Shaik, J.; Smelkinson, M.; Tamoutounour, S.; et al. Non-classical immunity controls microbiota impact on skin immunity and tissue repair. Cell 2018, 172, 784–796.e18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szczesny, G.; Olszewski, W.L.; Zaleska, M. Limb lymph node response to bone fracture. Lymphat. Res. Biol. 2004, 2, 155–164. [Google Scholar] [CrossRef]
- Olszewski, W.L.; Jain, P.; Zaleska, M.; Stelmach, E.; Swoboda, E. Chronic lower limb wounds evoke systemic response of the lymphatic (immune) system. Indian J. Plast. Surg. 2012, 45, 255–260. [Google Scholar] [CrossRef]
- Masamatti, S.S.; Narasimha, A.; Janardhan, J.V.; Chowdappa, V. Lymph node fibrosis in a case of primary lymphoedema—A report of two cases. J. Clin. Diagn. Res. 2016, 10, ED08–ED09. [Google Scholar] [CrossRef]
- Olszewski, W.L.; Zagozda, M.; Zaleska, M.T.; Durlik, M. Predilection to dermato-lymphangio-adenitis in obstructive lymphedema of lower limbs depending on genetic polymorphisms at TNFα-308G>A, CCR2-190G>A, CD14-159C>T, TLR2 2029C>T, TLR4 1063A>G, TLR4 1363C>T, TGFβ 74G>C, and TGFβ 29T>C. Lymphat. Res. Biol. 2018, 16, 109–116. [Google Scholar] [CrossRef]
- Gunderson, C.G. Cellulitis: Definition, etiology, and clinical features. Am. J. Med. 2011, 124, 1113–1122. [Google Scholar] [CrossRef]
- Mortimer, P.S.; Rockson, S.G. New developments in clinical aspects of lymphatic disease. J. Clin. Investig. 2014, 124, 915–921. [Google Scholar] [CrossRef]
- Olszewski, W.L.; Zaleska, M.T. Diagnosis and management of infection in lymphedema. In Lymphedema; Lee, B.B., Rockson, S., Bergan, J., Eds.; Springer: Cham, Switzerland, 2018; pp. 465–480. [Google Scholar] [CrossRef]
- Wireko, S.; Asiedu, S.O.; Kini, P.; Aglomasa, B.C.; Amewu, E.K.A.; Asiedu, E.; Osei-Akoto, F.; Boahen, K.G.; Obiri-Yeboah, D.; Amato, K.R.; et al. Prevalence of methicillin-resistant Staphylococcus species among filarial lymphedema patients in Ahanta West District of Ghana. Front. Tropical Dis. 2021, 2, 41. [Google Scholar] [CrossRef]
- Hara, H.; Mihara, M. Bacterial flora in the genital area of patients with lower limb lymphedema. Lymphat. Res. Biol. 2020, 18, 31–34. [Google Scholar] [CrossRef] [PubMed]
- Hon, K.L.; Tsang, Y.C.; Pong, N.H.; Leung, T.F.; Ip, M. Exploring Staphylococcus epidermidis in atopic eczema: Friend or foe? Clin. Exp. Dermatol. 2016, 41, 659–663. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Speer, C.P.; Glaser, K. Beyond sepsis: Staphylococcus epidermidis is an underestimated but significant contributor to neonatal morbidity. Virulence 2018, 9, 621–633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cąkała, M.; Olszewski, W.L. The response of the lymphatic system to the human skin resident bacteria. Ann. Transplant. 2002, 7, 30–35. [Google Scholar]
- Netea, M.G.; Schlitzer, A.; Placek, K.; Joosten, L.A.B.; Schultze, J.L. Innate and adaptive immune memory: An evolutionary continuum in the host’s response to pathogens. Cell Host Microbe. 2019, 25, 13–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbas, K.B.; Lichtman, A.H.; Pillai, S. Properties and overview of immune responses. Innate and adaptive immunity. In Cellular and Molecular Immunology, 10th ed.; South Asia Edition-E-Book; Elsevier Health Sciences: New Delhi, India, 2021; pp. 1–11. [Google Scholar]
