

Noradrenergic Modulation of Learned and Innate Behaviors in Dopamine Transporter Knockout Rats by Guanfacine

 ,
,  and
and 
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Animals and Their Maintenance
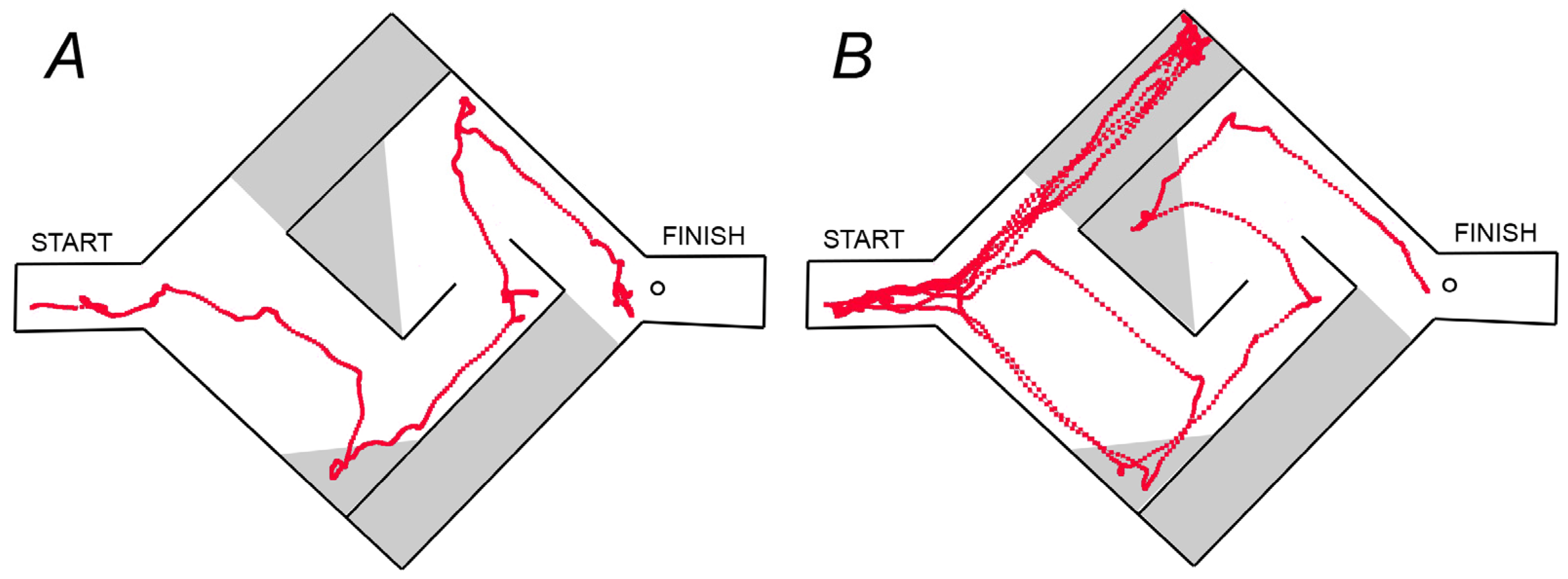
2.2. Hebb–Williams Maze
2.2.1. Apparatus
2.2.2. Experimental Setup
2.3. Acoustic Startle Reaction (ASR) and the Pre-Pulse Inhibition (PPI)
2.3.1. Apparatus
2.3.2. Experimental Setup
2.4. LFP Power Spectra and Coherence
2.4.1. Surgical Procedures
2.4.2. EEG and LFP Recordings
2.4.3. Analysis of EEG and LFP Activity
2.5. Statistical Analysis
3. Results
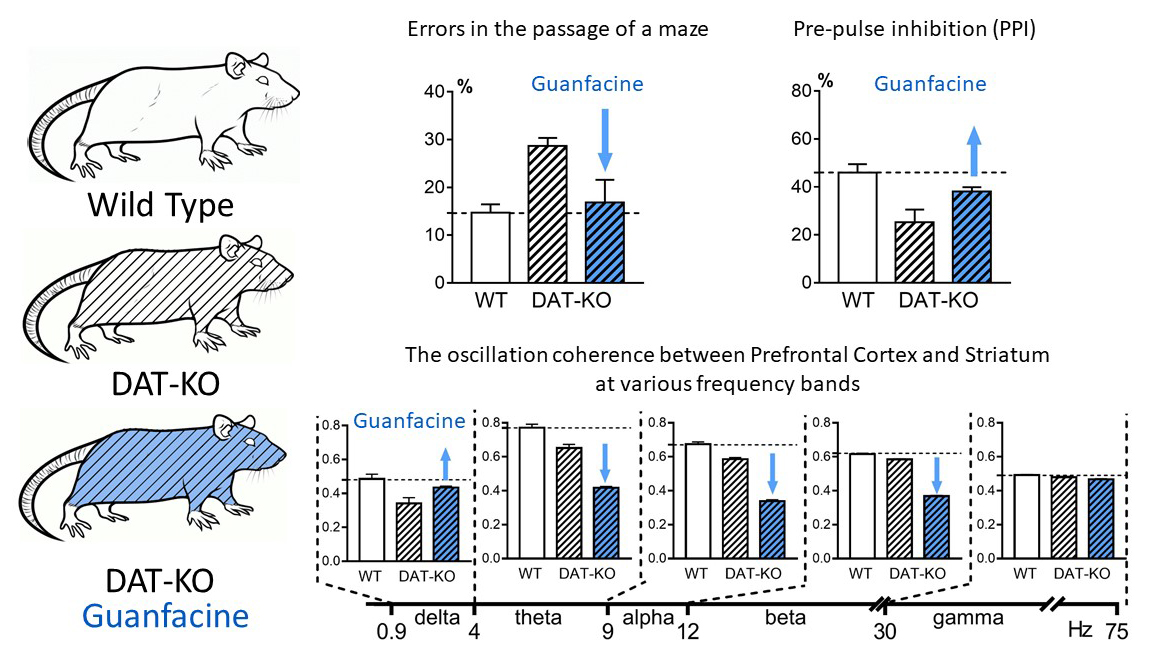
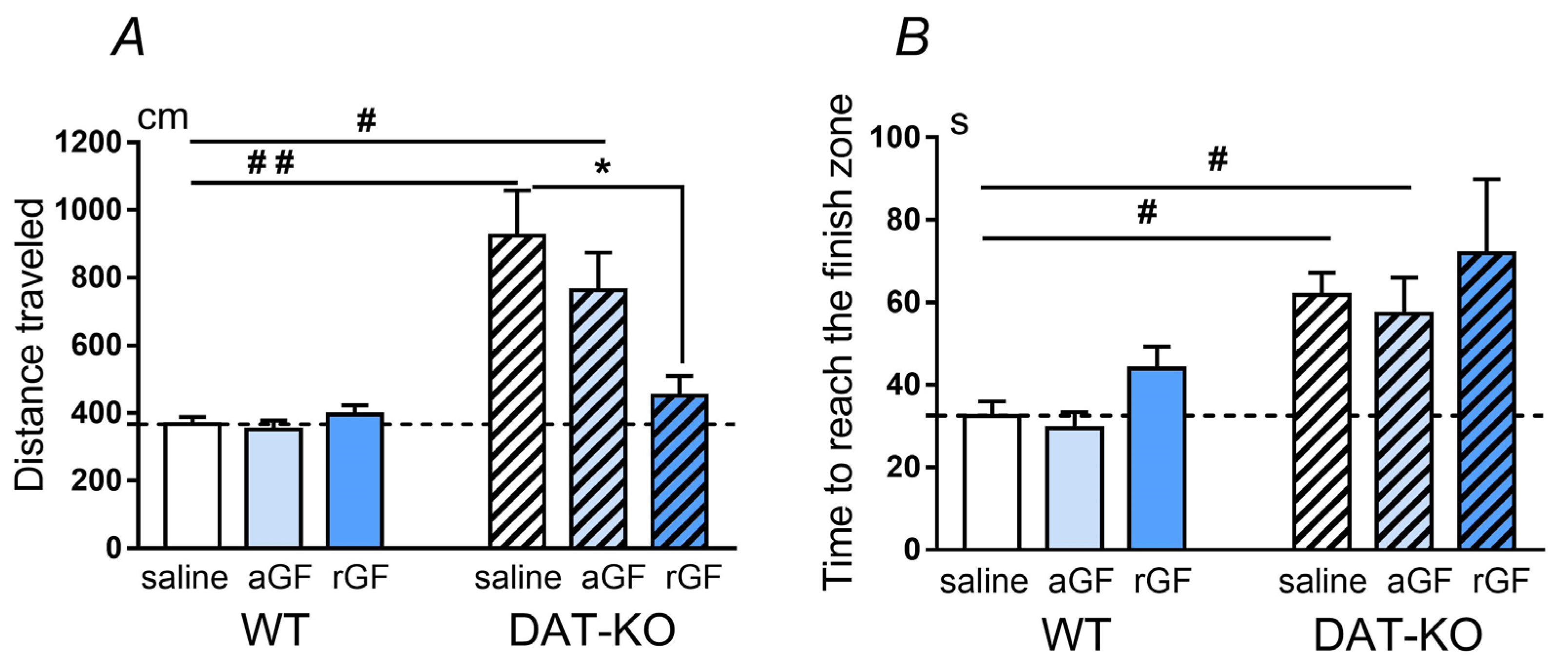
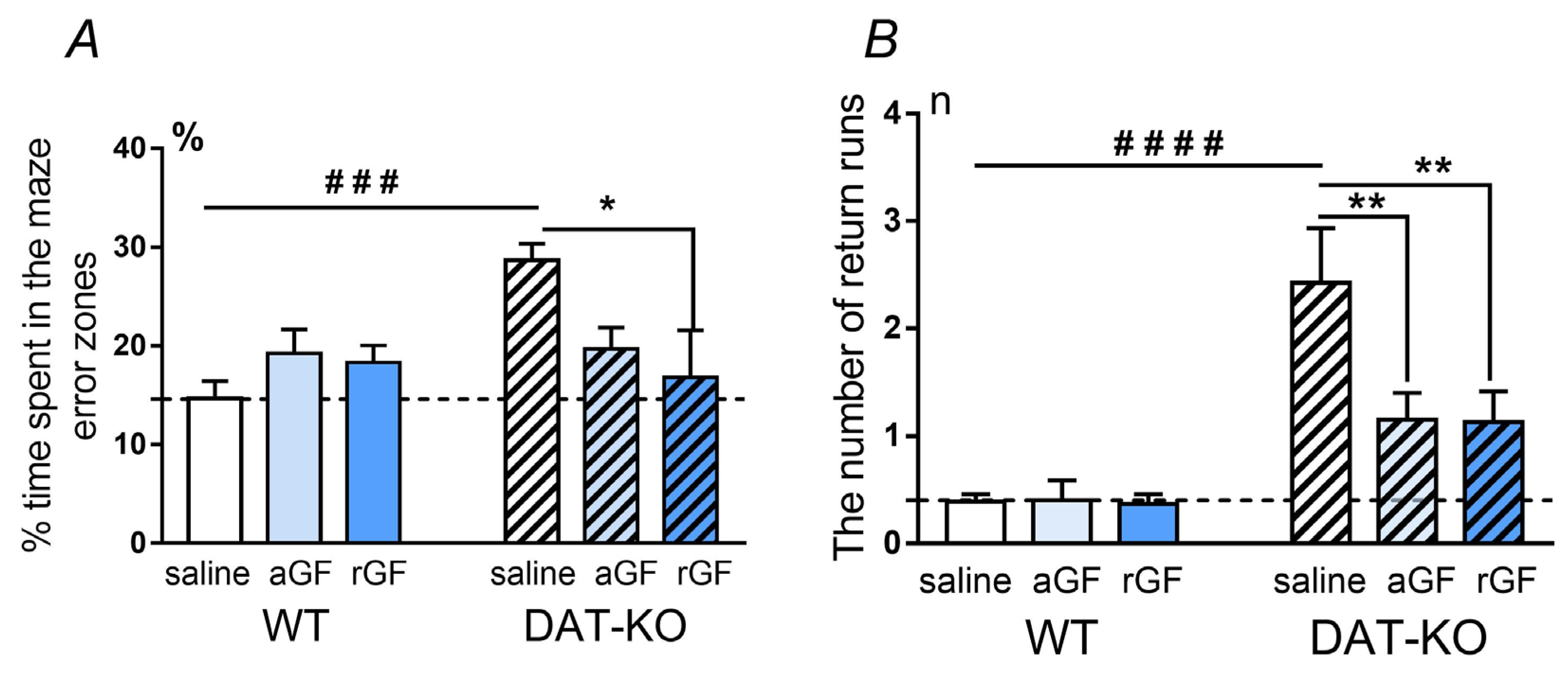
3.1. Hebb–Williams Maze
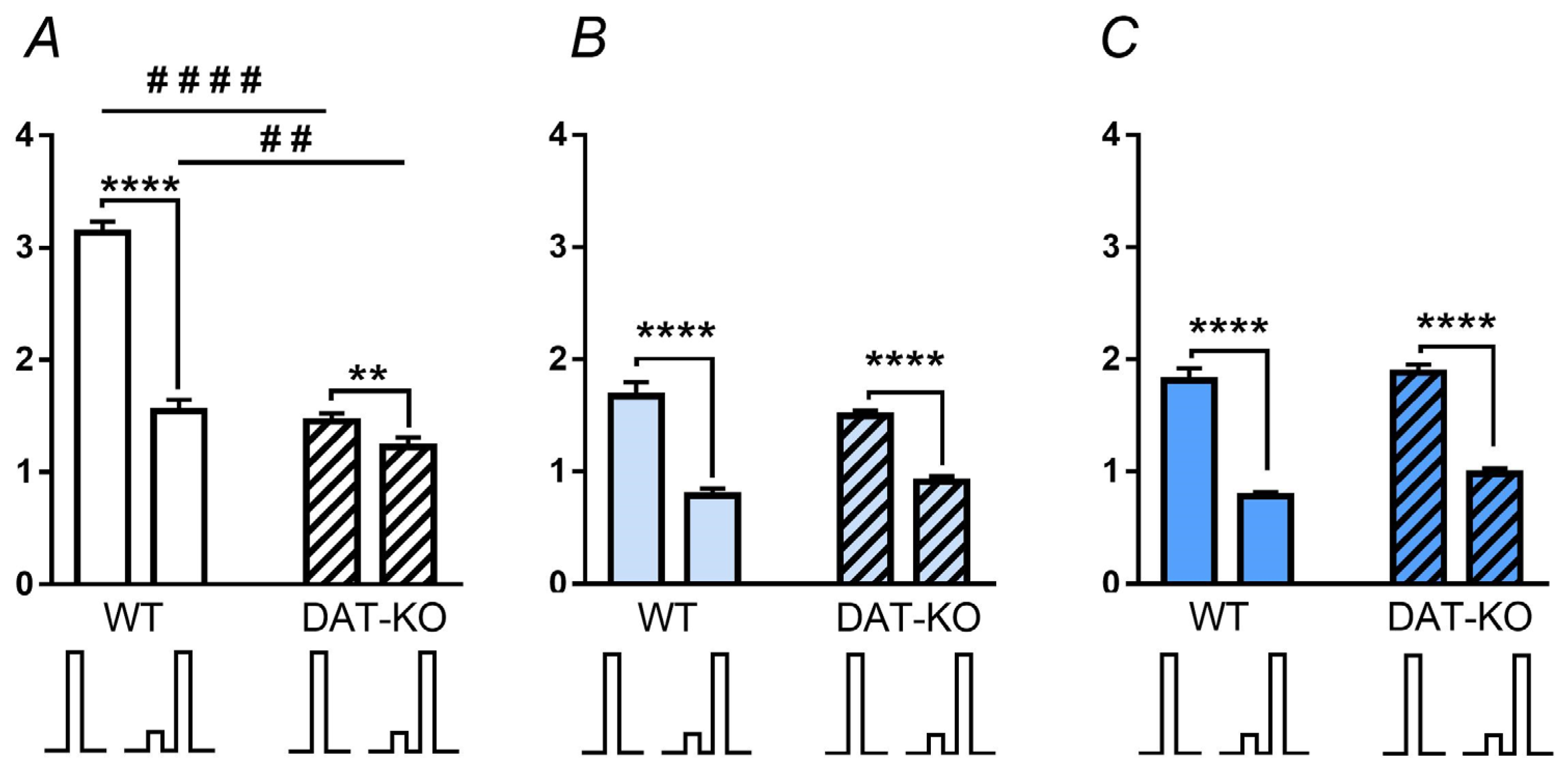
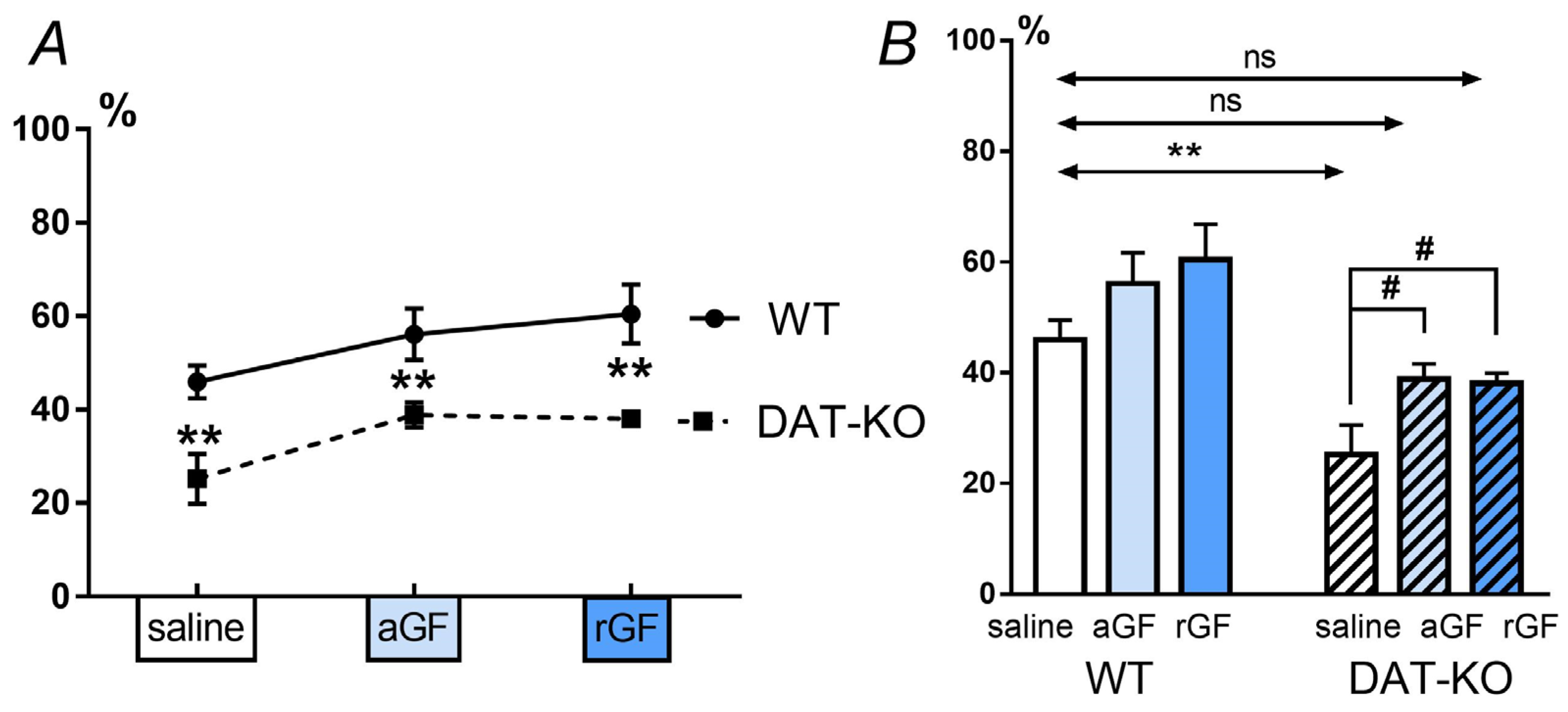
3.2. Acoustic Startle Reaction (ASR) and the Pre-Pulse Inhibition (PPI)
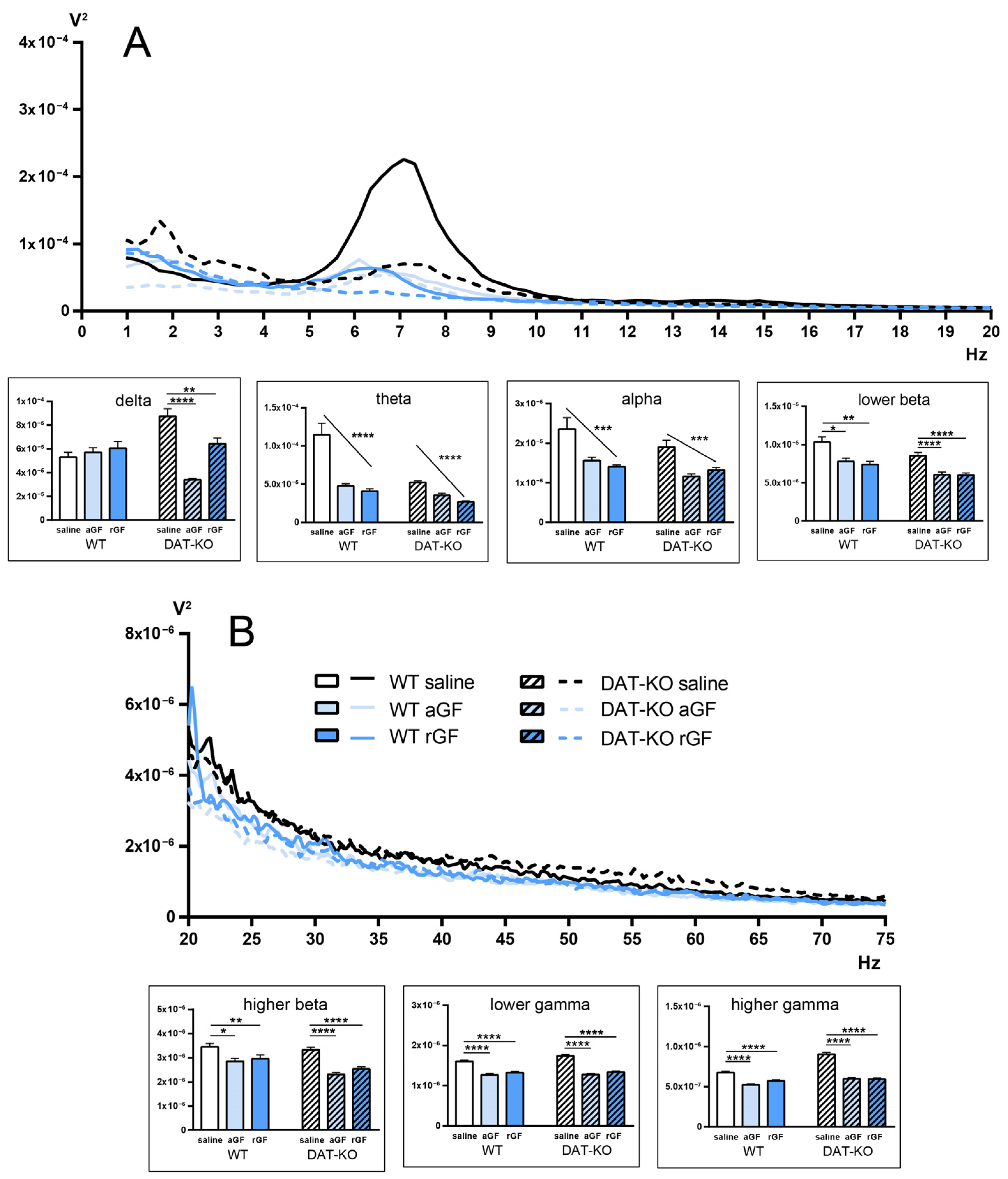
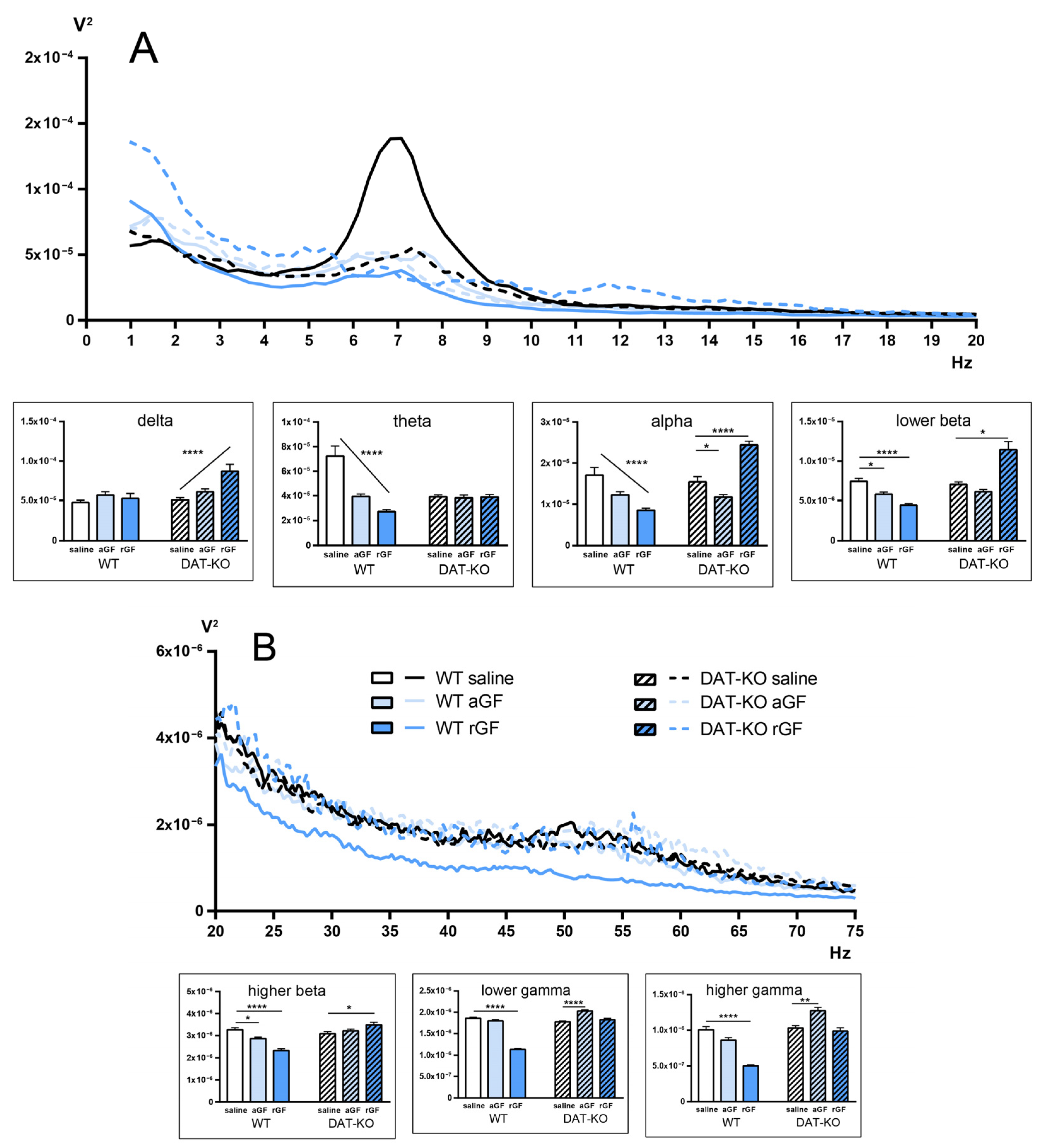
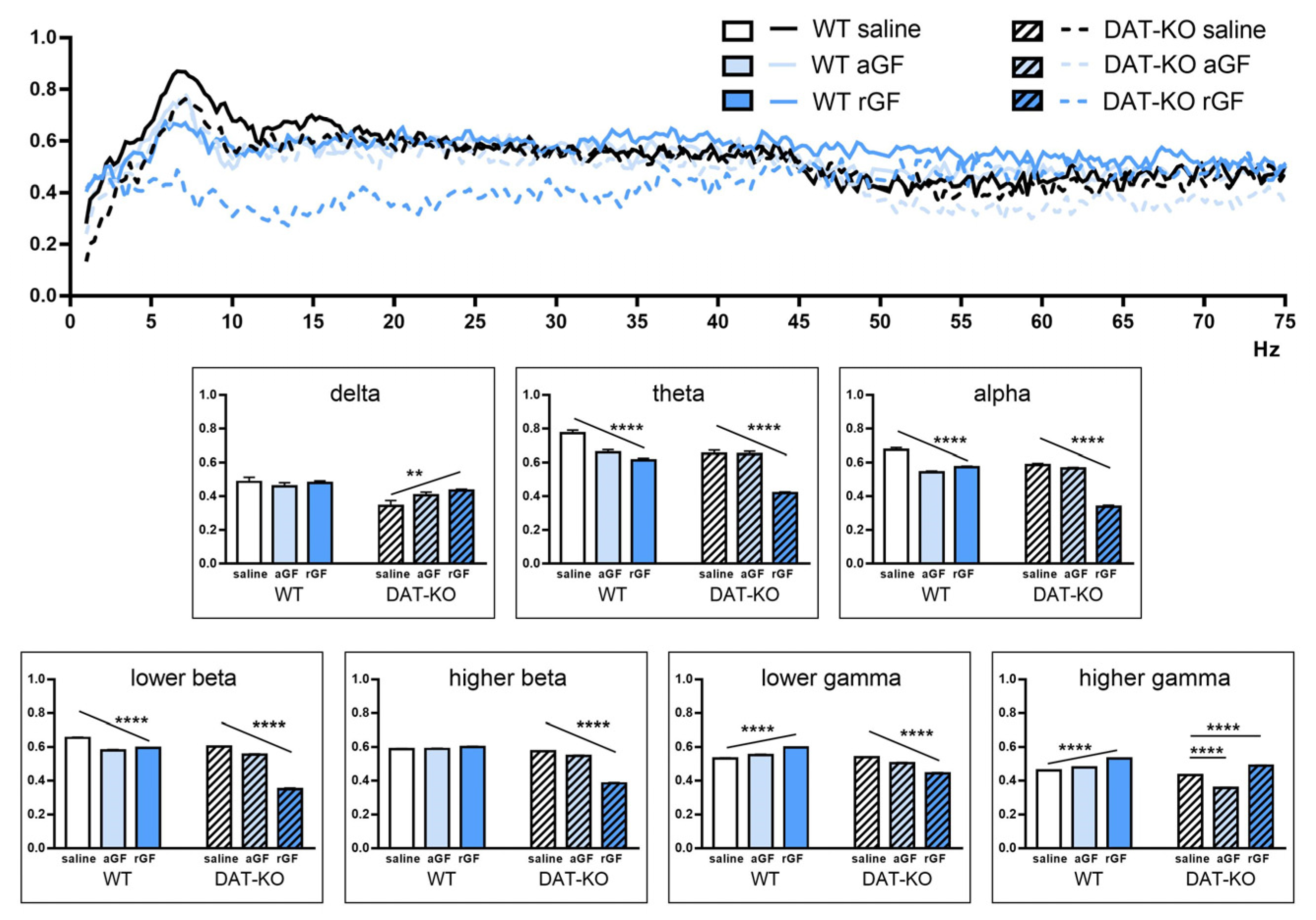
3.3. Power Spectra and Coherence of the Brain Activity
4. Discussion
5. Conclusions
6. Limitations and Future Directions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nikolaus, S.; Antke, C.; Beu, M.; Müller, H.W.; Nikolaus, S. Cortical GABA, Striatal Dopamine and Midbrain Serotonin as the Key Players in Compulsiveand Anxiety Disorders—Results from in Vivo Imaging Studies. Rev. Neurosci. 2010, 21, 119–140. [Google Scholar] [CrossRef] [PubMed]
- Grace, A.A. Dysregulation of the Dopamine System in the Pathophysiology of Schizophrenia and Depression. Nat. Rev. Neurosci. 2016, 17, 524–532. [Google Scholar] [CrossRef] [PubMed]
