
Obliviate! Reviewing Neural Fundamentals of Intentional Forgetting from a Meta-Analytic Perspective


Abstract
:1. Introduction
1.1. Experimental Paradigms
1.2. Hypotheses of Brain Mechanisms: Thought Suppression and/or Substitution
1.3. Current Meta-Analysis
2. Materials and Methods
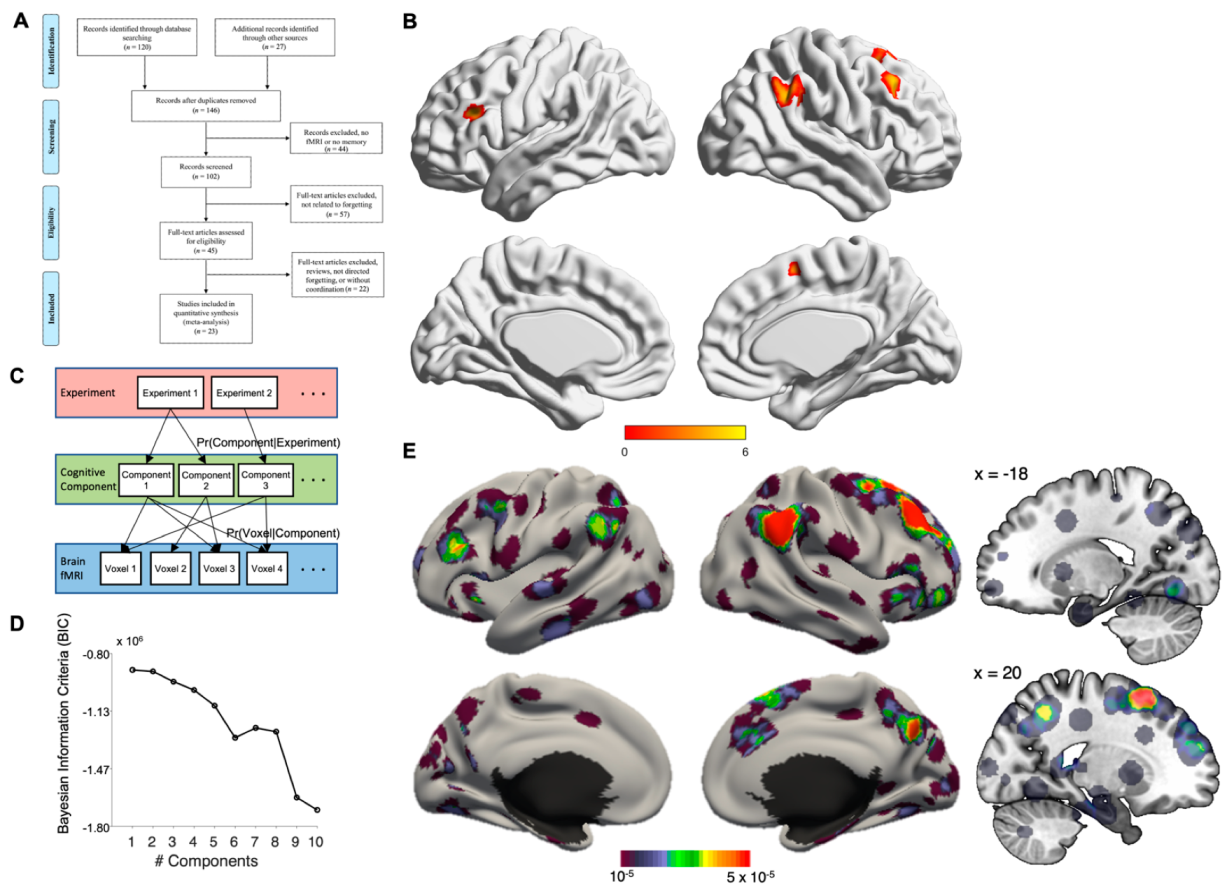
2.1. Literature Search and Article Selection
2.2. Activation Likelihood Estimation (ALE) Analysis
2.3. Latent Dirichlet Allocation (LDA) Analysis
3. Results
4. Discussion
4.1. Core IF Brain Regions
4.2. Supportive IF Network
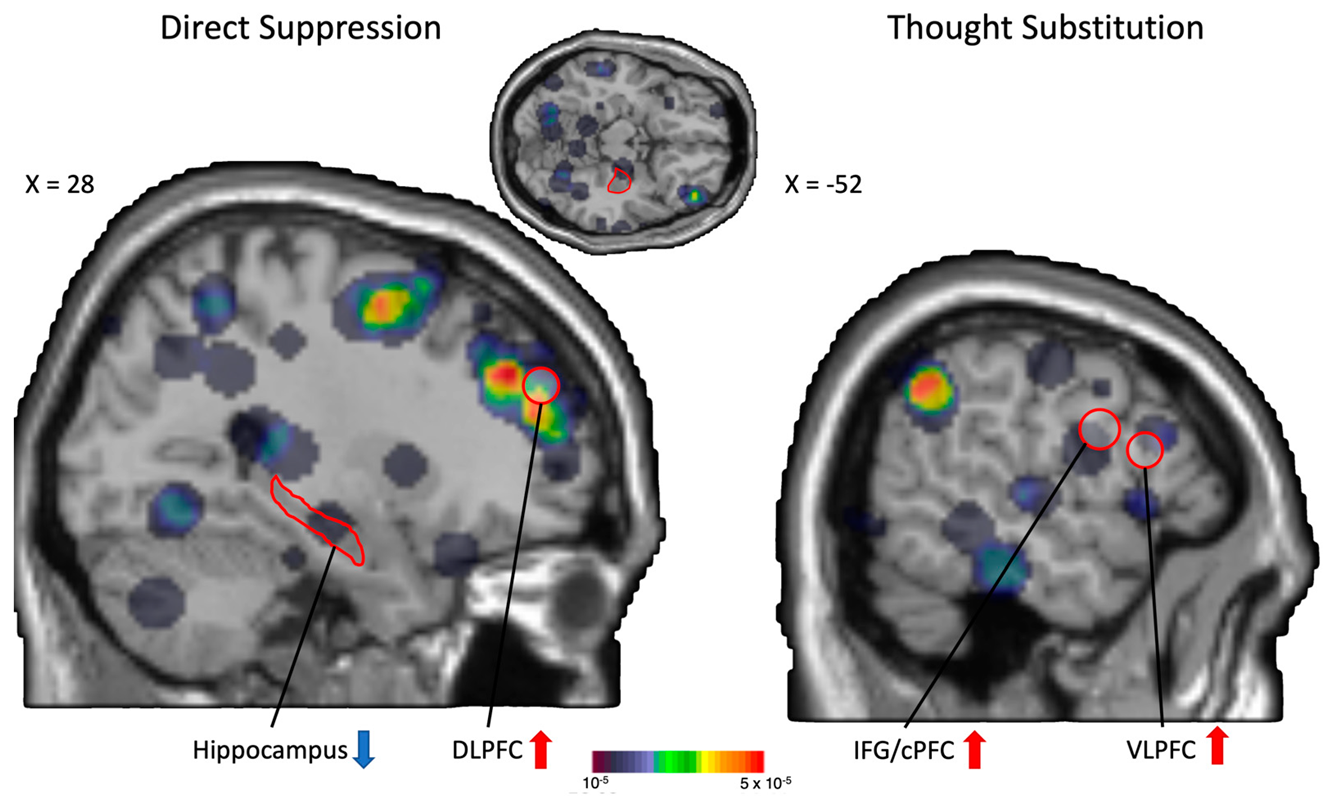
4.3. Thought Suppression and/or Substitution?
4.4. Applicability of the Findings
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bjork, R. Retrieval inhibition as an adaptive mechanism in human memory. In Varieties of Memory and Consciousness: Essays in Honour of Endel Tulving; Roediger, H.L., III, Craik, F., Eds.; Erlbaum: Hillsdale, NJ, USA, 1989; pp. 309–330. [Google Scholar]
- Norby, S. Why Forget? On the Adaptive Value of Memory Loss. Perspect. Psychol. Sci. J. Assoc. Psychol. Sci. 2015, 10, 551–578. [Google Scholar] [CrossRef] [PubMed]
- Nader, K.; Schafe, G.; Le Doux, J. Fear memories require protein synthesis in the amygdala for reconsolidation after retrieval. Nature 2000, 406, 722–726. [Google Scholar] [CrossRef] [PubMed]
- Ryan, T.J.; Frankland, P.W. Forgetting as a form of adaptive engram cell plasticity. Nat. Rev. Neurosci. 2022, 23, 173–186. [Google Scholar] [CrossRef] [PubMed]
- Haaker, J.; Gaburro, S.; Sah, A.; Gartmann, N.; Lonsdorf, T.B.; Meier, K.; Singewald, N.; Pape, H.C.; Morellini, F.; Kalisch, R. Single dose of L-dopa makes extinction memories context-independent and prevents the return of fear. Proc. Natl. Acad. Sci. USA 2018, 110, E2428–E2436. [Google Scholar] [CrossRef] [Green Version]
- Salinas-Hernandez, X.I.; Vogel, P.; Betz, S.; Kalisch, R.; Sigurdsson, T.; Duvarci, S. Dopamine neurons drive fear extinction learning by signaling the omission of expected aversive outcomes. eLife 2018, 7, e38818. [Google Scholar] [CrossRef]
- Kalisch, R.; Korenfeld, E.; Stephan, K.E.; Weiskopf, N.; Seymour, B.; Dolan, R.J. Context-dependent human extinction memory is mediated by a ventromedial prefrontal and hippocampal network. J. Neurosci. Off. J. Soc. Neurosci. 2016, 26, 9503–9511. [Google Scholar] [CrossRef] [Green Version]
- Lonsdorf, T.B.; Haaker, J.; Kalisch, R. Long-term expression of human contextual fear and extinction memories involves amygdala, hippocampus and ventromedial prefrontal cortex: A reinstatement study in two independent samples. Soc. Cogn. Affect. Neurosci. 2014, 9, 1973–1983. [Google Scholar] [CrossRef] [Green Version]
- Gerlicher, A.; Tüscher, O.; Kalisch, R. Dopamine-dependent prefrontal reactivations explain long-term benefit of fear extinction. Nat. Commun. 2018, 9, 4294. [Google Scholar] [CrossRef] [Green Version]
- Thiele, M.; Yuen, K.; Gerlicher, A.; Kalisch, R. A ventral striatal prediction error signal in human fear extinction learning. NeuroImage 2021, 229, 117709. [Google Scholar] [CrossRef]
