
Inflammation of the Human Dental Pulp Induces Phosphorylation of eNOS at Thr495 in Blood Vessels


 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Tissue Sample Collection, Ethic Statement, and Tissue Preparation
2.2. Hematoxylin and Eosin Staining
2.3. Immunohistochemical Methods
2.3.1. Specificity of the Antibodies
2.3.2. Avidin–Biotin–Peroxidase Complex Method
2.3.3. Double Immunofluorescence and Confocal Microscopy
2.3.4. Quantification of Immunohistochemical Results
2.3.5. Statistical Analysis
3. Results
3.1. Characterization of Healthy and Inflamed Human Dentin–Pulp Complex
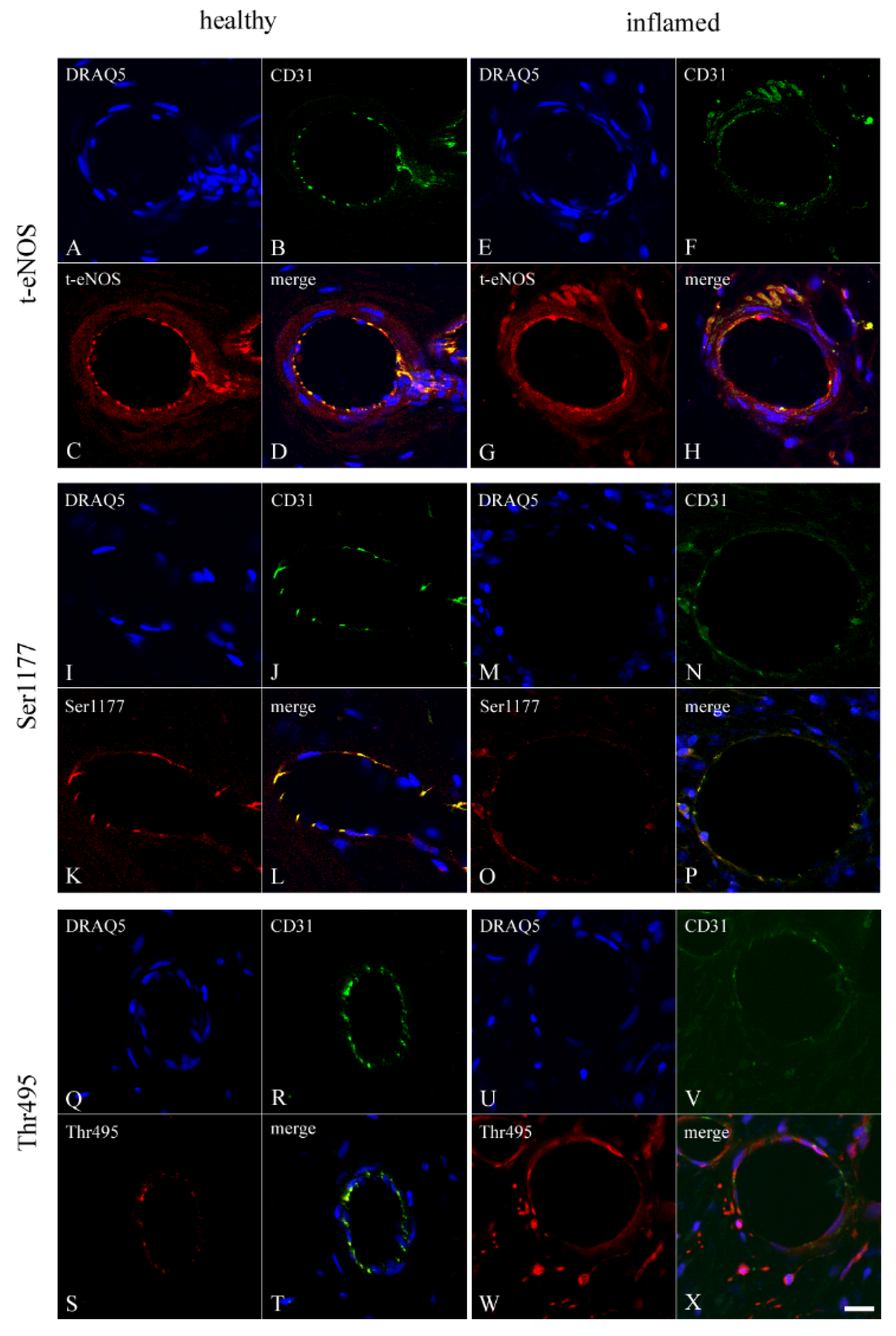
3.2. Co-Localization of CD31 with t-eNOS, Ser1177, and Thr495 in Healthy and Inflamed Endothelium of the Human Dental Pulp
3.3. Localization of t-eNOS, Ser1177, and Thr495 in Healthy Endothelium of the Human Dental Pulp
3.4. Localization of t-eNOS, Ser1177, and Thr495 in Inflamed Endothelium of the Human Dental Pulp
3.5. Immunohistochemical Controls
3.6. Statistical Analysis of t-eNOS, Ser1177, and Thr495 in Blood Vessels of the Healthy and Inflamed Human Dental Pulp
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Babaahmady, K.G.; Challacombe, S.J.; Marsh, P.D.; Newman, H.N. Ecological study of Streptococcus mutans, Streptococcus sobrinus and Lactobacillus spp. at sub-sites from approximal dental plaque from children. Caries Res. 1998, 32, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, N.; Nyvad, B. Caries ecology revisited: Microbial dynamics and the caries process. Caries Res. 2008, 42, 409–418. [Google Scholar] [CrossRef] [PubMed]
- Jenkinson, H.F. Beyond the oral microbiome. Environ. Microbiol. 2011, 13, 3077–3087. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Barraza, J.P.; Arthur, R.A.; Hara, A.; Lewis, K.; Liu, Y.; Scisci, E.L.; Hajishengallis, E.; Whiteley, M.; Koo, H. Spatial mapping of polymicrobial communities reveals a precise biogeography associated with human dental caries. Proc. Natl. Acad. Sci. USA 2020, 117, 12375–12386. [Google Scholar] [CrossRef]
- Damé-Teixeira, N.; Ev, L.D.; Bitello-Firmino, L.; Soares, V.K.; Dalalba, R.S.; Rup, A.G.; Maltz, M.; Parolo, C.C.F. Characterization of Lactobacilli isolated from carious dentin after selective caries removal and cavity sealing. Arch. Oral Biol. 2021, 121, 104988. [Google Scholar] [CrossRef]
- Goldberg, M.; Farges, J.C.; Lacerda-Pinheiro, S.; Six, N.; Jegat, N.; Decup, F.; Septier, D.; Carrouel, F.; Durand, S.; Chaussain-Miller, C.; et al. Inflammatory and immunological aspects of dental pulp repair. Pharmacol. Res. 2008, 58, 137–147. [Google Scholar] [CrossRef] [Green Version]
- Korkmaz, Y.; Lang, H.; Beikler, T.; Cho, B.; Behrends, S.; Bloch, W.; Addicks, K.; Raab, W.H. Irreversible inflammation is associated with decreased levels of the α1-, β1- and α2-subunit of sGC in human odontoblasts. J. Dent. Res. 2011, 90, 517–522. [Google Scholar] [CrossRef]
