
Purkinje Cell Activity Resonation Generates Rhythmic Behaviors at the Preferred Frequency of 8 Hz

 , , , ,
, , , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Data and code availability
2.2. Experimental Model and Subject Details
Mice
2.3. Method Details
2.3.1. Surgeries
2.3.2. Whisker Movement Recording and Tracking
2.3.3. Electrophysiology
2.3.4. Viral Injections
2.3.5. Histology and Microscopy
2.3.6. Optogenetic Stimulation
2.4. Quantification and Statistical Analysis
2.4.1. Experimental Paradigm
2.4.2. Simple Spike Analysis
2.4.3. Whisker Analysis
3. Results
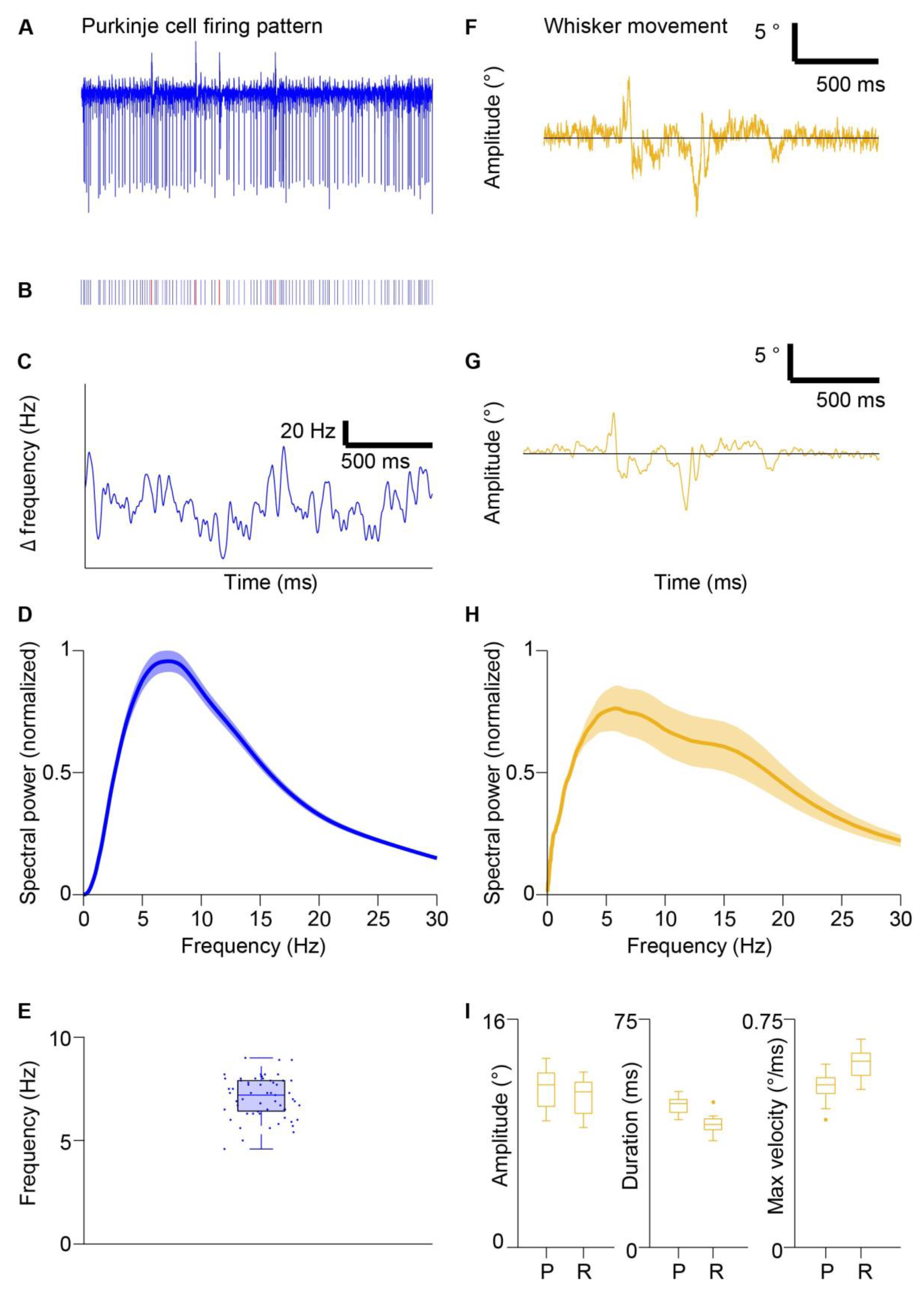
3.1. Oscillatory Simple Spike Frequency and Whisking Kinematics
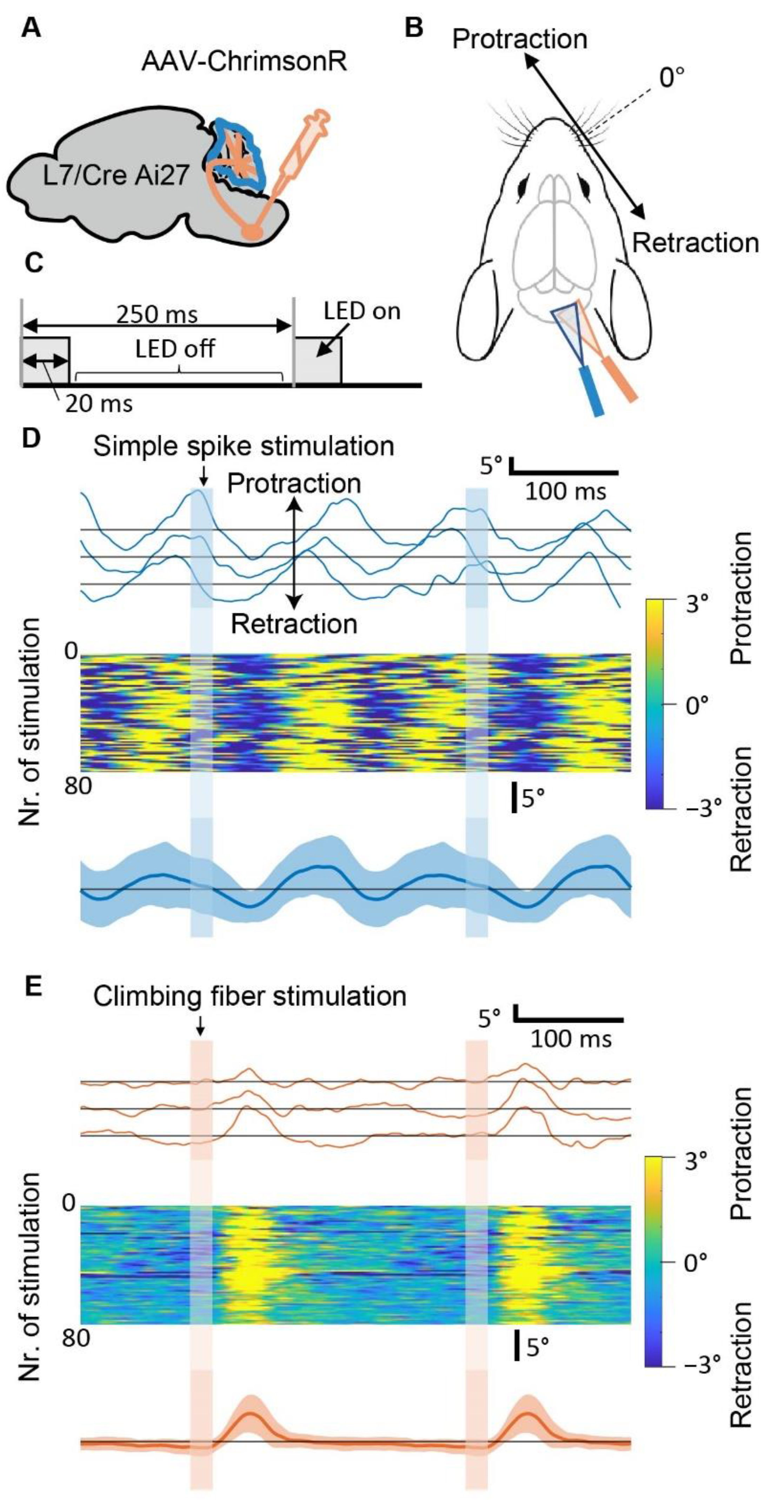
3.2. Optogenetic Stimulation of Purkinje Cells can Induce Specific Patterns of Whisker Movement
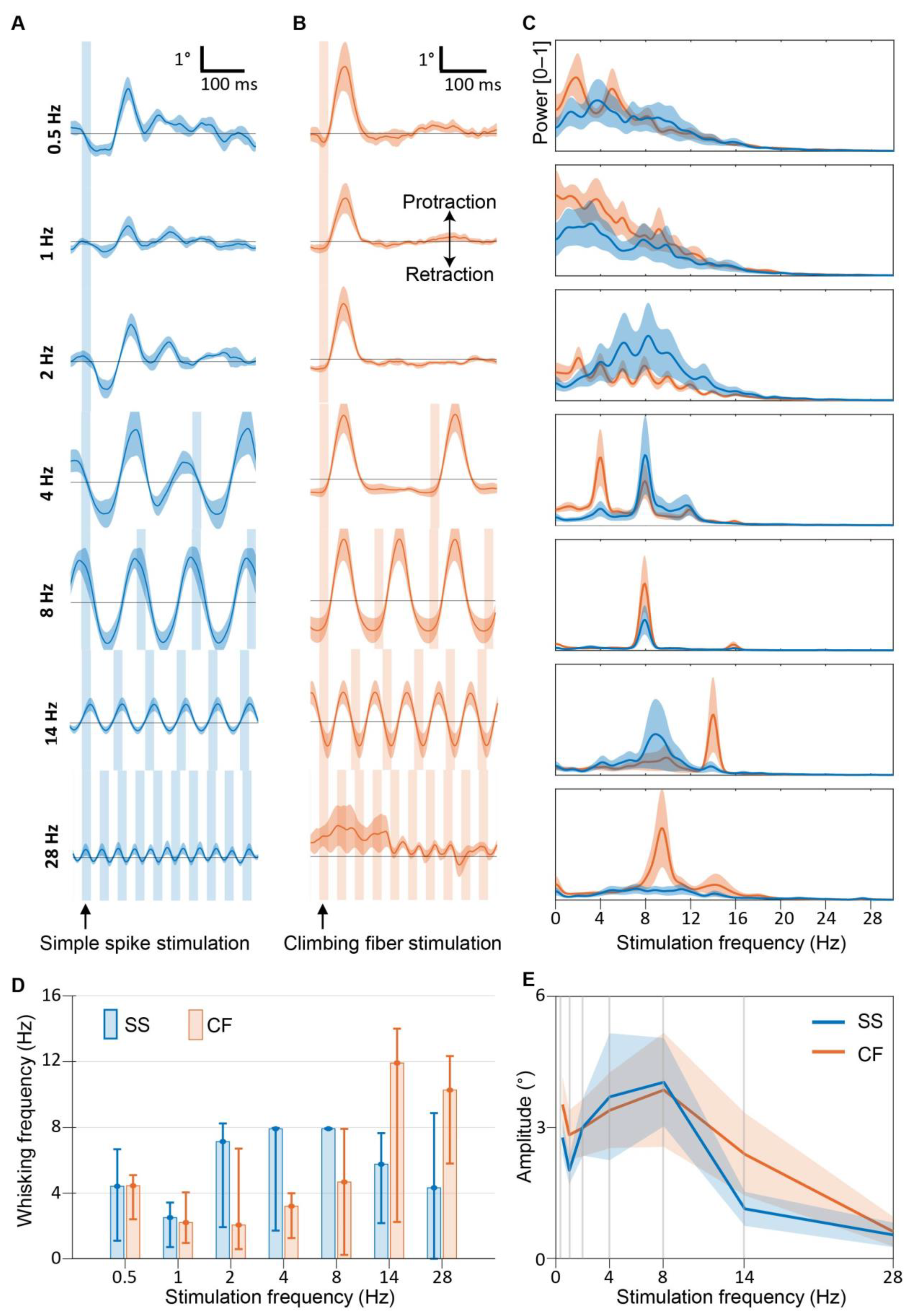
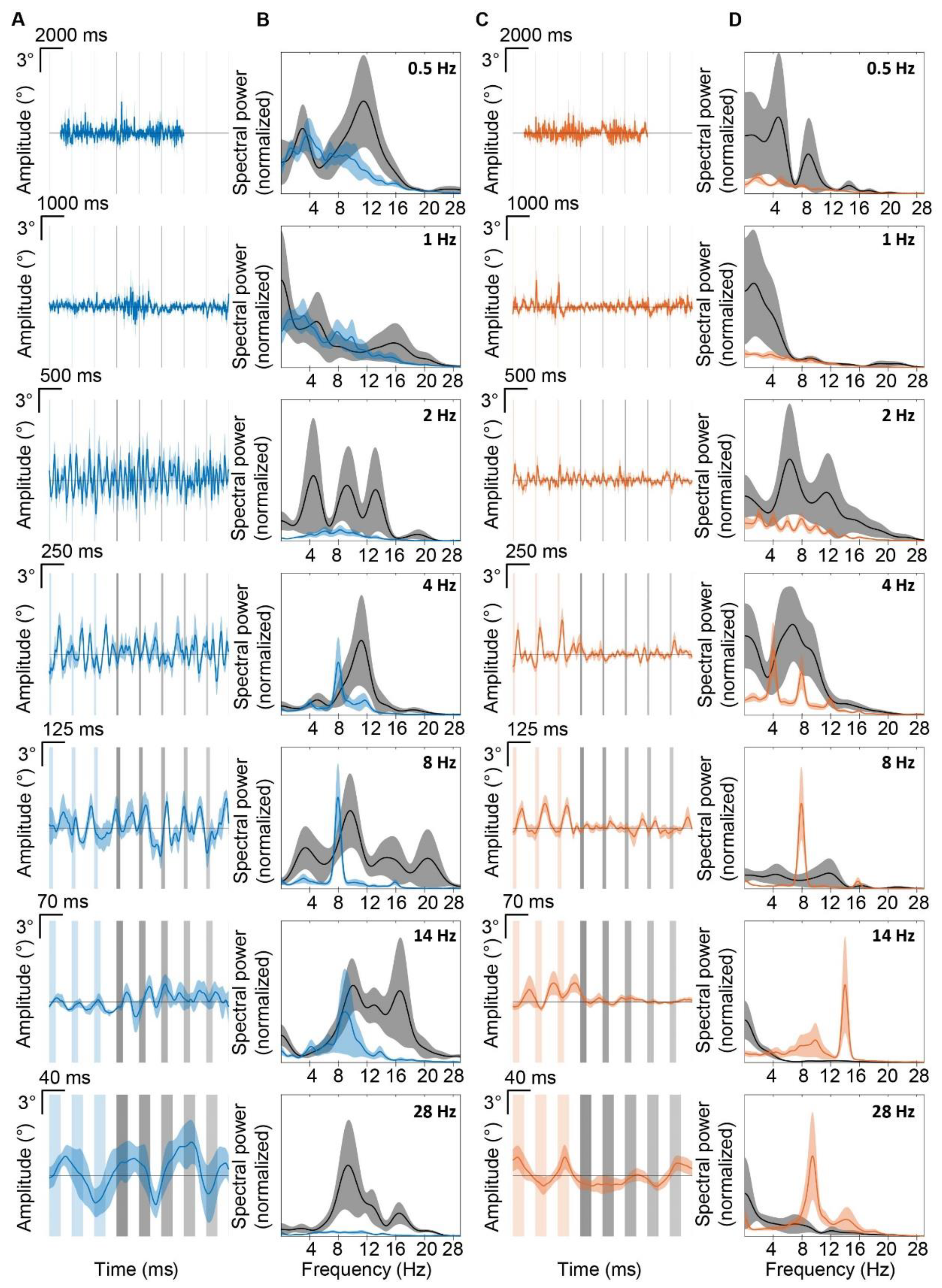
3.3. Purkinje Cell Activity Induces Whisker Movements at the Preferred Frequency of 8 Hz
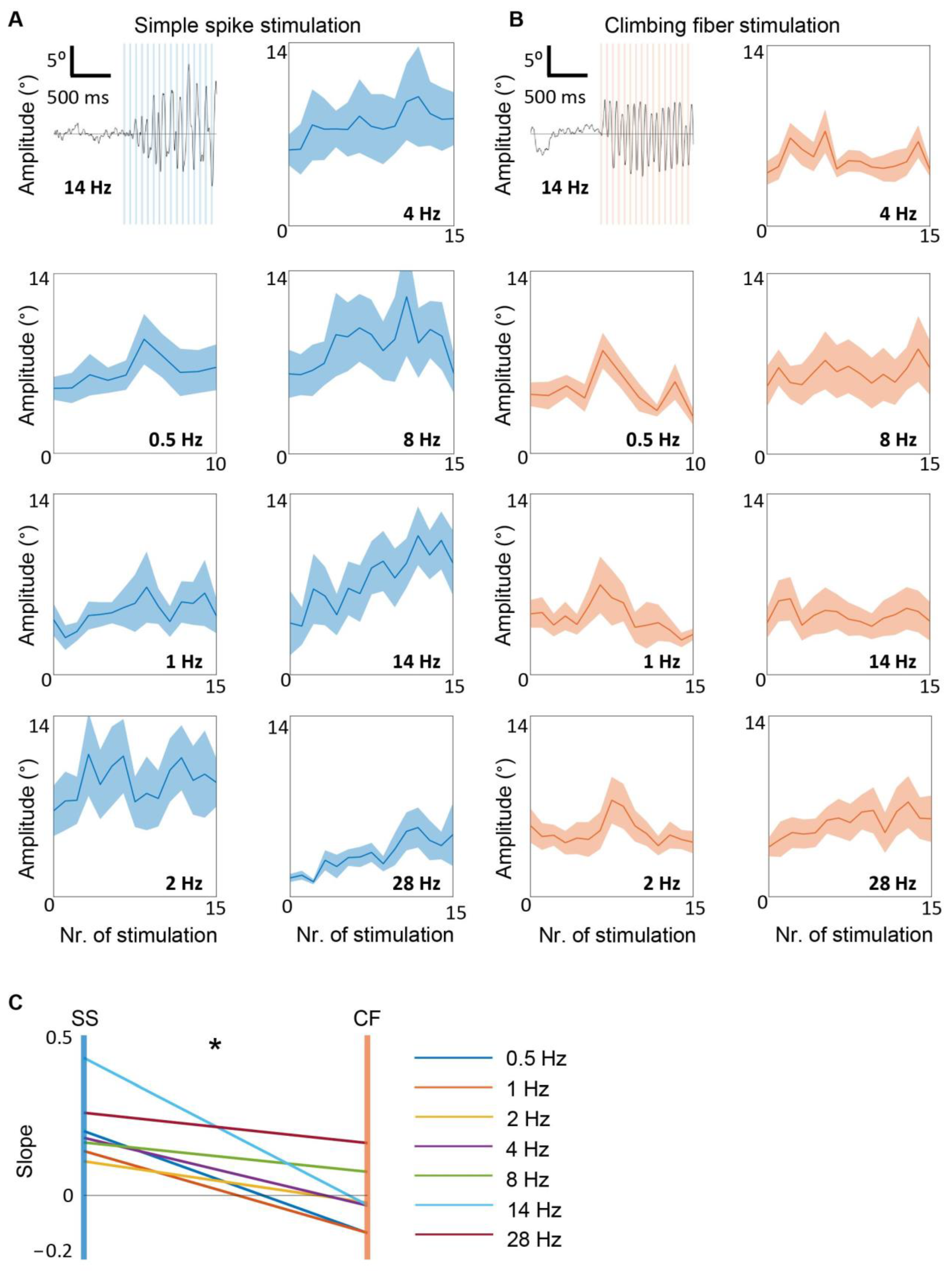
3.4. Simple Spike, but Not Climbing Fiber Stimulation, Entrains Whisker Movement
4. Discussion
4.1. Simple Spike Activation of Reverberating Circuits Sustaining Rhythmic Whisks
4.2. Multiple Cerebellar–Whisker Pathways
4.3. The Olivocerebellar Contribution to Rhythmic Whisking
4.4. Olivocerebellar Rhythmic Activity beyond Mouse Whisking
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Zeeuw, C.I.; Wylie, D.R.; Stahl, J.S.; Simpson, J.I. Phase relations of Purkinje cells in the rabbit flocculus during compensatory eye movements. J. Neurophysiol. 1995, 74, 2051–2064. [Google Scholar] [CrossRef] [PubMed]
- Welsh, J.P.; Lang, E.J.; Suglhara, I.; Llinas, R. Dynamic organization of motor control within the olivocerebellar system. Nature 1995, 374, 453–457. [Google Scholar] [CrossRef] [PubMed]
- Morton, S.M.; Bastian, A.J. Cerebellar control of balance and locomotion. Neuroscientist 2004, 10, 247–259. [Google Scholar] [CrossRef] [PubMed]
- Schmahmann, J.D. Disorders of the cerebellum: Ataxia, dysmetria of thought, and the cerebellar cognitive affective syndrome. J. Neuropsychiatry Clin. Neurosci. 2004, 16, 367–378. [Google Scholar] [CrossRef] [PubMed]
