
ROCK and PDE-5 Inhibitors for the Treatment of Dementia: Literature Review and Meta-Analysis

 ,
,  , , ,
, , ,  , and
, and
Abstract
:1. Introduction
2. Pathophysiology of Dementia
3. Rho-Associated Protein Kinase (ROCK) and Dementia
3.1. ROCK and AD
3.2. ROCK and Vascular Remodeling
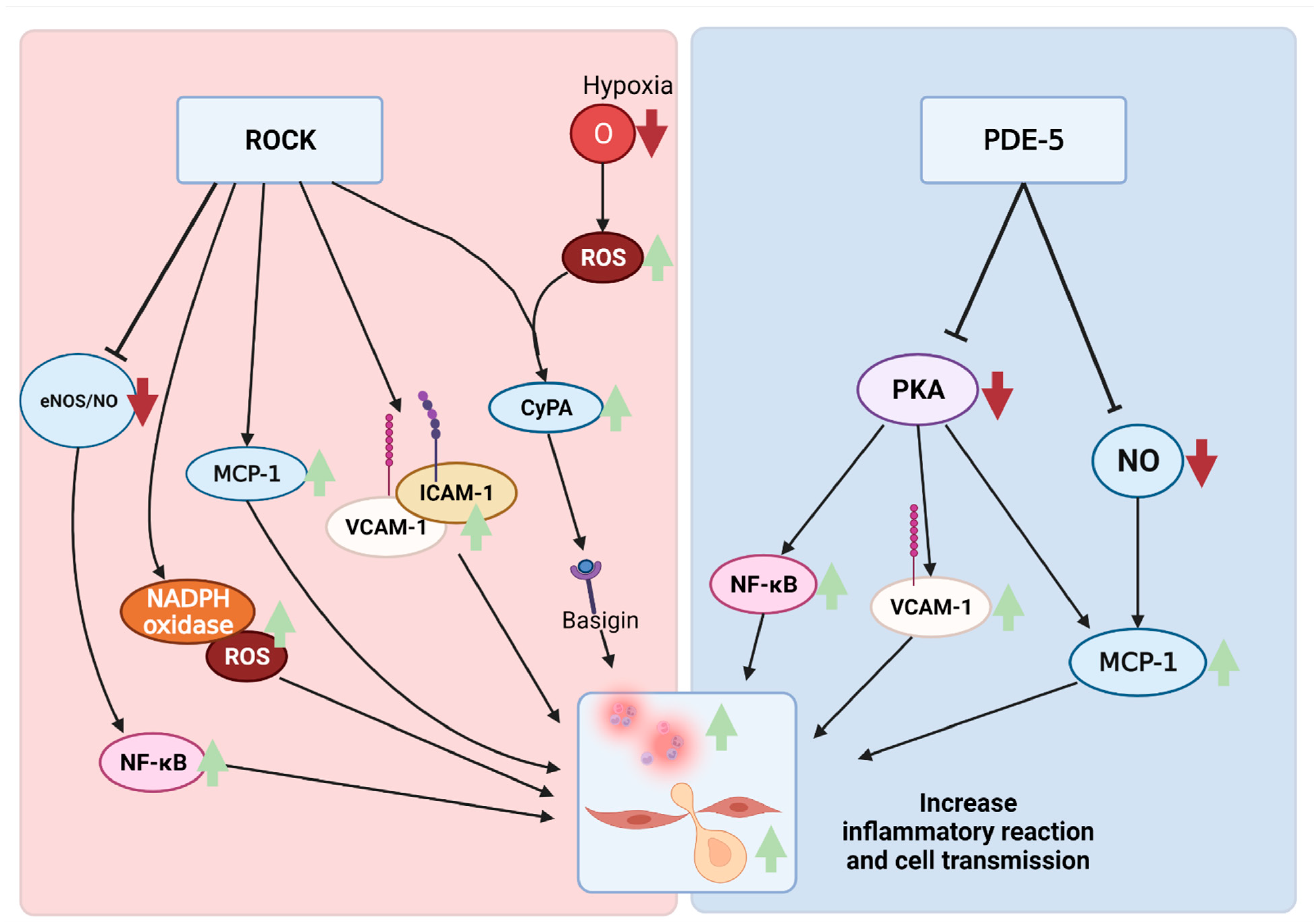
3.3. ROCK and Neuroinflammation
4. Phosphodiesterase-5 (PDE-5) and Dementia
4.1. PDE-5 and AD
4.2. PDE-5 and Vascular Remodeling
4.3. PDE-5 and Neuroinflammation
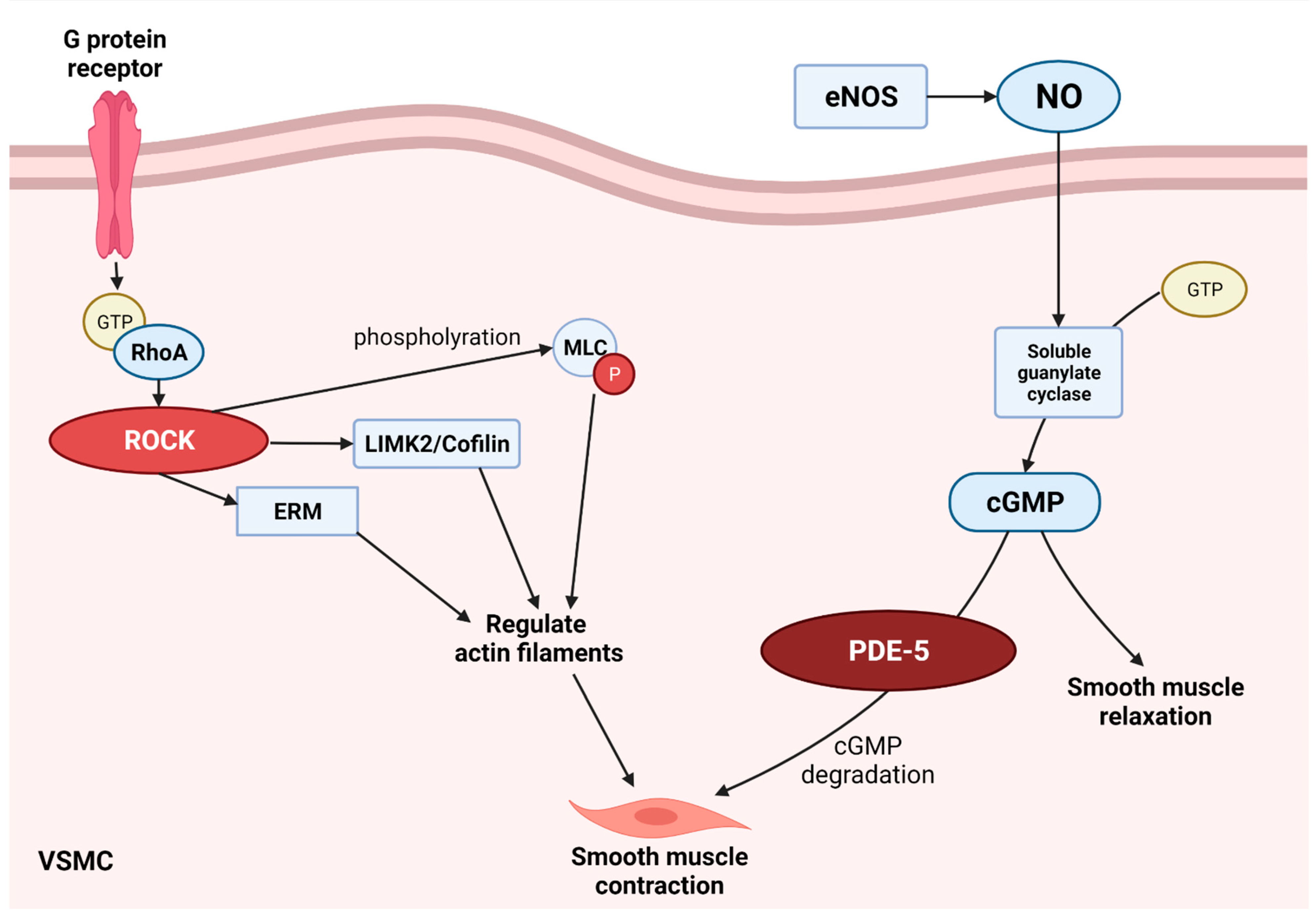
5. Inhibition of ROCK and PDE-5 Pathway for Neuroprotection
6. Meta-Analysis of ROCK Inhibitors and PDE-5 Inhibitors in Animal Experiments
6.1. Methods
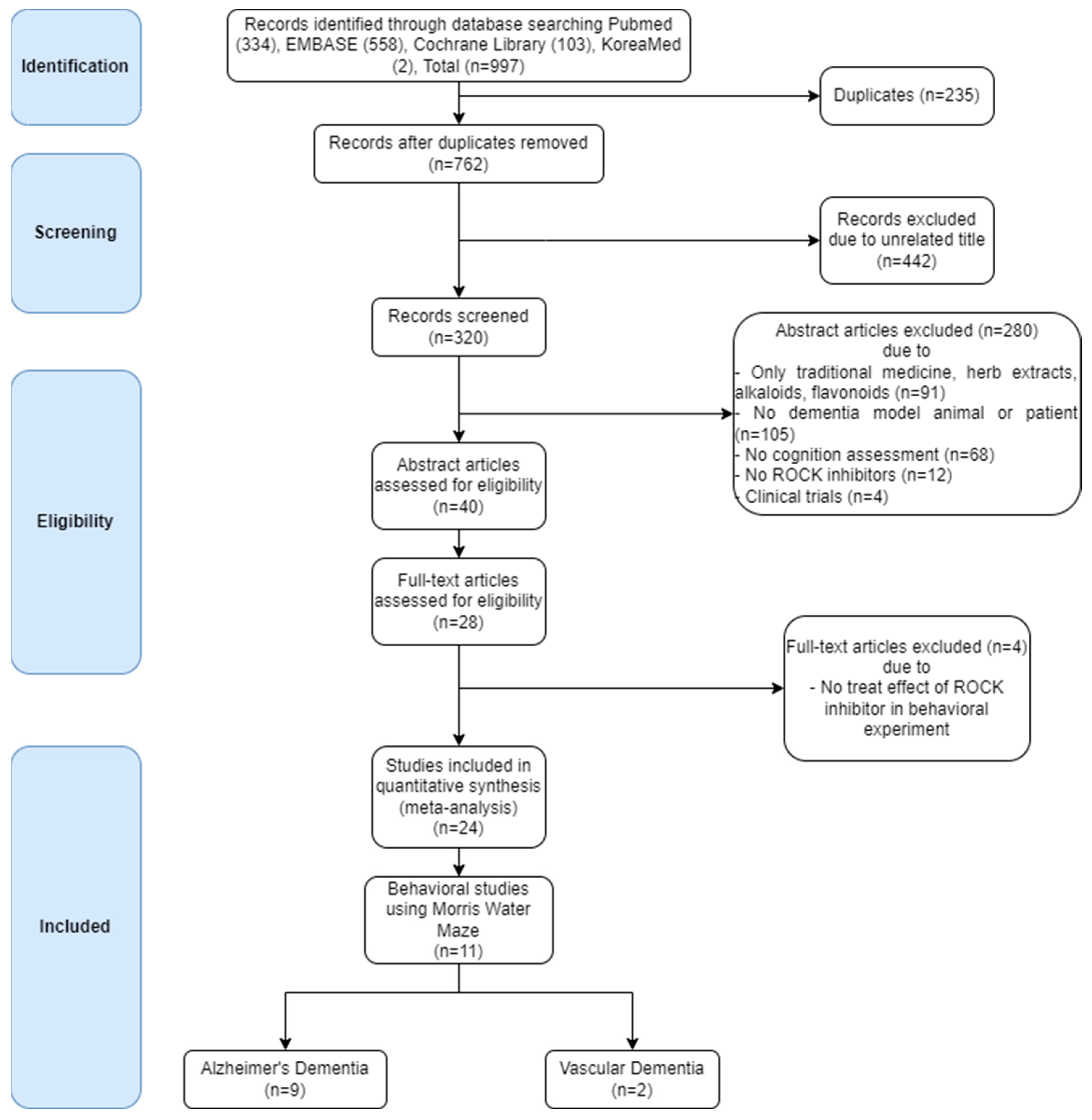
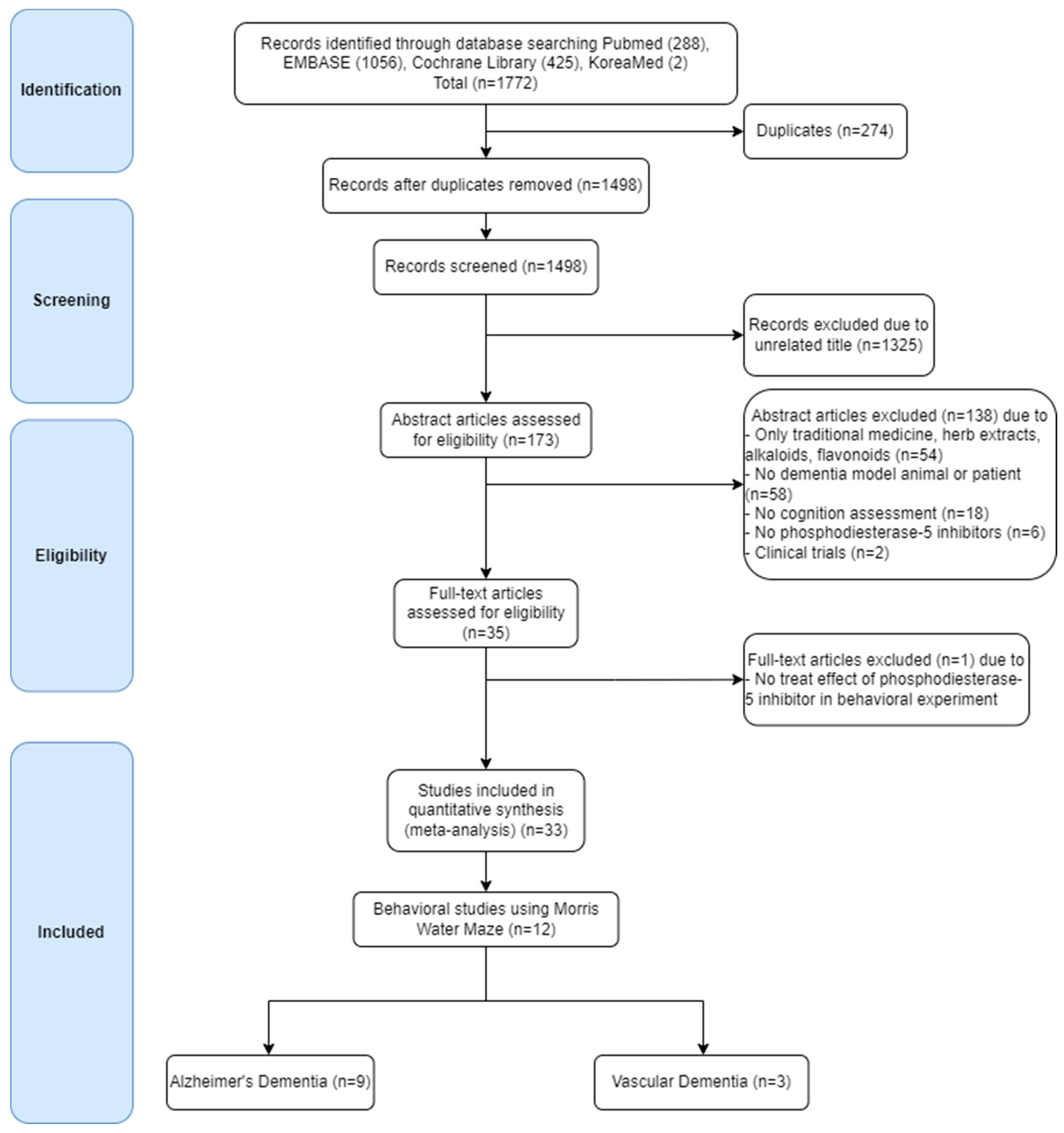
6.1.1. Search Strategy and Selection Criteria
6.1.2. Data Extraction
6.1.3. Data Analysis
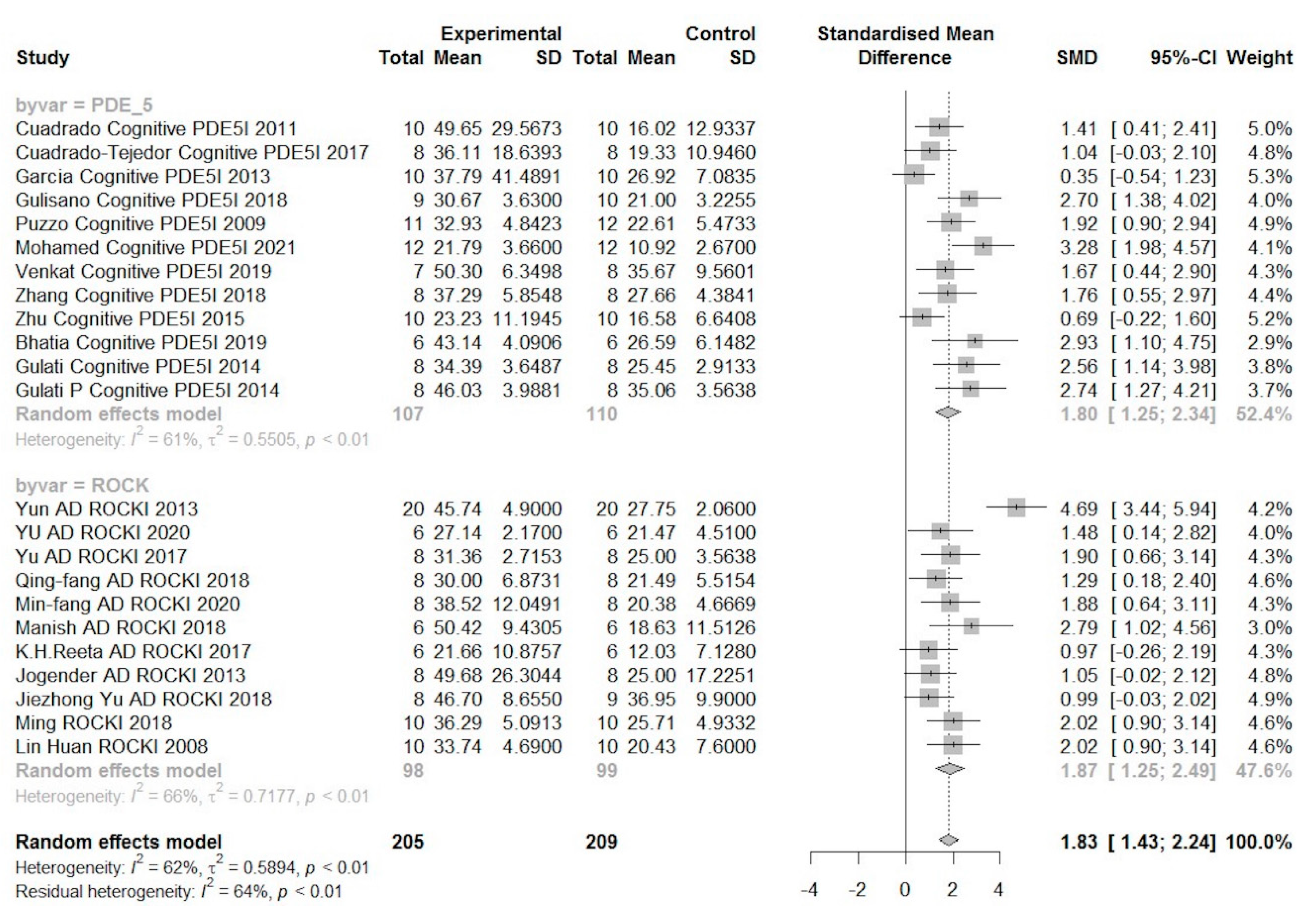
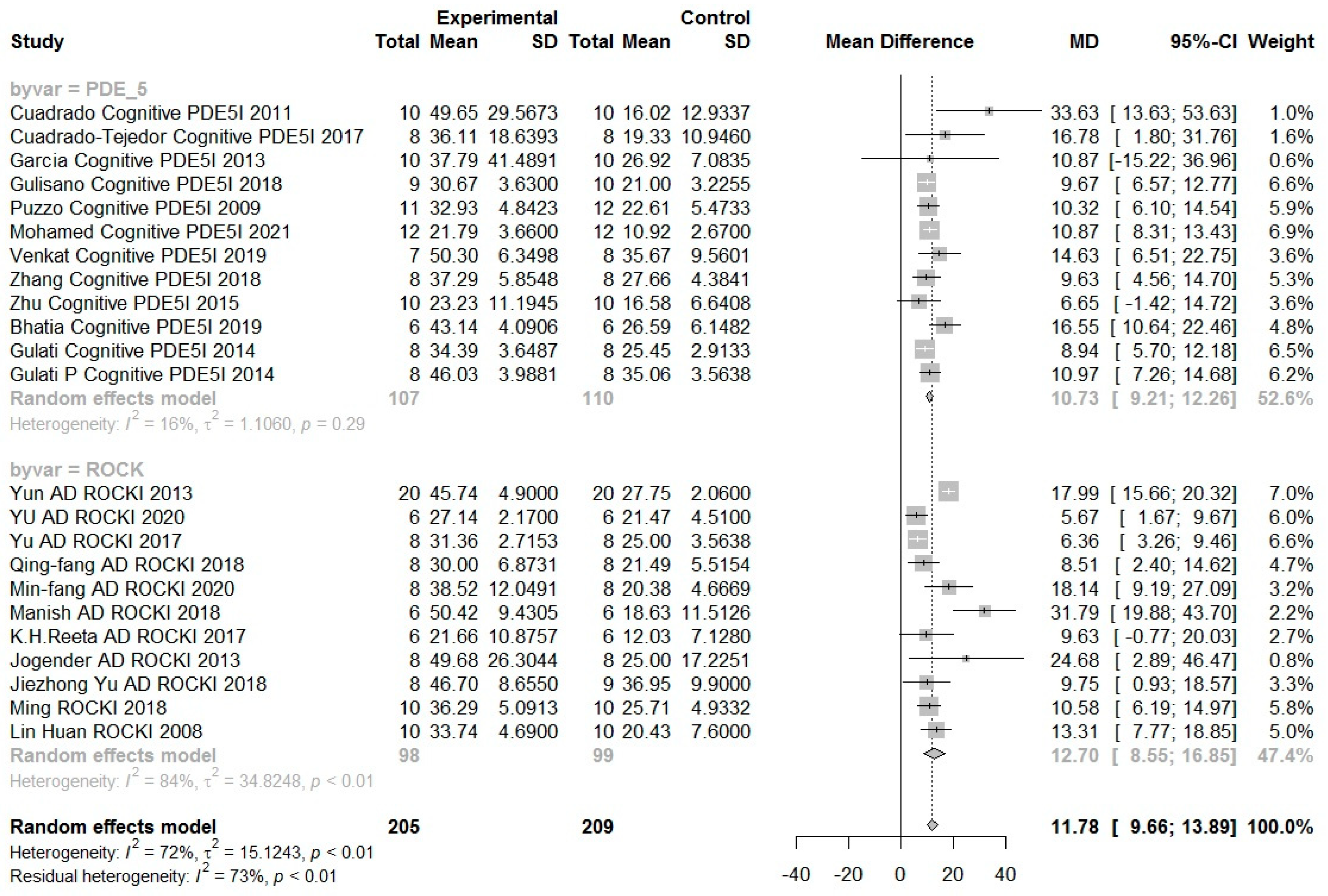
6.2. Results
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gale, S.A.; Acar, D.; Daffner, K.R. Dementia. Am. J. Med. 2018, 131, 1161–1169. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Hou, D.; Lin, F.; Luo, J.; Xie, J.; Wang, Y.; Tian, Y. The role of neurovascular unit damage in the occurrence and development of Alzheimer′s disease. Rev. Neurosci. 2019, 30, 477–484. [Google Scholar] [CrossRef] [PubMed]
- Tackenberg, C.; Kulic, L.; Nitsch, R.M. Familial Alzheimer′s disease mutations at position 22 of the amyloid beta-peptide sequence differentially affect synaptic loss, tau phosphorylation and neuronal cell death in an ex vivo system. PLoS ONE 2020, 15, e0239584. [Google Scholar] [CrossRef] [PubMed]
- Atri, A. The Alzheimer′s disease clinical spectrum: Diagnosis and management. Med. Clin. N. Am. 2019, 103, 263–293. [Google Scholar] [CrossRef]
- Förstl, H.; Kurz, A. Clinical features of Alzheimer’s disease. Eur. Arch. Psychiatry Clin. Neurosci. 1999, 249, 288–290. [Google Scholar] [CrossRef]
- Carlesimo, G.A.; Oscar-Berman, M. Memory deficits in Alzheimer’s patients: A comprehensive review. Neuropsychol. Rev. 1992, 3, 119–169. [Google Scholar] [CrossRef]
- Jelicic, M.; Bonebakker, A.E.; Bonke, B. Implicit memory performance of patients with Alzheimer’s disease: A brief review. Int. Psychogeriatr. 1995, 7, 385–392. [Google Scholar] [CrossRef]
- Taler, V.; Phillips, N.A. Language performance in Alzheimer’s disease and mild cognitive impairment: A comparative review. J. Clin. Exp. Neuropsychol. 2008, 30, 501–556. [Google Scholar] [CrossRef]
- Frank, E.M. Effect of Alzheimer’s disease on communication function. J. S. Carol. Med. Assoc. 1994, 90, 417–423. [Google Scholar]
- Volicer, L.; Harper, D.G.; Manning, B.C.; Goldstein, R.; Satlin, A. Sundowning and circadian rhythms in Alzheimer′s disease. Am. J. Psychiatry 2001, 158, 704–711. [Google Scholar] [CrossRef]
- Cunningham, E.L.; McGuinness, B.; Herron, B.; Passmore, A.P. Dementia. Ulster Med. J. 2015, 84, 79–87. [Google Scholar] [PubMed]
- Karantzoulis, S.; Galvin, J.E. Distinguishing Alzheimer′s disease from other major forms of dementia. Expert Rev. Neurother. 2011, 11, 1579–1591. [Google Scholar] [CrossRef] [PubMed]
- Rajan, K.B.; Weuve, J.; Barnes, L.L.; McAninch, E.A.; Wilson, R.S.; Evans, D.A. Population estimate of people with clinical Alzheimer’s disease and mild cognitive impairment in the United States (2020–2060). Alzheimer’s Dement. 2021, 17, 1966–1975. [Google Scholar] [CrossRef] [PubMed]
- Plassman, B.L.; Langa, K.M.; Fisher, G.G.; Heeringa, S.G.; Weir, D.R.; Ofstedal, M.B.; Burke, J.R.; Hurd, M.D.; Potter, G.G.; Rodgers, W.L.; et al. Prevalence of dementia in the United States: The aging, demographics, and memory study. Neuroepidemiology 2007, 29, 125–132. [Google Scholar] [CrossRef]
- Jorm, A.F.; Korten, A.E.; Henderson, A.S. The prevalence of dementia: A quantitative integration of the literature. Acta Psychiatr. Scand. 1987, 76, 465–479. [Google Scholar] [CrossRef]
- Moore, M.J.; Zhu, C.W.; Clipp, E.C. Informal costs of dementia care: Estimates from the National Longitudinal Caregiver Study. J. Gerontol. Ser. B Psychol. Sci. Soc. Sci. 2001, 56, S219–S228. [Google Scholar] [CrossRef]
- De Deyn, P.P. Treatment of Alzheimer′s disease. N. Engl. J. Med. 2000, 342, 821–822. [Google Scholar]
- Abyadeh, M.; Gupta, V.; Gupta, V.; Chitranshi, N.; Wu, Y.; Amirkhani, A.; Meyfour, A.; Sheriff, S.; Shen, T.; Dhiman, K.; et al. Comparative analysis of aducanumab, zagotenemab and pioglitazone as targeted treatment strategies for Alzheimer′s disease. Aging Dis. 2021, 12, 1964–1976. [Google Scholar] [CrossRef]
- Toyn, J. What lessons can be learned from failed Alzheimer′s disease trials? Expert Rev. Clin. Pharmacol. 2015, 8, 267–269. [Google Scholar] [CrossRef] [Green Version]
- Kang, S.; Lee, Y.-H.; Lee, J.E. Metabolism-centric overview of the pathogenesis of Alzheimer’s disease. Yonsei Med. J. 2017, 58, 479–488. [Google Scholar] [CrossRef]
- Lott, I.T.; Head, E. Dementia in Down syndrome: Unique insights for Alzheimer disease research. Nat. Rev. Neurol. 2019, 15, 135–147. [Google Scholar] [CrossRef] [PubMed]
- Hardy, J.; Allsop, D. Amyloid deposition as the central event in the aetiology of Alzheimer’s disease. Trends Pharmacol. Sci. 1991, 12, 383–388. [Google Scholar] [CrossRef]
- Mudher, A.; Lovestone, S. Alzheimer′s disease-do tauists and baptists finally shake hands? Trends Neurosci. 2002, 25, 22–26. [Google Scholar] [CrossRef]
- Yankner, B.A. Mechanisms of neuronal degeneration in Alzheimer’s Disease. Neuron 1996, 16, 921–932. [Google Scholar] [CrossRef] [Green Version]