- Farges, J.C.; Alliot-Licht, B.; Baudouin, C.; Msika, P.; Bleicher, F.; Carrouel, F. Odontoblast control of dental pulp inflammation triggered by cariogenic bacteria. Front. Physiol. 2013, 4, 326. [Google Scholar] [CrossRef] [Green Version]
- Galler, K.M.; Weber, M.; Korkmaz, Y.; Widbiller, M.; Feuerer, M. Inflammatory response mechanisms of the dentine-pulp complex and the periapical tissues. Int. J. Mol. Sci. 2021, 22, 1480. [Google Scholar] [CrossRef]
- Cooper, P.R.; Takahashi, Y.; Graham, L.W.; Simon, S.; Imazato, S.; Smith, A.J. Inflammation-regeneration interplay in the dentine-pulp complex. J. Dent. 2010, 38, 687–697. [Google Scholar] [CrossRef]
- Cooper, P.R.; Holder, M.J.; Smith, A.J. Inflammation and regeneration in the dentin-pulp complex: A double-edged sword. J. Endod. 2014, 40, S46–S51. [Google Scholar] [CrossRef] [PubMed]
- Ricucci, D.; Loghin, S.; Siqueira, J.F., Jr. Correlation between clinical and histologic pulp diagnoses. J. Endod. 2014, 40, 1932–1939. [Google Scholar] [CrossRef] [PubMed]
- Hahn, C.L.; Falkler, W.A., Jr.; Siegel, M.A. A study of T and B cells in pulpal pathosis. J. Endod. 1989, 15, 20–26. [Google Scholar] [CrossRef]
- Izumi, T.; Kobayashi, I.; Okamura, K.; Sakai, H. Immunohistochemical study on the immunocompetent cells of the pulp in human non-carious and carious teeth. Arch. Oral. Biol. 1995, 40, 609–614. [Google Scholar] [CrossRef]
- Iwasaki, Y.; Otsuka, H.; Yanagisawa, N.; Hisamitsu, H.; Manabe, A.; Nonaka, N.; Nakamura, M. In situ proliferation and differentiation of macrophages in dental pulp. Cell Tissue Res. 2011, 346, 99–109. [Google Scholar] [CrossRef] [Green Version]
- Mazzoni, A.; Tjäderhane, L.; Checchi, V.; Di Lenarda, R.; Salo, T.; Tay, F.R.; Pashley, D.H.; Breschi, L. Role of dentin MMPs in caries progression and bond stability. J. Dent. Res. 2015, 94, 241–251. [Google Scholar] [CrossRef] [Green Version]
- Ballal, N.V.; Duncan, H.F.; Wiedemeier, D.B.; Rai, N.; Jalan, P.; Bhat, V.; Belle, V.S.; Zehnder, M. MMP-9 levels and NaOCl lavage in randomized trial on direct pulp capping. J. Dent. Res. 2022, 101, 414–419. [Google Scholar] [CrossRef]
- Li, X.; Hu, L.; Ma, L.; Chang, S.; Wang, W.; Feng, Y.; Xu, Y.; Hu, J.; Zhang, C.; Wang, S. Severe periodontitis may influence cementum and dental pulp through inflammation, oxidative stress, and apoptosis. J. Periodontol. 2019, 90, 1297–1306. [Google Scholar] [CrossRef]
- Lee, D.H.; Lim, B.S.; Lee, Y.K.; Yang, H.C. Effects of hydrogen peroxide (H2O2) on alkaline phosphatase activity and matrix mineralization of odontoblast and osteoblast cell lines. Cell. Biol. Toxicol. 2006, 22, 39–46. [Google Scholar] [CrossRef]
- Smith, A.J. Pulpal responses to caries and dental repair. Caries Res. 2002, 36, 223–232. [Google Scholar] [CrossRef]