- Kalisch, R.; Gerlicher, A.; Duvarci, S. A Dopaminergic Basis for Fear Extinction. Trends Cogn. Sci. 2019, 23, 274–277. [Google Scholar] [CrossRef]
- Wagner, A.D.; Schacter, D.L.; Rotte, M.; Koutstaal, W.; Maril, A.; Dale, A.M.; Rosen, B.R.; Buckner, R.L. Building memories: Remembering and forgetting of verbal experiences as predicted by brain activity. Science 1998, 281, 1188–1191. [Google Scholar] [CrossRef] [Green Version]
- Kim, H. Neural activity that predicts subsequent memory and forgetting: A meta-analysis of 74 fMRI studies. Neuroimage 2011, 54, 2446–2461. [Google Scholar] [CrossRef] [PubMed]
- Golding, J.M.; Long, D.L. There’s more to intentional forgetting than directed forgetting: An integrative review. In Intentional Forgetting: Interdisciplinary Approaches; Lawrence Erlbaum Associates Publishers: Mahwah, NJ, USA, 1998; pp. 59–102. [Google Scholar]
- Freud, S. Resistance and suppression. In A General Introduction to Psychoanalysis; Horace Liveright: New York, NY, USA, 1920; pp. 248–261. [Google Scholar]
- Anderson, M.C.; Green, C. Suppressing unwanted memories by executive control. Nature 2001, 410, 366–369. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.C.; Ochsner, K.N.; Kuhl, B.; Cooper, J.; Robertson, E.; Gabrieli, S.W.; Glover, G.H.; Gabrieli, J.D.E. Neural systems underlying the suppression of unwanted memories. Science 2004, 303, 232–235. [Google Scholar] [CrossRef] [PubMed]
- Chalkia, A.; Schroyens, N.; Leng, L.; Vanhasbroeck, N.; Zenses, A.-K.; Van Oudenhove, L.; Beckers, T. No persistent attenuation of fear memories in humans: A registered replication of the reactivation-extinction effect. Cortex 2020, 129, 496–509. [Google Scholar] [CrossRef]
- Basden, B.H.; Basden, D.R. Directed forgetting: Further comparisons of the item and list methods. Memory 1996, 4, 633–653. [Google Scholar] [CrossRef]
- MacLeod, C.M. The item and list methods of directed forgetting: Test differences and the role of demand characteristics. Psychon. Bull. Rev. 1999, 6, 123–129. [Google Scholar] [CrossRef]
- Depue, B.E.; Banich, M.T.; Curran, T. Suppression of emotional and nonemotional content in memory: Effects of repetition on cognitive control. Psychol. Sci. 2006, 17, 441–447. [Google Scholar] [CrossRef] [Green Version]
- Bulevich, J.B.; Roediger, H.L.; Balota, D.A.; Butler, A.C. Failures to find suppression of episodic memories in the think/no-think paradigm. Mem. Cogn. 2006, 34, 1569–1577. [Google Scholar] [CrossRef] [Green Version]
- Hertel, P.T.; Calcaterra, G. Intentional forgetting benefits from thought substitution. Psychon. Bull. Rev. 2005, 12, 484–489. [Google Scholar] [CrossRef] [Green Version]
- Geiselman, R.E.; Bjork, R.A.; Fishman, D.L. Disrupted retrieval in directed forgetting: A link with posthypnotic amnesia. J. Exp. Psychol. Gen. 1983, 112, 58–72. [Google Scholar] [CrossRef] [PubMed]
- Basden, B.H.; Basden, D.R.; Gargano, G.J. Directed forgetting in implicit and explicit memory tests: A comparison of methods. J. Exp. Psychol. Learn. Mem. Cogn. 1993, 19, 603–616. [Google Scholar] [CrossRef]