- Glenner, G.G.; Wong, C.W. Alzheimer′s disease: Initial report of the purification and characterization of a novel cerebrovascular amyloid protein. Biochem. Biophys. Res. Commun. 1984, 120, 885–890. [Google Scholar] [CrossRef]
- Kang, J.; Lemaire, H.-G.; Unterbeck, A.; Salbaum, J.M.; Masters, C.L.; Grzeschik, K.-H.; Multhaup, G.; Beyreuther, K.; Müller-Hill, B. The precursor of Alzheimer’s disease amyloid A4 protein resembles a cell-surface receptor. Nature 1987, 325, 733–736. [Google Scholar] [CrossRef]
- Masters, C.L.; Simms, G.; Weinman, N.A.; Multhaup, G.; McDonald, B.L.; Beyreuther, K. Amyloid plaque core protein in Alzheimer disease and Down syndrome. Proc. Natl. Acad. Sci. USA 1985, 82, 4245–4249. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, K.; Alonso, A.D.C.; Chen, S.; Chohan, M.O.; El-Akkad, E.; Gong, C.-X.; Khatoon, S.; Li, B.; Liu, F.; Rahman, A.; et al. Tau pathology in Alzheimer disease and other tauopathies. Biochim. Biophys. Acta Mol. Basis Dis. 2005, 1739, 198–210. [Google Scholar] [CrossRef] [Green Version]
- Manzano, S.; Agüera, L.; Aguilar, M.; Olazarán, J. A review on tramiprosate (homotaurine) in Alzheimer’s disease and other neurocognitive disorders. Front. Neurol. 2020, 11, 614. [Google Scholar] [CrossRef]
- Leng, F.; Edison, P. Neuroinflammation and microglial activation in Alzheimer disease: Where do we go from here? Nat. Rev. Neurol. 2021, 17, 157–172. [Google Scholar] [CrossRef]
- Heneka, M.T.; Carson, M.J.; Khoury, J.E.; Landreth, G.E.; Brosseron, F.; Feinstein, D.L.; Jacobs, A.H.; Wyss-Coray, T.; Vitorica, J.; Ransohoff, R.M.; et al. Neuroinflammation in Alzheimer′s disease. Lancet Neurol. 2015, 14, 388–405. [Google Scholar] [CrossRef] [Green Version]
- Uddin, S.; Kabir, T.; Jalouli, M.; Rahman, A.; Jeandet, P.; Behl, T.; Alexiou, A.; Albadrani, G.M.; Abdel-Daim, M.M.; Perveen, A.; et al. Neuroinflammatory signaling in the pathogenesis of Alzheimer′s disease. Curr. Neuropharmacol. 2021, 20, 126–146. [Google Scholar] [CrossRef]
- Kwon, H.S.; Koh, S.-H. Neuroinflammation in neurodegenerative disorders: The roles of microglia and astrocytes. Transl. Neurodegener. 2020, 9, 42. [Google Scholar] [CrossRef] [PubMed]
- Iemolo, F.; Duro, G.; Rizzo, C.; Castiglia, L.; Hachinski, V.; Caruso, C. Pathophysiology of vascular dementia. Immun. Ageing 2009, 6, 13. [Google Scholar] [CrossRef] [Green Version]
- Markus, H.S.; Barley, J.; Lunt, R.; Bland, J.M.; Jeffery, S.; Carter, N.D.; Brown, M.M. Angiotensin-converting enzyme gene deletion polymorphism. A new risk factor for lacunar stroke but not carotid atheroma. Stroke 1995, 26, 1329–1333. [Google Scholar] [CrossRef]
- Lassen, N.A. Incomplete cerebral infarction—Focal incomplete ischemic tissue necrosis not leading to emollision. Stroke 1982, 13, 522–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia, J.H.; Lassen, N.A.; Weiller, C.; Sperling, B.; Nakagawara, J. Ischemic stroke and incomplete infarction. Stroke 1996, 27, 761–765. [Google Scholar] [CrossRef] [PubMed]
- Sakaguchi, T.; Takefuji, M.; Wettschureck, N.; Hamaguchi, T.; Amano, M.; Kato, K.; Tsuda, T.; Eguchi, S.; Ishihama, S.; Mori, Y.; et al. Protein kinase n promotes stress-induced cardiac dysfunction through phosphorylation of myocardin-related transcription factor A and disruption of its interaction with actin. Circulation 2019, 140, 1737–1752. [Google Scholar] [CrossRef] [PubMed]
- Kimura, K.; Ito, M.; Amano, M.; Chihara, K.; Fukata, Y.; Nakafuku, M.; Yamamori, B.; Feng, J.; Nakano, T.; Okawa, K.; et al. Regulation of myosin phosphatase by Rho and Rho-associated kinase (Rho-kinase). Science 1996, 273, 245–248. [Google Scholar] [CrossRef] [PubMed]
- Amano, M.; Ito, M.; Kimura, K.; Fukata, Y.; Chihara, K.; Nakano, T.; Matsuura, Y.; Kaibuchi, K. Phosphorylation and activation of myosin by Rho-associated kinase (Rho-kinase). J. Biol. Chem. 1996, 271, 20246–20249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahbazi, R.; Baradaran, B.; Khordadmehr, M.; Safaei, S.; Baghbanzadeh, A.; Jigari, F.; Ezzati, H. Targeting ROCK signaling in health, malignant and non-malignant diseases. Immunol. Lett. 2020, 219, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Secinti, B.B.; Tatar, G.; Tok, T.T. Determination of potential selective inhibitors for ROCKI and ROCKII isoforms with molecular modeling techniques: Structure based docking, ADMET and molecular dynamics simulation. J. Biomol. Struct. Dyn. 2019, 37, 2457–2463. [Google Scholar] [CrossRef] [PubMed]
- Guazzi, M.; Vicenzi, M.; Arena, R.; Guazzi, M.D. PDE5 inhibition with sildenafil improves left ventricular diastolic function, cardiac geometry, and clinical status in patients with stable systolic heart failure: Results of a 1-year, prospective, randomized, placebo-controlled study. Circ. Heart Fail. 2011, 4, 8–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nave, A.H.; Mižíková, I.; Niess, G.; Steenbock, H.; Reichenberger, F.; Talavera, M.L.; Veit, F.; Herold, S.; Mayer, K.; Vadász, I.; et al. Lysyl oxidases play a causal role in vascular remodeling in clinical and experimental pulmonary arterial hypertension. Arter. Thromb. Vasc. Biol. 2014, 34, 1446–1458. [Google Scholar] [CrossRef] [Green Version]
- Hahmann, C.; Schroeter, T. Rho-kinase inhibitors as therapeutics: From pan inhibition to isoform selectivity. Cell. Mol. Life Sci. 2010, 67, 171–177. [Google Scholar] [CrossRef]
- Riento, K.; Ridley, A.J. ROCKs: Multifunctional kinases in cell behaviour. Nat. Rev. Mol. Cell Biol. 2003, 4, 446–456. [Google Scholar] [CrossRef]
