
TGF-Beta Modulates the Integrity of the Blood Brain Barrier In Vitro, and Is Associated with Metabolic Alterations in Pericytes
 , , , , and
, , , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Cell Culture
2.2. Isolation of Primary Porcine Brain Microvascular Endothelial Cells
2.3. The In Vitro BBB Model
2.4. Immunocytochemistry and Immunofluorescence
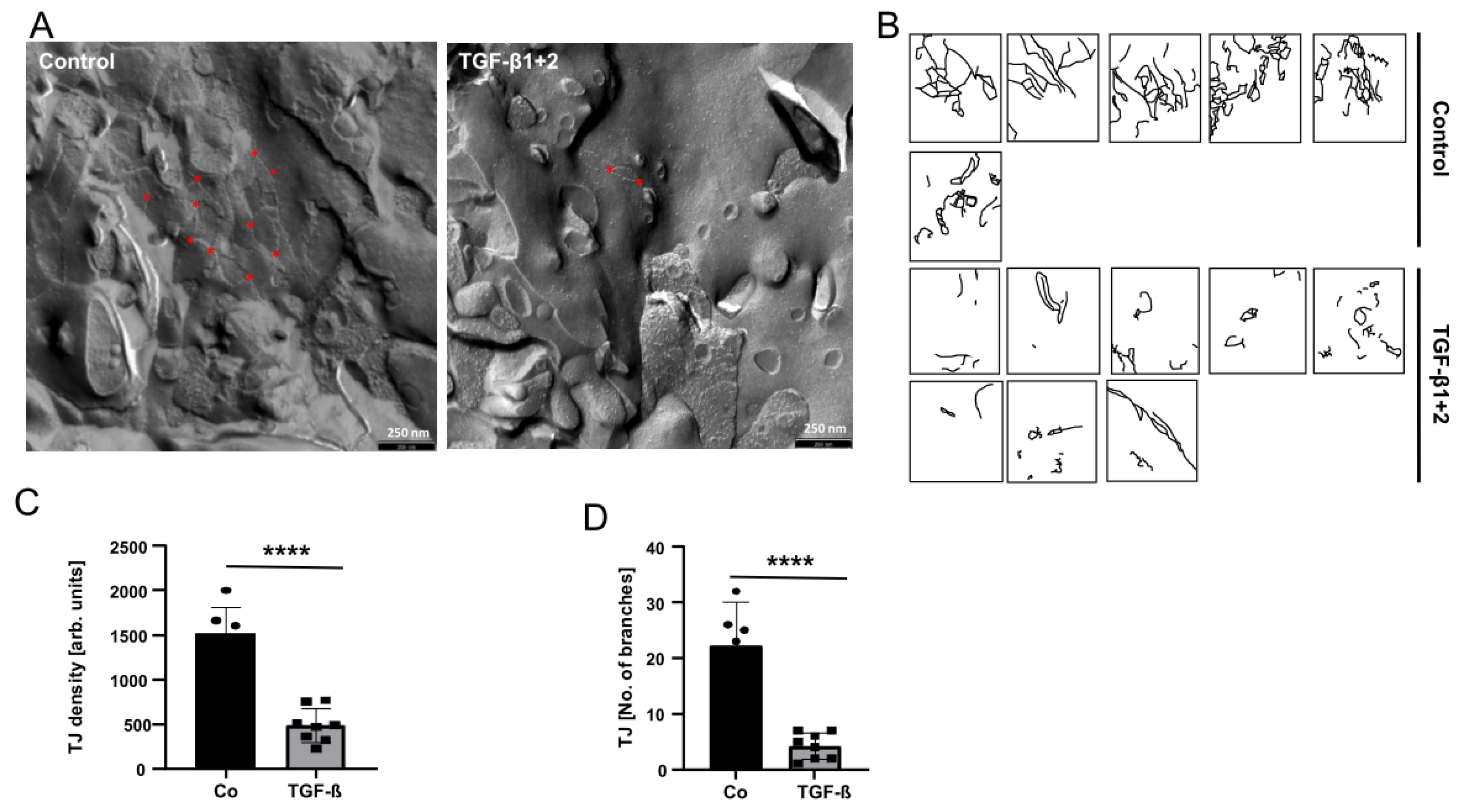
2.5. Freeze Fracture Electron Microscopy
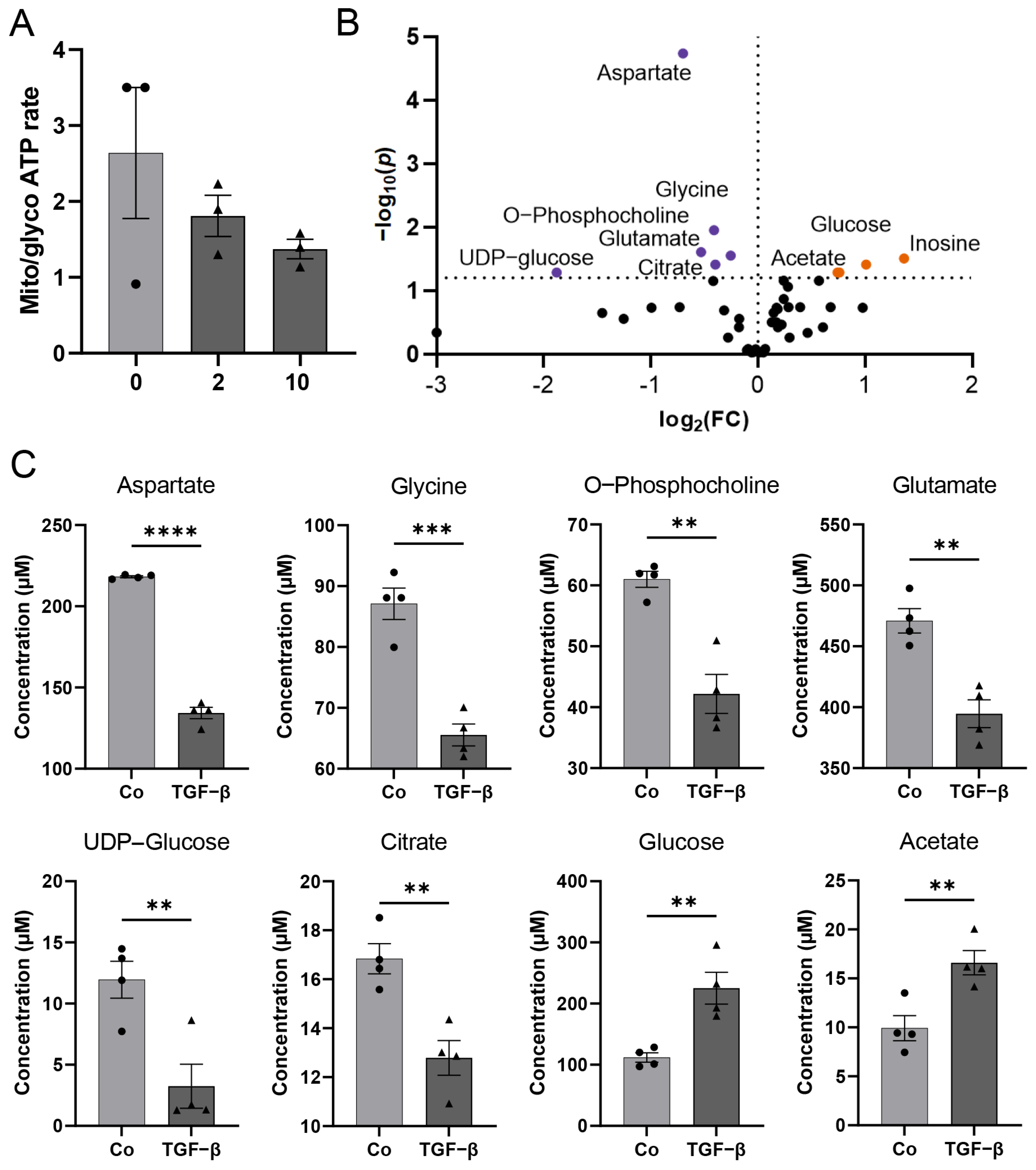
2.6. Metabolic Analysis
2.7. 1H-NMR Spectroscopy-Based Metabolomics of HBVP Intracellular Metabolites
2.8. RNA Sequencing
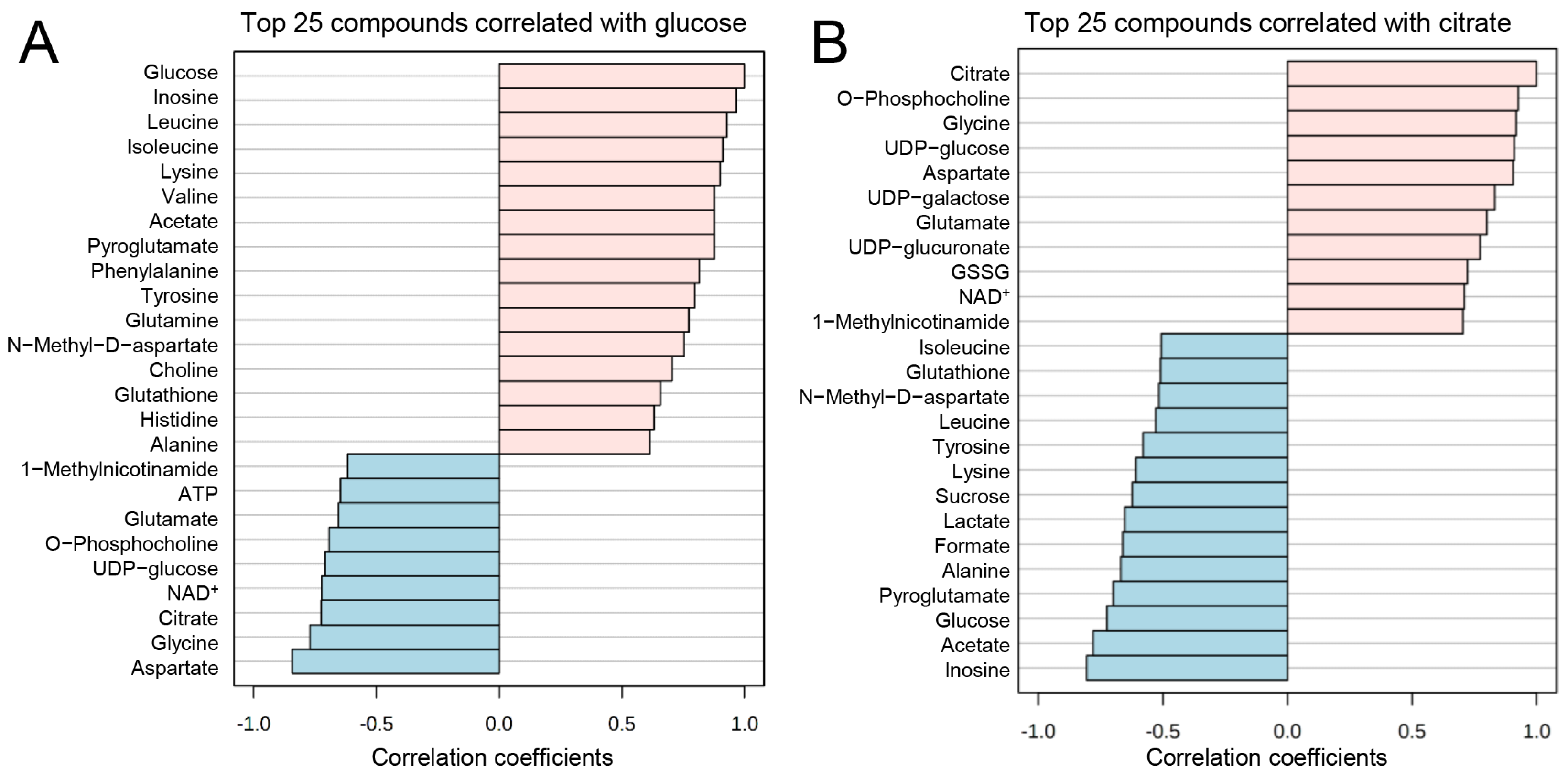
2.9. Combined Metabolomics and RNA-Seq Pathway Analysis
2.10. Statistics
3. Results
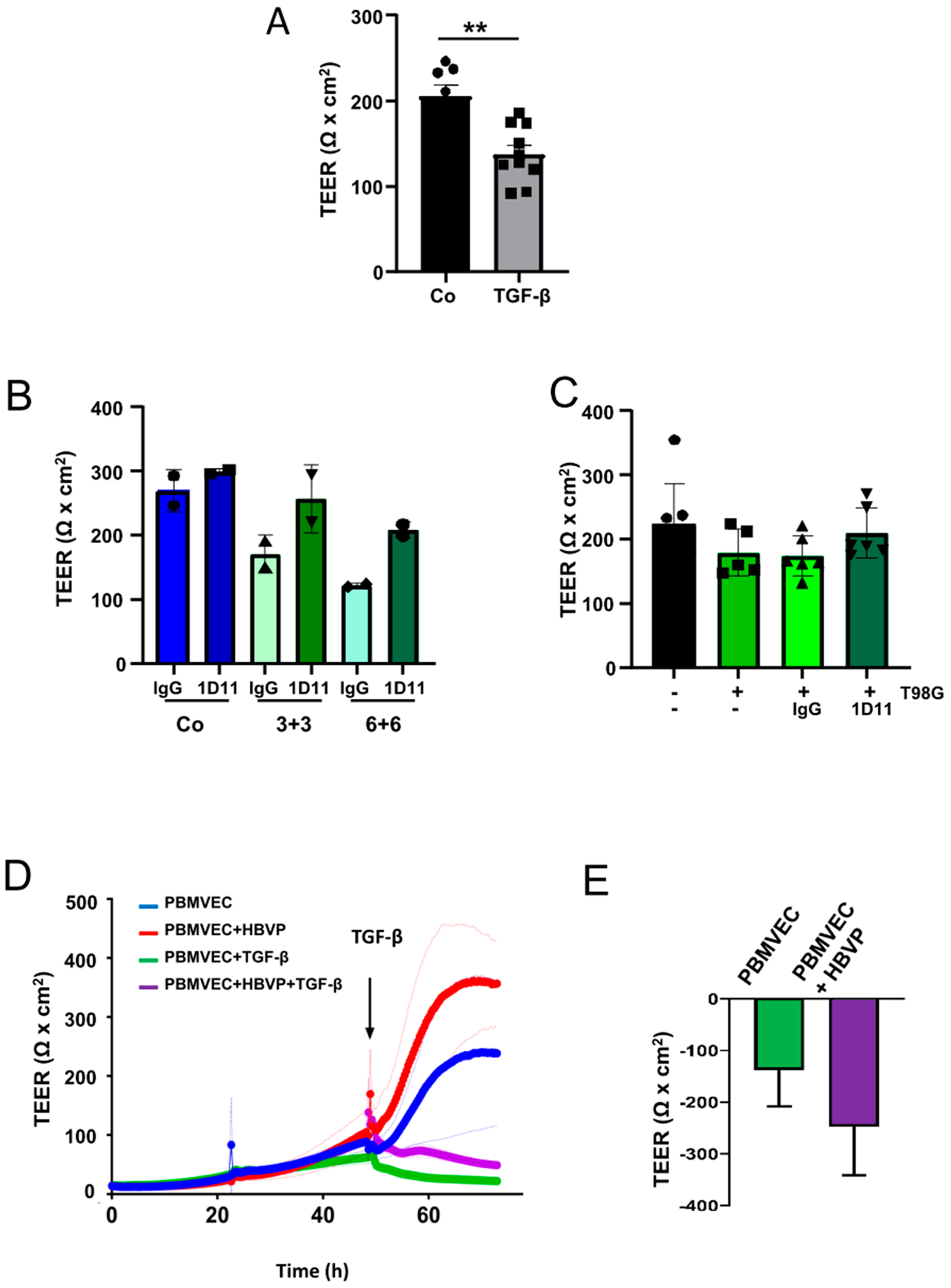
3.1. Co-Cultures of Primary Endothelial Cells and Pericytes Develop a Tight BBB
3.2. TGF-β Treatment of HBVP Negativels Modelates the Integrity of the BBB
3.3. TGF-β Treatment Leads to a Distinct Metabolite Profile, Pathway Alterations, and Gene Expression Patterns in HBVP
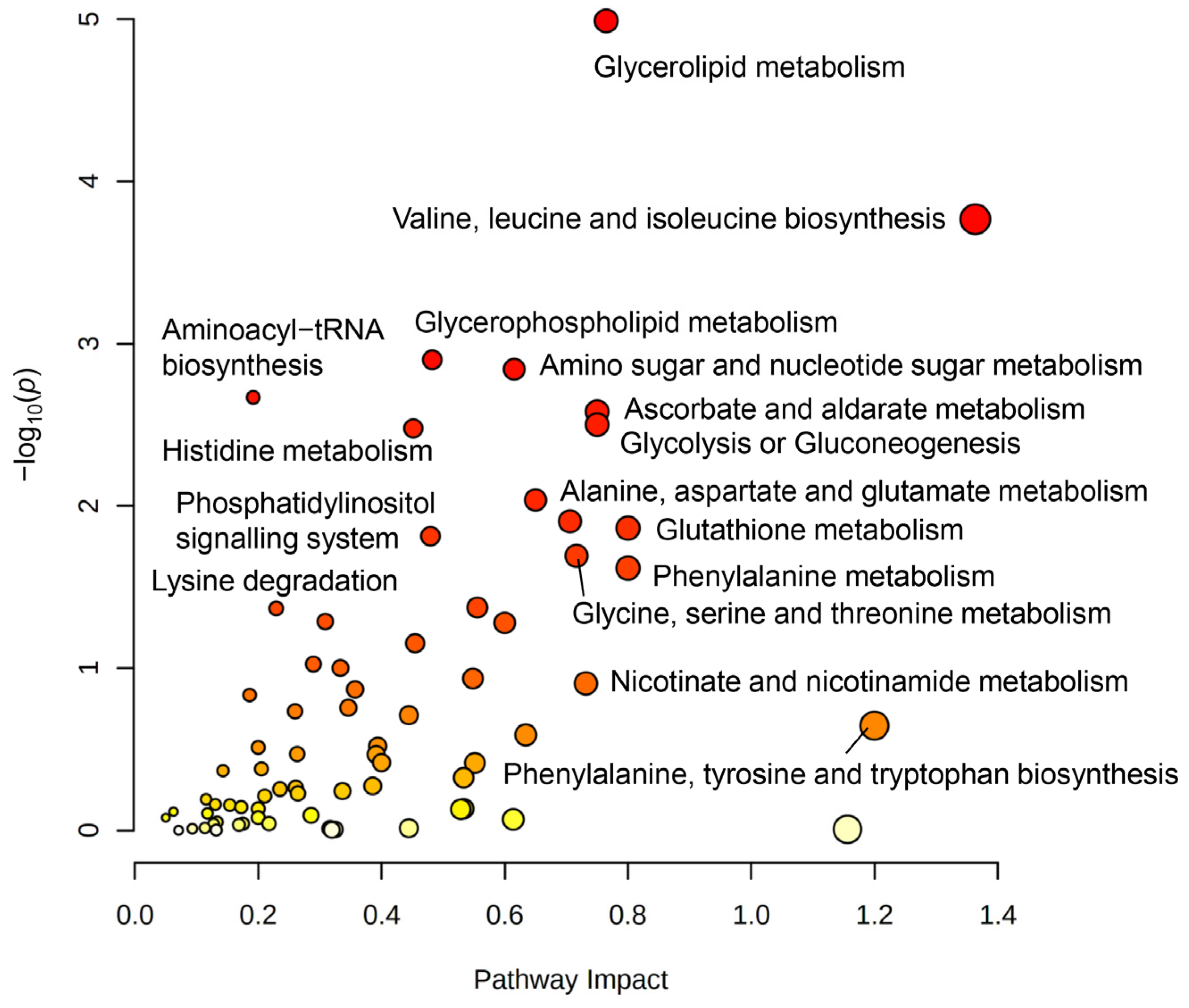
3.4. Joint-Pathway Analysis Illustrates Further Pathways Dysregulated upon TGF-β Treatment
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ADAMTS6 | ADAM metallopeptidase with thrombospondin type 1 motif, 6 |
| AJ | adherent junction |
| ANOVA | analysis of variance |
| BBB | blood–brain barrier |
| BCAA | branched amino acid |
| BMVEC | brain microvascular endothelial cells |
| BSA | bovine serum albumin |
| CM | conditioned medium |
| CNS | central nerve system |
| DMEM | Dulbecco’s Modified Eagle’s Medium |
| ECGS | endothelial cell growth supplements |
| EGF | epidermal growth factor |
| EM | epithelial cell growth medium |
| EMT | epithelial-to-mesenchymal transition |
| ESM1 | endothelial cell-specific molecule 1/endocan |
| FC | fold change |
| FCS | fetal calf serum |
| FGF | fibroblast growth factor |
| FDR | false discovery rate |
| FOXS1 | forkhead box S1 |
| GBM | glioblastoma |
| GJ | gap junction |
| GPR183 | G protein-coupled receptor 183 |
| HBVP | human brain microvascular pericytes |
| HRP | horseradish peroxidase |