- Bosman, L.W.; Koekkoek, S.K.E.; Shapiro, J.; Rijken, B.; Zandstra, F.; Van Der Ende, B.; Owens, C.; Potters, J.W.; De Gruijl, J.; Ruigrok, T.; et al. Encoding of whisker input by cerebellar Purkinje cells. J. Physiol. 2010, 588, 3757–3783. [Google Scholar] [CrossRef] [PubMed]
- Witter, L.; Canto, C.B.; Hoogland, T.M.; de Gruijl, J.R.; de Zeeuw, C.I. Strength and timing of motor responses mediated by rebound firing in the cerebellar nuclei after Purkinje cell activation. Front. Neural Circuits 2013, 7, 133. [Google Scholar] [CrossRef] [PubMed]
- Heiney, S.A.; Kim, J.; Augustine, G.J.; Medina, J.F. Precise control of movement kinematics by optogenetic inhibition of Purkinje cell activity. J. Neurosci. 2014, 34, 2321–2330. [Google Scholar] [CrossRef]
- Rahmati, N.; Owens, C.B.; Bosman, L.; Spanke, J.K.; Lindeman, S.; Gong, W.; Potters, J.-W.; Romano, V.; Voges, K.; Moscato, L.; et al. Cerebellar potentiation and learning a whisker-based object localization task with a time response window. J. Neurosci. 2014, 34, 1949–1962. [Google Scholar] [CrossRef]
- Herzfeld, D.J.; Kojima, Y.; Soetedjo, R.; Shadmehr, R. Encoding of action by the Purkinje cells of the cerebellum. Nature 2015, 526, 439–442. [Google Scholar] [CrossRef]
- Hoogland, T.M.; de Gruijl, J.R.; Witter, L.; Canto, C.B.; de Zeeuw, C.I. Role of Synchronous Activation of Cerebellar Purkinje Cell Ensembles in Multi-joint Movement Control. Curr. Biol. 2015, 25, 1157–1165. [Google Scholar] [CrossRef]
- Chen, S.; Augustine, G.J.; Chadderton, P. The cerebellum linearly encodes whisker position during voluntary movement. Elife 2016, 5, e10509. [Google Scholar] [CrossRef]
- White, J.J.; Sillitoe, R.V. Genetic silencing of olivocerebellar synapses causes dystonia-like behaviour in mice. Nat. Commun. 2017, 8, 14912. [Google Scholar] [CrossRef]
- Brown, S.T.; Raman, I.M. Sensorimotor Integration and Amplification of Reflexive Whisking by Well-Timed Spiking in the Cerebellar Corticonuclear Circuit. Neuron 2018, 99, 564–575.e2. [Google Scholar] [CrossRef]
- Gao, Z.; Davis, C.; Thomas, A.M.; Economo, M.; Abrego, A.M.; Svoboda, K.; De Zeeuw, C.I.; Li, N. A cortico-cerebellar loop for motor planning. Nature 2018, 563, 113–116. [Google Scholar] [CrossRef]