- Lu, W.; Wen, J.; Chen, Z. Distinct roles of ROCK1 and ROCK2 on the cerebral ischemia injury and subsequently neurodegenerative changes. Pharmacology 2020, 105, 3–8. [Google Scholar] [CrossRef]
- Kloc, M.; Uosef, A.; Villagran, M.; Zdanowski, R.; Kubiak, J.Z.; Wosik, J.; Ghobrial, R.M. Ghobrial, RhoA- and Actin-Dependent functions of macrophages from the rodent cardiac transplantation model perspective—Timing is the essence. Biology 2021, 10, 70. [Google Scholar] [CrossRef]
- Magalhaes, Y.T.; Farias, J.O.; Silva, L.E.; Forti, F.L. GTPases, genome, actin: A hidden story in DNA damage response and repair mechanisms. DNA Repair 2021, 100, 103070. [Google Scholar] [CrossRef]
- Herskowitz, J.H.; Seyfried, N.T.; Gearing, M.; Kahn, R.A.; Peng, J.; Levey, A.I.; Lah, J.J. Rho kinase II phosphorylation of the lipoprotein receptor LR11/SORLA alters amyloid-beta production. J. Biol Chem. 2011, 286, 6117–6127. [Google Scholar] [CrossRef] [Green Version]
- Weber, A.J.; Herskowitz, J.H. Perspectives on ROCK2 as a therapeutic target for Alzheimer′s Disease. Front. Cell. Neurosci. 2021, 15, 636017. [Google Scholar] [CrossRef] [PubMed]
- Zheng, K.; Hu, F.; Zhou, Y.; Zhang, J.; Zheng, J.; Lai, C.; Xiong, W.; Cui, K.; Hu, Y.Z.; Han, Z.T.; et al. miR-135a-5p mediates memory and synaptic impairments via the Rock2/Adducin1 signaling pathway in a mouse model of Alzheimer′s disease. Nat. Commun. 2021, 12, 1903. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Pallares, J.; Rodriguez-Perez, A.I.; Muñoz, A.; Parga, J.A.; Toledo-Aral, J.J.; Labandeira-Garcia, J.L. Effects of rho kinase inhibitors on grafts of dopaminergic cell precursors in a rat model of Parkinson’s disease. Stem Cells Transl. Med. 2016, 5, 804–815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sherrington, R.; Rogaev, E.I.; Liang, Y.; Rogaeva, E.A.; Levesque, G.; Ikeda, M.; Chi, H.; Lin, C.; Li, G.; Holman, K.; et al. Cloning of a gene bearing missense mutations in early-onset familial Alzheimer′s disease. Nature 1995, 375, 754–760. [Google Scholar] [CrossRef] [PubMed]
- Bekris, L.M.; Yu, C.E.; Bird, T.D.; Tsuang, D.W. Genetics of Alzheimer disease. J. Geriatr. Psychiatry Neurol. 2010, 23, 213–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedrini, S.; Carter, T.L.; Prendergast, G.; Petanceska, S.; Ehrlich, M.E.; Gandy, S. Modulation of statin-activated shedding of alzheimer APP ectodomain by ROCK. PLoS Med. 2005, 2, e18. [Google Scholar] [CrossRef]
- Hamano, T.; Shirafuji, N.; Yen, S.-H.; Yoshida, H.; Kanaan, N.M.; Hayashi, K.; Ikawa, M.; Yamamura, O.; Fujita, Y.; Kuriyama, M.; et al. Rho-kinase ROCK inhibitors reduce oligomeric tau protein. Neurobiol. Aging 2020, 89, 41–54. [Google Scholar] [CrossRef]
- Somlyo, A.P.; Somlyo, A.V. Signal transduction and regulation in smooth muscle. Nature 1994, 372, 231–236. [Google Scholar] [CrossRef]
- Laufs, U.; Liao, J.K. Post-transcriptional Regulation of Endothelial Nitric Oxide Synthase mRNA Stability by Rho GTPase. J. Biol. Chem. 1998, 273, 24266–24271. [Google Scholar] [CrossRef] [Green Version]
- Lee, E.C.; Hong, D.-Y.; Lee, D.-H.; Park, S.-W.; Lee, J.Y.; Jeong, J.H.; Kim, E.-Y.; Chung, H.-M.; Hong, K.-S.; Park, S.-P.; et al. Inflammation and rho-associated protein kinase-induced brain changes in vascular dementia. Biomedicines 2022, 10, 446. [Google Scholar] [CrossRef]
- Aizawa, T.; Wei, H.; Miano, J.M.; Abe, J.I.; Berk, B.C.; Yan, C. Role of phosphodiesterase 3 in NO/cGMP-mediated anti-inflammatory effects in vascular smooth muscle cells. Circ. Res. 2003, 93, 406–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamatovic, S.M.; Keep, R.; Kunkel, S.L.; Andjelkovic, A.V. Potential role of MCP-1 in endothelial cell tight junction ‘opening‘: Signaling via Rho and Rho kinase. J. Cell Sci. 2003, 116, 4615–4628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teramoto, H.; Salem, P.; Robbins, K.C.; Bustelo, X.R.; Gutkind, J.S. Tyrosine phosphorylation of the vav proto-oncogene product links FcepsilonRI to the Rac1-JNK pathway. J. Biol. Chem. 1997, 272, 10751–10755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perona, R.; Montaner, S.; Saniger, L.; Sanchez-Perez, I.; Bravo, R.; Lacal, J.C. Activation of the nuclear factor-kappaB by Rho, CDC42, and Rac-1 proteins. Genes Dev. 1997, 11, 463–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biswas, P.S.; Gupta, S.; Chang, E.; Song, L.; Stirzaker, R.A.; Liao, J.K.; Bhagat, G.; Pernis, A.B. Phosphorylation of IRF4 by ROCK2 regulates IL-17 and IL-21 production and the development of autoimmunity in mice. J. Clin. Investig. 2010, 120, 3280–3295. [Google Scholar] [CrossRef] [Green Version]
- Jin, R.; Yang, G.; Li, G. Inflammatory mechanisms in ischemic stroke: Role of inflammatory cells. J. Leukoc. Biol. 2010, 87, 779–789. [Google Scholar] [CrossRef] [Green Version]
- Satoh, K.; Satoh, T.; Kikuchi, N.; Omura, J.; Kurosawa, R.; Suzuki, K.; Sugimura, K.; Aoki, T.; Nochioka, K.; Tatebe, S.; et al. Basigin mediates pulmonary hypertension by promoting inflammation and vascular smooth muscle cell proliferation. Circ. Res. 2014, 115, 738–750. [Google Scholar] [CrossRef] [Green Version]
- Damsker, J.M.; Bukrinsky, M.I.; Constant, S.L. Preferential chemotaxis of activated human CD4+ T cells by extracellular cyclophilin A. J. Leukoc. Biol. 2007, 82, 613–618. [Google Scholar] [CrossRef] [Green Version]
- Katsumata, N.; Shimokawa, H.; Seto, M.; Kozai, T.; Yamawaki, T.; Kuwata, K.; Egashira, K.; Ikegaki, I.; Asano, T.; Sasaki, Y.; et al. Enhanced myosin light chain phosphorylations as a central mechanism for coronary artery spasm in a swine model with interleukin-1beta. Circulation 1997, 96, 4357–4363. [Google Scholar] [CrossRef]