- MacLeod, C. Directed forgetting affects both direct and indirect tests of memory. J. Exp. Psychol. Learn. Mem. Cogn. 1989, 15, 13–21. [Google Scholar] [CrossRef]
- Lee, Y.S. Cognitive load hypothesis of item-method directed forgetting. Q. J. Exp. Psychol. 2012, 65, 1110–1122. [Google Scholar] [CrossRef] [PubMed]
- Nowicka, A.; Marchewka, A.; Jednoróg, K.; Tacikowski, P.; Brechmann, A. Forgetting of emotional information is hard: An fMRI study of directed forgetting. Cereb. Cortex 2011, 21, 539–549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gamboa, O.L.; Yuen, K.S.L.; von Wegner, F.; Behrens, M.; Steinmetz, H. The challenge of forgetting: Neurobiological mechanisms of auditory directed forgetting. Hum. Brain Mapp. 2018, 39, 249–263. [Google Scholar] [CrossRef] [Green Version]
- Wierzba, M.; Riegel, M.; Wypych, M.; Jednoróg, K.; Grabowska, A.; Marchewka, A. Cognitive control over memory–individual differences in memory performance for emotional and neutral material. Sci. Rep. 2018, 8, 3808. [Google Scholar] [CrossRef] [Green Version]
- Yang, T.; Lei, X.; Anderson, M. Decreased inhibitory control of negative information in directed forgetting. Int. J. Psychophysiol. 2016, 100, 44–51. [Google Scholar] [CrossRef]
- Yang, W.; Liu, P.; Cui, Q.; Wei, D.; Li, W.; Qiu, J.; Zhang, Q. Directed forgetting of negative self-referential information is difficult: An FMRI study. PLoS ONE 2013, 8, e75190. [Google Scholar] [CrossRef]
- Zacks, R.T.; Radvansky, G.; Hasher, L. Studies of directed forgetting in older adults. J. Exp. Psychol. Learn. Mem. Cogn. 1996, 22, 143–156. [Google Scholar] [CrossRef]
- Hourihan, K.L.; Taylor, T.L. Cease remembering: Executive control processes in directed forgetting. J. Exp. Psychol. Hum. Percept. Perform. 2006, 32, 1354–1365. [Google Scholar] [CrossRef] [PubMed]
- Taylor, T.L. Inhibition of return following instructions to remember and forget. Q. J. Exp. Psychol. A 2005, 58, 613–629. [Google Scholar] [CrossRef] [PubMed]
- Wylie, G.R.; Foxe, J.J.; Taylor, T.L. Forgetting as an active process: An FMRI investigation of item-method-directed forgetting. Cereb. Cortex 2008, 18, 670–682. [Google Scholar] [CrossRef] [PubMed]
- Gamboa, O.L.; Garcia-Campayo, J.; Müller, T.; von Wegner, F. Suppress to Forget: The Effect of a Mindfulness-Based Strategy during an Emotional Item-Directed Forgetting Paradigm. Front. Psychol. 2017, 8, 432. [Google Scholar] [CrossRef] [Green Version]
- Benoit, R.G.; Anderson, M.C. Opposing mechanisms support the voluntary forgetting of unwanted memories. Neuron 2012, 76, 450–460. [Google Scholar] [CrossRef] [Green Version]
- Noreen, S.; O’Connor, A.R.; MacLeod, M.D. Neural Correlates of Direct and Indirect Suppression of Autobiographical Memories. Front. Psychol. 2016, 7, 379. [Google Scholar] [CrossRef] [Green Version]
- Ngo, G.H.; Eickhoff, S.B.; Nguyen, M.; Sevinc, G.; Fox, P.T.; Spreng, R.N.; Yeo, B.T. Beyond consensus: Embracing heterogeneity in curated neuroimaging meta-analysis. Neuroimage 2019, 200, 142–158. [Google Scholar] [CrossRef]