- Romano, V.; Reddington, A.L.; Cazzanelli, S.; Mazza, R.; Ma, Y.; Strydis, C.; Negrello, M.; Bosman, L.W.; De Zeeuw, C.I. Functional Convergence of Autonomic and Sensorimotor Processing in the Lateral Cerebellum. Cell Rep. 2020, 32, 107867. [Google Scholar] [CrossRef]
- Tsutsumi, S.; Chadney, O.; Yiu, T.-L.; Bäumler, E.; Faraggiana, L.; Beau, M.; Häusser, M. Purkinje Cell Activity Determines the Timing of Sensory-Evoked Motor Initiation. Cell Rep. 2020, 33, 108537. [Google Scholar] [CrossRef]
- De Zeeuw, C.I. Bidirectional learning in upbound and downbound microzones of the cerebellum. Nat. Rev. Neurosci. 2021, 22, 92–110. [Google Scholar] [CrossRef]
- Reeber, S.L.; Otis, T.S.; Sillitoe, R.V. New roles for the cerebellum in health and disease. Front. Syst. Neurosci. 2013, 7, 83. [Google Scholar] [CrossRef]
- Geschwind, D.H.; Perlman, S.; Figueroa, C.P.; Treiman, L.J.; Pulst, S.M. The prevalence and wide clinical spectrum of the spinocerebellar ataxia type 2 trinucleotide repeat in patients with autosomal dominant cerebellar ataxia. Am. J. Hum. Genet. 1997, 60, 842–850. [Google Scholar]
- Smeets, C.J.; Verbeek, D.S. Climbing fibers in spinocerebellar ataxia: A mechanism for the loss of motor control. Neurobiol. Dis. 2016, 88, 96–106. [Google Scholar] [CrossRef]
- D’Angelo, E.; Koekkoek, S.; Lombardo, P.; Solinas, S.; Ros, E.; Garrido, J.; Schonewille, M.; De Zeeuw, C. Timing in the cerebellum: Oscillations and resonance in the granular layer. Neuroscience 2009, 162, 805–815. [Google Scholar] [CrossRef]
- Hattox, A.M.; Priest, C.A.; Keller, A. Functional circuitry involved in the regulation of whisker movements. J. Comp. Neurol. 2002, 442, 266–276. [Google Scholar] [CrossRef]
- Kleinfeld, D.; Berg, R.W.; O’Connor, S.M. Anatomical loops and their electrical dynamics in relation to whisking by rat. Somatosens. Mot. Res. 1999, 16, 69–88. [Google Scholar] [CrossRef]
- Lang, E.J.; Sugihara, I.; Llinas, R. Olivocerebellar modulation of motor cortex ability to generate vibrissal movements in rat. J. Physiol. 2006, 571, 101–120. [Google Scholar] [CrossRef]
- Teune, T.M.; van der Burg, J.; van der Moer, J.; Voogd, J.; Ruigrok, T.J. Topography of cerebellar nuclear projections to the brain stem in the rat. Prog. Brain Res. 2000, 124, 141–172. [Google Scholar]