- Klein, M.O.; Battagello, D.S.; Cardoso, A.R.; Hauser, D.N.; Bittencourt, J.C.; Correa, R.G. Dopamine: Functions, Signaling, and Association with Neurological Diseases. Cell. Mol. Neurobiol. 2018, 39, 31–59. [Google Scholar] [CrossRef]
- Beaulieu, J.-M.; Gainetdinov, R.R. The Physiology, Signaling, and Pharmacology of Dopamine Receptors. Pharmacol. Rev. 2011, 63, 182–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galéra, C.; Cortese, S.; Orri, M.; Collet, O.; van der Waerden, J.; Melchior, M.; Boivin, M.; Tremblay, R.E.; Côté, S.M. Medical Conditions and Attention-Deficit/Hyperactivity Disorder Symptoms from Early Childhood to Adolescence. Mol. Psychiatry 2021, 27, 976–984. [Google Scholar] [CrossRef]
- Bonvicini, C.; Faraone, S.V.; Scassellati, C. Attention-Deficit Hyperactivity Disorder in Adults: A Systematic Review and Meta-Analysis of Genetic, Pharmacogenetic and Biochemical Studies. Mol. Psychiatry 2016, 21, 872–884. [Google Scholar] [CrossRef]
- Athanasiadou, A.; Buitelaar, J.K.; Brovedani, P.; Chorna, O.; Fulceri, F.; Guzzetta, A.; Scattoni, M.L. Early Motor Signs of Attention-Deficit Hyperactivity Disorder: A Systematic Review. Eur. Child Adolesc. Psychiatry 2020, 29, 903–916. [Google Scholar] [CrossRef] [Green Version]
- Posner, J.; Polanczyk, G.V.; Sonuga-Barke, E. Attention-Deficit Hyperactivity Disorder. Lancet 2020, 395, 450–462. [Google Scholar] [CrossRef]
- Cinque, S.; Zoratto, F.; Poleggi, A.; Leo, D.; Cerniglia, L.; Cimino, S.; Tambelli, R.; Alleva, E.; Gainetdinov, R.R.; Laviola, G.; et al. Behavioral Phenotyping of Dopamine Transporter Knockout Rats: Compulsive Traits, Motor Stereotypies, and Anhedonia. Front. Psychiatry 2018, 9, 43. [Google Scholar] [CrossRef] [PubMed]
