
Metabolic and Molecular Mechanisms of Diet and Physical Exercise in the Management of Polycystic Ovarian Syndrome

 ,
,  ,
,  ,
,  ,
,  ,
,
Abstract
:1. Polycystic Ovary Syndrome: Definition and Pathogenic Mechanisms
- -
- oligo/anovulation + hyperandrogenism + polycystic ovaries;
- -
- oligo/anovulation + hyperandrogenism;
- -
- hyperandrogenism + polycystic ovaries;
- -
- oligo/anovulation + polycystic ovaries [8].
2. Mechanisms Involved in PCOS Pathogenesis
2.1. Molecular Mechanisms
2.2. Gene Polymorphisms
2.3. Epigenetic Mechanisms
2.3.1. DNA Methylation
2.3.2. MicroRNA
2.3.3. Chromatin Modifications
2.4. Immune Mechanisms
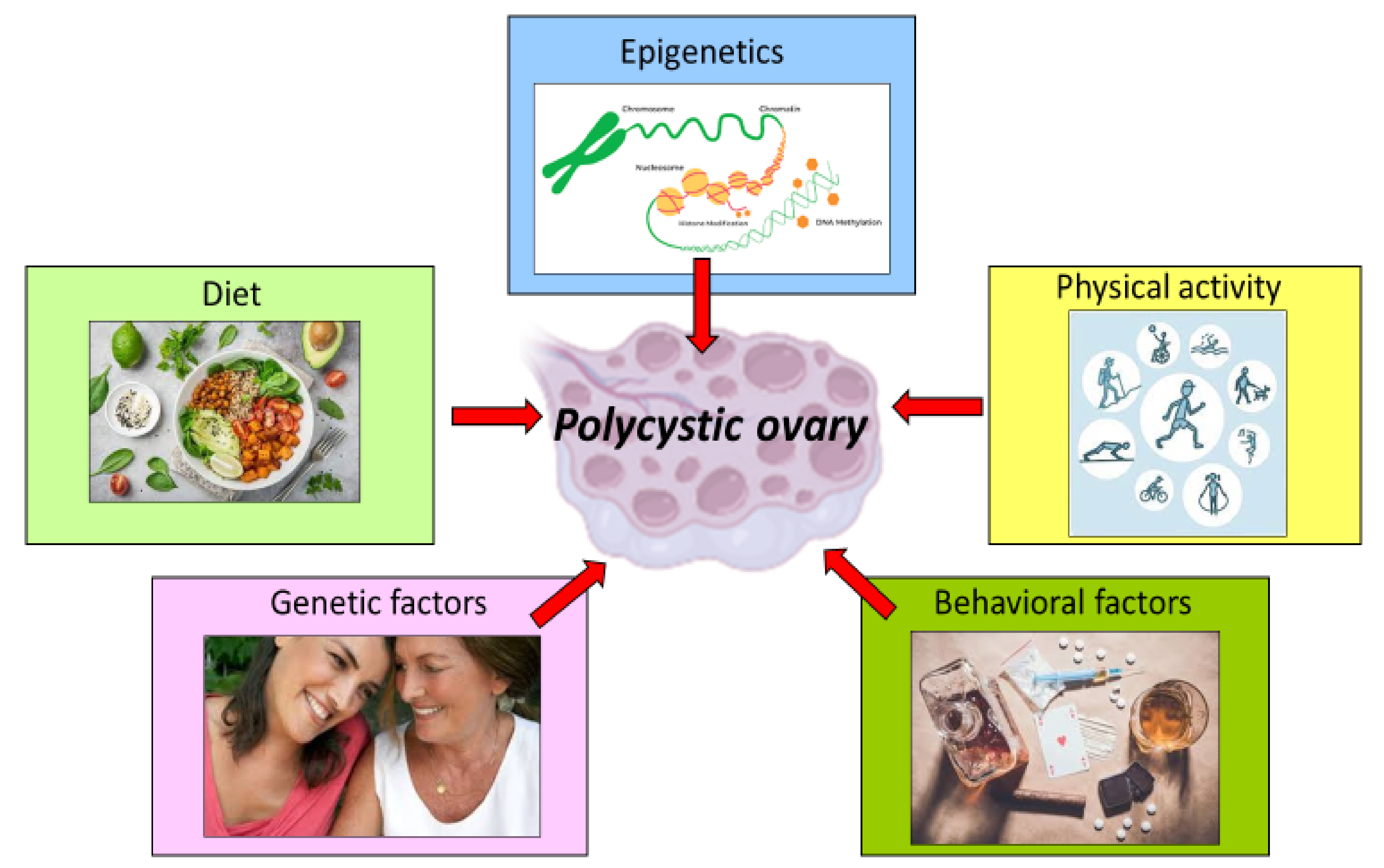
3. Impact of Lifestyle and Behavior in the Management of PCOS
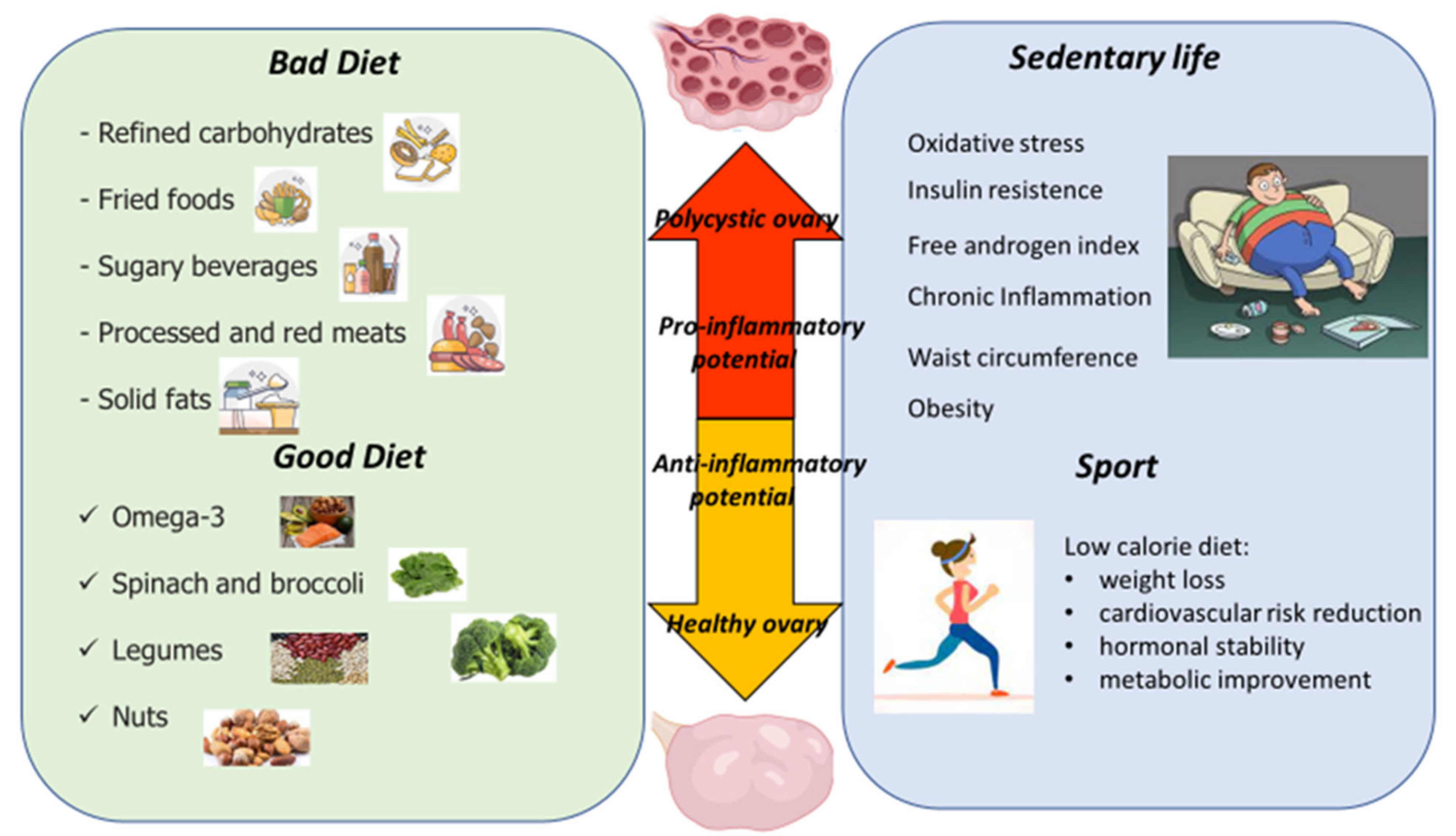
4. Dietary Modification
5. Exercise Interventions in PCOS Women
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Laven, J.S.E. Follicle Stimulating Hormone Receptor (FSHR) Polymorphisms and Polycystic Ovary Syndrome (PCOS). Front. Endocrinol. 2019, 10, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, S. Polycystic Ovary Syndrome (PCOS), an Inflammatory, Systemic, Lifestyle Endocrinopathy. J. Steroid Biochem. Mol. Biol. 2018, 182, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Witchel, S.F.; Oberfield, S.E.; Peña, A.S. Polycystic Ovary Syndrome: Pathophysiology, Presentation, and Treatment With Emphasis on Adolescent Girls. J. Endocr. Soc. 2019, 3, 1545–1573. [Google Scholar] [CrossRef] [PubMed]
- Wojciechowska, A.; Osowski, A.; Jóźwik, M.; Górecki, R.; Rynkiewicz, A.; Wojtkiewicz, J. Inositols’ Importance in the Improvement of the Endocrine–Metabolic Profile in PCOS. IJMS 2019, 20, 5787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ajmal, N.; Khan, S.Z.; Shaikh, R. Polycystic Ovary Syndrome (PCOS) and Genetic Predisposition: A Review Article. Eur. J. Obstet. Gynecol. Reprod. Biol. X 2019, 3, 100060. [Google Scholar] [CrossRef] [PubMed]
- Costello, M.F.; Misso, M.L.; Balen, A.; Boyle, J.; Devoto, L.; Garad, R.M.; Hart, R.; Johnson, L.; Jordan, C.; Legro, R.S.; et al. A Brief Update on the Evidence Supporting the Treatment of Infertility in Polycystic Ovary Syndrome. Aust. N. Z. J. Obstet. Gynaecol. 2019, 59, 867–873. [Google Scholar] [CrossRef]
- Hart, R. PCOS and Infertility. Panminerva Med. 2008, 50, 305–314. [Google Scholar]
- Revised 2003 Consensus on Diagnostic Criteria and Long-Term Health Risks Related to Polycystic Ovary Syndrome. Fertil. Steril. 2004, 81, 19–25. [CrossRef]
- Bednarska, S.; Siejka, A. The Pathogenesis and Treatment of Polycystic Ovary Syndrome: What’s New? Adv. Clin. Exp. Med. 2017, 26, 359–367. [Google Scholar] [CrossRef] [Green Version]
- Ganie, M.; Vasudevan, V.; Wani, I.; Baba, M.; Arif, T.; Rashid, A. Epidemiology, Pathogenesis, Genetics & Management of Polycystic Ovary Syndrome in India. Indian J. Med. Res. 2019, 150, 333. [Google Scholar] [CrossRef]
- Ruiz-Manriquez, L.M.; Ledesma Pacheco, S.J.; Medina-Gomez, D.; Uriostegui-Pena, A.G.; Estrada-Meza, C.; Bandyopadhyay, A.; Pathak, S.; Banerjee, A.; Chakraborty, S.; Srivastava, A.; et al. A Brief Review on the Regulatory Roles of MicroRNAs in Cystic Diseases and Their Use as Potential Biomarkers. Genes 2022, 13, 191. [Google Scholar] [CrossRef] [PubMed]
- Cai, G.; Ma, X.; Chen, B.; Huang, Y.; Liu, S.; Yang, H.; Zou, W. MicroRNA-145 Negatively Regulates Cell Proliferation Through Targeting IRS1 in Isolated Ovarian Granulosa Cells From Patients With Polycystic Ovary Syndrome. Reprod. Sci. 2017, 24, 902–910. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Huang, J.; Li, L.; Chen, Y.; Chen, X.; Zhao, X.; Yang, D. MicroRNA-93 Promotes Ovarian Granulosa Cells Proliferation through Targeting CDKN1A in Polycystic Ovarian Syndrome. J. Clin. Endocrinol. Metab. 2015, 100, E729–E738. [Google Scholar] [CrossRef] [Green Version]
- Yi, S.; Zheng, B.; Zhu, Y.; Cai, Y.; Sun, H.; Zhou, J. Melatonin Ameliorates Excessive PINK1/Parkin-Mediated Mitophagy by Enhancing SIRT1 Expression in Granulosa Cells of PCOS. Am. J. Physiol. Endocrinol. Metab. 2020, 319, E91–E101. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Luo, M.; Wang, R.; Ye, J.; Wang, X. Mitochondria in Sex Hormone-Induced Disorder of Energy Metabolism in Males and Females. Front. Endocrinol. 2021, 12, 749451. [Google Scholar] [CrossRef]
- Ye, J. Mechanism of Insulin Resistance in Obesity: A Role of ATP. Front. Med. 2021, 15, 372–382. [Google Scholar] [CrossRef]
- Wu, S.; Divall, S.; Nwaopara, A.; Radovick, S.; Wondisford, F.; Ko, C.; Wolfe, A. Obesity-Induced Infertility and Hyperandrogenism Are Corrected by Deletion of the Insulin Receptor in the Ovarian Theca Cell. Diabetes 2014, 63, 1270–1282. [Google Scholar] [CrossRef] [Green Version]
- Van Leckwyck, M.; Kong, W.; Burton, K.J.; Amati, F.; Vionnet, N.; Pralong, F.P. Decreasing Insulin Sensitivity in Women Induces Alterations in LH Pulsatility. J. Clin. Endocrinol. Metab. 2016, 101, 3240–3249. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Wang, H.; Liu, W.; Zhang, Z.; Zhang, Y.; Zhang, W.; Chen, Z.; Xia, G.; Wang, C. High Level of C-Type Natriuretic Peptide Induced by Hyperandrogen-Mediated Anovulation in Polycystic Ovary Syndrome Mice. Clin. Sci. 2018, 132, 759–776. [Google Scholar] [CrossRef] [Green Version]
- Azhary, J.M.K.; Harada, M.; Kunitomi, C.; Kusamoto, A.; Takahashi, N.; Nose, E.; Oi, N.; Wada-Hiraike, O.; Urata, Y.; Hirata, T.; et al. Androgens Increase Accumulation of Advanced Glycation End Products in Granulosa Cells by Activating ER Stress in PCOS. Endocrinology 2020, 161, bqaa015. [Google Scholar] [CrossRef]
- Diamanti-Kandarakis, E.; Chatzigeorgiou, A.; Papageorgiou, E.; Koundouras, D.; Koutsilieris, M. Advanced Glycation End-Products and Insulin Signaling in Granulosa Cells. Exp. Biol. Med. 2016, 241, 1438–1445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, M.; Du, J.; Lu, D.; Ren, C.; Shen, H.; Qiao, J.; Chen, X.; Zhang, H. Increased Expression of Kindlin 2 in Luteinized Granulosa Cells Correlates with Androgen Receptor Level in Patients with Polycystic Ovary Syndrome Having Hyperandrogenemia. Reprod. Sci. 2014, 21, 696–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, M.; Zhang, Y.; Li, X.; Cui, P.; Li, J.; Brännström, M.; Shao, L.R.; Billig, H. Alterations of Endometrial Epithelial-Mesenchymal Transition and MAPK Signalling Components in Women with PCOS Are Partially Modulated by Metformin in Vitro. Mol. Hum. Reprod. 2020, 26, 312–326. [Google Scholar] [CrossRef] [PubMed]