- Berretta, S.; Perciavalle, V.; Poppele, R.E. Origin of spinal projections to the anterior and posterior lobes of the rat cerebellum. J. Comp. Neurol. 1991, 305, 273–281. [Google Scholar] [CrossRef]
- Sengul, G.; Fu, Y.; Yu, Y.; Paxinos, G. Spinal cord projections to the cerebellum in the mouse. Brain Struct. Funct. 2015, 220, 2997–3009. [Google Scholar] [CrossRef]
- D’Angelo, E.; Nieus, T.; Maffei, A.; Armano, S.; Rossi, P.; Taglietti, V.; Fontana, A.; Naldi, G. Theta-frequency bursting and resonance in cerebellar granule cells: Experimental evidence and modeling of a slow k+-dependent mechanism. J. Neurosci. 2001, 21, 759–770. [Google Scholar] [CrossRef]
- Gandolfi, D.; Lombardo, P.; Mapelli, J.; Solinas, S.; D’Angelo, E. theta-Frequency resonance at the cerebellum input stage improves spike timing on the millisecond time-scale. Front. Neural Circuits 2013, 7, 64. [Google Scholar] [CrossRef]
- Solinas, S.; Forti, L.; Cesana, E.; Mapelli, J.; De Schutter, E.; D’Angelo, E. Fast-reset of pacemaking and theta-frequency resonance patterns in cerebellar golgi cells: Simulations of their impact in vivo. Front. Cell. Neurosci. 2007, 1, 4. [Google Scholar] [CrossRef]
- D’Angelo, E. The critical role of Golgi cells in regulating spatio-temporal integration and plasticity at the cerebellum input stage. Front. Neurosci. 2008, 2, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Marquez-Ruiz, J.; Cheron, G. Sensory stimulation-dependent plasticity in the cerebellar cortex of alert mice. PLoS ONE 2012, 7, e36184. [Google Scholar] [CrossRef] [PubMed]
- Ramakrishnan, K.B.; Voges, K.; de Propris, L.; de Zeeuw, C.I.; D’Angelo, E. Tactile Stimulation Evokes Long-Lasting Potentiation of Purkinje Cell Discharge In Vivo. Front. Cell. Neurosci. 2016, 10, 36. [Google Scholar] [CrossRef] [PubMed]
- Roggeri, L.; Rivieccio, B.; Rossi, P.; D’Angelo, E. Tactile stimulation evokes long-term synaptic plasticity in the granular layer of cerebellum. J. Neurosci. 2008, 28, 6354–6359. [Google Scholar] [CrossRef]
- Chadderton, P.; Margrie, T.W.; Hausser, M. Integration of quanta in cerebellar granule cells during sensory processing. Nature 2004, 428, 856–860. [Google Scholar] [CrossRef]