- Kocaturk, R.; Ozcan, O.; Karahan, M. Animal Models of Attention Deficit and Hyperactivity Disorder: A Critical Overview and Suggestions. J. Neurobehav. Sci. 2021, 8, 1. [Google Scholar] [CrossRef]
- Leo, D.; Sukhanov, I.; Zoratto, F.; Illiano, P.; Caffino, L.; Sanna, F.; Messa, G.; Emanuele, M.; Esposito, A.; Dorofeikova, M.; et al. Pronounced Hyperactivity, Cognitive Dysfunctions, and BDNF Dysregulation in Dopamine Transporter Knock-out Rats. J. Neurosci. 2018, 38, 1959–1972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Natsheh, J.Y.; Shiflett, M.W. Dopaminergic Modulation of Goal-Directed Behavior in a Rodent Model of Attention-Deficit/Hyperactivity Disorder. Front. Integr. Neurosci. 2018, 12, 45. [Google Scholar] [CrossRef] [Green Version]
- Russell, V.A.; Sagvolden, T.; Johansen, E.B. Animal Models of Attention-Deficit Hyperactivity Disorder. Behav. Brain Funct. 2005, 1, 9. [Google Scholar] [CrossRef] [Green Version]
- Sagvolden, T.; Russell, V.A.; Aase, H.; Johansen, E.B.; Farshbaf, M. Rodent Models of Attention-Deficit/Hyperactivity Disorder. Biol. Psychiatry 2005, 57, 1239–1247. [Google Scholar] [CrossRef]
- Tanaka, M.; Spekker, E.; Szabó, Á.; Polyák, H.; Vécsei, L. Modelling the Neurodevelopmental Pathogenesis in Neuropsychiatric Disorders. Bioactive Kynurenines and Their Analogues as Neuroprotective Agents—In Celebration of 80th Birthday of Professor Peter Riederer. J. Neural Transm. 2022, 129, 627–642. [Google Scholar] [CrossRef] [PubMed]