| IGF-1 | insulin-like growth factor 1 |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| mTORC1 | mammalian target of rapamycin complex 1 |
| NOX4 | NADPH oxidase 4 |
| NVU | neurovascular unit |
| o/n | overnight |
| OxPhos | oxidative phosphorylation |
| PBMVEC | porcine brain microvascular endothelial cells |
| PCA | principal component analysis |
| PGS | pericyte growth supplement |
| PLL | poly-L-lysine |
| PM | pericyte medium |
| PMET | plasma membrane electron transport |
| PPARGC1AC | peroxisome proliferator-activated receptor gamma, coactivator 1 alpha |
| PRR5 | proline rich 5 |
| P/S | penicillin/streptomycin |
| RT | room temperature |
| SLC46A3 | solute carrier family 46, member 3 |
| SD | standard deviation |
| SEM | standard error of the mean |
| SPOCK1 | sparc/osteonectin, cwcv and kazal-like domains proteoglycan 1 |
| SV-GA | SV40 large T-antigene immortalized astrocytic cells |
| TEER | transendothelial electric resistance |
| TGF-β | transforming growth factor beta |
| TJ | tight junction |
| VEGF | vascular endothelial growth factor |
| ZO | zonula occludens |
References
- Langen, U.H.; Ayloo, S.; Gu, C. Development and Cell Biology of the Blood-Brain Barrier. Annu. Rev. Cell Dev. Biol. 2019, 35, 591–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.X.; Lin, G.S.; Lin, Z.X.; Zhang, J.D. Peritumoral edema shown by MRI predicts poor clinical outcome in glioblastoma. World J. Surg. Oncol. 2015, 13, 97. [Google Scholar] [CrossRef] [Green Version]
- Iorgulescu, J.B.; Gokhale, P.C.; Speranza, M.C.; Eschle, B.K.; Portras, M.J.; Wilken, M.K.; Soroko, K.M.; Chhoeu, C.; Knott, A.; Gao, Y.; et al. Concurrent Dexamethasone Limits the Clinical Benefit of Immune Checkpoint Blockade in Glioblastoma. Clin. Cancer Res. 2021, 27, 276–287. [Google Scholar] [CrossRef] [PubMed]
- Jackson, S.; ElALi, A.; Virgintino, D.; Gilbert, M.R. Blood-brain barrier pericyte importance in malignant gliomas: What we can learn from stroke and Alzheimer’s disease. Neuro Oncol. 2017, 19, 1173–1182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nwadozi, E.; Rudnicki, M.; Haas, T.L. Metabolic Coordination of Pericyte Phenotypes: Therapeutic Implications. Front. Cell Dev. Biol. 2020, 8, 77. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.F.; Ogunshola, O.O. Metabolomic profiling provides new insights into blood-brain barrier regulation. Neural Regen. Res. 2021, 16, 1786–1787. [Google Scholar]