- Duncan, H.F.; Cooper, P.R.; Smith, A.J. Dissecting dentine-pulp injury and wound healing responses: Consequences for regenerative endodontics. Int. Endod. J. 2019, 52, 261–266. [Google Scholar] [CrossRef] [Green Version]
- Duncan, H.F. Present status and future directions-Vital pulp treatment and pulp preservation strategies. Int. Endod. J. 2022, 55, 497–511. [Google Scholar] [CrossRef] [PubMed]
- Aguilar, P.; Linsuwanont, P. Vital pulp therapy in vital permanent teeth with cariously exposed pulp: A systematic review. J. Endod. 2011, 37, 581–587. [Google Scholar] [CrossRef] [PubMed]
- Demant, S.; Dabelsteen, S.; Bjørndal, L. A macroscopic and histological analysis of radiographically well-defined deep and extremely deep carious lesions: Carious lesion characteristics as indicators of the level of bacterial penetration and pulp response. Int. Endod. J. 2021, 54, 319–330. [Google Scholar] [CrossRef] [PubMed]
- Taha, N.A.; Khazali, M.A. Partial pulpotomy in mature permanent teeth with clinical signs indicative of irreversible pulpitis: A Randomized Clinical Trial. J. Endod. 2017, 43, 1417–1421. [Google Scholar] [CrossRef]
- Simon, S.; Perard, M.; Zanini, M.; Smith, A.J.; Charpentier, E.; Djole, S.X.; Lumley, P.J. Should pulp chamber pulpotomy be seen as a permanent treatment? Some preliminary thoughts. Int. Endod. J. 2013, 46, 79–87. [Google Scholar] [CrossRef]
- Santos, J.M.; Marques, J.A.; Diogo, P.; Messias, A.; Sousa, V.; Sequeira, D.; Palma, P.J. Influence of preoperative pulp inflammation in the outcome of full pulpotomy using a dog model. J. Endod. 2021, 47, 1417–1426. [Google Scholar] [CrossRef]
- Fleming, I.; Busse, R. Molecular mechanisms involved in the regulation of the endothelial nitric oxide synthase. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2003, 284, R1–R12. [Google Scholar] [CrossRef] [Green Version]
- Fleming, I. Molecular mechanisms underlying the activation of eNOS. Pflug. Arch Eur. J. Physiol. 2010, 459, 793–806. [Google Scholar] [CrossRef]
- Förstermann, U.; Sessa, W.C. Nitric oxide synthases: Regulation and function. Eur. Heart. J. 2012, 33, 829–837. [Google Scholar] [CrossRef] [Green Version]
- Karbach, S.; Wenzel, P.; Waisman, A.; Munzel, T.; Daiber, A. eNOS uncoupling in cardiovascular diseases-the role of oxidative stress and inflammation. Curr. Pharm. Des. 2014, 20, 3579–3594. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Förstermann, U. Pharmacological prevention of eNOS uncoupling. Curr Pharm Des. 2014, 20, 3595–3606. [Google Scholar] [CrossRef] [PubMed]
- Förstermann, U.; Münzel, T. Endothelial nitric oxide synthase in vascular disease: From marvel to menace. Circulation 2006, 113, 1708–1714. [Google Scholar] [CrossRef] [Green Version]