- Bardgett, M.E.; Depenbrock, M.; Downs, N.; Points, M.; Green, L. Dopamine Modulates Effort-Based Decision Making in Rats. Behav. Neurosci. 2009, 123, 242–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cagniard, B.; Balsam, P.D.; Brunner, D.; Zhuang, X. Mice with Chronically Elevated Dopamine Exhibit Enhanced Motivation, but Not Learning, for a Food Reward. Neuropsychopharmacology 2006, 31, 1362–1370. [Google Scholar] [CrossRef] [Green Version]
- Chudasama, Y.; Robbins, T.W. Dopaminergic Modulation of Visual Attention and Working Memory in the Rodent Prefrontal Cortex. Neuropsychopharmacology 2004, 29, 1628–1636. [Google Scholar] [CrossRef] [Green Version]
- Tripp, G.; Wickens, J.R. Neurobiology of ADHD. Neuropharmacology 2009, 57, 579–589. [Google Scholar] [CrossRef] [PubMed]
- Volkow, N.D.; Wang, G.J.; Newcorn, J.H.; Kollins, S.H.; Wigal, T.L.; Telang, F.; Fowler, J.S.; Goldstein, R.Z.; Klein, N.; Logan, J.; et al. Motivation Deficit in ADHD Is Associated with Dysfunction of the Dopamine Reward Pathway. Mol. Psychiatry 2011, 16, 1147–1154. [Google Scholar] [CrossRef]
- Wise, R.A. Dopamine, Learning and Motivation. Nat. Rev. Neurosci. 2004, 5, 483–494. [Google Scholar] [CrossRef] [PubMed]
- Kurzina, N.P.; Aristova, I.Y.; Volnova, A.B.; Gainetdinov, R.R. Deficit in Working Memory and Abnormal Behavioral Tactics in Dopamine Transporter Knockout Rats during Training in the 8-Arm Maze. Behav. Brain Res. 2020, 390, 112642. [Google Scholar] [CrossRef] [PubMed]
- Kurzina, N.P.; Volnova, A.B.; Aristova, I.Y.; Gainetdinov, R.R. A New Paradigm for Training Hyperactive Dopamine Transporter Knockout Rats: Influence of Novel Stimuli on Object Recognition. Front. Behav. Neurosci. 2021, 15, 654469. [Google Scholar] [CrossRef] [PubMed]