- Fisch, S.C.; Nikou, A.F.; Wright, E.A.; Phan, J.D.; Leung, K.L.; Grogan, T.R.; Abbott, D.H.; Chazenbalk, G.D.; Dumesic, D.A. Precocious Subcutaneous Abdominal Stem Cell Development to Adipocytes in Normal-Weight Women with Polycystic Ovary Syndrome. Fertil. Steril. 2018, 110, 1367–1376. [Google Scholar] [CrossRef]
- Shaaban, Z.; Khoradmehr, A.; Amiri-Yekta, A.; Nowzari, F.; Jafarzadeh Shirazi, M.R.; Tamadon, A. Pathophysiologic Mechanisms of Insulin Secretion and Signaling-Related Genes in Etiology of Polycystic Ovary Syndrome. Genet. Res. 2021, 2021, 1–13. [Google Scholar] [CrossRef]
- Wawrzkiewicz-Jałowiecka, A.; Kowalczyk, K.; Trybek, P.; Jarosz, T.; Radosz, P.; Setlak, M.; Madej, P. In Search of New Therapeutics-Molecular Aspects of the PCOS Pathophysiology: Genetics, Hormones, Metabolism and Beyond. Int. J. Mol. Sci. 2020, 21, E7054. [Google Scholar] [CrossRef]
- Urbanek, M.; Woodroffe, A.; Ewens, K.G.; Diamanti-Kandarakis, E.; Legro, R.S.; Strauss, J.F.; Dunaif, A.; Spielman, R.S. Candidate Gene Region for Polycystic Ovary Syndrome on Chromosome 19p13.2. J. Clin. Endocrinol. Metab. 2005, 90, 6623–6629. [Google Scholar] [CrossRef] [Green Version]
- Anastasia, K.; Koika, V.; Roupas, N.D.; Armeni, A.; Marioli, D.; Panidis, D.; George, A.; Georgopoulos, N.A. Association of Calpain (CAPN) 10 (UCSNP-43, Rs3792267) Gene Polymorphism with Elevated Serum Androgens in Young Women with the Most Severe Phenotype of Polycystic Ovary Syndrome (PCOS). Gynecol. Endocrinol. 2015, 31, 630–634. [Google Scholar] [CrossRef]
- Dilek, S.; Ertunc, D.; Tok, E.C.; Erdal, E.M.; Aktas, A. Association of Gly972Arg Variant of Insulin Receptor Substrate-1 with Metabolic Features in Women with Polycystic Ovary Syndrome. Fertil. Steril. 2005, 84, 407–412. [Google Scholar] [CrossRef]
- Hatfield, J.; Kasliwal, A. Response to “The FSRH Guideline on Conscientious Objection Disrespects Patient Rights and Endangers Their Health”. BMJ Sex. Reprod Health 2018, 44, 146. [Google Scholar] [CrossRef] [Green Version]
- Chambers, A.E.; Griffin, C.; Naif, S.A.; Mills, I.; Mills, W.E.; Syngelaki, A.; Nicolaides, K.H.; Banerjee, S. Quantitative ELISAs for Serum Soluble LHCGR and HCG-LHCGR Complex: Potential Diagnostics in First Trimester Pregnancy Screening for Stillbirth, Down’s Syndrome, Preterm Delivery and Preeclampsia. Reprod. Biol. Endocrinol. 2012, 10, 113. [Google Scholar] [CrossRef] [Green Version]
- Zheng, M.-X.; Li, Y.; Hu, R.; Wang, F.-M.; Zhang, X.-M.; Guan, B. Anti-Müllerian Hormone Gene Polymorphism Is Associated with Androgen Levels in Chinese Polycystic Ovary Syndrome Patients with Insulin Resistance. J. Assist. Reprod. Genet. 2016, 33, 199–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Witchel, S.F.; Kahsar-Miller, M.; Aston, C.E.; White, C.; Azziz, R. Prevalence of CYP21 Mutations and IRS1 Variant among Women with Polycystic Ovary Syndrome and Adrenal Androgen Excess. Fertil. Steril. 2005, 83, 371–375. [Google Scholar] [CrossRef] [PubMed]
- Diamanti-Kandarakis, E.; Bartzis, M.I.; Bergiele, A.T.; Tsianateli, T.C.; Kouli, C.R. Microsatellite Polymorphism (Tttta)(n) at -528 Base Pairs of Gene CYP11alpha Influences Hyperandrogenemia in Patients with Polycystic Ovary Syndrome. Fertil. Steril. 2000, 73, 735–741. [Google Scholar] [CrossRef]
- Mehdizadeh, A.; Kalantar, S.M.; Sheikhha, M.H.; Aali, B.S.; Ghanei, A. Association of SNP Rs.2414096 CYP19 Gene with Polycystic Ovarian Syndrome in Iranian Women. Int. J. Reprod. Biomed. 2017, 15, 491–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chua, A.K.; Azziz, R.; Goodarzi, M.O. Association Study of CYP17 and HSD11B1 in Polycystic Ovary Syndrome Utilizing Comprehensive Gene Coverage. Mol. Hum. Reprod. 2012, 18, 320–324. [Google Scholar] [CrossRef]
- Wickenheisser, J.K.; Quinn, P.G.; Nelson, V.L.; Legro, R.S.; Strauss, J.F.; McAllister, J.M. Differential Activity of the Cytochrome P450 17alpha-Hydroxylase and Steroidogenic Acute Regulatory Protein Gene Promoters in Normal and Polycystic Ovary Syndrome Theca Cells. J. Clin. Endocrinol. Metab. 2000, 85, 2304–2311. [Google Scholar] [CrossRef] [Green Version]
- Deepika, M.L.N.; Reddy, K.R.; Yashwanth, A.; Rani, V.U.; Latha, K.P.; Jahan, P. TNF-α Haplotype Association with Polycystic Ovary Syndrome–a South Indian Study. J. Assist. Reprod. Genet. 2013, 30, 1493–1503. [Google Scholar] [CrossRef] [Green Version]
- Deswal, R.; Yadav, A.; Dang, A.S. Sex Hormone Binding Globulin–an Important Biomarker for Predicting PCOS Risk: A Systematic Review and Meta-Analysis. Syst. Biol. Reprod. Med. 2018, 64, 12–24. [Google Scholar] [CrossRef]
- Dasgupta, S.; Sirisha, P.V.S.; Neelaveni, K.; Anuradha, K.; Reddy, B.M. Association of CAPN10 SNPs and Haplotypes with Polycystic Ovary Syndrome among South Indian Women. PLoS ONE 2012, 7, e32192. [Google Scholar] [CrossRef] [Green Version]
- Wiltgen, D.; Furtado, L.; Kohek, M.B.F.; Spritzer, P.M. CAPN10 UCSNP-43, UCSNP-19 and UCSNP-63 Polymorphisms and Metabolic Syndrome in Polycystic Ovary Syndrome. Gynecol. Endocrinol. 2007, 23, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Mutib, M.T.; Hamdan, F.B.; Al-Salihi, A.R. INSR Gene Variation Is Associated with Decreased Insulin Sensitivity in Iraqi Women with PCOs. Iran. J. Reprod. Med. 2014, 12, 499–506. [Google Scholar]
- Daghestani, M.H. Rs1799817 in INSR Associates with Susceptibility to Polycystic Ovary Syndrome. J. Med. Biochem. 2020, 39, 149. [Google Scholar] [CrossRef] [PubMed]
- Song, D.K.; Lee, H.; Oh, J.-Y.; Hong, Y.S.; Sung, Y.-A. FTO Gene Variants Are Associated with PCOS Susceptibility and Hyperandrogenemia in Young Korean Women. Diabetes Metab. J. 2014, 38, 302. [Google Scholar] [CrossRef] [Green Version]