- Romano, V.; De Propris, L.; Bosman, L.W.; Warnaar, P.; Brinke, M.M.T.; Lindeman, S.; Ju, C.; Velauthapillai, A.; Spanke, J.K.; Guerra, E.M.; et al. Potentiation of cerebellar Purkinje cells facilitates whisker reflex adaptation through increased simple spike activity. Elife 2018, 7, e38852. [Google Scholar] [CrossRef]
- De Zeeuw, C.I.; Hoebeek, F.E.; Bosman, L.; Schonewille, M.; Witter, L.; Koekkoek, S.K. Spatiotemporal firing patterns in the cerebellum. Nat. Rev. Neurosci. 2011, 12, 327–344. [Google Scholar] [CrossRef]
- Bazzigaluppi, P.; De Gruijl, J.R.; van der Giessen, R.S.; Khosrovani, S.; De Zeeuw, C.I.; de Jeu, M.T.G. Olivary subthreshold oscillations and burst activity revisited. Front. Neural Circuits 2012, 6, 91. [Google Scholar] [CrossRef]
- De Gruijl, J.R.; Bazzigaluppi, P.; de Jeu, M.T.; de Zeeuw, C.I. Climbing fiber burst size and olivary sub-threshold oscillations in a network setting. PLoS Comput. Biol. 2012, 8, e1002814. [Google Scholar] [CrossRef]
- Lang, E.J. Excitatory afferent modulation of complex spike synchrony. Cerebellum 2003, 2, 165–170. [Google Scholar] [CrossRef]
- Khosrovani, S.; van der Giessen, R.S.; de Zeeuw, C.I.; de Jeu, M.T. In vivo mouse inferior olive neurons exhibit heterogeneous subthreshold oscillations and spiking patterns. Proc. Natl. Acad. Sci. USA 2007, 104, 15911–15916. [Google Scholar] [CrossRef]
- Llinas, R.; Yarom, Y. Oscillatory properties of guinea-pig inferior olivary neurones and their pharmacological modulation: An in vitro study. J. Physiol. 1986, 376, 163–182. [Google Scholar] [CrossRef]
- Negrello, M.; Warnaar, P.; Romano, V.; Owens, C.B.; Lindeman, S.; Iavarone, E.; Spanke, J.K.; Bosman, L.W.J.; De Zeeuw, C.I. Quasiperiodic rhythms of the inferior olive. PLoS Comput. Biol. 2019, 15, e1006475. [Google Scholar] [CrossRef]
- Van der Giessen, R.S.; Koekkoek, S.K.; van Dorp, S.; De Gruijl, J.R.; Cupido, A.; Khosrovani, S.; Dortland, B.; Wellershaus, K.; Degen, J.; Deuchars, J.; et al. Role of olivary electrical coupling in cerebellar motor learning. Neuron 2008, 58, 599–612. [Google Scholar] [CrossRef]
- Hutcheon, B.; Yarom, Y. Resonance, oscillation and the intrinsic frequency preferences of neurons. Trends Neurosci. 2000, 23, 216–222. [Google Scholar] [CrossRef]