- Savchenko, A.; Müller, C.; Lubec, J.; Leo, D.; Korz, V.; Afjehi-Sadat, L.; Malikovic, J.; Sialana, F.J.; Lubec, G.; Sukhanov, I. The Lack of Dopamine Transporter Is Associated With Conditional Associative Learning Impairments and Striatal Proteomic Changes. Front. Psychiatry 2022, 13, 269. [Google Scholar] [CrossRef] [PubMed]
- Jahn, C.I.; Varazzani, C.; Sallet, J.; Walton, M.E.; Bouret, S. Noradrenergic But Not Dopaminergic Neurons Signal Task State Changes and Predict Reengagement After a Failure. Cereb. Cortex 2020, 30, 4979–4994. [Google Scholar] [CrossRef]
- Robbins, T.W.; Arnsten, A.F.T. The Neuropsychopharmacology of Fronto-Executive Function: Monoaminergic Modulation. Annu. Rev. Neurosci. 2009, 32, 267–287. [Google Scholar] [CrossRef] [Green Version]
- Levy, F. Pharmacological and Therapeutic Directions in ADHD: Specificity in the PFC. Behav. Brain Funct. 2008, 4, 12. [Google Scholar] [CrossRef] [Green Version]
- Gao, W.-J.; Wang, H.-X.; Snyder, M.A.; Li, Y.-C. The Unique Properties of the Prefrontal Cortex and Mental Illness. In When Things Go Wrong: Diseases and Disorders of the Human Brain; InTech: London, UK, 2012; pp. 1–26. [Google Scholar] [CrossRef] [Green Version]
- Bokor, G.; Anderson, P.D. Attention-Deficit/Hyperactivity Disorder. J. Pharm. Pract. 2014, 27, 336–349. [Google Scholar] [CrossRef]
- Zametkin, A.J.; Rapoport, J.L. Neurobiology of Attention Deficit Disorder with Hyperactivity: Where Have We Come in 50 Years? J. Am. Acad. Child Adolesc. Psychiatry 1987, 26, 676–686. [Google Scholar] [CrossRef]
- Pliszka, S.R.; McCracken, J.T.; Maas, J.W. Catecholamines in Attention-Deficit Hyperactivity Disorder: Current Perspectives. J. Am. Acad. Child Adolesc. Psychiatry 1996, 35, 264–272. [Google Scholar] [CrossRef]