- Chaudhary, H.; Patel, J.; Jain, N.K.; Joshi, R. The Role of Polymorphism in Various Potential Genes on Polycystic Ovary Syndrome Susceptibility and Pathogenesis. J. Ovarian Res. 2021, 14, 125. [Google Scholar] [CrossRef]
- Ashraf, S.; Rasool, S.U.A.; Nabi, M.; Ganie, M.A.; Masoodi, S.R.; Amin, S. Impact of Rs2414096 Polymorphism of CYP19 Gene on Susceptibility of Polycystic Ovary Syndrome and Hyperandrogenism in Kashmiri Women. Sci. Rep. 2021, 11, 12942. [Google Scholar] [CrossRef]
- Jin, J.-L.; Sun, J.; Ge, H.-J.; Cao, Y.-X.; Wu, X.-K.; Liang, F.-J.; Sun, H.-X.; Ke, L.; Yi, L.; Wu, Z.-W.; et al. Association between CYP19 Gene SNP Rs2414096 Polymorphism and Polycystic Ovary Syndrome in Chinese Women. BMC Med. Genet. 2009, 10, 139. [Google Scholar] [CrossRef] [Green Version]
- Munawar Lone, N.; Babar, S.; Sultan, S.; Malik, S.; Nazeer, K.; Riaz, S. Association of the CYP17 and CYP19 Gene Polymorphisms in Women with Polycystic Ovary Syndrome from Punjab, Pakistan. Gynecol. Endocrinol. 2021, 37, 456–461. [Google Scholar] [CrossRef]
- Astapova, O.; Minor, B.M.N.; Hammes, S.R. Physiological and Pathological Androgen Actions in the Ovary. Endocrinology 2019, 160, 1166–1174. [Google Scholar] [CrossRef]
- Polat, S.; Karaburgu, S.; Unluhizarci, K.; Dündar, M.; Özkul, Y.; Arslan, Y.K.; Karaca, Z.; Kelestimur, F. The Role of Androgen Receptor CAG Repeat Polymorphism in Androgen Excess Disorder and Idiopathic Hirsutism. J. Endocrinol. Invest. 2020, 43, 1271–1281. [Google Scholar] [CrossRef]
- Li, Y.; Fang, L.; Yan, Y.; Wang, Z.; Wu, Z.; Jia, Q.; Cheng, J.-C.; Sun, Y.-P. Association between Human SHBG Gene Polymorphisms and Risk of PCOS: A Meta-Analysis. Reprod. BioMed. Online 2021, 42, 227–236. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Chen, Z.; Feng, W.; Long, S.; Mo, Z.-C. Sex Hormone-Binding Globulin and Polycystic Ovary Syndrome. Clin. Chim. Acta 2019, 499, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Deswal, R.; Nanda, S.; Dang, A.S. Association of Luteinizing Hormone and LH Receptor Gene Polymorphism with Susceptibility of Polycystic Ovary Syndrome. Syst. Biol. Reprod. Med. 2019, 65, 400–408. [Google Scholar] [CrossRef]
- Abutorabi, E.S.; Rashidi, B.H.; Irani, S.; Haghollahi, F.; Bagheri, M. Investigation of the FSHR, CYP11, and INSR Mutations and Polymorphisms in Iranian Infertile Women with Polycystic Ovary Syndrome (PCOS). Rep. Biochem. Mol. Biol. 2021, 9, 470–477. [Google Scholar] [CrossRef] [PubMed]
- Ferk, P.; Teran, N.; Gersak, K. The (TAAAA)n Microsatellite Polymorphism in the SHBG Gene Influences Serum SHBG Levels in Women with Polycystic Ovary Syndrome. Hum. Reprod. 2007, 22, 1031–1036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Concha, F.; Recabarren, S.E.; Pérez, B.F. Epigenética Del Síndrome de Ovario Poliquístico. Rev. Méd. Chile 2017, 145, 907–915. [Google Scholar] [CrossRef] [Green Version]
- Illingworth, R.; Kerr, A.; Desousa, D.; Jørgensen, H.; Ellis, P.; Stalker, J.; Jackson, D.; Clee, C.; Plumb, R.; Rogers, J.; et al. A Novel CpG Island Set Identifies Tissue-Specific Methylation at Developmental Gene Loci. PLoS Biol. 2008, 6, e22. [Google Scholar] [CrossRef] [Green Version]
- Vázquez-Martínez, E.R.; Gómez-Viais, Y.I.; García-Gómez, E.; Reyes-Mayoral, C.; Reyes-Muñoz, E.; Camacho-Arroyo, I.; Cerbón, M. DNA Methylation in the Pathogenesis of Polycystic Ovary Syndrome. Reproduction 2019, 158, R27–R40. [Google Scholar] [CrossRef]
- Cui, P.; Ma, T.; Tamadon, A.; Han, S.; Li, B.; Chen, Z.; An, X.; Shao, L.R.; Wang, Y.; Feng, Y. Hypothalamic DNA Methylation in Rats with Dihydrotestosterone-Induced Polycystic Ovary Syndrome: Effects of Low-Frequency Electro-Acupuncture. Exp. Physiol. 2018, 103, 1618–1632. [Google Scholar] [CrossRef]
- Kokosar, M.; Benrick, A.; Perfilyev, A.; Fornes, R.; Nilsson, E.; Maliqueo, M.; Behre, C.J.; Sazonova, A.; Ohlsson, C.; Ling, C.; et al. Epigenetic and Transcriptional Alterations in Human Adipose Tissue of Polycystic Ovary Syndrome. Sci. Rep. 2016, 6, 22883. [Google Scholar] [CrossRef] [Green Version]
- Shen, H.-R.; Qiu, L.-H.; Zhang, Z.-Q.; Qin, Y.-Y.; Cao, C.; Di, W. Genome-Wide Methylated DNA Immunoprecipitation Analysis of Patients with Polycystic Ovary Syndrome. PLoS ONE 2013, 8, e64801. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.-X.; Wei, J.-Z.; Jiao, J.; Jiang, S.-Y.; Yu, D.-H.; Li, D. Genome-Wide DNA Methylation and Gene Expression Patterns Provide Insight into Polycystic Ovary Syndrome Development. Oncotarget 2014, 5, 6603–6610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, F.; Wang, F.-F.; Yin, R.; Ding, G.-L.; El-prince, M.; Gao, Q.; Shi, B.-W.; Pan, H.-H.; Huang, Y.-T.; Jin, M.; et al. A Molecular Mechanism Underlying Ovarian Dysfunction of Polycystic Ovary Syndrome: Hyperandrogenism Induces Epigenetic Alterations in the Granulosa Cells. Mol. Med. 2012, 90, 911–923. [Google Scholar] [CrossRef]
- Jiang, L.-L.; Xie, J.-K.; Cui, J.-Q.; Wei, D.; Yin, B.-L.; Zhang, Y.-N.; Chen, Y.-H.; Han, X.; Wang, Q.; Zhang, C.-L. Promoter Methylation of Yes-Associated Protein (YAP1) Gene in Polycystic Ovary Syndrome. Medicine 2017, 96, e5768. [Google Scholar] [CrossRef]
- Sagvekar, P.; Kumar, P.; Mangoli, V.; Desai, S.; Mukherjee, S. DNA Methylome Profiling of Granulosa Cells Reveals Altered Methylation in Genes Regulating Vital Ovarian Functions in Polycystic Ovary Syndrome. Clin. Epigenet. 2019, 11, 61. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Zhu, D.; Duan, H.; Ren, A.; Glintborg, D.; Andersen, M.; Skov, V.; Thomassen, M.; Kruse, T.; Tan, Q. Differential DNA Methylation Patterns of Polycystic Ovarian Syndrome in Whole Blood of Chinese Women. Oncotarget 2017, 8, 20656–20666. [Google Scholar] [CrossRef] [Green Version]
- Sang, Q.; Zhang, S.; Zou, S.; Wang, H.; Feng, R.; Li, Q.; Jin, L.; He, L.; Xing, Q.; Wang, L. Quantitative Analysis of Follistatin (FST) Promoter Methylation in Peripheral Blood of Patients with Polycystic Ovary Syndrome. Reprod. BioMed. Online 2013, 26, 157–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sang, Q.; Li, X.; Wang, H.; Wang, H.; Zhang, S.; Feng, R.; Xu, Y.; Li, Q.; Zhao, X.; Xing, Q.; et al. Quantitative Methylation Level of the EPHX1 Promoter in Peripheral Blood DNA Is Associated with Polycystic Ovary Syndrome. PLoS ONE 2014, 9, e88013. [Google Scholar] [CrossRef]