- Peixoto, C.A.; Nunes, A.K.S.; Garcia-Osta, A. Phosphodiesterase-5 inhibitors: Action on the signaling pathways of neuroinflammation, neurodegeneration, and cognition. Mediators Inflamm. 2015, 2015, 940207. [Google Scholar] [CrossRef] [Green Version]
- Schlossmann, J.; Schinner, E. cGMP becomes a drug target. Naunyn Schmiedebergs Arch. Pharmacol. 2012, 385, 243–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Y.; Huang, Y.; Mai, C.; Pan, H.; Luo, H.-B.; Liu, L.; Xie, Y. The immunomodulatory role of PDEs inhibitors in immune cells: Therapeutic implication in rheumatoid arthritis. Pharmacol. Res. 2020, 161, 105134. [Google Scholar] [CrossRef] [PubMed]
- Schäfer, A.; Fraccarollo, D.; Werner, L.; Bauersachs, J. Guanylyl cyclase activator ataciguat improves vascular function and reduces platelet activation in heart failure. Pharmacol. Res. 2010, 62, 432–438. [Google Scholar] [CrossRef] [PubMed]
- Yao, M.; Roberts, D.D.; Isenberg, J.S. Thrombospondin-1 inhibition of vascular smooth muscle cell responses occurs via modulation of both cAMP and cGMP. Pharmacol. Res. 2011, 63, 13–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Sun, X.; Li, Z.; Liu, Y. Inhibition of cGMP phosphodiesterase 5 suppresses serotonin signalling in pulmonary artery smooth muscles cells. Pharmacol. Res. 2009, 59, 312–318. [Google Scholar] [CrossRef] [PubMed]
- Rohini, A.; Agrawal, N.; Koyani, C.N.; Singh, R. Molecular targets and regulators of cardiac hypertrophy. Pharmacol. Res. 2010, 61, 269–280. [Google Scholar] [CrossRef]
- Francis, S.H.; Blount, M.A.; Corbin, J.D. Mammalian cyclic nucleotide phosphodiesterases: Molecular mechanisms and physiological functions. Physiol. Rev. 2011, 91, 651–690. [Google Scholar] [CrossRef] [Green Version]
- Baillie, G.S.; Tejeda, G.S.; Kelly, M.P. Therapeutic targeting of 3′,5′-cyclic nucleotide phosphodiesterases: Inhibition and beyond. Nat. Rev. Drug. Discov. 2019, 18, 770–796. [Google Scholar] [CrossRef]
- Bhat, A.; Ray, B.; Mahalakshmi, A.M.; Tuladhar, S.; Nandakumar, D.; Srinivasan, M.; Essa, M.M.; Chidambaram, S.B.; Guillemin, G.J.; Sakharkar, M.K. Phosphodiesterase-4 enzyme as a therapeutic target in neurological disorders. Pharmacol. Res. 2020, 160, 105078. [Google Scholar] [CrossRef]
- Malinski, T. Nitric Oxide and Nitroxidative Stress in Alzheimer’s Disease. J. Alzheimer’s Dis. 2007, 11, 207–218. [Google Scholar] [CrossRef]
- Puzzo, D.; Vitolo, O.; Trinchese, F.; Jacob, J.P.; Palmeri, A.; Arancio, O. Amyloid-beta peptide inhibits activation of the nitric oxide/cGMP/cAMP-responsive element-binding protein pathway during hippocampal synaptic plasticity. J. Neurosci. 2005, 25, 6887–6897. [Google Scholar] [CrossRef]
- Acquarone, E.; Argyrousi, E.K.; Berg, M.V.D.; Gulisano, W.; Fà, M.; Staniszewski, A.; Calcagno, E.; Zuccarello, E.; D′Adamio, L.; Deng, S.-X.; et al. Synaptic and memory dysfunction induced by tau oligomers is rescued by up-regulation of the nitric oxide cascade. Mol. Neurodegener. 2019, 14, 1–19. [Google Scholar] [CrossRef]
- Law, A.; Gauthier, S.; Quirion, R. Say NO to Alzheimer′s disease: The putative links between nitric oxide and dementia of the Alzheimer′s type. Brain. Res. Rev. 2001, 35, 73–96. [Google Scholar] [CrossRef]
- Austin, S.A.; Santhanam, A.V.; Hinton, D.J.; Choi, D.-S.; Katusic, Z.S. Endothelial nitric oxide deficiency promotes Alzheimer’s disease pathology. J. Neurochem. 2013, 127, 691–700. [Google Scholar] [CrossRef] [Green Version]
- Austin, S.A.; Santhanam, A.V.; Katusic, Z.S. Endothelial nitric oxide modulates expression and processing of amyloid precursor protein. Circ. Res. 2010, 107, 1498–1502. [Google Scholar] [CrossRef] [Green Version]
- Colton, C.A.; Vitek, M.P.; Wink, D.A.; Xu, Q.; Cantillana, V.; Previti, M.L.; Van Nostrand, W.E.; Weinberg, J.B.; Dawson, H. NO synthase 2 (NOS2) deletion promotes multiple pathologies in a mouse model of Alzheimer’s disease. Proc. Natl. Acad. Sci. USA 2006, 103, 12867–12872. [Google Scholar] [CrossRef] [Green Version]
- Ugarte, A.; Gil-Bea, F.; García-Barroso, C.; Cedazo-Minguez, Á.; Ramírez, M.J.; Franco, R.; Garcia-Osta, A.; Oyarzabal, J.; Cuadrado-Tejedor, M. Decreased levels of guanosine 3′, 5′-monophosphate (cGMP) in cerebrospinal fluid (CSF) are associated with cognitive decline and amyloid pathology in Alzheimer′s disease. Neuropathol. Appl. Neurobiol. 2015, 41, 471–482. [Google Scholar] [CrossRef]
- Xu, L.L.; Warren, M.K.; Rose, W.L.; Gong, W.; Wang, J.M. Human recombinant monocyte chemotactic protein and other c-c chemokines bind and induce directional migration of dendritic cells in vitro. J. Leukoc. Biol. 1996, 60, 365–371. [Google Scholar] [CrossRef]
- Zeiher, A.M.; Fisslthaler, B.; Schray-Utz, B.; Busse, R. Nitric oxide modulates the expression of monocyte chemoattractant protein 1 in cultured human endothelial cells. Circ. Res. 1995, 76, 980–986. [Google Scholar] [CrossRef]
- Figley, C.R.; Stroman, P.W. The role(s) of astrocytes and astrocyte activity in neurometabolism, neurovascular coupling, and the production of functional neuroimaging signals. Eur. J. Neurosci. 2011, 33, 577–588. [Google Scholar] [CrossRef]
- Santello, M.; Volterra, A. TNFalpha in synaptic function: Switching gears. Trends Neurosci. 2012, 35, 638–647. [Google Scholar] [CrossRef]
- Freeman, M.R.; Rowitch, D.H. Evolving concepts of gliogenesis: A Look Way Back and Ahead to the Next 25 Years. Neuron 2013, 80, 613–623. [Google Scholar] [CrossRef] [Green Version]