- Ulke, C.; Rullmann, M.; Huang, J.; Luthardt, J.; Becker, G.A.; Patt, M.; Meyer, P.M.; Tiepolt, S.; Hesse, S.; Sabri, O.; et al. Adult Attention-Deficit/Hyperactivity Disorder Is Associated with Reduced Norepinephrine Transporter Availability in Right Attention Networks: A (S,S)-O-[11C]Methylreboxetine Positron Emission Tomography Study. Transl. Psychiatry 2019, 9, 301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biederman, J.; Spencer, T. Attention-Deficit/Hyperactivity Disorder (Adhd) as a Noradrenergic Disorder. Biol. Psychiatry 1999, 46, 1234–1242. [Google Scholar] [CrossRef]
- Braff, D.L.; Geyer, M.A.; Swerdlow, N.R. Human Studies of Prepulse Inhibition of Startle: Normal Subjects, Patient Groups, and Pharmacological Studies. Psychopharmacology 2001, 156, 234–258. [Google Scholar] [CrossRef]
- Valsamis, B.; Schmid, S. Habituation and Prepulse Inhibition of Acoustic Startle in Rodents. J Vis Exp. 2011, 55, e3446. [Google Scholar] [CrossRef] [Green Version]
- Swerdlow, N.R.; Weber, M.; Qu, Y.; Light, G.A.; Braff, D.L. Realistic Expectations of Prepulse Inhibition in Translational Models for Schizophrenia Research. Psychopharmacology 2008, 199, 331–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swerdlow, N.R.; Geyer, M.A.; Braff, D.L. Neural Circuit Regulation of Prepulse Inhibition of Startle in the Rat: Current Knowledge and Future Challenges. Psychopharmacology 2001, 156, 194–215. [Google Scholar] [CrossRef]
- Swerdlow, N.R.; Braff, D.L.; Geyer, M.A. Sensorimotor Gating of the Startle Reflex: What We Said 25 Years Ago, What Has Happened since Then, and What Comes Next. J. Psychopharmacol. 2016, 30, 1072–1081. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, H.; Kamio, Y. Acoustic Startle Response and Its Modulation in Schizophrenia and Autism Spectrum Disorder in Asian Subjects. Schizophr. Res. 2018, 198, 16–20. [Google Scholar] [CrossRef] [PubMed]