- Förstermann, U.; Li, H. Therapeutic effect of enhancing endothelial nitric oxide synthase (eNOS) expression and preventing eNOS uncoupling. Br. J. Pharmacol. 2011, 164, 213–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolluru, G.K.; Siamwala, J.H.; Chatterjee, S. eNOS phosphorylation in health and disease. Biochimie 2010, 92, 1186–1198. [Google Scholar] [CrossRef] [PubMed]
- Heiss, E.H.; Dirsch, V.M. Regulation of eNOS enzyme activity by posttranslational modification. Curr. Pharm. Des. 2014, 20, 3503–3513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Förstermann, U. Prevention of atherosclerosis by interference with the vascular nitric oxide system. Curr. Pharm. Des. 2009, 15, 3133–3145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Förstermann, U. Uncoupling of endothelial NO synthase in atherosclerosis and vascular disease. Curr. Opin. Pharmacol. 2013, 13, 161–167. [Google Scholar] [CrossRef]
- Chen, D.D.; Chen, L.Y.; Xie, J.B.; Shu, C.; Yang, T.; Zhou, S.; Yuan, H.; Chen, A.F. Tetrahydrobiopterin regulation of eNOS redox function. Curr. Pharm. Des. 2014, 20, 3554–3562. [Google Scholar] [CrossRef]
- Li, H.; Förstermann, U. Nitric oxide in the pathogenesis of vascular disease. J. Pathol. 2000, 190, 244–254. [Google Scholar] [CrossRef]
- Förstermann, U.; Xia, N.; Li, H. Roles of vascular oxidative stress and nitric oxide in the pathogenesis of atherosclerosis. Circ. Res. 2017, 120, 713–735. [Google Scholar] [CrossRef] [PubMed]
- Di Nardo Di Maio, F.; Lohinai, Z.; D’Arcangelo, C.; De Fazio, P.E.; Speranza, L.; De Lutiis, M.A.; Patruno, A.; Grilli, A.; Felaco, M. Nitric oxide synthase in healthy and inflamed human dental pulp. J. Dent. Res. 2004, 83, 312–316. [Google Scholar] [CrossRef] [PubMed]
- Korkmaz, Y.; Baumann, M.A.; Steinritz, D.; Schroder, H.; Behrends, S.; Addicks, K.; Schneider, K.; Raab, W.H.; Bloch, W. NO-cGMP signaling molecules in cells of the rat molar dentin-pulp complex. J. Dent. Res. 2005, 84, 618–623. [Google Scholar] [CrossRef]
- Korkmaz, Y.; Bloch, W.; Steinritz, D.; Baumann, M.A.; Addicks, K.; Schneider, K.; Raab, W.H. Bradykinin mediates phosphorylation of eNOS in odontoblasts. J. Dent. Res. 2006, 85, 536–541. [Google Scholar] [CrossRef] [PubMed]
- Korkmaz, Y.; Imhof, T.; Kämmerer, P.W.; Bloch, W.; Rink-Notzon, S.; Möst, T.; Weber, M.; Kesting, M.; Galler, K.M.; Deschner, J. The colocalizations of pulp neural stem cells markers with dentin matrix protein-1, dentin sialoprotein and dentin phosphoprotein in human denticle (pulp stone) lining cells. Ann. Anat. 2021, 239, 151815. [Google Scholar] [CrossRef] [PubMed]
- Burry, R.W. Specificity controls for immunocytochemical methods. J. Histochem. Cytochem. 2000, 48, 163–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burry, R.W. Controls for immunocytochemistry: An update. J. Histochem. Cytochem. 2011, 59, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Vo, P.A.; Lad, B.; Tomlinson, J.A.P.; Francis, S.; Ahluwalia, A. Autoregulatory role of endothelium-derived nitric oxide (NO) on lipopolysaccharide-induced vascular inducible NO synthase expression and function. J. Biol. Chem. 2005, 280, 7236–7243. [Google Scholar] [CrossRef] [Green Version]