- Moscato, L.; Montagna, I.; De Propris, L.; Tritto, S.; Mapelli, L.; D’Angelo, E. Long-Lasting Response Changes in Deep Cerebellar Nuclei in vivo Correlate With Low-Frequency Oscillations. Front. Cell Neurosci. 2019, 13, 84. [Google Scholar] [CrossRef]
- Perkon, I.; Kosir, A.; Itskov, P.M.; Tasic, J.; Diamond, M.E. Unsupervised quantification of whisking and head movement in freely moving rodents. J. Neurophysiol. 2011, 105, 1950–1962. [Google Scholar] [CrossRef]
- Betting, J.L.F.; Romano, V.; Al-Ars, Z.; Bosman, L.W.J.; Strydis, C.; De Zeeuw, C.I. WhiskEras: A New Algorithm for Accurate Whisker Tracking. Front. Cell Neurosci. 2020, 14, 588445. [Google Scholar] [CrossRef]
- Romano, V.; Zhai, P.; van der Horst, A.; Mazza, R.; Jacobs, T.; Bauer, S.; Wang, X.; White, J.J.; De Zeeuw, C. Olivocerebellar control of movement symmetry. Curr. Biol. 2022, 32, 654–670.e4. [Google Scholar] [CrossRef]
- Cao, Y.; Roy, S.; Sachdev, R.N.; Heck, D.H. Dynamic correlation between whisking and breathing rhythms in mice. J. Neurosci. 2012, 32, 1653–1659. [Google Scholar] [CrossRef]
- Kleinfeld, D.; Deschenes, M.; Ulanovsky, N. Whisking, Sniffing, and the Hippocampal theta-Rhythm: A Tale of Two Oscillators. PLoS Biol. 2016, 14, e1002385. [Google Scholar] [CrossRef]
- Moore, J.D.; Deschênes, M.; Furuta, T.; Huber, D.; Smear, M.C.; Demers, M.; Kleinfeld, D. Hierarchy of orofacial rhythms revealed through whisking and breathing. Nature 2013, 497, 205–210. [Google Scholar] [CrossRef]
- Proville, R.D.; Spolidoro, M.; Guyon, N.; Dugué, G.P.; Selimi, F.; Isope, P.; Popa, D.; Léna, C.; Proville, R.D.; Selimi, F.; et al. Cerebellum involvement in cortical sensorimotor circuits for the control of voluntary movements. Nat. Neurosci. 2014, 17, 1233–1239. [Google Scholar] [CrossRef]
- Pellerin, J.P.; Lamarre, Y. Local field potential oscillations in primate cerebellar cortex during voluntary movement. J. Neurophysiol. 1997, 78, 3502–3507. [Google Scholar] [CrossRef]
- Marshall, S.P.; Lang, E.J. Inferior olive oscillations gate transmission of motor cortical activity to the cerebellum. J. Neurosci. 2004, 24, 11356–11367. [Google Scholar] [CrossRef]