- Swerdlow, N.R.; Light, G.A. Sensorimotor Gating Deficits in Schizophrenia: Advancing Our Understanding of the Phenotype, Its Neural Circuitry and Genetic Substrates. Schizophr. Res. 2018, 198, 1–5. [Google Scholar] [CrossRef]
- Kurzina, N.; Belskaya, A.; Gromova, A.; Ignashchenkova, A.; Gainetdinov, R.R.; Volnova, A. Modulation of Spatial Memory Deficit and Hyperactivity in Dopamine Transporter Knockout Rats via A2A-Adrenoceptors. Front. Psychiatry 2022, 13, 503. [Google Scholar] [CrossRef]
- Huss, M.; Chen, W.; Ludolph, A.G. Guanfacine Extended Release: A New Pharmacological Treatment Option in Europe. Clin. Drug Investig. 2016, 36, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Pritchett, K.; Mulder, G.B. Hebb-Williams Mazes. Contemp. Top. Lab. Anim. Sci. 2004, 43, 44–45. [Google Scholar]
- Raut, S.B.; Jadhav, K.S.; Marathe, P.A. Role of Dopamine—D2 Receptor in Spatial Memory Retention and Retrieval Determined Using Hebb-Williams Complex Maze. Indian J. Physiol. Pharmacol. 2014, 58, 191–195. [Google Scholar]
- Ptukha, M.; Fesenko, Z.; Belskaya, A.; Gromova, A.; Pelevin, A.; Kurzina, N.; Gainetdinov, R.R.; Volnova, A. Effects of Atomoxetine on Motor and Cognitive Behaviors and Brain Electrophysiological Activity of Dopamine Transporter Knockout Rats. Biomolecules 2022, 12, 1484. [Google Scholar] [CrossRef] [PubMed]
- Leahy, L.G. Diagnosis and Treatment of ADHD in Children vs Adults: What Nurses Should Know. Arch. Psychiatr. Nurs. 2018, 32, 890–895. [Google Scholar] [CrossRef] [PubMed]