- Borán, M.S.; Baltrons, M.A.; García, A. The ANP-cGMP-protein kinase G pathway induces a phagocytic phenotype but decreases inflammatory gene expression in microglial cells. Glia 2008, 56, 394–411. [Google Scholar] [CrossRef]
- Shin, C.Y.; Lee, W.J.; Choi, J.W.; Choi, M.S.; Ryu, J.R.; Oh, S.J.; Cheong, J.H.; Choi, E.Y.; Ko, K.H. Down-regulation of matrix metalloproteinase-9 expression by nitric oxide in lipopolysaccharide-stimulated rat primary astrocytes. Nitric Oxide 2007, 16, 425–432. [Google Scholar] [CrossRef]
- Tonges, L.; Frank, T.; Tatenhorst, L.; Saal, K.A.; Koch, J.C.; Szego, E.M.; Bahr, M.; Weishaupt, J.H.; Lingor, P. Inhibition of rho kinase enhances survival of dopaminergic neurons and attenuates axonal loss in a mouse model of Parkinson’s disease. Brain 2012, 135, 3355–3370. [Google Scholar] [CrossRef]
- Yamashita, K.; Kotani, Y.; Nakajima, Y.; Shimazawa, M.; Yoshimura, S.-I.; Nakashima, S.; Iwama, T.; Hara, H. Fasudil, a Rho kinase (ROCK) inhibitor, protects against ischemic neuronal damage in vitro and in vivo by acting directly on neurons. Brain Res. 2007, 1154, 215–224. [Google Scholar] [CrossRef]
- Koch, J.C.; Tönges, L.; Barski, E.; Michel, U.; Bahr, M.; Lingor, P. ROCK2 is a major regulator of axonal degeneration, neuronal death and axonal regeneration in the CNS. Cell Death Dis. 2014, 5, e1225. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.W.; Liu, P.Y.; Oyama, N.; Rikitake, Y.; Kitamoto, S.; Gitlin, J.; Liao, J.K.; Boisvert, W.A. Deficiency of ROCK1 in bone marrow-derived cells protects against atherosclerosis in LDLR-/- mice. FASEB J. 2008, 22, 3561–3570. [Google Scholar] [CrossRef] [Green Version]
- Matoba, K.; Kawanami, D.; Ishizawa, S.; Kanazawa, Y.; Yokota, T.; Utsunomiya, K. Rho-kinase mediates TNF-α-induced MCP-1 expression via p38 MAPK signaling pathway in mesangial cells. Biochem. Biophys. Res. Commun. 2010, 402, 725–730. [Google Scholar] [CrossRef]
- Yu, Y.H.; Kim, S.W.; Kang, J.; Song, Y.; Im, H.; Kim, S.J.; Yoo, D.Y.; Lee, M.R.; Park, D.K.; Oh, J.S.; et al. Phosphodiesterase-5 inhibitor attenuates anxious phenotypes and movement disorder induced by mild ischemic stroke in rats. J. Korean Neurosurg. Soc. 2022. [Google Scholar] [CrossRef]
- Song, Y.; Chen, X.; Wang, L.Y.; Gao, W.; Zhu, M.J. Rho kinase inhibitor fasudil protects against beta-amyloid-induced hippocampal neurodegeneration in rats. CNS Neurosci. Ther. 2013, 19, 603–610. [Google Scholar] [CrossRef]
- Yu, N.; Huang, Y.; Jiang, Y.; Zou, L.; Liu, X.; Liu, S.; Chen, F.; Luo, J.; Zhu, Y. Ganoderma lucidum triterpenoids (GLTs) reduce neuronal apoptosis via inhibition of ROCK signal pathway in APP/PS1 transgenic alzheimer’s disease mice. Oxidative Med. Cell. Longev. 2020, 2020, 9894037. [Google Scholar] [CrossRef]
- Yu, J.-Z.; Li, Y.-H.; Liu, C.-Y.; Wang, Q.; Gu, Q.-F.; Wang, H.-Q.; Zhang, G.-X.; Xiao, B.-G.; Ma, C.-G. Multitarget Therapeutic Effect of Fasudil in APP/PS1transgenic Mice. CNS Neurol. Disord.—Drug Targets 2017, 16, 199–209. [Google Scholar] [CrossRef]
- Gu, Q.; Yu, J.; Wu, H.; Li, Y.; Liu, C.-Y.; Feng, L.; Zhang, G.; Xiao, B.; Ma, C. Therapeutic effect of Rho kinase inhibitor FSD-C10 in a mouse model of Alzheimer’s disease. Exp. Ther. Med. 2018, 16, 3929–3938. [Google Scholar] [CrossRef] [Green Version]
- Guo, M.F.; Zhang, H.Y.; Li, Y.H.; Gu, Q.F.; Wei, W.Y.; Wang, Y.Y.; Zhang, X.J.; Liu, X.Q.; Song, L.J.; Chai, Z.; et al. Fasudil inhibits the activation of microglia and astrocytes of transgenic Alzheimer′s disease mice via the downregulation of TLR4/Myd88/NF-kappaB pathway. J. Neuroimmunol. 2020, 346, 577284. [Google Scholar] [CrossRef]
- Kumar, M.; Bansal, N. Fasudil hydrochloride ameliorates memory deficits in rat model of streptozotocin-induced Alzheimer′s disease: Involvement of PI3-kinase, eNOS and NFkappaB. Behav. Brain. Res. 2018, 351, 4–16. [Google Scholar] [CrossRef]
- Reeta, K.H.; Singh, D.; Gupta, Y.K. Edaravone attenuates intracerebroventricular streptozotocin-induced cognitive impairment in rats. Eur. J. Neurosci. 2017, 45, 987–997. [Google Scholar] [CrossRef]
- Mehla, J.; Pahuja, M.; Gupta, P.; Dethe, S.; Agarwal, A.; Gupta, Y.K. Clitoria ternatea ameliorated the intracerebroventricularly injected streptozotocin induced cognitive impairment in rats: Behavioral and biochemical evidence. Psychopharmacology 2013, 230, 589–605. [Google Scholar] [CrossRef]
- Yu, J.; Yan, Y.; Gu, Q.; Kumar, G.; Yu, H.; Zhao, Y.; Liu, C.; Gao, Y.; Chai, Z.; Chumber, J.; et al. Fasudil in combination with bone marrow stromal cells (BMSCs) attenuates Alzheimer′s disease-related changes through the regulation of the peripheral immune system. Front. Aging Neurosci. 2018, 10, 216. [Google Scholar] [CrossRef] [Green Version]
- Ni, M.; Zhang, J.; Huang, L.; Liu, G.; Li, Q. A Rho-kinase inhibitor reverses learning and memory deficits in a Rat model of chronic cerebral ischemia by altering Bcl-2/Bax-NMDAR signaling in the cerebral cortex. J. Pharmacol. Sci. 2018, 138, 107–115. [Google Scholar] [CrossRef]
- Huang, L.; He, Z.; Guo, L.; Wang, H. Improvement of cognitive deficit and neuronal damage in rats with chronic cerebral ischemia via relative long-term inhibition of rho-kinase. Cell. Mol. Neurobiol. 2008, 28, 757–768. [Google Scholar] [CrossRef]
- Cuadrado-Tejedor, M.; Hervias, I.; Ricobaraza, A.; Puerta, E.; Pérez-Roldán, J.M.; García-Barroso, C.; Franco, R.; Aguirre, N.; Garcia-Osta, A. Sildenafil restores cognitive function without affecting beta-amyloid burden in a mouse model of Alzheimer′s disease. Br. J. Pharmacol. 2011, 164, 2029–2041. [Google Scholar] [CrossRef] [Green Version]