- Kistler, W.M.; de Zeeuw, C.I. Time windows and reverberating loops: A reverse-engineering approach to cerebellar function. Cerebellum 2003, 2, 44–54. [Google Scholar] [CrossRef]
- Bellavance, M.A.; Takatoh, J.; Lu, J.; Demers, M.; Kleinfeld, D.; Wang, F.; Deschênes, M. Parallel Inhibitory and Excitatory Trigemino-Facial Feedback Circuitry for Reflexive Vibrissa Movement. Neuron 2017, 95, 722–723. [Google Scholar] [CrossRef]
- Komiyama, M.; Shibata, H.; Suzuki, T. Somatotopic representation of facial muscles within the facial nucleus of the mouse. A study using the retrograde horseradish peroxidase and cell degeneration techniques. Brain Behav. Evol. 1984, 24, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Bosman, L.W.; Houweling, A.R.; Owens, C.B.; Tanke, N.; Shevchouk, O.T.; Rahmati, N.; Teunissen, W.H.T.; Ju, C.; Gong, W.; Koekkoek, S.K.E.; et al. Anatomical pathways involved in generating and sensing rhythmic whisker movements. Front. Integr. Neurosci. 2011, 5, 53. [Google Scholar] [CrossRef] [PubMed]
- Gao, P.; Bermejo, R.; Zeigler, H.P. Whisker deafferentation and rodent whisking patterns: Behavioral evidence for a central pattern generator. J. Neurosci. 2001, 21, 5374–5380. [Google Scholar] [CrossRef] [PubMed]
- Lindeman, S.; Hong, S.; Kros, L.; De Zeeuw, C.I. Cerebellar Purkinje cells can differentially modulate coherence between sensory and motor cortex depending on region and behavior. Proc. Natl. Acad. Sci. USA 2021, 118, e2015292118. [Google Scholar] [CrossRef]
- Chaumont, J.; Guyon, N.; Valera, A.M.; Dugué, G.P.; Popa, D.; Marcaggi, P.; Gautheron, V.; Reibel-Foisset, S.; Dieudonné, S.; Stephan, A.; et al. Clusters of cerebellar Purkinje cells control their afferent climbing fiber discharge. Proc. Natl. Acad. Sci. USA 2013, 110, 16223–16228. [Google Scholar] [CrossRef]
- Wang, X.; Novello, M.; Gao, Z.; Ruigrok, T.J.H.; de Zeeuw, C.I. Input and output organization of the mesodiencephalic junction for cerebro-cerebellar communication. J. Neurosci. Res. 2022, 100, 620–637. [Google Scholar] [CrossRef]
- Buzsaki, G.; Draguhn, A. Neuronal oscillations in cortical networks. Science 2004, 304, 1926–1929. [Google Scholar] [CrossRef]