- Eickhoff, S.B.; Laird, A.; Grefkes, C.; Wang, L.; Zilles, K.; Fox, P.T. Coordinate-based activation likelihood estimation meta-analysis of neuroimaging data: A random-effects approach based on empirical estimates of spatial uncertainty. Hum. Brain Mapp. 2009, 30, 2907–2926. [Google Scholar] [CrossRef] [Green Version]
- Turkeltaub, P.E.; Eden, G.F.; Jones, K.M.; Zeffiro, T.A. Meta-analysis of the functional neuroanatomy of single-word reading: Method and validation. Neuroimage 2002, 16, 765–780. [Google Scholar] [CrossRef]
- Yeo, B.T.T.; Krienen, F.M.; Eickhoff, S.B.; Yaakub, S.N.; Fox, P.T.; Buckner, R.L.; Asplund, C.; Chee, M. Functional specialization and flexibility in human association cortex. Cereb. Cortex 2015, 25, 3654–3672. [Google Scholar] [CrossRef]
- Zhang, X.; Mormino, E.C.; Sun, N.; Sperling, R.A.; Sabuncu, M.R.; Yeo, B.T.T.; Weiner, M.W.; Aisen, P.; Petersen, R.; Jack, C.R.; et al. Bayesian model reveals latent atrophy factors with dissociable cognitive trajectories in Alzheimer’s disease. Proc. Natl. Acad. Sci. USA 2016, 113, E6535–E6544. [Google Scholar] [CrossRef] [Green Version]
- Sun, N.; Mormino, E.C.; Chen, J.; Sabuncu, M.R.; Yeo, B.T.; Alzheimer’s Disease Neuroimaging Initiative. Multi-modal latent factor exploration of atrophy, cognitive and Tau heterogeneity in Alzheimer’s Disease. Neuroimage 2019, 201, 116043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Groot, C.; Yeo, B.T.; Vogel, J.W.; Zhang, X.; Sun, N.; Mormino, E.C.; Pijnenburg, Y.A.; Miller, B.L.; Rosen, H.J.; La Joie, R.; et al. Latent atrophy factors related to phenotypical variants of posterior cortical atrophy. Neurology 2020, 95, e1672–e1685. [Google Scholar] [CrossRef] [PubMed]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; The PRISMA Group. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. Int. J. Surg. 2010, 8, 336–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turkeltaub, P.E.; Eickhoff, S.B.; Laird, A.; Fox, M.; Wiener, M.; Fox, P. Minimizing within-experiment and within-group effects in activation likelihood estimation meta-analyses. Hum. Brain Mapp. 2012, 33, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Eickhoff, S.B.; Bzdok, D.; Laird, A.; Kurth, F.; Fox, P.T. Activation likelihood estimation meta-analysis revisited. Neuroimage 2012, 59, 2349–2361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Qiu, B.; Giles, C.L.; Foley, H.C.; Yen, J. An LDA-based community structure discovery approach for large-scale social networks. In Proceedings of the 2007 IEEE Intelligence and Security Informatics, Brunswick, NJ, USA, 23–24 May 2007; pp. 200–207. [Google Scholar]
- Eickhoff, S.B.; Nichols, T.E.; Laird, A.R.; Hoffstaedter, F.; Amunts, K.; Fox, P.T.; Bzdok, D.; Eickhoff, C.R. Behavior, sensitivity, and power of activation likelihood estimation characterized by massive empirical simulation. Neuroimage 2016, 137, 70–85. [Google Scholar] [CrossRef] [Green Version]
- Hu, S.; Ide, J.S.; Zhang, S.; Li, C.-S.R. The Right Superior Frontal Gyrus and Individual Variation in Proactive Control of Impulsive Response. J. Neurosci. 2016, 36, 12688–12696. [Google Scholar] [CrossRef] [Green Version]