- SAS Institute Inc. SAS/STAT® 15.1 User’s Guide; SAS Institute Inc.: Cary, NC, USA, 2018. [Google Scholar]
- Cakala, M.; Olszewski, W.L. The reaction of the regional lymph nodes on the skin bacterial and allogeneic antigens. Ann. Transplant. 2004, 9, 59–62. [Google Scholar]
- Desvars-Larrive, A.; Ruppitsch, W.; Lepuschitz, S.; Szostak, M.P.; Spergser, J.; Feßler, A.T.; Schwarz, S.; Monecke, S.; Ehricht, R.; Walzer, C.; et al. Urban brown rats (Rattus norvegicus) as possible source of multidrug-resistant Enterobacteriaceae and meticillin-resistant Staphylococcus spp., Vienna, Austria, 2016 and 2017. Euro Surveill. 2019, 24, 1900149. [Google Scholar] [CrossRef] [Green Version]
- Banchereau, J.; Briere, F.; Caux, C.; Davoust, J.; Lebecque, S.; Liu, Y.J.; Pulendran, B.; Palucka, K. Immunobiology of dendritic cells. Annu. Rev. Immunol. 2000, 18, 767–811. [Google Scholar] [CrossRef]
- Britschgi, M.R.; Favre, S.; Luther, S.A. CCL21 is sufficient to mediate DC migration, maturation and function in the absence of CCL19. Eur. J. Immunol. 2010, 40, 1266–1271. [Google Scholar] [CrossRef]
- Teijeira, A.; Russo, E.; Halin, C. Taking the lymphatic route: Dendritic cell migration to draining lymph nodes. Semin. Immunopathol. 2014, 36, 261–274. [Google Scholar] [CrossRef]
- Luther, S.; Tang, H.L.; Hyman, P.L.; Farr, A.G.; Cyster, J.G. Coexpression of the chemokines ELC and SLC by T zone stromal cells and deletion of the ELC gene in the plt/plt mouse. Proc. Natl. Acad. Sci. USA 2000, 97, 12694–12699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaiser, A.; Donnadieu, E.; Abastado, J.P.; Trautmann, A.; Nardin, A. CC chemokine ligand 19 secreted by mature dendritic cells increases naive T cell scanning behavior and their response to rare cognate antigen. J. Immunol. 2005, 175, 2349–2356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egawa, G.; Roediger, B.; Tay, S.S.; Cavanagh, L.; Guy, T.V.; de Fazekas, B.; Brzoska, A.J.; Firth, N.; Weninger, W. Bacterial antigen is directly delivered to the draining lymph nodes and activates CD8+ T cells during Staphylococcus aureus skin infection. Immunol. Cell Biol. 2021, 99, 299–308. [Google Scholar] [CrossRef] [PubMed]
- Pepper, M.; Jenkins, M.K. Origins of CD4+ effector and central memory T cells. Nat. Immunol. 2011, 12, 467–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krawczyk, C.M.; Shen, H.; Pearce, E.J. Functional plasticity in memory T helper cell responses. J. Immunol. 2007, 178, 4080–4088. [Google Scholar] [CrossRef] [Green Version]
- Bell, E.B.; Westermann, J. CD4 memory T cells on trial: Immunological memory without a memory T cell. Trends Immunol. 2008, 29, 405–411. [Google Scholar] [CrossRef]
- MacLeod, M.K.; Clambey, E.T.; Kappler, J.W.; Marrack, P. CD4 memory T cells: What are they and what can they do? Semin. Immunol. 2009, 21, 53–61. [Google Scholar] [CrossRef] [Green Version]
- Janssen, E.M.; Lemmens, E.E.; Wolfe, T.; Christen, U.; von Herrath, M.G.; Schoenberger, S.P. CD4+ T cells are required for secondary expansion and memory in CD8+ T lymphocytes. Nature 2003, 421, 852–856. [Google Scholar] [CrossRef]