- Ting, W.; Yanyan, Q.; Jian, H.; Keqin, H.; Duan, M. The Relationship Between Insulin Resistance and CpG Island Methylation of LMNA Gene in Polycystic Ovary Syndrome. Cell Biochem. Biophys. 2013, 67, 1041–1047. [Google Scholar] [CrossRef]
- Teede, H.; Ng, S.; Hedger, M.; Moran, L. Follistatin and Activins in Polycystic Ovary Syndrome: Relationship to Metabolic and Hormonal Markers. Metabolism 2013, 62, 1394–1400. [Google Scholar] [CrossRef]
- Hai, L.; McGee, S.R.; Rabideau, A.C.; Paquet, M.; Narayan, P. Infertility in Female Mice with a Gain-of-Function Mutation in the Luteinizing Hormone Receptor Is Due to Irregular Estrous Cyclicity, Anovulation, Hormonal Alterations, and Polycystic Ovaries1. Biol. Reprod. 2015, 93, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korhonen, S. Two Exonic Single Nucleotide Polymorphisms in the Microsomal Epoxide Hydrolase Gene Are Associated with Polycystic Ovary Syndrome. Fertil. Steril. 2003, 79, 1353–1357. [Google Scholar] [CrossRef]
- Vandenbeek, R.; Khan, N.P.; Estall, J.L. Linking Metabolic Disease With the PGC-1α Gly482Ser Polymorphism. Endocrinology 2018, 159, 853–865. [Google Scholar] [CrossRef]
- Titus, A.J.; Gallimore, R.M.; Salas, L.A.; Christensen, B.C. Cell-Type Deconvolution from DNA Methylation: A Review of Recent Applications. Hum. Mol. Genet. 2017, 26, R216–R224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luense, L.J.; Veiga-Lopez, A.; Padmanabhan, V.; Christenson, L.K. Developmental Programming: Gestational Testosterone Treatment Alters Fetal Ovarian Gene Expression. Endocrinology 2011, 152, 4974–4983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eini, F.; Novin, M.G.; Joharchi, K.; Hosseini, A.; Nazarian, H.; Piryaei, A.; Bidadkosh, A. Intracytoplasmic Oxidative Stress Reverses Epigenetic Modifications in Polycystic Ovary Syndrome. Reprod. Fertil. Dev. 2017, 29, 2313. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.-N.; Guo, L.; Hou, Y.; Ou, X.-H.; Liu, Z.; Sun, Q.-Y. The DNA Methylation Profile of Oocytes in Mice with Hyperinsulinaemia and Hyperandrogenism as Detected by Single-Cell Level Whole Genome Bisulphite Sequencing (SC-WGBS) Technology. Reprod. Fertil. Dev. 2018, 30, 1713. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Li, T.; Zhao, H.; Mao, Z.; Ding, C.; Kang, Y. Integrated Transcriptomic and Epigenetic Study of PCOS: Impact of Map3k1 and Map1lc3a Promoter Methylation on Autophagy. Front. Genet. 2021, 12, 620241. [Google Scholar] [CrossRef]
- Mimouni, N.E.H.; Paiva, I.; Barbotin, A.-L.; Timzoura, F.E.; Plassard, D.; Le Gras, S.; Ternier, G.; Pigny, P.; Catteau-Jonard, S.; Simon, V.; et al. Polycystic Ovary Syndrome Is Transmitted via a Transgenerational Epigenetic Process. Cell Metab. 2021, 33, 513–530.e8. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.-X.; Tan, Y.-J.; Wang, F.-F.; Hou, N.-N.; Xiang, Y.-Q.; Zhang, J.-Y.; Liu, Y.; Qu, F.; Meng, Q.; Xu, J.; et al. Aberrant Expression and DNA Methylation of Lipid Metabolism Genes in PCOS: A New Insight into Its Pathogenesis. Clin. Epigenet. 2018, 10, 6. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Jiao, J.; Zhou, Y.-M.; Wang, X.-X. Epigenetic Regulation of Traf2- and Nck-Interacting Kinase (TNIK) in Polycystic Ovary Syndrome. Am. J. Transl. Res. 2015, 7, 1152–1160. [Google Scholar] [PubMed]
- Jones, M.R.; Brower, M.A.; Xu, N.; Cui, J.; Mengesha, E.; Chen, Y.-D.I.; Taylor, K.D.; Azziz, R.; Goodarzi, M.O. Systems Genetics Reveals the Functional Context of PCOS Loci and Identifies Genetic and Molecular Mechanisms of Disease Heterogeneity. PLoS Genet. 2015, 11, e1005455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumariya, S.; Ubba, V.; Jha, R.K.; Gayen, J.R. Autophagy in Ovary and Polycystic Ovary Syndrome: Role, Dispute and Future Perspective. Autophagy 2021, 17, 2706–2733. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Heikkinen, L.; Wang, C.; Yang, Y.; Sun, H.; Wong, G. Trends in the Development of MiRNA Bioinformatics Tools. Brief. Bioinform. 2019, 20, 1836–1852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, Z.; Li, T.; Zhao, H.; Qin, Y.; Wang, X.; Kang, Y. Identification of Epigenetic Interactions between MicroRNA and DNA Methylation Associated with Polycystic Ovarian Syndrome. J. Hum. Genet. 2021, 66, 123–137. [Google Scholar] [CrossRef]
- Jiang, X.; Li, J.; Zhang, B.; Hu, J.; Ma, J.; Cui, L.; Chen, Z.-J. Differential Expression Profile of Plasma Exosomal MicroRNAs in Women with Polycystic Ovary Syndrome. Fertil. Steril. 2021, 115, 782–792. [Google Scholar] [CrossRef]
- Chen, Z.; Ou, H.; Wu, H.; Wu, P.; Mo, Z. Role of MicroRNA in the Pathogenesis of Polycystic Ovary Syndrome. DNA Cell Biol. 2019, 38, 754–762. [Google Scholar] [CrossRef]
- Long, W.; Zhao, C.; Ji, C.; Ding, H.; Cui, Y.; Guo, X.; Shen, R.; Liu, J. Characterization of Serum MicroRNAs Profile of PCOS and Identification of Novel Non-Invasive Biomarkers. Cell Physiol. Biochem. 2014, 33, 1304–1315. [Google Scholar] [CrossRef]
- Mao, Z.; Fan, L.; Yu, Q.; Luo, S.; Wu, X.; Tang, J.; Kang, G.; Tang, L. Abnormality of Klotho Signaling Is Involved in Polycystic Ovary Syndrome. Reprod. Sci. 2018, 25, 372–383. [Google Scholar] [CrossRef]
- Xu, B.; Zhang, Y.-W.; Tong, X.-H.; Liu, Y.-S. Characterization of MicroRNA Profile in Human Cumulus Granulosa Cells: Identification of MicroRNAs That Regulate Notch Signaling and Are Associated with PCOS. Mol. Cell Endocrinol. 2015, 404, 26–36. [Google Scholar] [CrossRef]
- Lin, L.; Du, T.; Huang, J.; Huang, L.-L.; Yang, D.-Z. Identification of Differentially Expressed MicroRNAs in the Ovary of Polycystic Ovary Syndrome with Hyperandrogenism and Insulin Resistance. Chin. Med. J. 2015, 128, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Luo, S.; Li, S.-W. MiRNA-592 Is Downregulated and May Target LHCGR in Polycystic Ovary Syndrome Patients. Reprod. Biol. 2015, 15, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, E.; Shahhoseini, M.; Afsharian, P.; Karimian, L.; Ashrafi, M.; Mehraein, F.; Afatoonian, R. Role of Epigenetic Modifications in the Aberrant CYP19A1 Gene Expression in Polycystic Ovary Syndrome. Arch. Med. Sci. AMS 2019, 15, 887–895. [Google Scholar] [CrossRef] [PubMed]