- Van Dun, K.; Bodranghien, F.C.; Marien, P.; Manto, M.U. tDCS of the Cerebellum: Where Do We Stand in 2016? Technical Issues and Critical Review of the Literature. Front. Hum. Neurosci. 2016, 10, 199. [Google Scholar] [CrossRef]
- Hooper, J.; Simpson, P.; Whittle, I.R. Chronic posttraumatic movement disorder alleviated by insertion of meso-diencephalic deep brain stimulating electrode. Br. J. Neurosurg. 2001, 15, 435–438. [Google Scholar] [CrossRef]
- Turner, R.C.; VanGilder, R.L.; Naser, Z.J.; Lucke-Wold, B.P.; Bailes, J.E.; Matsumoto, R.R. Elucidating the severity of preclinical traumatic brain injury models: A role for functional assessment? Neurosurgery 2014, 74, 382–394; discussion 394. [Google Scholar] [CrossRef]
- Webster, B.R.; Celnik, P.A.; Cohen, L.G. Noninvasive brain stimulation in stroke rehabilitation. NeuroRX 2006, 3, 474–481. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bauer, S.; van Wingerden, N.; Jacobs, T.; van der Horst, A.; Zhai, P.; Betting, J.-H.L.F.; Strydis, C.; White, J.J.; De Zeeuw, C.I.; Romano, V. Purkinje Cell Activity Resonation Generates Rhythmic Behaviors at the Preferred Frequency of 8 Hz. Biomedicines 2022, 10, 1831. https://doi.org/10.3390/biomedicines10081831
Bauer S, van Wingerden N, Jacobs T, van der Horst A, Zhai P, Betting J-HLF, Strydis C, White JJ, De Zeeuw CI, Romano V. Purkinje Cell Activity Resonation Generates Rhythmic Behaviors at the Preferred Frequency of 8 Hz. Biomedicines. 2022; 10(8):1831. https://doi.org/10.3390/biomedicines10081831
Chicago/Turabian StyleBauer, Staf, Nathalie van Wingerden, Thomas Jacobs, Annabel van der Horst, Peipei Zhai, Jan-Harm L. F. Betting, Christos Strydis, Joshua J. White, Chris I. De Zeeuw, and Vincenzo Romano. 2022. "Purkinje Cell Activity Resonation Generates Rhythmic Behaviors at the Preferred Frequency of 8 Hz" Biomedicines 10, no. 8: 1831. https://doi.org/10.3390/biomedicines10081831
APA StyleBauer, S., van Wingerden, N., Jacobs, T., van der Horst, A., Zhai, P., Betting, J. -H. L. F., Strydis, C., White, J. J., De Zeeuw, C. I., & Romano, V. (2022). Purkinje Cell Activity Resonation Generates Rhythmic Behaviors at the Preferred Frequency of 8 Hz. Biomedicines, 10(8), 1831. https://doi.org/10.3390/biomedicines10081831