- Bastin, C.; Feyers, D.; Majerus, S.; Balteau, E.; Degueldre, C.; Luxen, A.; Maquet, P.; Salmon, E.; Collette, F. The neural substracts of memory suppression: A fMRI exploration of directed forgetting. PLoS ONE 2012, 7, e29905. [Google Scholar] [CrossRef]
- Benoit, R.G.; Hulbert, J.C.; Huddleston, E.; Anderson, M.C. Adaptive top-down suppression of hippocampal activity and the purging of intrusive memories from consciousness. J. Cog. Neurosci. 2015, 27, 96–111. [Google Scholar] [CrossRef]
- Butler, A.J.; James, K.H. The neural correlates of attempting to suppress negative versus neutral memories. Cogn. Affect. Behav. Neurosci. 2010, 10, 182–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Depue, B.E.; Curran, T.; Banich, M.T. Prefrontal regions orchestrate suppression of emotional memories via a two-phase process. Science 2007, 317, 215–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Depue, B.E.; Orr, J.M.; Smolker, H.R.; Naaz, F.; Banich, M.T. The organization of right prefrontal networks reveals common mechanisms of inhibitory regulation across cognitive, emotional, and motor processes. Cereb. Cortex 2016, 26, 1634–1646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gagnepain, P.; Henson, R.N.; Anderson, M.C. Suppressing unwanted memories reduces their unconscious influence via targeted cortical inhibition. Proc. Natl. Acad. Sci. USA 2014, 111, E1310–E1319. [Google Scholar] [CrossRef] [Green Version]
- Gagnepain, P.; Hulbert, J.; Anderson, M.C. Parallel regulation of memory and emotional supports the suppression of intrusive memories. J. Neurosci. 2017, 37, 6423–6441. [Google Scholar] [CrossRef] [Green Version]
- Hanslmayr, S.; Volberg, G.; Wimber, M.; Oehler, N.; Staudigl, T.; Hartmann, T.; Raabe, M.; Greenlee, M.W.; Bäuml, K.-H.T. Prefrontally driven downregulation of neural synchrony mediates goal-directed forgetting. J. Neurosci. 2012, 32, 14742–14751. [Google Scholar] [CrossRef] [Green Version]
- Marchewka, A.; Wypych, M.; Michałowski, J.; Sińczuk, M.; Wordecha, M.; Jednoróg, K.; Nowicka, A. What is the effect of basic emotions on directed forgetting? Investigating the role of basic emotions in memory. Front. Hum. Neurosci. 2016, 10, 378. [Google Scholar] [CrossRef] [Green Version]
- Reber, P.J.; Siwiec, R.M.; Gitleman, D.R.; Parrish, T.; Mesulam, M.-M.; Paller, K. Neural correlates of successful encoding identifies using functional magnetic resonance imaging. J. Neurosci. 2002, 22, 9541–9548. [Google Scholar] [CrossRef]
- Rizio, A.; Dennis, N.A. The neural correlates of cognitive control: Successful remembering and intentional forgetting. J. Cogn. Neurosci. 2013, 25, 297–312. [Google Scholar] [CrossRef]
- Sacchet, M.D.; Levy, B.J.; Hamilton, J.P.; Maksimovskiy, A.; Hertel, P.T.; Joormann, J.; Anderson, M.; Wagner, A.D.; Gotlib, I.H. Cognitive and neural consequences of memory suppression in major depressive disorder. Cogn. Affect. Behav. Neurosci. 2017, 17, 77–93. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.H.; Placek, K.; Lewis-Peacock, J.A. More is less: Increased processing of unwanted memories facilitates forgetting. J. Neurosci. 