- Cuadrado-Tejedor, M.; Garcia-Barroso, C.; Sanchez-Arias, J.A.; Rabal, O.; González, M.P.; Mederos, S.; Ugarte, A.; Franco, R.; Segura, V.; Perea, G.; et al. A First-in-class small-molecule that acts as a dual inhibitor of HDAC and PDE5 and that rescues hippocampal synaptic impairment in Alzheimer′s disease mice. Neuropsychopharmacology 2017, 42, 524–539. [Google Scholar] [CrossRef] [Green Version]
- García-Barroso, C.; Ricobaraza, A.; Pascual-Lucas, M.; Unceta, N.; Rico, A.J.; Goicolea, M.A.; Sallés, J.; Lanciego, J.L.; Oyarzabal, J.; Franco, R.; et al. Tadalafil crosses the blood–brain barrier and reverses cognitive dysfunction in a mouse model of AD. Neuropharmacology 2013, 64, 114–123. [Google Scholar] [CrossRef]
- Gulisano, W.; Tropea, M.R.; Arancio, O.; Palmeri, A.; Puzzo, D. Sub-efficacious doses of phosphodiesterase 4 and 5 inhibitors improve memory in a mouse model of Alzheimer’s disease. Neuropharmacology 2018, 138, 151–159. [Google Scholar] [CrossRef]
- Puzzo, D.; Staniszewski, A.; Deng, S.X.; Privitera, L.; Leznik, E.; Liu, S.; Zhang, H.; Feng, Y.; Palmeri, A.; Laundry, D.W.; et al. Phosphodiesterase 5 inhibition improves synaptic function, memory, and amyloid-beta load in an Alzheimer′s disease mouse model. J. Neurosci. 2009, 29, 8075–8086. [Google Scholar] [CrossRef]
- Salem, M.; Budzyńska, B.; Kowalczyk, J.; El Sayed, N.S.; Mansour, S.M. Tadalafil and bergapten mitigate streptozotocin-induced sporadic Alzheimer′s disease in mice via modulating neuroinflammation, PI3K/Akt, Wnt/beta-catenin, AMPK/mTOR signaling pathways. Toxicol. Appl. Pharmacol. 2021, 429, 115697. [Google Scholar] [CrossRef]
- Venkat, P.; Chopp, M.; Zacharek, A.; Cui, C.; Landschoot-Ward, J.; Qian, Y.; Chen, Z.; Chen, J. Sildenafil treatment of vascular dementia in aged rats. Neurochem. Int. 2019, 127, 103–112. [Google Scholar] [CrossRef]
- ZZhang, L.; Seo, J.H.; Li, H.; Nam, G.; Yang, H.O. The phosphodiesterase 5 inhibitor, KJH-1002, reverses a mouse model of amnesia by activating a cGMP/cAMP response element binding protein pathway and decreasing oxidative damage. J. Cereb. Blood Flow Metab. 2018, 175, 3347–3360. [Google Scholar]
- Zhu, L.; Yang, J.-Y.; Xue, X.; Dong, Y.-X.; Liu, Y.; Miao, F.-R.; Wang, Y.-F.; Xue, H.; Wu, C.-F. A novel phosphodiesterase-5 Inhibitor: Yonkenafil modulates neurogenesis, gliosis to improve cognitive function and ameliorates amyloid burden in an APP/PS1 transgenic mice model. Mech. Ageing Dev. 2015, 150, 34–45. [Google Scholar] [CrossRef]
- Bhatia, P.; Singh, N. Ameliorative effect of phosphodiesterase-5 inhibitor in rat model of vascular dementia. Curr. Neurovascular. Res. 2019, 16, 27–39. [Google Scholar] [CrossRef]
- Gulati, P.; Singh, N. Neuroprotective effect of tadalafil, a PDE-5 inhibitor, and its modulation by L-NAME in mouse model of ischemia-reperfusion injury. J. Surg. Res. 2014, 186, 475–483. [Google Scholar] [CrossRef]
- Gulati, P.; Singh, N. Tadalafil enhances the neuroprotective effects of ischemic postconditioning in mice, probably in a nitric oxide associated manner. Can. J. Physiol. Pharmacol. 2014, 92, 418–426. [Google Scholar] [CrossRef]
- D′Hooge, R.; De Deyn, P.P. Applications of the Morris water maze in the study of learning and memory. Brain Res. Rev. 2001, 36, 60–90. [Google Scholar] [CrossRef]
- Rogers, D.C.; Hunter, A. Photothrombotic lesions of the rat cortex impair acquisition of the water maze. Pharmacol. Biochem. Behav. 1997, 56, 747–754. [Google Scholar] [CrossRef]
- Uchiyama-Tsuyuki, Y.; Kawashima, K.; Araki, H.; Otomo, S. Prostacyclin analogue TTC-909 reduces memory impairment in rats with cerebral embolism. Pharmacol. Biochem. Behav. 1995, 52, 555–559. [Google Scholar] [CrossRef]
- Yonemori, F.; Yamada, H.; Yamaguchi, T.; Uemura, A.; Tamura, A. spatial memory disturbance after focal cerebral ischemia in rats. J. Cereb. Blood Flow Metab. 1996, 16, 973–980. [Google Scholar] [CrossRef] [Green Version]
- Lyden, P.; Zivin, J.; Chabolla, D.; Jacobs, M.; Gage, F. Quantitative effects of cerebral infarction on spatial learning in rats. Exp. Neurol. 1992, 116, 122–132. [Google Scholar] [CrossRef]
- Block, F. Global ischemia and behavioural deficits. Prog. Neurobiol. 1999, 58, 279–295. [Google Scholar] [CrossRef]
- Smith, D.H.; Soares, H.D.; Pierce, J.S.; Perlman, K.G.; Saatman, K.E.; Meaney, D.F.; Dixon, C.E.; McIntosh, T.K. A Model of parasagittal controlled cortical impact in the mouse: Cognitive and histopathologic effects. J. Neurotrauma 1995, 12, 169–178. [Google Scholar] [CrossRef] [Green Version]
- Dell′Anna, E.; Iuvone, L.; Calzolari, S.; Geloso, M.C. Effect of acetyl-l-carnitine on hyperactivity and spatial memory deficits of rats exposed to neonatal anoxia. Neurosci. Lett. 1997, 223, 201–205. [Google Scholar] [CrossRef]
- Cremer, H.; Lange, R.; Christoph, A.; Plomann, M.; Vopper, G.; Roes, J.; Brown, R.; Baldwin, S.; Kraemer, P.; Scheff, S.; et al. Inactivation of the N-CAM gene in mice results in size reduction of the olfactory bulb and deficits in spatial learning. Nature 1994, 367, 455–459. [Google Scholar] [CrossRef]
- Fransen, E.; D′Hooge, R.; Van Camp, G.; Verhoye, M.; Sijbers, M.; Reyniers, E.; Soriano, P.; Kamiguchi, H.; Willemsen, R.; Koekkoek, S.K.E.; et al. L1 knockout mice show dilated ventricles, vermis hypoplasia and impaired exploration patterns. Hum. Mol. Genet. 1998, 7, 999–1009. [Google Scholar] [CrossRef]