- Geng, B.; Cui, Y.; Zhao, J.; Yu, F.; Zhu, Y.; Xu, G.; Zhang, Z.; Tang, C.; Du, J. Hydrogen sulfide downregulates the aortic L-arginine/nitric oxide pathway in rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 293, R1608–R1618. [Google Scholar] [CrossRef]
- Tepavcevic, S.; Milutinovic, D.V.; Macut, D.; Stanisic, J.; Nikolic, M.; Bozic-Antic, I.; Rodaljevic, S.; Bijekic-Macut, J.; Matic, G.; Koricanac, G. Cardiac nitric oxide synthases and Na+/K+-ATPase in the rat model of polycystic ovary syndrome induced by dihydrotestosterone. Exp. Clin. Endocrinol. Diabetes 2015, 123, 303–307. [Google Scholar]
- Förstermann, U. Oxidative stress in vascular disease: Causes, defense mechanisms and potential therapies. Nat. Clin. Pract. Cardiovasc. Med. 2008, 5, 338–349. [Google Scholar] [CrossRef] [PubMed]
- Landmesser, U.; Dikalov, S.; Price, S.R.; McCann, L.; Fukai, T.; Holland, S.M.; Mitch, W.E.; Harrison, D.G. Oxidation of tetrahydrobiopterin leads to uncoupling of endothelial cell nitric oxide synthase in hypertension. J. Clin. Investig. 2003, 111, 1201–1209. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.; Lei, H.; Qin, H.; Xia, Y. Molecular mechanisms of endothelial NO synthase uncoupling. Curr. Pharm. Des. 2014, 20, 3548–3553. [Google Scholar] [CrossRef] [PubMed]
- Murad, F. Nitric oxide and cyclic guanosine monophosphate signaling in the eye. Can. J. Ophthalmol. 2008, 43, 291–294. [Google Scholar] [CrossRef] [PubMed]
- Friebe, A.; Koesling, D. Regulation of nitric oxide-sensitive guanylyl cyclase. Circ. Res. 2003, 93, 96–105. [Google Scholar] [CrossRef]
- Murad, F. Shattuck Lecture. Nitric oxide and cyclic GMP in cell signaling and drug development. N. Engl. J. Med. 2006, 355, 2003–2011. [Google Scholar] [CrossRef]
- Stasch, J.P.; Pacher, P.; Evgenov, O.V. Soluble guanylate cyclase as an emerging therapeutic target in cardiopulmonary disease. Circulation 2011, 123, 2263–2273. [Google Scholar] [CrossRef] [Green Version]
- Stasch, J.P.; Schmidt, P.M.; Nedvetsky, P.I.; Nedvetskaya, T.Y.; Arun, K.H.S.; Meurer, S.; Deile, M.; Taye, A.; Knorr, A.; Lapp, H.; et al. Targeting the heme-oxidized nitric oxide receptor for selective vasodilatation of diseased blood vessels. J. Clin. Investig. 2006, 116, 2552–2561. [Google Scholar] [CrossRef] [Green Version]
- Rahaman, M.M.; Nguyen, A.T.; Miller, M.P.; Hahn, S.A.; Sparacino-Watkins, C.; Jobbagy, S.; Carew, N.T.; Cantu-Medellin, N.; Wood, K.C.; Baty, C.J.; et al. Cytochrome b5 Reductase 3 Modulates Soluble Guanylate Cyclase Redox State and cGMP Signaling. Circ. Res. 2017, 121, 137–148. [Google Scholar] [CrossRef]