- Leung, K.L.; Sanchita, S.; Pham, C.T.; Davis, B.A.; Okhovat, M.; Ding, X.; Dumesic, P.; Grogan, T.R.; Williams, K.J.; Morselli, M.; et al. Dynamic Changes in Chromatin Accessibility, Altered Adipogenic Gene Expression, and Total versus de Novo Fatty Acid Synthesis in Subcutaneous Adipose Stem Cells of Normal-Weight Polycystic Ovary Syndrome (PCOS) Women during Adipogenesis: Evidence of Cellular Programming. Clin. Epigenet. 2020, 12, 181. [Google Scholar] [CrossRef]
- Romitti, M.; Fabris, V.C.; Ziegelmann, P.K.; Maia, A.L.; Spritzer, P.M. Association between PCOS and Autoimmune Thyroid Disease: A Systematic Review and Meta-Analysis. Endocr. Connect. 2018, 7, 1158–1167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serin, A.N.; Birge, Ö.; Uysal, A.; Görar, S.; Tekeli, F. Hashimoto’s Thyroiditis Worsens Ovaries in Polycystic Ovary Syndrome Patients Compared to Anti-Müllerian Hormone Levels. BMC Endocr. Disord. 2021, 21, 44. [Google Scholar] [CrossRef] [PubMed]
- Adamska, A.; Łebkowska, A.; Krentowska, A.; Hryniewicka, J.; Adamski, M.; Leśniewska, M.; Polak, A.M.; Kowalska, I. Ovarian Reserve and Serum Concentration of Thyroid Peroxidase Antibodies in Euthyroid Women With Different Polycystic Ovary Syndrome Phenotypes. Front. Endocrinol. 2020, 11, 440. [Google Scholar] [CrossRef]
- Krishna, M.B.; Joseph, A.; Subramaniam, A.G.; Gupta, A.; Pillai, S.M.; Laloraya, M. Reduced Tregs in Peripheral Blood of PCOS Patients–a Consequence of Aberrant Il2 Signaling. J. Clin. Endocrinol. Metab. 2015, 100, 282–292. [Google Scholar] [CrossRef] [Green Version]
- Rashid, A.; Bhat, J.A.; Ganie, M.A.; Wani, I.A.; Bhat, M.H.; Shah, Z.A.; Masoodi, S.R.; Marwaha, R.K. Evaluation of Serum Anti-Nuclear Antibody among Women with PCOS: A Hospital Based Single Center Cross Sectional Study. Gynecol. Endocrinol. 2018, 34, 965–969. [Google Scholar] [CrossRef]
- Palacio, J.R.; Iborra, A.; Ulcova-Gallova, Z.; Badia, R.; Martínez, P. The Presence of Antibodies to Oxidative Modified Proteins in Serum from Polycystic Ovary Syndrome Patients. Clin. Exp. Immunol. 2006, 144, 217–222. [Google Scholar] [CrossRef]
- Gingnell, M.; Dahlbom, I.; Lindholm, Å.; Hudecova, M.; Arnadottir, R.; Hansson, T.; Sundstrom-Poromaa, I. Patients with Polycystic Ovary Syndrome Have Lower Levels of IgM Anti-Phosphorylcholine Antibodies than Healthy Women. Gynecol. Endocrinol. 2011, 27, 486–490. [Google Scholar] [CrossRef]
- Li, H.; Guo, Y.; Deng, J.; Fischer, H.; Weedin, E.A.; Burks, H.R.; Craig, L.B.; Yu, X. Increased Testosterone and Proinflammatory Cytokines in Patients with Polycystic Ovary Syndrome Correlate with Elevated GnRH Receptor Autoantibody Activity Assessed by a Fluorescence Resonance Energy Transfer-Based Bioassay. Endocrine 2021, 74, 163–171. [Google Scholar] [CrossRef]
- Weedin, E.A.; Burks, H.R.; Yu, X.; Li, H.L.; Aston, C.E.; Kem, D.C.; Craig, L.B. Elevated Activity Levels of Activating Autoantibodies to the GnRH Receptor in Patients with Polycystic Ovary Syndrome. F&S Rep. 2020, 1, 299–304. [Google Scholar] [CrossRef]
- Li, H.; Zhang, G.; Guo, Y.; Deng, J.; Fischer, H.; Craig, L.B.; Kem, D.C.; Yu, X. Autoimmune Activation of the GnRH Receptor Induces Insulin Resistance Independent of Obesity in a Female Rat Model. Physiol. Rep. 2021, 8, e14672. [Google Scholar] [CrossRef]
- Schniewind, H.A.; Sattler, L.-M.; Haudum, C.W.; Münzker, J.; Minich, W.B.; Obermayer-Pietsch, B.; Schomburg, L. Autoimmunity to the Follicle-Stimulating Hormone Receptor (FSHR) and Luteinizing Hormone Receptor (LHR) in Polycystic Ovarian Syndrome. IJMS 2021, 22, 13667. [Google Scholar] [CrossRef] [PubMed]
- Panidis, D.; Tziomalos, K.; Papadakis, E.; Vosnakis, C.; Chatzis, P.; Katsikis, I. Lifestyle Intervention and Anti-Obesity Therapies in the Polycystic Ovary Syndrome: Impact on Metabolism and Fertility. Endocrine 2013, 44, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Cena, H.; Chiovato, L.; Nappi, R.E. Obesity, Polycystic Ovary Syndrome, and Infertility: A New Avenue for GLP-1 Receptor Agonists. J. Clin. Endocrinol. Metab. 2020, 105, e2695–e2709. [Google Scholar] [CrossRef] [PubMed]
- Silvestris, E.; de Pergola, G.; Rosania, R.; Loverro, G. Obesity as Disruptor of the Female Fertility. Reprod. Biol. Endocrinol. 2018, 16, 22. [Google Scholar] [CrossRef]
- Asdaq, S.M.B.; Jomah, S.; Hasan, R.; Al-Baroudi, D.; Alharbi, M.; Alsubaie, S.; Buhamad, M.H.; Alyahya, B.; Al-Yamani, M.J. Impact of Polycystic Ovary Syndrome on Eating Behavior, Depression and Health Related Quality of Life: A Cross-Sectional Study in Riyadh. Saudi J. Biol. Sci. 2020, 27, 3342–3347. [Google Scholar] [CrossRef]
- Sadeeqa, S.; Mustafa, T.; Latif, S. Polycystic Ovarian Syndrome–Related Depression in Adolescent Girls: A Review. J. Pharm. Bioall. Sci. 2018, 10, 55. [Google Scholar] [CrossRef]
- Kolhe, J.V.; Chhipa, A.S.; Butani, S.; Chavda, V.; Patel, S.S. PCOS and Depression: Common Links and Potential Targets. Reprod. Sci. 2021, 1–18, Epub ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Simon, S.; Rahat, H.; Carreau, A.-M.; Garcia-Reyes, Y.; Halbower, A.; Pyle, L.; Nadeau, K.J.; Cree-Green, M. Poor Sleep Is Related to Metabolic Syndrome Severity in Adolescents With PCOS and Obesity. J. Clin. Endocrinol. Metab. 2020, 105, e1827–e1834. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, R.; Azevedo, I. Chronic Inflammation in Obesity and the Metabolic Syndrome. Mediat. Inflamm. 2010, 2010, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Mesarwi, O.A.; Loomba, R.; Malhotra, A. Obstructive Sleep Apnea, Hypoxia, and Nonalcoholic Fatty Liver Disease. Am. J. Respir. Crit. Care Med. 2019, 199, 830–841. [Google Scholar] [CrossRef] [PubMed]