2019, 39, 3551–3560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, W.; Chen, Q.; Liu, P.; Cheng, H.; Cui, Q.; Wei, D.; Zhang, Q.; Qiu, J. Abnormal brain activation during directed forgetting of negative memory in depressed patients. J. Affect. Disord. 2016, 190, 880–888. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, S.; Honda, M.; Sadato, N. Modality-specific cognitive function of medial and lateral human Brodmann area 6. J. Neurosci. 2005, 25, 496–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Qin, W.; Liu, H.; Fan, L.; Wang, J.; Jiang, T.; Yu, C. Subregions of the human superior frontal gyrus and their connections. Neuroimage 2013, 78, 46–58. [Google Scholar] [CrossRef] [PubMed]
- Rajah, M.N.; Languay, R.; Grady, C.L. Age-related changes in right middle frontal gyrus volume correlate with altered episodic retrieval activity. J. Neurosci. 2011, 31, 17941–17954. [Google Scholar] [CrossRef] [Green Version]
- Rajah, M.N.; Ames, B.; D’Esposito, M. Prefrontal contributions to domain-general executive control processes during temporal context retrieval. Neuropsychologia 2008, 46, 1088–1103. [Google Scholar] [CrossRef] [Green Version]
- Japee, S.; Holiday, K.; Satyshur, M.D.; Mukai, I.; Ungerleider, L.G. A role of right middle frontal gyrus in reorienting of attention: A case study. Front. Syst. Neurosci. 2015, 9, 23. [Google Scholar] [CrossRef] [Green Version]
- Corbetta, M.; Patel, G.; Shulman, G.L. The reorienting system of the human brain: From environment to theory of mind. Neuron 2008, 58, 306–324. [Google Scholar] [CrossRef] [Green Version]
- Conway, M.A.; Fthenaki, A. Disruption of inhibitory control of memory following lesions to the frontal and temporal lobes. Cortex 2003, 39, 667–686. [Google Scholar] [CrossRef]
- Singh-Curry, V.; Husain, M. The functional role of the inferior parietal lobe in the dorsal and ventral stream dichotomy. Neuropsychologia 2009, 47, 1434–1448. [Google Scholar] [CrossRef] [Green Version]
- Uncapher, M.R.; Wagner, A.D. Posterior parietal cortex and episodic encoding: Insights from fMRI subsequent memory effects and dual-attention theory. Neurobiol. Learn. Mem. 2009, 91, 139–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabeza, R. Role of parietal regions in episodic memory retrieval: The dual attentional processes hypothesis. Neuropsychologia 2008, 46, 1813–1827. [Google Scholar] [CrossRef] [Green Version]
- Daselaar, S.M.; Prince, S.E.; Cabeza, R. When less means more: Deactivations during encoding that predict subsequent memory. Neuroimage 2004, 23, 921–927. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, T.W.; Correia, M.M.; Ferreira, C.; Prescot, A.P.; Anderson, M.C. Hippocampal GABA enables inhibitory control over unwanted thoughts. Nat. Commun. 2017, 8, 1311. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.; Johnson, N.F.; Cilles, S.E.; Gold, B.T. Common and distinct mechanisms of cognitive flexibility in prefrontal cortex. J. Neurosci. 2011, 31, 4771–4779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, C.; Cilles, S.E.; Johnson, N.F.; Gold, B.T. Domain general and domain preferential brain regions associated with different types of task switching: A Meta-analysis. Hum. Brain Mapp. 2012, 33, 130–142. [Google Scholar] [CrossRef] [Green Version]