- Ariotti, S.; Hogenbirk, M.A.; Dijkgraaf, F.E.; Visser, L.L.; Hoekstra, M.E.; Song, J.Y.; Jacobs, H.; Haanen, J.B.; Schumacher, T.N. T cell memory. Skin-resident memory CD8+ T cells trigger a state of tissue-wide pathogen alert. Science 2014, 346, 101–105. [Google Scholar] [CrossRef]
- Chen, M.; Wang, Y.H.; Wang, Y.; Huang, L.; Sandoval, H.; Liu, Y.J.; Wang, J. Dendritic cell apoptosis in the maintenance of immune tolerance. Science 2006, 311, 1160–1164. [Google Scholar] [CrossRef]
- Guarda, G.; Hons, M.; Soriano, S.F.; Huang, A.Y.; Polley, R.; Martín-Fontecha, A.; Stein, J.V.; Germain, R.N.; Lanzavecchia, A.; Sallusto, F. L-selectin-negative CCR7- effector and memory CD8+ T cells enter reactive lymph nodes and kill dendritic cells. Nat. Immunol. 2007, 8, 743–752. [Google Scholar] [CrossRef] [PubMed]
- Björck, P.; Axelsson, B.; Paulie, S. Expression of CD40 and CD43 during activation of human B lymphocytes. Scand. J. Immunol. 1991, 33, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Inui, M.; Hirota, S.; Hirano, K.; Fujii, H.; Sugahara-Tobinai, A.; Ishii, T.; Harigae, H.; Takai, T. Human CD43+ B cells are closely related not only to memory B cells phenotypically but also to plasmablasts developmentally in healthy individuals. Int. Immunol. 2015, 27, 345–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clavarino, G.; Delouche, N.; Vettier, C.; Laurin, D.; Pernollet, M.; Raskovalova, T.; Cesbron, J.Y.; Dumestre-Pérard, C.; Jacob, M.C. Novel strategy for phenotypic characterization of human B lymphocytes from precursors to effector cells by flow cytometry. PLoS ONE. 2016, 11, e0162209. [Google Scholar] [CrossRef] [PubMed]
- Raman, V.S.; Akondy, R.S.; Rath, S.; Bal, V.; George, A. Ligation of CD27 on B cells in vivo during primary immunization enhances commitment to memory B cell responses. J. Immunol. 2003, 171, 5876–5881. [Google Scholar] [CrossRef] [Green Version]
- Sallusto, F.; Lanzavecchia, A.; Araki, K.; Ahmed, R. From vaccines to memory and back. Immunity 2010, 33, 451–463. [Google Scholar] [CrossRef] [Green Version]
- Westermann, J.; Geismar, U.; Sponholz, A.; Bode, U.; Sparshott, S.M.; Bell, E.B. CD4+ T cells of both the naive and the memory phenotype enter rat lymph nodes and Peyer’s patches via high endothelial venules: Within the tissue their migratory behavior differs. Eur. J. Immunol. 1997, 27, 3174–3181. [Google Scholar] [CrossRef]
- Klinger, A.; Gebert, A.; Bieber, K.; Kalies, K.; Ager, A.; Bell, E.B.; Westermann, J. Cyclical expression of L-selectin (CD62L) by recirculating T cells. Int. Immunol. 2009, 21, 443–455. [Google Scholar] [CrossRef] [Green Version]
- van der Aar, A.M.; Picavet, D.I.; Muller, F.J.; de Boer, L.; van Capel, T.M.; Zaat, S.A.; Bos, J.D.; Janssen, H.; George, T.C.; Kapsenberg, M.L.; et al. Langerhans cells favor skin flora tolerance through limited presentation of bacterial antigens and induction of regulatory T cells. J. Investig. Dermatol. 2013, 133, 1240–1249. [Google Scholar] [CrossRef] [Green Version]
- Poirot, J.; Medvedovic, J.; Trichot, C.; Soumelis, V. Compartmentalized multicellular crosstalk in lymph nodes coordinates the generation of potent cellular and humoral immune responses. Eur. J. Immunol. 2021, 51, 3146–3160. [Google Scholar] [CrossRef]