- Leproult, R.; Van Cauter, E. Role of Sleep and Sleep Loss in Hormonal Release and Metabolism. In Endocrine Development; Loche, S., Cappa, M., Ghizzoni, L., Maghnie, M., Savage, M.O., Eds.; KARGER: Basel, Switzerland, 2009; Volume 17, pp. 11–21. ISBN 978-3-8055-9302-1. [Google Scholar]
- Kite, C.; Lahart, I.M.; Afzal, I.; Broom, D.R.; Randeva, H.; Kyrou, I.; Brown, J.E. Exercise, or Exercise and Diet for the Management of Polycystic Ovary Syndrome: A Systematic Review and Meta-Analysis. Syst. Rev. 2019, 8, 51. [Google Scholar] [CrossRef]
- Szczuko, M.; Kikut, J.; Szczuko, U.; Szydłowska, I.; Nawrocka-Rutkowska, J.; Ziętek, M.; Verbanac, D.; Saso, L. Nutrition Strategy and Life Style in Polycystic Ovary Syndrome—Narrative Review. Nutrients 2021, 13, 2452. [Google Scholar] [CrossRef] [PubMed]
- Mehrabani, H.H.; Salehpour, S.; Amiri, Z.; Farahani, S.J.; Meyer, B.J.; Tahbaz, F. Beneficial Effects of a High-Protein, Low-Glycemic-Load Hypocaloric Diet in Overweight and Obese Women with Polycystic Ovary Syndrome: A Randomized Controlled Intervention Study. J. Am. Coll. Nutr. 2012, 31, 117–125. [Google Scholar] [CrossRef] [Green Version]
- Gower, B.A.; Chandler-Laney, P.C.; Ovalle, F.; Goree, L.L.; Azziz, R.; Desmond, R.A.; Granger, W.M.; Goss, A.M.; Bates, G.W. Favourable Metabolic Effects of a Eucaloric Lower-Carbohydrate Diet in Women with PCOS. Clin. Endocrinol. 2013, 79, 550–557. [Google Scholar] [CrossRef] [Green Version]
- Kazemi, M.; Hadi, A.; Pierson, R.A.; Lujan, M.E.; Zello, G.A.; Chilibeck, P.D. Effects of Dietary Glycemic Index and Glycemic Load on Cardiometabolic and Reproductive Profiles in Women with Polycystic Ovary Syndrome: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Adv. Nutr. 2021, 12, 161–178. [Google Scholar] [CrossRef]
- Cutler, D.A.; Pride, S.M.; Cheung, A.P. Low Intakes of Dietary Fiber and Magnesium Are Associated with Insulin Resistance and Hyperandrogenism in Polycystic Ovary Syndrome: A Cohort Study. Food Sci. Nutr. 2019, 7, 1426–1437. [Google Scholar] [CrossRef] [Green Version]
- Cincione, R.I.; Losavio, F.; Ciolli, F.; Valenzano, A.; Cibelli, G.; Messina, G.; Polito, R. Effects of Mixed of a Ketogenic Diet in Overweight and Obese Women with Polycystic Ovary Syndrome. IJERPH 2021, 18, 12490. [Google Scholar] [CrossRef] [PubMed]
- Mavropoulos, J.C.; Yancy, W.S.; Hepburn, J.; Westman, E.C. The Effects of a Low-Carbohydrate, Ketogenic Diet on the Polycystic Ovary Syndrome: A Pilot Study. Nutr. Metab. 2005, 2, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rudnicka, E.; Suchta, K.; Grymowicz, M.; Calik-Ksepka, A.; Smolarczyk, K.; Duszewska, A.M.; Smolarczyk, R.; Meczekalski, B. Chronic Low Grade Inflammation in Pathogenesis of PCOS. IJMS 2021, 22, 3789. [Google Scholar] [CrossRef] [PubMed]
- Youm, Y.-H.; Nguyen, K.Y.; Grant, R.W.; Goldberg, E.L.; Bodogai, M.; Kim, D.; D’Agostino, D.; Planavsky, N.; Lupfer, C.; Kanneganti, T.D.; et al. The Ketone Metabolite β-Hydroxybutyrate Blocks NLRP3 Inflammasome–Mediated Inflammatory Disease. Nat. Med. 2015, 21, 263–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maktabi, M.; Jamilian, M.; Asemi, Z. Magnesium-Zinc-Calcium-Vitamin D Co-Supplementation Improves Hormonal Profiles, Biomarkers of Inflammation and Oxidative Stress in Women with Polycystic Ovary Syndrome: A Randomized, Double-Blind, Placebo-Controlled Trial. Biol. Trace Elem. Res. 2018, 182, 21–28. [Google Scholar] [CrossRef]
- Mas-Bargues, C.; Escrivá, C.; Dromant, M.; Borrás, C.; Viña, J. Lipid Peroxidation as Measured by Chromatographic Determination of Malondialdehyde. Human Plasma Reference Values in Health and Disease. Arch. Biochem. Biophys. 2021, 709, 108941. [Google Scholar] [CrossRef]
- Afshar Ebrahimi, F.; Foroozanfard, F.; Aghadavod, E.; Bahmani, F.; Asemi, Z. The Effects of Magnesium and Zinc Co-Supplementation on Biomarkers of Inflammation and Oxidative Stress, and Gene Expression Related to Inflammation in Polycystic Ovary Syndrome: A Randomized Controlled Clinical Trial. Biol. Trace Elem. Res. 2018, 184, 300–307. [Google Scholar] [CrossRef]
- Hamilton, K.P.; Zelig, R.; Parker, A.R.; Haggag, A. Insulin Resistance and Serum Magnesium Concentrations among Women with Polycystic Ovary Syndrome. Curr. Dev. Nutr. 2019, 3, nzz108. [Google Scholar] [CrossRef]
- Guo, S.; Tal, R.; Jiang, H.; Yuan, T.; Liu, Y. Vitamin D Supplementation Ameliorates Metabolic Dysfunction in Patients with PCOS: A SystematicReview of RCTs and Insight into the Underlying Mechanism. Int. J. Endocrinol. 2020, 2020, 1–18. [Google Scholar] [CrossRef]
- Zhao, J.-F.; Li, B.-X.; Zhang, Q. Vitamin D Improves Levels of Hormonal, Oxidative Stress and Inflammatory Parameters in Polycystic Ovary Syndrome: A Meta-Analysis Study. Ann. Palliat. Med. 2021, 10, 169–183. [Google Scholar] [CrossRef]
- Costantini, L.; Molinari, R.; Farinon, B.; Merendino, N. Impact of Omega-3 Fatty Acids on the Gut Microbiota. IJMS 2017, 18, 2645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, J.; Wen, X.; Jia, M. Efficacy of Omega-3 Polyunsaturated Fatty Acids on Hormones, Oxidative Stress, and Inflammatory Parameters among Polycystic Ovary Syndrome: A Systematic Review and Meta-Analysis. Ann. Palliat. Med. 2021, 10, 8991–9001. [Google Scholar] [CrossRef] [PubMed]
- Unfer, V. A Deeper Assessment of Ω3-Poly-Unsaturated Fatty Acids in Polycystic Ovary Syndrome Management. Comment on Regidor et al. Chronic Inflammation in PCOS: The Potential Benefits of Specialized Pro-Resolving Lipid Mediators (SPMs) in the Improvement of the Resolutive Response. Int. J. Mol. Sci. 2021, 22, 10114. [Google Scholar] [CrossRef]
- Salek, M.; Clark, C.C.T.; Taghizadeh, M.; Jafarnejad, S. N-3 Fatty Acids as Preventive and Therapeutic Agents in Attenuating PCOS Complications. EXCLI J. 2019, 18, 558. [Google Scholar] [CrossRef] [PubMed]
- Nordio, M.; Basciani, S.; Camajani, E. The 40:1 Myo-Inositol/D-Chiro-Inositol Plasma Ratio Is Able to Restore Ovulation in PCOS Patients: Comparison with Other Ratios. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 5512–5521. [Google Scholar] [CrossRef]
- Kamenov, Z.; Gateva, A. Inositols in PCOS. Molecules 2020, 25, 5566. [Google Scholar] [CrossRef]