- Mäder, L.; Blank, A.E.; Capper, D.; Jansong, J.; Baumgarten, P.; Wirsik, N.M.; Zachskorn, C.; Ehlers, J.; Seifert, M.; Klink, B.; et al. Pericytes/vessel-associated mural cells (VAMCs) are the major source of key epithelial-mesenchymal transition (EMT) factors SLUG and TWIST in human glioma. Oncotarget 2018, 9, 24041–24053. [Google Scholar] [CrossRef] [Green Version]
- Ehlers, J.; Mäder, L.; Ilina, E.; Blank, A.-E.; Grote, A.; Feuerhake, F.; Baumgarten, P.; Devraj, K.; Harter, P.; Mittelbronn, M.; et al. TGF-beta activates pericytes via induction of the epithelial to mesenchymal transition protein SLUG in glioblastoma. Neuropathol. Appl. Neurobiol. 2021, 47, 768–780. [Google Scholar]
- Ständer, M.; Naumann, U.; Dumitrescu, L.; Heneka, M.; Gulbins, E.; Dichgans, J.; Weller, M. Decorin gene transfer-mediated suppression of TGF-beta synthesis abrogates experimental malignant glioma growth in vivo. Gene Ther. 1998, 5, 1187–1194. [Google Scholar] [CrossRef] [Green Version]
- Major, E.O.; Miller, A.E.; Mourrain, P.; Traub, R.G.; de Widt, E.; Sever, J. Establishment of a line of human fetal glial cells that supports JC virus multiplication. Proc. Natl. Acad. Sci. USA 1985, 82, 1257–1261. [Google Scholar] [CrossRef] [Green Version]
- Mantwill, K.; Naumann, U.; Seznec, J.; Girbinger, V.; Lage, H.; Surowiak, P.; Beier, D.; Mittelbronn, M.; Schlegel, J.; Holm, P.S. YB-1 dependent oncolytic adenovirus efficiently inhibits tumor growth of glioma cancer stem like cells. J. Transl. Med. 2013, 11, 216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartussek, C.; Naumann, U.; Weller, M. Accumulation of mutant p53(V143A) modulates the growth, clonogenicity, and radiochemosensitivity of malignant glioma cells independent of endogenous p53 status. Exp. Cell Res. 1999, 253, 432–439. [Google Scholar] [CrossRef]
- Thomsen, L.B.; Burkhart, A.; Moos, T. A Triple Culture Model of the Blood-Brain Barrier Using Porcine Brain Endothelial cells, Astrocytes and Pericytes. PLoS ONE 2015, 10, e0134765. [Google Scholar] [CrossRef] [Green Version]