- Daiber, A.; Oelze, M.; Wenzel, P.; Wickramanayake, J.M.; Schuhmacher, S.; Jansen, T.; Lackner, K.J.; Torzewski, M.; Münzel, T. Nitrate tolerance as a model of vascular dysfunction: Roles for mitochondrial aldehyde dehydrogenase and mitochondrial oxidative stress. Pharmacol. Rep. 2009, 61, 33–48. [Google Scholar] [CrossRef]
- Münzel, T.; Gori, T.; Bruno, R.M.; Taddei, S. Is oxidative stress a therapeutic target in cardiovascular disease? Eur. Heart. J. 2010, 31, 2741–2748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paulo, M.; Costa, D.E.; Bonaventura, D.; Lunardi, C.N.; Bendhack, L.M. Nitric Oxide Donors as Potential Drugs for the Treatment of Vascular Diseases Due to Endothelium Dysfunction. Curr. Pharm. Des. 2020, 26, 3748–3759. [Google Scholar] [CrossRef] [PubMed]
- Münzel, T.; Daiber, A. Novel Concept for the Regulation of eNOS (Endothelial Nitric Oxide Synthase) Activity: Inhibitory Effects of the Enigma Homolog Protein and the PHLPP (Pleckstrin Homology Domain and Leucine-Rich Repeat Protein Phosphatase)-2 on Akt (Protein Kinase B)-Dependent Nitric Oxide Synthase Activation. Arterioscler. Thromb. Vasc. Biol. 2020, 40, 1608–1610. [Google Scholar]
- Caballano-Infantes, E.; Cahuana, G.M.; Bedoya, F.J.; Salguero-Aranda, C.; Tejedo, J.R. The role of nitric oxide in stem cell biology. Antioxidants 2022, 11, 497. [Google Scholar] [CrossRef]
- Felaco, M.; Di Maio, F.D.; De Fazio, P.; D’Arcangelo, C.; De Lutiis, M.A.; Varvara, G.; Grilli, A.; Barbacane, R.C.; Reale, M.; Conti, P. Localization of the e-NOS enzyme in endothelial cells and odontoblasts of healthy human dental pulp. Life Sci. 2000, 68, 297–306. [Google Scholar] [CrossRef]
- Bogdan, C. Nitric oxide and the immune response. Nat. Immunol. 2001, 2, 907–916. [Google Scholar] [CrossRef] [PubMed]
- Ibiza, S.; Víctor, V.M.; Boscá, I.; Ortega, A.; Urzainqui, A.; O’Connor, J.E.; Sánchez-Madrid, F.; Esplugues, J.V.; Serrador, J.M. Endothelial nitric oxide synthase regulates T cell receptor signaling at the immunological synapse. Immunity 2006, 24, 753–765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, M.B.; Ju, H.; Venema, V.J.; Liang, H.; Zou, R.; Michell, B.J.; Chen, Z.P.; Kemp, B.E.; Venema, R.C. Reciprocal phosphorylation and regulation of endothelial nitric-oxide synthase in response to bradykinin stimulation. J. Biol. Chem. 2001, 276, 16587–16591. [Google Scholar] [CrossRef] [Green Version]
- Michell, B.J.; Chen, Z.P.; Tiganis, T.; Stapleton, D.; Katsis, F.; Power, D.A.; Sim, A.T.; Kemp, B.E. Coordinated control of endothelial nitric-oxide synthase phosphorylation by protein kinase C and the cAMP-dependent protein kinase. J. Biol. Chem. 2001, 276, 17625–17628. [Google Scholar] [CrossRef] [Green Version]
- Schmitt, C.A.; Heiss, E.H.; Aristei, Y.; Severin, T.; Dirsch, V.M. Norfuraneol dephosphorylates eNOS at threonine 495 and enhances eNOS activity in human endothelial cells. Cardiovasc. Res. 2009, 81, 750–757. [Google Scholar] [CrossRef] [Green Version]