- D′Hooge, R.; Marescau, B.; Qureshi, I.A.; De Deyn, P.P. Impaired cognitive performance in ornithine transcarbamylase-deficient mice on arginine-free diet. Brain Res. 2000, 876, 1–9. [Google Scholar] [CrossRef]
- Vorhees, C.V.; Acuff-Smith, K.D.; Weisenburger, W.P.; Minck, D.R.; Berry, H.K. Branched chain amino acids improve radial-arm maze acquisition and water maze forced-choice learning in rat offspring exposed in utero to hyperphenylalaninemia. Neurotoxicol. Teratol. 1992, 14, 35–41. [Google Scholar] [CrossRef]
- Nabeshima, T.; Nitta, A. Memory impairment and neuronal dysfunction induced by β-amyloid protein in rats. Tohoku J. Exp. Med. 1994, 174, 241–249. [Google Scholar] [CrossRef] [Green Version]
- Nitta, A.; Itoh, A.; Hasegawa, T.; Nabeshima, T. beta-Amyloid protein-induced Alzheimer′s disease animal model. Neurosci. Lett. 1994, 170, 63–66. [Google Scholar] [CrossRef]
- Maurice, T.; Lockhart, B.P.; Privat, A. Amnesia induced in mice by centrally administered beta-amyloid peptides involves cholinergic dysfunction. Brain Res. 1996, 706, 181–193. [Google Scholar] [CrossRef]
- Chen, S.Y.; Wright, J.W.; Barnes, C.D. The neurochemical and behavioral effects of beta-amyloid peptide(25-35). Brain Res. 1996, 720, 54–60. [Google Scholar] [CrossRef]
- Lukoyanov, N.V.; Madeira, M.; Paula–Barbosa, M.M. Behavioral and neuroanatomical consequences of chronic ethanol intake and withdrawal. Physiol. Behav. 1999, 66, 337–346. [Google Scholar] [CrossRef]
- Fernandez-Teruel, A.; Escorihuela, R.M.; Castellano, B.; González, B.; Tobena, A. Neonatal handling and environmental enrichment effects on emotionality, novelty/reward seeking, and age-related cognitive and hippocampal impairments: Focus on the roman rat lines. Behav. Genet. 1997, 27, 513–526. [Google Scholar] [CrossRef]
- Iuvone, L.; Geloso, M.C.; Dell’Anna, E. Changes in open field behavior, spatial memory, and hippocampal parvalbumin immunoreactivity following enrichment in rats exposed to neonatal anoxia. Exp. Neurol. 1996, 139, 25–33. [Google Scholar] [CrossRef]
- Decker, M.W.; Curzon, P.; Brioni, J.D.; Arnerić, S.P. Effects of ABT-418, a novel cholinergic channel ligand, on place learning in septal-lesioned rats. Eur. J. Pharmacol. 1994, 261, 217–222. [Google Scholar] [CrossRef]
- Socci, D.; Crandall, B.; Arendash, G. Chronic antioxidant treatment improves the cognitive performance of aged rats. Brain Res. 1995, 693, 88–94. [Google Scholar] [CrossRef]
- Winkler, J.; Suhr, S.T.; Gage, F.H.; Thal, L.J.; Fisher, L.J. Essential role of neocortical acetylcholine in spatial memory. Nature 1995, 375, 484–487. [Google Scholar] [CrossRef]
- Dickinson-Anson, H.; Aubert, I.; Gage, F.; Fisher, L. Hippocampal grafts of acetylcholine-producing cells are sufficient to improve behavioural performance following a unilateral fimbria–fornix lesion. Neuroscience 1998, 84, 771–781. [Google Scholar] [CrossRef]
- Xiang, X.; Wang, X.; Jin, S.; Hu, J.; Wu, Y.; Li, Y.; Wu, X. Activation of GPR55 attenuates cognitive impairment and neurotoxicity in a mouse model of Alzheimer′s disease induced by Abeta1-42 through inhibiting RhoA/ROCK2 pathway. Prog. Neuropsychopharmacol. Biol. Psychiatry 2022, 112, 110423. [Google Scholar] [CrossRef]
- Saxena, G.; Patro, I.K.; Nath, C. ICV STZ induced impairment in memory and neuronal mitochondrial function: A protective role of nicotinic receptor. Behav. Brain Res. 2011, 224, 50–57. [Google Scholar] [CrossRef]
- Chen, Y.; Liang, Z.; Tian, Z.; Blanchard, J.; Dai, C.L.; Chalbot, S.; Iqbal, K.; Liu, F.; Gong, C.X. Intracerebroventricular streptozotocin exacerbates Alzheimer-like changes of 3xTg-AD mice. Mol. Neurobiol. 2014, 49, 547–562. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study | Model | Selection | Comparability | Outcome | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| ROCK Inhibitor | 1 | 2 | 3 | 4 | 1a | 1b | 1 | 2 | 3 | |
| Yun AD ROCKI 2013 [101] | AD | * | * | * | * | * | * | * | * | |
| YU AD ROCKI 2020 [102] | AD | * | * | * | * | * | * | * | * | * |
| Yu AD ROCKI 2017 [103] | AD | * | * | * | * | * | * | * | * | * |
| Qing-fang AD ROCKI 2018 [104] | AD | * | * | * | * | * | * | * | * | * |
| Min-fang AD ROCKI 2020 [105] | AD | * | * | * | * | * | * | * | * | |
| Manish AD ROCKI 2018 [106] | AD | * | * | * | * | * | * | * | * | * |
| K.H. Reeta AD ROCKI 2017 [107] | AD | * | * | * | * | * | * | * | * | * |
| Jogender AD ROCKI 2013 [108] | AD | * | * | * | * | * | * | * | * | |
| Jiezhong Yu AD ROCKI 2018 [109] | AD | * | * | * | * | * | * | * | * | * |
| Ming ROCKI 2018 [110] | VD | * | * | * | * | * | * | * | * | |
| Lin Huan ROCKI 2008 [111] | VD | * | * | * | * | * | * | * | ||
| PDE-5 inhibitor | ||||||||||
| Cuadrado Cognitive PDE5I 2011 [112] | AD | * | * | * | * | * | * | * | * | |
| Cuadrado-Tejedor Cognitive PDE5I 2017 [113] | AD | * | * | * | * | * | * | * | * | |
| Garcia Cognitive PDE5I 2013 [114] | AD | * | * | * | * | * | * | * | * | |
| Gulisano Cognitive PDE5I 2018 [115] | AD | * | * | * | * | * | * | * | ||
| Puzzo Cognitive PDE5I 2009 [116] | AD | * | * | * | * | * | * | * | ||
| Mohamed Cognitive PDE5I 2021 [117] | AD | * | * | * | * | * | * | * | * | |
| Venkat Cognitive PDE5I 2019 [118] | AD | * | * | * | * | * | * | * | * | |
| Zhang Cognitive PDE5I 2018 [119] | AD | * | * | * | * | * | * | * | ||
| Zhu Cognitive PDE5I 2015 [120] | AD | * | * | * | * | * | * | * | ||
| Bhatia Cognitive PDE5I 2019 [121] | VD | * | * | * | * | * | * | * | ||
| Gulati Cognitive PDE5I 2014 [122] | VD | * | * | * | * | * | * | * | ||
| Gulati P Cognitive PDE5I 2014 [123] | VD | * | * | * | * | * | * | * | ||