- Colak, E.; Ozcimen, E.E.; Tohma, Y.A.; Ceran, M.U. May Myo-inositol and d -chiro-inositol (40:1) Treatment Be a Good Option on Normal-weighted Polycystic Ovary Syndrome Patients without Insulin Resistance? J. Obstet. Gynaecol. Res. 2020, 46, 2605–2611. [Google Scholar] [CrossRef]
- Artini, P.G.; Malacarne, E.; Tomatis, V.; Genazzani, A.D. The Relevance of Inositols Treatment for PCOS before and during ART. Eur. Rev. Med. Pharmacol. Sci. 2021, 25, 4799–4809. [Google Scholar] [CrossRef]
- Di Emidio, G.; Rea, F.; Placidi, M.; Rossi, G.; Cocciolone, D.; Virmani, A.; Macchiarelli, G.; Palmerini, M.G.; D’Alessandro, A.M.; Artini, P.G.; et al. Regulatory Functions of L-Carnitine, Acetyl, and Propionyl L-Carnitine in a PCOS Mouse Model: Focus on Antioxidant/Antiglycative Molecular Pathways in the Ovarian Microenvironment. Antioxidants 2020, 9, 867. [Google Scholar] [CrossRef]
- Tauqir, S.; Israr, M.; Rauf, B.; Malik, M.O.; Habib, S.H.; Shah, F.A.; Usman, M.; Raza, M.A.; Shah, I.; Badshah, H.; et al. Acetyl-l-Carnitine Ameliorates Metabolic and Endocrine Alterations in Women with PCOS: A Double-Blind Randomized Clinical Trial. Adv. Ther. 2021, 38, 3842–3856. [Google Scholar] [CrossRef]
- Akbaribazm, M.; Goodarzi, N.; Rahimi, M. Female Infertility and Herbal Medicine: An Overview of the New Findings. Food Sci. Nutr. 2021, 9, 5869–5882. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Lv, L.; Liu, Q.; Song, J. Total Flavonoids Extracted from Nervilia Fordii Function in Polycystic Ovary Syndrome through IL-6 Mediated JAK2/STAT3 Signaling Pathway. Biosci. Rep. 2019, 39, BSR20181380. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Jiang, Y.; Xi, H.; Chen, L.; Feng, X. Exploration of the Relationship Between Gut Microbiota and Polycystic Ovary Syndrome (PCOS): A Review. Geburtshilfe Frauenheilkd 2020, 80, 161–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Y.; Yu, J.; Liang, C.; Li, S.; Wen, X.; Li, Y. Characterization on Gut Microbiome of PCOS Rats and Its Further Design by Shifts in High-Fat Diet and Dihydrotestosterone Induction in PCOS Rats. Bioprocess. Biosyst. Eng. 2021, 44, 953–964. [Google Scholar] [CrossRef]
- Giampaolino, P.; Foreste, V.; Di Filippo, C.; Gallo, A.; Mercorio, A.; Serafino, P.; Improda, F.P.; Verrazzo, P.; Zara, G.; Buonfantino, C.; et al. Microbiome and PCOS: State-of-Art and Future Aspects. IJMS 2021, 22, 2048. [Google Scholar] [CrossRef]
- Wang, X.; Xu, T.; Liu, R.; Wu, G.; Gu, L.; Zhang, Y.; Zhang, F.; Fu, H.; Ling, Y.; Wei, X.; et al. High-Fiber Diet or Combined With Acarbose Alleviates Heterogeneous Phenotypes of Polycystic Ovary Syndrome by Regulating Gut Microbiota. Front. Endocrinol. 2022, 12, 806331. [Google Scholar] [CrossRef]
- Wang, T.; Sha, L.; Li, Y.; Zhu, L.; Wang, Z.; Li, K.; Lu, H.; Bao, T.; Guo, L.; Zhang, X.; et al. Dietary α-Linolenic Acid-Rich Flaxseed Oil Exerts Beneficial Effects on Polycystic Ovary Syndrome Through Sex Steroid Hormones—Microbiota—Inflammation Axis in Rats. Front. Endocrinol. 2020, 11, 284. [Google Scholar] [CrossRef]
- Woodward, A.; Klonizakis, M.; Broom, D. Exercise and Polycystic Ovary Syndrome. In Physical Exercise for Human Health; Xiao, J., Ed.; Advances in Experimental Medicine and Biology; Springer Singapore: Singapore, 2020; Volume 1228, pp. 123–136. ISBN 9789811517914. [Google Scholar]
- Benham, J.L.; Booth, J.E.; Corenblum, B.; Doucette, S.; Friedenreich, C.M.; Rabi, D.M.; Sigal, R.J. Exercise Training and Reproductive Outcomes in Women with Polycystic Ovary Syndrome: A Pilot Randomized Controlled Trial. Clin. Endocrinol. 2021, 95, 332–343. [Google Scholar] [CrossRef]
- Vigorito, C.; Giallauria, F.; Palomba, S.; Cascella, T.; Manguso, F.; Lucci, R.; De Lorenzo, A.; Tafuri, D.; Lombardi, G.; Colao, A.; et al. Beneficial Effects of a Three-Month Structured Exercise Training Program on Cardiopulmonary Functional Capacity in Young Women with Polycystic Ovary Syndrome. J. Clin. Endocrinol. Metab. 2007, 92, 1379–1384. [Google Scholar] [CrossRef] [Green Version]
- Moro, C.; Pasarica, M.; Elkind-Hirsch, K.; Redman, L.M. Aerobic Exercise Training Improves Atrial Natriuretic Peptide and Catecholamine-Mediated Lipolysis in Obese Women with Polycystic Ovary Syndrome. J. Clin. Endocrinol. Metab. 2009, 94, 2579–2586. [Google Scholar] [CrossRef] [Green Version]
- Lionett, S.; Kiel, I.A.; Camera, D.M.; Vanky, E.; Parr, E.B.; Lydersen, S.; Hawley, J.A.; Moholdt, T. Circulating and Adipose Tissue MiRNAs in Women With Polycystic Ovary Syndrome and Responses to High-Intensity Interval Training. Front. Physiol. 2020, 11, 904. [Google Scholar] [CrossRef] [PubMed]
- Visser, M.; Bouter, L.M.; McQuillan, G.M.; Wener, M.H.; Harris, T.B. Elevated C-Reactive Protein Levels in Overweight and Obese Adults. JAMA 1999, 282, 2131–2135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Covington, J.D.; Tam, C.S.; Pasarica, M.; Redman, L.M. Higher Circulating Leukocytes in Women with PCOS Is Reversed by Aerobic Exercise. Biochimie 2016, 124, 27–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, N.; Yang, C.; Xie, H.; Liu, Y.; Liao, Y. Effects of Aerobic Exercise on Rats with Hyperandrogenic Polycystic Ovarian Syndrome. Int. J. Endocrinol. 2021, 2021, 1–12. [Google Scholar] [CrossRef]
- Ribeiro, V.B.; Pedroso, D.C.C.; Kogure, G.S.; Lopes, I.P.; Santana, B.A.; Dutra de Souza, H.C.; Ferriani, R.A.; Calado, R.T.; Furtado, C.L.M.; Reis, R.M.D. Short-Term Aerobic Exercise Did Not Change Telomere Length While It Reduced Testosterone Levels and Obesity Indexes in PCOS: A Randomized Controlled Clinical Trial Study. Int. J. Environ. Res. Public Health 2021, 18, 11274. [Google Scholar] [CrossRef]
- Shele, G.; Genkil, J.; Speelman, D. A Systematic Review of the Effects of Exercise on Hormones in Women with Polycystic Ovary Syndrome. JFMK 2020, 5, 35. [Google Scholar] [CrossRef]
- Ribeiro, V.B.; Kogure, G.S.; Reis, R.M.; Gastaldi, A.C.; DE Araújo, J.E.; Mazon, J.H.; Borghi, A.; Souza, H.C.D. Polycystic Ovary Syndrome Presents Higher Sympathetic Cardiac Autonomic Modulation That Is Not Altered by Strength Training. Int. J. Exerc. Sci. 2016, 9, 554–566. [Google Scholar]