- Heyeraas, K.J. Pulpal, microvascular, and tissue pressure. J. Dent. Res. 1985, 64, 585–589. [Google Scholar] [CrossRef] [PubMed]
- Heyeraas, K.J. Pulpal hemodynamics and interstitial fluid pressure: Balance of transmicrovascular fluid transport. J. Endod. 1989, 15, 468–472. [Google Scholar] [CrossRef]
- Kim, S. Neurovascular interactions in the dental pulp in health and inflammation. J. Endod. 1990, 16, 48–53. [Google Scholar] [CrossRef]
- Heyeraas, K.J.; Kvinnsland, I. Tissue pressure and blood flow in pulpal inflammation. Proc. Finn. Dent. Soc. 1992, 88, 393–401. [Google Scholar] [PubMed]
- Heyeraas, K.J.; Berggreen, E. Interstitial fluid pressure in normal and inflamed pulp. Crit. Rev. Oral. Biol. Med. 1999, 10, 328–336. [Google Scholar] [CrossRef] [Green Version]
- Berggreen, E.; Bletsa, A.; Heyeraas, K.J. Circulation in normal and inflamed dental pulp. Endod. Top. 2010, 17, 2–11. [Google Scholar] [CrossRef]
- Shay-Salit, A.; Shusy, M.; Wolfovitz, E.; Yahav, H.; Breviario, F.; Dejana, E.; Resnick, N. VEGF receptor 2 and the adherens junction as a mechanical transducer in vascular endothelial cells. Proc. Natl. Acad. Sci. USA 2002, 99, 9462–9467. [Google Scholar] [CrossRef] [Green Version]
- Sieminski, A.L.; Hebbel, R.P.; Gooch, K.J. The relative magnitudes of endothelial force generation and matrix stiffness modulate capillary morphogenesis in vitro. Exp. Cell Res. 2004, 297, 574–584. [Google Scholar] [CrossRef]
- Dimmeler, S.; Fleming, I.; Fisslthaler, B.; Hermann, C.; Busse, R.; Zeiher, A.M. Activation of nitric oxide synthase in endothelial cells by Akt-dependent phosphorylation. Nature 1999, 399, 60–65. [Google Scholar] [CrossRef]
- Fleming, I.; Fisslthaler, B.; Dimmeler, S.; Kemp, B.E.; Busse, R. Phosphorylation of Thr(495) regulates Ca(2+)/calmodulin-dependent endothelial nitric oxide synthase activity. Circ. Res. 2001, 88, E68–E75. [Google Scholar] [CrossRef] [Green Version]
- Lin, M.I.; Fulton, D.; Babbitt, R.; Fleming, I.; Busse, R.; Pritchard, K.A., Jr.; Sessa, W.C. Phosphorylation of threonine 497 in endothelial nitric-oxide synthase coordinates the coupling of L-arginine metabolism to efficient nitric oxide production. J. Biol. Chem. 2003, 278, 44719–44726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Zhu, J.; Li, B.; Gao, X.; Liu, Y.; Mao, W.; Chen, S.L. Resveratrol ameliorates low shear stress induced oxidative stress by suppressing ERK/eNOS Thr495 in endothelial cells. Mol. Med. Rep. 2014, 10, 1964–1972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Zhang, J.; Wang, Z.; Chen, S. Ivabradine prevents low shear stress induced endothelial inflammation and oxidative stress via mTOR/eNOS pathway. PLoS ONE 2016, 11, e0149694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, X.; Qu, X.; Li, B.; Wang, Z.; Chao, Y.; Jiang, X.; Wu, W.; Chen, S.L. Modulation of low shear stress-induced eNOS multi-site phosphorylation and nitric oxide production via protein kinase and ERK1/2 signaling. Mol. Med. Rep. 2017, 15, 908–914. [Google Scholar] [CrossRef]