- Paolinelli, R.; Corada, M.; Ferrarini, L.; Devraj, K.; Artus, C.; Czupalla, C.J.; Rudini, N.; Maddaluna, L.; Papa, E.; Engelhardt, B.; et al. Wnt activation of immortalized brain endothelial cells as a tool for generating a standardized model of the blood brain barrier in vitro. PLoS ONE 2013, 8, e70233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czupalla, C.J.; Liebner, S.; Devraj, K. In vitro models of the blood-brain barrier. Methods Mol. Biol. 2014, 1135, 415–437. [Google Scholar] [PubMed]
- Srinivasan, B.; Kolli, A.R.; Esch, M.B.; Abaci, H.E.; Shuler, M.L.; Hickman, J.J. TEER measurement techniques for in vitro barrier model systems. J. Lab Autom. 2015, 20, 107–126. [Google Scholar] [CrossRef] [Green Version]
- Wolburg, H.; Liebner, S.; Lippoldt, A. Freeze-fracture studies of cerebral endothelial tight junctions. Methods Mol. Med. 2003, 89, 51–66. [Google Scholar]
- Schindelin, J.; Aganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Bus, C.; Zizmare, L.; Feldkämper, M.; Geisler, S.; Zarani, M.; Schädler, A.; Klose, F.; Admard, J.; Mageean, C.J.; Arena, G.; et al. Human Dopaminergic Neurons Lacking PINK1 Exhibit Disrupted Dopamine Metabolism Related to Vitamin B6 Co-Factors. iScience 2020, 23, 101797. [Google Scholar] [CrossRef] [PubMed]
- Pang, Z.; Zhou, G.; Ewald, J.; Hacariz, O.; Basu, N.; Xia, L. Using MetaboAnalyst 5.0 for LC-HRMS spectra processing, multi-omics integration and covariate adjustment of global metabolomics data. Nat. Protoc. 2022, 17, 1735–1761. [Google Scholar] [CrossRef] [PubMed]
- Malinovskaya, N.A.; Komleva, Y.K.; Samin, V.V.; Morgun, A.V.; Shuvaev, A.N.; Panina, Y.A.; Boitsova, E.B.; Samina, A.B. Endothelial Progenitor Cells Physiology and Metabolic Plasticity in Brain Angiogenesis and Blood-Brain Barrier Modeling. Front. Physiol. 2016, 7, 599. [Google Scholar] [CrossRef] [Green Version]
- Cantelmo, A.R.; Conradi, L.C.; Brajic, A.; Goveia, J.; Kalucke, J.; Pircher, A.; Chaturredi, P.; Hol, J.; Thienpont, B.; Teuwen, L.A.; et al. Inhibition of the Glycolytic Activator PFKFB3 in Endothelium Induces Tumor Vessel Normalization, Impairs Metastasis, and Improves Chemotherapy. Cancer Cell 2016, 30, 968–985. [Google Scholar] [CrossRef] [Green Version]