- Cyster, J.G. B cell follicles and antigen encounters of the third kind. Nat. Immunol. 2010, 11, 989–996. [Google Scholar] [CrossRef] [PubMed]
- Moran, I.; Grootveld, A.K.; Nguyen, A.; Phan, T.G. Subcapsular sinus macrophages: The seat of innate and adaptive memory in murine lymph nodes. Trends Immunol. 2019, 40, 35–48. [Google Scholar] [CrossRef] [PubMed]
- Gatto, D.; Brink, R. The germinal center reaction. J. Allergy Clin. Immunol. 2010, 126, 898–907. [Google Scholar] [CrossRef]
- Card, C.M.; Yu, S.S.; Swartz, M.A. Emerging roles of lymphatic endothelium in regulating adaptive immunity. J. Clin. Investig. 2014, 124, 943–952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamenyeva, O.; Boularan, C.; Kabat, J.; Cheung, G.Y.; Cicala, C.; Yeh, A.J.; Chan, J.L.; Periasamy, S.; Otto, M.; Kehrl, J.H. Neutrophil recruitment to lymph nodes limits local humoral response to Staphylococcus aureus. PLoS Pathog. 2015, 11, e1004827. [Google Scholar] [CrossRef]
- Geng, S.; Matsushima, H.; Okamoto, T.; Yao, Y.; Lu, R.; Page, K.; Blumenthal, R.M.; Ward, N.L.; Miyazaki, T.; Takashima, A. Emergence, origin, and function of neutrophil-dendritic cell hybrids in experimentally induced inflammatory lesions in mice. Blood 2013, 121, 1690–1700. [Google Scholar] [CrossRef] [Green Version]
- Matsushima, H.; Geng, S.; Lu, R.; Okamoto, T.; Yao, Y.; Mayuzumi, N.; Kotol, P.F.; Chojnacki, B.J.; Miyazaki, T.; Gallo, R.L.; et al. Neutrophil differentiation into a unique hybrid population exhibiting dual phenotype and functionality of neutrophils and dendritic cells. Blood 2013, 121, 1677–1689. [Google Scholar] [CrossRef] [Green Version]
- Takashima, A.; Yao, Y. Neutrophil plasticity: Acquisition of phenotype and functionality of antigen-presenting cell. J. Leukoc. Biol. 2015, 98, 489–496. [Google Scholar] [CrossRef] [Green Version]
- Hampton, H.R.; Chtanova, T. The lymph node neutrophil. Semin. Immunol. 2016, 28, 129–136. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Bhanja, A.; Bhattacharyya, J.; Jaganathan, B.G. Multiple roles of CD90 in cancer. Tumour Biol. 2016, 37, 11611–11622. [Google Scholar] [CrossRef]
- Agematsu, K.; Hokibara, S.; Nagumo, H.; Komiyama, A. CD27: A memory B-cell marker. Immunol. Today. 2000, 21, 204–206. [Google Scholar] [CrossRef]
- Vugmeyster, Y.; Howell, K.; Bakshi, A.; Flores, C.; Hwang, O.; McKeever, K. B-cell subsets in blood and lymphoid organs in Macaca fascicularis. Cytometry A. 2004, 61, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Byrd, A.L.; Belkaid, Y.; Segre, J.A. The human skin microbiome. Nat. Rev. Microbiol. 2018, 16, 143–155. [Google Scholar] [CrossRef] [PubMed]
- Galkowska, H.; Podbielska, A.; Olszewski, W.L.; Stelmach, E.; Luczak, M.; Rosinski, G.; Karnafel, W. Epidemiology and prevalence of methicillin-resistant Staphylococcus aureus and Staphylococcus epidermidis in patients with diabetic foot ulcers: Focus on the differences between species isolated from individuals with ischemic vs. neuropathic foot ulcers. Diabetes Res. Clin. Pract. 2009, 84, 187–193. [Google Scholar] [CrossRef]