- Yoo, K.; Rosenberg, M.D.; Kwon, Y.H.; Lin, Q.; Avery, E.W.; Sheinost, D.; Constable, R.T.; Chun, M.M. A brain-based general measure of attentions. Nat. Hum. Behav. 2022, 6, 782–795. [Google Scholar] [CrossRef]
- Gottwald, B.; Mihajlovic, Z.; Wilde, B.; Mehdorn, H.M. Does the cerebellum contribute to specific aspect of attention? Neuropsychologia 2003, 11, 1452–1460. [Google Scholar] [CrossRef]
- Courchesne, E.; Allen, G. Prediction and preparation, fundamental functions of the cerebellum. Learn. Mem. 1997, 4, 1–35. [Google Scholar] [CrossRef] [Green Version]
- Geraerts, E.; Merckelbach, H.; Jelicic, M.; Smeets, E. Long term consequences of suppression of intrusive anxious thoughts and repressive coping. Behav. Res. Ther. 2006, 44, 1451–1460. [Google Scholar] [CrossRef]
- Dalgleish, T.; Hauer, B.; Kuyken, W. The mental regulation of autobiographical recollection in the aftermath of trauma. Curr. Dir. Psychol. Sci. 2008, 17, 259–263. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Studies | n | Age | Software 1 | Paradigm 2 | Stimuli | Contrast |
|---|---|---|---|---|---|---|
| Anderson et al., 2004 [17] | 24 (10F) | 29–31 | SPM99 | T/NT | word pairs | suppression > recall |
| Bastin et al., 2012 [52] | 17 (8F) | 20–32 | SPM5 | DF | 6-letter words | To be forget-forget > To be remember-forget |
| Benoit et al., 2012 [37] | 18 (12F) | 23.7 | SPM8 | T/NT | word pairs | suppression > recall |
| Benoit et al., 2015 [53] | 16 (8F) | 22 | SPM8 | T/NT | Picture | suppression > recall |
| Butler et al., 2010 [54] | 14 (7F) | 22.6 | BV | T/NT | emotion pictures | NT > T (neutral) |
| Depue et al., 2007 [55] | 16 (8F) | 19–29 | FSL | T/NT | face-picture pairs | Suppression > recall |
| Depue et al., 2016 [56] | 21 (10F) | 21.5 | FSL | T/NT | neutral face pictures | Suppression > recall |
| Gagnepain et al., 2014 [57] | 24 (11F) | 22. | SPM8 | T/NT | word-object pairs | Suppression > recall |
| Gagnepain et al., 2017 [58] | 22 (8F) | 18–35 | SPM12 | T/NT | face-scene pairs | NT > T |
| Gamboa et al., 2018 [28] | 31 (15F) | 27.5 | SPM12 | DF | vocal words | To be Forget > to be remember |
| Hanslmayr et al., 2012 [59] | 22 (15F) | 23.05 | SPM5 | DF | words | To be Forget > to be remember |
| Marchewka et al., 2016 [60] | 18 (18F) | 22.02 | SPM12 | DF | emotional pictures | TBF-F > TBR-F |
| Noreen et al., 2016 [38] | 22 (18F) | 18–29 | SPM8 | T/NT | word-autobiographic-memory pairs | no-think > think |
| Nowicka et al., 2011 [27] | 16 (8F) | 26.6 | SPM8 | DF | emotional pictures | TBF > TBR for neutral pictures |
| Reber et al., 2002 [61] | 12 (9F) | 20 | NA | DF | faces | TBF > TBR |
| Rizio et al., 2013 [62] | 24 (NA) | 21.11 | SPM8 | DF | words | TBF > TBR |
| Sacchet et al., 2017 [63] | 16 (8F) | 31.7 | AFNI | T/NT | word-pairs | no-think > think |
| Wang et al., 2019 [64] | 20 (10F) | 23.6 | SPM 12 | DF | pictures (scene, faces, objects) | TBF > TBR |