- Ochs, K.; Sahm, F.; Opitz, C.A.; Lanz, T.V.; Oezen, I.; Couraud, P.O.; von Deimling, A.; Wick, W.; Platten, M. Immature mesenchymal stem cell-like pericytes as mediators of immunosuppression in human malignant glioma. J. Neuroimmunol. 2013, 265, 106–116. [Google Scholar] [CrossRef] [PubMed]
- Svensson, A.; Ozen, I.; Genove, G.; Paul, G.; Bengzon, J. Endogenous brain pericytes are widely activated and contribute to mouse glioma microvasculature. PLoS ONE 2015, 10, e0123553. [Google Scholar] [CrossRef] [Green Version]
- Thanabalasundaram, G.; Pieper, C.; Lischper, M.; Galla, H.J. Regulation of the blood-brain barrier integrity by pericytes via matrix metalloproteinases mediated activation of vascular endothelial growth factor in vitro. Brain Res. 2010, 1347, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Patabendige, A.; Abbott, N.J. Primary porcine brain microvessel endothelial cell isolation and culture. Curr. Protoc. Neurosci. 2014, 69, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Gericke, B.; Romermann, K.; Noack, A.; Noacke, S.; Kronenberg, J.; Blasig, I.E.; Loscher, W. A face-to-face comparison of claudin-5 transduced human brain endothelial (hCMEC/D3) cells with porcine brain endothelial cells as blood-brain barrier models for drug transport studies. Fluids Barriers CNS 2020, 17, 53. [Google Scholar] [CrossRef]
- Thomsen, M.S.; Humle, N.; Hede, E.; Moos, T.; Burkhart, A.; Thomsen, L.B. The blood-brain barrier studied in vitro across species. PLoS ONE 2021, 16, e0236770. [Google Scholar] [CrossRef]
- Papetti, M.; Shujath, J.; Riley, K.N.; Herman, I.M. FGF-2 antagonizes the TGF-beta1-mediated induction of pericyte alpha-smooth muscle actin expression: A role for myf-5 and Smad-mediated signaling pathways. Investig. Ophthalmol. Vis. Sci. 2003, 44, 4994–5005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thanabalasundaram, G.; Schneidewind, J.; Pieper, C.; Gall, H.J. The impact of pericytes on the blood-brain barrier integrity depends critically on the pericyte differentiation stage. Int. J. Biochem. Cell Biol. 2011, 43, 1284–1293. [Google Scholar] [CrossRef]