- Chao, Y.; Ye, P.; Kong, X.; Qu, X.; Zhang, J.; Luo, J.; Yang, H.; Chen, S. Low shear stress induces endothelial reactive oxygen species via the AT1R/eNOS/NO pathway. J. Cell Physiol. 2018, 233, 1384–1395. [Google Scholar] [CrossRef]
- Takahashi, K. Vascular architecture of dog pulp using corrosion resin cast examined under a scanning electron microscope. J. Dent. Res. 1985, 64, 579–584. [Google Scholar] [CrossRef]
- Kim, S. Microcirculation of the dental pulp in health and disease. J. Endod. 1985, 11, 465–471. [Google Scholar] [CrossRef]
- Yoshida, S.; Ohshima, H. Distribution and organization of peripheral capillaries in dental pulp and their relationship to odontoblasts. Anat. Record 1996, 245, 313–326. [Google Scholar] [CrossRef]
- Cooper, P.R.; Smith, A.J. Molecular mediators of pulp inflammation and regeneration. Endod. Top. 2013, 28, 90–105. [Google Scholar] [CrossRef]
- Mittal, M.; Siddiqui, M.R.; Tran, K.; Reddy, S.P.; Malik, A.B. Reactive oxygen species in inflammation and tissue injury. Antioxid. Redox Signal. 2014, 20, 1126–1167. [Google Scholar] [CrossRef] [Green Version]
- Baumgardner, K.R.; Sulfaro, M.A. The anti-inflammatory effects of human recombinant copper-zinc superoxide dismutase on pulp inflammation. J. Endod. 2001, 27, 190–195. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Erdek, Ö.; Bloch, W.; Rink-Notzon, S.; Roggendorf, H.C.; Uzun, S.; Meul, B.; Koch, M.; Neugebauer, J.; Deschner, J.; Korkmaz, Y. Inflammation of the Human Dental Pulp Induces Phosphorylation of eNOS at Thr495 in Blood Vessels. Biomedicines 2022, 10, 1586. https://doi.org/10.3390/biomedicines10071586
Erdek Ö, Bloch W, Rink-Notzon S, Roggendorf HC, Uzun S, Meul B, Koch M, Neugebauer J, Deschner J, Korkmaz Y. Inflammation of the Human Dental Pulp Induces Phosphorylation of eNOS at Thr495 in Blood Vessels. Biomedicines. 2022; 10(7):1586. https://doi.org/10.3390/biomedicines10071586
Chicago/Turabian StyleErdek, Özlem, Wilhelm Bloch, Svenja Rink-Notzon, Hubert C. Roggendorf, Senem Uzun, Britta Meul, Manuel Koch, Jörg Neugebauer, James Deschner, and Yüksel Korkmaz. 2022. "Inflammation of the Human Dental Pulp Induces Phosphorylation of eNOS at Thr495 in Blood Vessels" Biomedicines 10, no. 7: 1586. https://doi.org/10.3390/biomedicines10071586
APA StyleErdek, Ö., Bloch, W., Rink-Notzon, S., Roggendorf, H. C., Uzun, S., Meul, B., Koch, M., Neugebauer, J., Deschner, J., & Korkmaz, Y. (2022). Inflammation of the Human Dental Pulp Induces Phosphorylation of eNOS at Thr495 in Blood Vessels. Biomedicines, 10(7), 1586. https://doi.org/10.3390/biomedicines10071586