- Zhang, J.; Bao, Y.; Zhou, X.; Zheng, L. Polycystic Ovary Syndrome and Mitochondrial Dysfunction. Reprod. Biol. Endocrinol. 2019, 17, 67. [Google Scholar] [CrossRef]
- Malamouli, M.; Levinger, I.; McAinch, A.J.; Trewin, A.J.; Rodgers, R.J.; Moreno-Asso, A. The Mitochondrial Profile in Women with Polycystic Ovary Syndrome: Impact of Exercise. J. Mol. Endocrinol. 2022, 68, R11–R23. [Google Scholar] [CrossRef]
- Scarpulla, R.C. Metabolic Control of Mitochondrial Biogenesis through the PGC-1 Family Regulatory Network. Biochim. Biophys. Acta BBA Mol. Cell Res. 2011, 1813, 1269–1278. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Wu, H.; Sun, W.; Wang, C. Improvement of Anti-Müllerian Hormone and Oxidative Stress through Regular Exercise in Chinese Women with Polycystic Ovary Syndrome. Hormones 2021, 20, 339–345. [Google Scholar] [CrossRef]
- Polyzos, S.A.; Anastasilakis, A.D.; Efstathiadou, Z.A.; Makras, P.; Perakakis, N.; Kountouras, J.; Mantzoros, C.S. Irisin in Metabolic Diseases. Endocrine 2018, 59, 260–274. [Google Scholar] [CrossRef] [PubMed]
- Perakakis, N.; Triantafyllou, G.A.; Fernández-Real, J.M.; Huh, J.Y.; Park, K.H.; Seufert, J.; Mantzoros, C.S. Physiology and Role of Irisin in Glucose Homeostasis. Nat. Rev. Endocrinol. 2017, 13, 324–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pesce, M.; Ballerini, P.; Paolucci, T.; Puca, I.; Farzaei, M.H.; Patruno, A. Irisin and Autophagy: First Update. IJMS 2020, 21, 7587. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Gene | Encoded Protein | Implication | Reference |
|---|---|---|---|
| INSR | Insuline receptor | Insulin secretion and action | [27] |
| CAPN10 | Calpain 10 protein | [28] | |
| IRS1, IRS2 | Insulin receptor substrates IRS1 and IRS2 | [29] | |
| FSHR | Follicle-stimulating hormone receptor | Gonadotropin release regulation | [30] |
| LH | Beta subunit of luteinizing hormone | [31] | |
| AMH | Anti-Müllerian hormone | [32] | |
| CYP21 | 21-hydroxylase enzyme | Synthesis pathways of steroid hormones | [33] |
| CYP11a | A cytochrome P450 enzyme | [34] | |
| CYP19 | Cytochrome P450 aromatase | Androgen synthesis pathways | [35] |
| CYP17 | Enzyme cytochrome P450-C17 | [36,37] | |
| TNF-α | The cytokine Tumor Necrosis Factor | inflammation | [38] |
| SHBG | Sex Hormone–Binding Globulin | Regulation the androgen level in the body | [39] |
| Gene | Polymorphisms | Effects on PCOS Phenotype |
|---|---|---|
| Calpain 10 (CAPN10) | UCSNP-44 | PCOS pathogenesis [40] |
| UCSNP-43 | Worse metabolic profile [41] | |
| Insulin receptor gene (INSR) | rs1799817 | Worse glycemic pattern and obesity [42] |
| Fat mass and obesity-associated gene (FTO) | -rs1421085 -rs17817449 -rs8050136 | High androgen levels [44] |
| Cytochrome P450 enzymes (CYP): | ||
| -CYP 17 | -rs743572 | Steroidogenesis and alteration of hormonal pathways [46,47,48] |
| -CYP 19 | -rs2414096 | |
| Androgen receptor gene (AR) | poly-glutamine (CAG) repeat region | Amplified response to male hormones [50] |
| Sex hormone–binding globulin gene (SHBG) | short (TAAAA)n pentanucleotide repeat | Obesity, impaired lipid metabolism, hyperinsulinemia, hyperandrogenism and chronic inflammation [55] |
| Follicle-stimulating hormone receptor (FSHR) | -p.Ala307Thr -p.Asn680Ser | Impaired oocyte maturation, anovulation, infertility [54] |
| Genes | DNA Methylation Mechanism | Tissue | Clinical Effects Related to DNA Methylation Changes | References |
|---|---|---|---|---|
| LY6G6F, KCTD21, ADCY9, RABL2B, ZNF611, VASH1, FST, LMNA, PPARGC1A | Hypermethylation | Peripheral blood | Increased prolactin and estradiol levels in serum, increased free androgen index, insulin resistance, increased triglyceride levels in plasma and risk for metabolic syndrome | [61,62] |
| L-1, TMSB15B, RPF1, DNA2, EPHA8, LHCGR EPHX1 | Hypomethylation | |||
| MATN4, DLGAP2, CDH13, GAREM2, GSC, ANKRD34C, ATP8B2 PPARG | Hypermethylation | Granulosa cells | Hyperandrogenism | [62,63,64] |
| L-1, LHCGR, SMG6, CCR5, LHB, NTN1, ARFGAP1, MDGA1, NCOR1, YAP1, CD9, NR4A1, EDN2, BNIP3, LIF | Hypomethylation | |||
| ZZEF1, TPT1, STUB1, DMAP1, RAB5B, PPARG, SVEP1, SAV1, RORA, RAB6A CNST | Hypermethylation | Subcutaneous adipose tissue | Oligomenorrhea; Increased testosterone levels in circulation | [60] |
| PUM1, DIP2C, SNX8, SRGAP3, ZFHX3, OR52W1and BBX | Hypomethylation | |||
| TET1 ROBO 1 CDKN1A HDC IGFBPL1 IRS4 | Hypomethylation | Ovarian tissue | Pathogenesis of PCOS | [65] |
| TNIK | Hypermethylation | Granulosa cells | Altered metabolic profile | [61] |
| TNF | Hypermethylation | Granulosa cells | Inflammation and hyperandrogenism | [66] |
| AKR1C3 CASR GHRHR RETN MAMLD1 | Hypomethylation | Granulosa cells | Hyperandrogenism and hormonal disbalance | [66] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scarfò, G.; Daniele, S.; Fusi, J.; Gesi, M.; Martini, C.; Franzoni, F.; Cela, V.; Artini, P.G. Metabolic and Molecular Mechanisms of Diet and Physical Exercise in the Management of Polycystic Ovarian Syndrome. Biomedicines 2022, 10, 1305. https://doi.org/10.3390/biomedicines10061305
Scarfò G, Daniele S, Fusi J, Gesi M, Martini C, Franzoni F, Cela V, Artini PG. Metabolic and Molecular Mechanisms of Diet and Physical Exercise in the Management of Polycystic Ovarian Syndrome. Biomedicines. 2022; 10(6):1305. https://doi.org/10.3390/biomedicines10061305
Chicago/Turabian StyleScarfò, Giorgia, Simona Daniele, Jonathan Fusi, Marco Gesi, Claudia Martini, Ferdinando Franzoni, Vito Cela, and Paolo Giovanni Artini. 2022. "Metabolic and Molecular Mechanisms of Diet and Physical Exercise in the Management of Polycystic Ovarian Syndrome" Biomedicines 10, no. 6: 1305. https://doi.org/10.3390/biomedicines10061305
APA StyleScarfò, G., Daniele, S., Fusi, J., Gesi, M., Martini, C., Franzoni, F., Cela, V., & Artini, P. G. (2022). Metabolic and Molecular Mechanisms of Diet and Physical Exercise in the Management of Polycystic Ovarian Syndrome. Biomedicines, 10(6), 1305. https://doi.org/10.3390/biomedicines10061305