- Atis, M.; Akcan, U.; Altunsu, D.; Ayvaz, E.; Ugur Yilmaz, C.; Sarikaya, D.; Temizyurek, A.; Ahishali, B.; Girouard, H.; Kaya, M. Targeting the blood-brain barrier disruption in hypertension by ALK5/TGF-Beta type I receptor inhibitor SB-431542 and dynamin inhibitor dynasore. Brain Res. 2022, 1794, 148071. [Google Scholar] [CrossRef]
- Gong, L.; Ji, L.; Xu, D.; Wang, J.; Zou, J. TGF-beta links glycolysis and immunosuppression in glioblastoma. Histol. Histopathol. 2021, 36, 1111–1124. [Google Scholar]
- Hattingen, E.; Bähr, O.; Rieger, J.; Blasel, S.; Steinbach, J.; Pilatus, H. Phospholipid metabolites in recurrent glioblastoma: In vivo markers detect different tumor phenotypes before and under antiangiogenic therapy. PLoS ONE 2013, 8, e56439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herst, P.M.; Berridge, M.V. Plasma membrane electron transport: A new target for cancer drug development. Curr. Mol. Med. 2006, 6, 895–904. [Google Scholar] [CrossRef]
- Gan, X.; Wang, J.; Wang, C.; Sommer, E.; Kozasa, T.; Srinivasula, S.; Alessi, D.; Offermanns, S.; Simon, M.I.; Wu, D. PRR5L degradation promotes mTORC2-mediated PKC-delta phosphorylation and cell migration downstream of Galpha12. Nat. Cell Biol. 2012, 14, 686–696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishimura, A.; Ago, T.; Kurada, J.; Avimura, K.; Tachibana, M.; Nakamura, K.; Wakisaka, Y.; Sadoshima, J.; Iihara, K.; Kitazono, T. Detrimental role of pericyte Nox4 in the acute phase of brain ischemia. J. Cereb. Blood Flow Metab. 2016, 36, 1143–1154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Underly, R.G.; Levy, M.; Hartmann, D.A.; Grant, R.I.; Watson, A.N.; Shih, A.Y. Pericytes as Inducers of Rapid, Matrix Metalloproteinase-9-Dependent Capillary Damage during Ischemia. J. Neurosci. 2017, 37, 129–140. [Google Scholar] [CrossRef] [Green Version]
- Sanchis-Gomar, F.; Garcia-Gimenez, J.L.; Gomez-Cabrera, M.C.; Pallardo, F.V. Mitochondrial biogenesis in health and disease. Molecular and therapeutic approaches. Curr. Pharm. Des. 2014, 20, 5619–5633. [Google Scholar]
- Liang, H.; Ward, W.F. PGC-1alpha: A key regulator of energy metabolism. Adv. Physiol. Educ. 2006, 30, 145–151. [Google Scholar] [CrossRef]