| Wierzba et al., 2018 [29] | 24 (24F) | 24.6 | SPM12 | DF | neutral/affective words | TBF > TBR |
| Wylie et al., 2008 [35] | 11 (6F) | 26 | AFNI | DF | word pairs | TBF > TBR |
| Yang, T. et al., 2016 [30] | 21 (13F) | 22.19 | SPM8 | DF | word pairs | TBF > TBR (neutral words) |
| Yang, W. et al., 2013 [31] | 25 (14F) | 30 | SPM8 | DF | word pairs | TBF > TBR (neutral words) |
| Yang, W. et al., 2016 [65] | 32 (10F) | 30 | SPM8 | DF | word pairs | TBF > TBR |
| Coordinates (MNI) | ||||||
|---|---|---|---|---|---|---|
| Cluster | X | Y | Z | Number of Voxels | L/R | Anatomical Structure |
| ALE | ||||||
| 1 | 16 | 16 | 60 | 221 | R | Superior Frontal Gyrus |
| 2 | 58 | −46 | 36 | 212 | R | Inferior Parietal Lobe |
| 3 | 42 | 24 | 44 | 160 | R | Middle Frontal Gyrus |
| 4 | −42 | 28 | 24 | 117 | L | Middle Frontal Gyrus |
| LDA | ||||||
| 1 | −45 | 15 | 1 | 4091 | L | Inferior Frontal Gyrus |
| 2 | −27 | 45 | 21 | 724 | L | Middle Frontal Gyrus |
| 3 | 37 | 27 | 41 | 16,556 | R | Middle Frontal Gyrus |
| 4 | −21 | 51 | −3 | 536 | L | Orbitofrontal Gyrus |
| 5 | −43 | −1 | 45 | 2122 | L | Precentral Gyrus |
| 6 | −15 | −3 | 45 | 498 | L | Middle Cingulum |
| 7 | 21 | −39 | 43 | 515 | R | Middle Cingulum |
| 8 | 57 | −37 | 13 | 1104 | R | Superior Temporal Gyrus |
| 9 | −55 | −37 | −17 | 2934 | L | Middle Temporal Gyrus |
| 10 | 69 | −25 | −17 | 1012 | R | Middle Temporal Gyrus |
| 11 | 53 | −25 | −33 | 1050 | R | Inferior Temporal Gyrus |
| 12 | −11 | −15 | −23 | 552 | L | Hippocampus |
| 13 | 25 | −25 | −17 | 1500 | R | Parahippocampal Gyrus |
| 14 | −55 | −59 | 39 | 1182 | L | Inferior Parietal Lobe |
| 15 | 57 | 47 | 41 | 13,717 | R | Inferior Parietal Lobe |
| 16 | −7 | −39 | 63 | 536 | L | Precuneus |
| 17 | −43 | −79 | −5 | 931 | L | Inferior Occipital Gyrus |
| 18 | 49 | −87 | −3 | 398 | R | Infeiror Occipital Gyrus |
| 19 | −15 | −75 | −7 | 1332 | L | Lingual Gyrus |
| 20 | 25 | −71 | −9 | 4213 | R | Lingual Gyrus |
| 21 | 13 | −93 | 3 | 1039 | R | Calcarine Gyrus |
| 22 | −33 | −59 | −23 | 552 | L | Cerebellum |
| 23 | 27 | −79 | −35 | 498 | R | Cerebellum |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gamboa, O.L.; Chuan-Peng, H.; Salas, C.E.; Yuen, K.S.L. Obliviate! Reviewing Neural Fundamentals of Intentional Forgetting from a Meta-Analytic Perspective. Biomedicines 2022, 10, 1555. https://doi.org/10.3390/biomedicines10071555
Gamboa OL, Chuan-Peng H, Salas CE, Yuen KSL. Obliviate! Reviewing Neural Fundamentals of Intentional Forgetting from a Meta-Analytic Perspective. Biomedicines. 2022; 10(7):1555. https://doi.org/10.3390/biomedicines10071555
Chicago/Turabian StyleGamboa, Olga Lucia, Hu Chuan-Peng, Christian E. Salas, and Kenneth S. L. Yuen. 2022. "Obliviate! Reviewing Neural Fundamentals of Intentional Forgetting from a Meta-Analytic Perspective" Biomedicines 10, no. 7: 1555. https://doi.org/10.3390/biomedicines10071555
APA StyleGamboa, O. L., Chuan-Peng, H., Salas, C. E., & Yuen, K. S. L. (2022). Obliviate! Reviewing Neural Fundamentals of Intentional Forgetting from a Meta-Analytic Perspective. Biomedicines, 10(7), 1555. https://doi.org/10.3390/biomedicines10071555