- Olszewski, W.L. Infection. In Lymphedema: A Concise Compedium of Theory and Practice; Lee, B.B., Bergan, J., Rockson, S.G., Eds.; Springer: Cham, Switzerland, 2011; Part VI, 25; pp. 207–218. [Google Scholar] [CrossRef]
- Andziak, P.; Olszewski, W.L.; Moscicka-Wesolowska, M.; Interewicz, B.; Swoboda, E.; Wastelmach, E. Skin own bacteria may aggravate inflammatory and occlusive changes in atherosclerotic arteries of lower limbs. Int. Angiol. 2012, 31, 474–482. [Google Scholar] [PubMed]
- Vu, B.G.; Stach, C.S.; Salgado-Pabón, W.; Diekema, D.J.; Gardner, S.E.; Schlievert, P.M. Superantigens of Staphylococcus aureus from patients with diabetic foot ulcers. J. Infect. Dis. 2014, 210, 1920–1927. [Google Scholar] [CrossRef] [Green Version]
- Toniolo, A.; Cassani, G.; Puggioni, A.; Rossi, A.; Colombo, A.; Onodera, T.; Ferrannini, E. The diabetes pandemic and associated infections: Suggestions for clinical microbiology. Rev. Med. Microbiol. 2019, 30, 1–17. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Type | Antigen (Rat Clone) |
|---|---|
| T lymphocytes | CD43+ (W3/13+) |
| T helper lymphocytes, monocytes | CD4+ (W3/25+) |
| T cytotoxic lymphocytes | CD8+ (OX8+) |
| B lymphocytes | B+ |
| B lymphocytes | OX12+ |
| B lymphocytes | CD45RA+ (OX33+) |
| Dendritic cells | OX62+ |
| Macrophages, monocytes | CD68+ (ED1+) |
| PECAM-1, endothelial cells | CD31+ |
| Stem cells, thymocytes, immature B lymphocytes | CD90+ (OX7+) |
| Granulocytes | HiS48+ |
| Activated antigen-presenting cells | MHC class II+ (OX6+) |
| ICAM-1, dendritic cells, endothelial cells | CD54+ |
| Activated T lymphocytes | CD43+ MHC class II+ |
| Activated helper T lymphocytes | CD4+ MHC class II+ |
| Activated cytotoxic T lymphocytes | CD8+ MHC class II+ |
| T regulatory cells (Treg) | CD4+ CD25+ |
| Memory B cells, plasmablasts | CD19+ CD27+ |
| Memory T helper lymphocytes | CD4+ CD45RC- |
| Central memory T helper lymphocytes | CD4+ CD45RC- CD62L+ |
| Effector memory T helper lymphocytes | CD4+ CD45RC- CD62L- |
| Memory T cytotoxic lymphocytes | CD8+ CD45RC- |
| Central memory T cytotoxic lymphocytes | CD8+ CD45RC- CD62L+ |
| Effector memory T cytotoxic lymphocytes | CD8+ CD45RC- CD62L- |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cąkała-Jakimowicz, M.; Puzianowska-Kuznicka, M. Towards Understanding the Lymph Node Response to Skin Infection with Saprophytic Staphylococcus epidermidis. Biomedicines 2022, 10, 1021. https://doi.org/10.3390/biomedicines10051021
Cąkała-Jakimowicz M, Puzianowska-Kuznicka M. Towards Understanding the Lymph Node Response to Skin Infection with Saprophytic Staphylococcus epidermidis. Biomedicines. 2022; 10(5):1021. https://doi.org/10.3390/biomedicines10051021
Chicago/Turabian StyleCąkała-Jakimowicz, Marta, and Monika Puzianowska-Kuznicka. 2022. "Towards Understanding the Lymph Node Response to Skin Infection with Saprophytic Staphylococcus epidermidis" Biomedicines 10, no. 5: 1021. https://doi.org/10.3390/biomedicines10051021
APA StyleCąkała-Jakimowicz, M., & Puzianowska-Kuznicka, M. (2022). Towards Understanding the Lymph Node Response to Skin Infection with Saprophytic Staphylococcus epidermidis. Biomedicines, 10(5), 1021. https://doi.org/10.3390/biomedicines10051021