- Zhenyukh, O.; Civantos, E.; Riuz-Ortega, M.; Sanchez, M.S.; Vasquez, C.; Peiro, C.; Egido, J.; Mas, S. High concentration of branched-chain amino acids promotes oxidative stress, inflammation and migration of human peripheral blood mononuclear cells via mTORC1 activation. Free Radic. Biol. Med. 2017, 104, 165–177. [Google Scholar] [CrossRef]
- Ding, Y.; Shan, L.; Nai, W.; Lin, X.; Zhou, L.; Dong, X.; Wu, H.; Xiao, M.; Zhou, X.; Wang, L.; et al. DEPTOR Deficiency-Mediated mTORc1 Hyperactivation in Vascular Endothelial Cells Promotes Angiogenesis. Cell Physiol. Biochem. 2018, 46, 520–531. [Google Scholar] [CrossRef]
- Banks, W.A.; Reed, M.J.; Logsdon, A.F.; Rhea, E.M.; Erickson, M.A. Healthy aging and the blood-brain barrier. Nat. Aging 2021, 1, 243–254. [Google Scholar] [CrossRef]
- Barnes, S.; Chowdhury, S.; Gatto, N.M.; Fraser, G.E.; Lee, G.J. Omega-3 fatty acids are associated with blood-brain barrier integrity in a healthy aging population. Brain Behav. 2021, 11, e2273. [Google Scholar] [CrossRef]
- May, J.M.; Qu, Z.C. Ascorbic acid efflux from human brain microvascular pericytes: Role of re-uptake. Biofactors 2015, 41, 330–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMillin, M.A.; Frampton, G.A.; Seiwell, A.P.; Patel, N.S.; Jacobs, N.S.; DeMorrow, S. TGFbeta1 exacerbates blood-brain barrier permeability in a mouse model of hepatic encephalopathy via upregulation of MMP9 and downregulation of claudin-5. Lab Investig. 2015, 95, 903–913. [Google Scholar] [CrossRef]
- Krizbai, I.A.; Gasparics, A.; Nagyoszi, P.; Fazakas, C.; Molnar, J.; Wilhelm, I.; Bencs, R.; Rosivall, L.; Sebe, A. Endothelial-mesenchymal transition of brain endothelial cells: Possible role during metastatic extravasation. PLoS ONE 2015, 10, e0123845. [Google Scholar] [CrossRef] [Green Version]
- Shen, W.; Li, S.; Chung, S.H.; Zhu, L.; Stayt, J.; Su, T.; Couraud, P.O.; Romero, I.A.; Weksler, B.; Gillies, M.C. Tyrosine phosphorylation of VE-cadherin and claudin-5 is associated with TGF-beta1-induced permeability of centrally derived vascular endothelium. Eur. J. Cell Biol. 2011, 90, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Dohgu, S.; Takata, F.; Yamauchi, A.; Nakagawa, S.; Egawa, T.; Naito, M.; Tsuruo, T.; Sawada, Y.; Niwa, M.; Kataoka, Y. Brain pericytes contribute to the induction and up-regulation of blood-brain barrier functions through transforming growth factor-beta production. Brain Res. 2005, 1038, 208–215. [Google Scholar] [CrossRef]