| Study | Model | Sex | Age | Treatment Drug Injection | Injected Volume | |
|---|---|---|---|---|---|---|
| ROCK Inhibitor | Control (Disease) | Experimental (Treatment) | ||||
| Yun AD ROCKI 2013 | Ab1–42 | Fasudil | Male | N.R | stereotaxic, left lateral ventricle | 10 mg/kg |
| YU AD ROCKI 2020 | APP/PS1 | Ganoderma lucidum Triterpenoids (ROCKI) | Male | 3 months | gavage | 1.4 g/kg |
| Yu AD ROCKI 2017 | APP/PS1 | Fasudil | N.R | N.R | I.P | 25 mg/kg/day |
| Qing-fang AD ROCKI 2018 | APP/PS1 | Fasudil | N.R | 8 months | I.P | 25 mg/kg/day |
| Min-fang AD ROCKI 2020 | APP/PS1 | Fasudil | N.R | 8 months | N.R | 25 mg/kg/day |
| Manish AD ROCKI 2018 | ICV-STZ | Fasudil | N.R | N.R | I.C.V | 3 mg/kg in 10 μL |
| K.H.Reeta AD ROCKI 2017 | ICV-STZ | Edaravone (ROCKI) | Male | N.R | I.C.V | 10 mg/kg |
| Jogender AD ROCKI 2013 | ICV-STZ | Clitoria ternatea (ROCKI) | Male | N.R | I.C.V | 500 mg/kg |
| Jiezhong Yu AD ROCKI 2018 | APP/PS1 | Fasudil | Male | 8 months | I.C.V | 25 mg/kg/day |
| Ming ROCKI 2018 | BCAO | Y-27632 | Male | N.R | I.P | 10 mg/kg |
| Lin Huan ROCKI 2008 | BCAL | Fasudil | Male | N.R | I.P | 10 mg/kg |
| PDE-5 inhibitor | ||||||
| Cuadrado Cognitive PDE5I 2011 | Tg2576 | Sildenafil | Female | 14–16 months | I.P | 15 mg/kg/day |
| Cuadrado-Tejedor Cognitive PDE5I 2017 | Tg2576 | CM-414 (PDE5i) | Female | 14–16 months | I.P | 40 mg/kg/day |
| Garcia Cognitive PDE5I 2013 | J20 | Sildenafil | Both | 3 months | oral gavage | 15 mg/kg |
| Gulisano Cognitive PDE5I 2018 | APPswe | Vardenafil | Both | 9–10 months | I.P | 0.01 mg/kg |
| Puzzo Cognitive PDE5I 2009 | APP/PS1 | Sildenafil, Tadalafil | Both | 3 months | I.P | 3 mg/kg/day |
| Mohamed Cognitive PDE5I 2021 | ICV-STZ | Tadalafil | Male | 2 months | I.C.V | 20 mg/kg/day |
| Venkat Cognitive PDE5I 2019 | Cholesterol crystal | Sildenafil | Male | 16–18 months | Internal carotid artery | 2 mg/kg/day |
| Zhang Cognitive PDE5I 2018 | Multiple micro infarction | KJH-1002 (PDE5i) | Male | 9 weeks | gavage | 20 mg/kg |
| Zhu Cognitive PDE5I 2015 | APP/PS1 | Sildenafil | Male | 7 months | I.P | 6 mg/kg |
| Bhatia Cognitive PDE5I 2019 | BCAO | Tadalafil | Both | 8–9 weeks | oral | 10 mg/kg |
| Gulati Cognitive PDE5I 2014 | BCAO | Tadalafil | Male | N. R | N. R | 20 mg/kg |
| Gulati P Cognitive PDE5I 2014 | BCAO | Tadalafil | Male | N. R | N. R | 20 mg/kg |
| Study | Model | Control (Disease) | Experimental (Treatment) | ||||
|---|---|---|---|---|---|---|---|
| ROCK Inhibitor | Mean (%) | SD (%) | Number | Mean (%) | SD (%) | Number | |
| Yun AD ROCKI 2013 | AD | 27.75 | 2.06 | 20 | 45.74 | 4.90 | 20 |
| YU AD ROCKI 2020 | AD | 21.47 | 4.51 | 6 | 27.14 | 2.17 | 6 |
| Yu AD ROCKI 2017 | AD | 25.00 | 1.26 | 8 | 31.36 | 0.96 | 8 |
| Qing-fang AD ROCKI 2018 | AD | 21.49 | 1.95 | 8 | 30.00 | 2.43 | 8 |
| Min-fang AD ROCKI 2020 | AD | 20.38 | 1.65 | 8 | 38.52 | 4.26 | 8 |
| Manish AD ROCKI 2018 | AD | 18.63 | 4.70 | 6 | 50.42 | 3.85 | 6 |
| K.H.Reeta AD ROCKI 2017 | AD | 12.03 | 2.91 | 6 | 21.66 | 4.44 | 6 |
| Jogender AD ROCKI 2013 | AD | 25.00 | 6.09 | 8 | 49.68 | 9.30 | 8 |
| Jiezhong Yu AD ROCKI 2018 | AD | 36.95 | 3.30 | 9 | 46.70 | 3.06 | 8 |
| Ming ROCKI 2018 | VD | 25.71 | 1.56 | 10 | 36.29 | 1.61 | 10 |
| Lin Huan ROCKI 2008 | VD | 20.43 | 7.60 | 10 | 33.74 | 4.69 | 10 |
| PDE-5 inhibitor | |||||||
| Cuadrado Cognitive PDE5I 2011 | AD | 16.02 | 4.09 | 10 | 49.65 | 9.35 | 10 |
| Cuadrado-Tejedor Cognitive PDE5I 2017 | AD | 19.33 | 3.87 | 8 | 36.11 | 6.59 | 8 |
| Garcia Cognitive PDE5I 2013 | AD | 26.92 | 2.24 | 10 | 37.79 | 13.12 | 10 |
| Gulisano Cognitive PDE5I 2018 | AD | 21.00 | 1.02 | 10 | 30.67 | 1.21 | 9 |
| Puzzo Cognitive PDE5I 2009 | AD | 22.61 | 1.58 | 12 | 32.93 | 1.46 | 11 |
| Mohamed Cognitive PDE5I 2021 | AD | 10.92 | 2.67 | 12 | 21.79 | 3.66 | 12 |
| Venkat Cognitive PDE5I 2019 | AD | 35.67 | 3.38 | 8 | 50.30 | 2.40 | 7 |
| Zhang Cognitive PDE5I 2018 | AD | 27.66 | 1.55 | 8 | 37.29 | 2.07 | 8 |
| Zhu Cognitive PDE5I 2015 | AD | 16.58 | 2.10 | 10 | 23.23 | 3.54 | 10 |
| Bhatia Cognitive PDE5I 2019 | VD | 26.59 | 2.51 | 6 | 43.14 | 1.67 | 6 |
| Gulati Cognitive PDE5I 2014 | VD | 25.45 | 1.03 | 8 | 34.39 | 1.29 | 8 |
| Gulati P Cognitive PDE5I 2014 | VD | 35.06 | 1.26 | 8 | 46.03 | 1.41 | 8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, D.-H.; Lee, J.Y.; Hong, D.-Y.; Lee, E.C.; Park, S.-W.; Jo, Y.N.; Park, Y.J.; Cho, J.Y.; Cho, Y.J.; Chae, S.H.; et al. ROCK and PDE-5 Inhibitors for the Treatment of Dementia: Literature Review and Meta-Analysis. Biomedicines 2022, 10, 1348. https://doi.org/10.3390/biomedicines10061348
Lee D-H, Lee JY, Hong D-Y, Lee EC, Park S-W, Jo YN, Park YJ, Cho JY, Cho YJ, Chae SH, et al. ROCK and PDE-5 Inhibitors for the Treatment of Dementia: Literature Review and Meta-Analysis. Biomedicines. 2022; 10(6):1348. https://doi.org/10.3390/biomedicines10061348
Chicago/Turabian StyleLee, Dong-Hun, Ji Young Lee, Dong-Yong Hong, Eun Chae Lee, Sang-Won Park, Yu Na Jo, Yu Jin Park, Jae Young Cho, Yoo Jin Cho, Su Hyun Chae, and et al. 2022. "ROCK and PDE-5 Inhibitors for the Treatment of Dementia: Literature Review and Meta-Analysis" Biomedicines 10, no. 6: 1348. https://doi.org/10.3390/biomedicines10061348
APA StyleLee, D. -H., Lee, J. Y., Hong, D. -Y., Lee, E. C., Park, S. -W., Jo, Y. N., Park, Y. J., Cho, J. Y., Cho, Y. J., Chae, S. H., Lee, M. R., & Oh, J. S. (2022). ROCK and PDE-5 Inhibitors for the Treatment of Dementia: Literature Review and Meta-Analysis. Biomedicines, 10(6), 1348. https://doi.org/10.3390/biomedicines10061348