- Jones, B. Targeted therapies: Early vessel normalization improves glioblastoma outcomes. Nat. Rev. Clin. Oncol. 2014, 11, 4. [Google Scholar] [CrossRef] [PubMed]
- Stylianopoulos, T.; Jain, R.K. Combining two strategies to improve perfusion and drug delivery in solid tumors. Proc. Natl. Acad. Sci. USA 2013, 110, 18632–18637. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schumacher, L.; Slimani, R.; Zizmare, L.; Ehlers, J.; Kleine Borgmann, F.; Fitzgerald, J.C.; Fallier-Becker, P.; Beckmann, A.; Grißmer, A.; Meier, C.; et al. TGF-Beta Modulates the Integrity of the Blood Brain Barrier In Vitro, and Is Associated with Metabolic Alterations in Pericytes. Biomedicines 2023, 11, 214. https://doi.org/10.3390/biomedicines11010214
Schumacher L, Slimani R, Zizmare L, Ehlers J, Kleine Borgmann F, Fitzgerald JC, Fallier-Becker P, Beckmann A, Grißmer A, Meier C, et al. TGF-Beta Modulates the Integrity of the Blood Brain Barrier In Vitro, and Is Associated with Metabolic Alterations in Pericytes. Biomedicines. 2023; 11(1):214. https://doi.org/10.3390/biomedicines11010214
Chicago/Turabian StyleSchumacher, Leonie, Rédouane Slimani, Laimdota Zizmare, Jakob Ehlers, Felix Kleine Borgmann, Julia C. Fitzgerald, Petra Fallier-Becker, Anja Beckmann, Alexander Grißmer, Carola Meier, and et al. 2023. "TGF-Beta Modulates the Integrity of the Blood Brain Barrier In Vitro, and Is Associated with Metabolic Alterations in Pericytes" Biomedicines 11, no. 1: 214. https://doi.org/10.3390/biomedicines11010214
APA StyleSchumacher, L., Slimani, R., Zizmare, L., Ehlers, J., Kleine Borgmann, F., Fitzgerald, J. C., Fallier-Becker, P., Beckmann, A., Grißmer, A., Meier, C., El-Ayoubi, A., Devraj, K., Mittelbronn, M., Trautwein, C., & Naumann, U. (2023). TGF-Beta Modulates the Integrity of the Blood Brain Barrier In Vitro, and Is Associated with Metabolic Alterations in Pericytes. Biomedicines, 11(1), 214. https://doi.org/10.3390/biomedicines11010214