- Leffa, D.T.; Caye, A.; Rohde, L.A. ADHD in Children and Adults: Diagnosis and Prognosis. Curr. Top. Behav. Neurosci. 2022, 57, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Saccaro, L.F.; Schilliger, Z.; Perroud, N.; Piguet, C. Inflammation, Anxiety, and Stress in Attention-Deficit/Hyperactivity Disorder. Biomedicines 2021, 9, 1313. [Google Scholar] [CrossRef] [PubMed]
- Liloia, D.; Crocetta, A.; Cauda, F.; Duca, S.; Costa, T.; Manuello, J. Seeking Overlapping Neuroanatomical Alterations between Dyslexia and Attention-Deficit/Hyperactivity Disorder: A Meta-Analytic Replication Study. Brain Sci. 2022, 12, 1367. [Google Scholar] [CrossRef]
- Reale, L.; Bartoli, B.; Cartabia, M.; Zanetti, M.; Costantino, M.A.; Canevini, M.P.; Termine, C.; Bonati, M.; Conte, S.; Renzetti, V.; et al. Comorbidity Prevalence and Treatment Outcome in Children and Adolescents with ADHD. Eur. Child Adolesc. Psychiatry 2017, 26, 1443–1457. [Google Scholar] [CrossRef]
- Gnanavel, S.; Sharma, P.; Kaushal, P.; Hussain, S. Attention Deficit Hyperactivity Disorder and Comorbidity: A Review of Literature. World J. Clin. Cases 2019, 7, 2420–2426. [Google Scholar] [CrossRef]
- Baboli, R.; Cao, M.; Halperin, J.M.; Li, X. Distinct Thalamic and Frontal Neuroanatomical Substrates in Children with Familial vs. Non-Familial Attention-Deficit/Hyperactivity Disorder (ADHD). Brain Sci. 2023, 13, 46. [Google Scholar] [CrossRef]
- Nyatega, C.; Qiang, L.; Adamu, M.; Kawuwa, H.B. Atypical Functional Connectivity of Limbic Network in Attention Deficit/Hyperactivity Disorder. Clin. Schizophr. Relat. Psychoses 2022, 16, 2022. [Google Scholar] [CrossRef]
- Arnsten, A.F. Catecholamine Modulation of Prefrontal Cortical Cognitive Function. Trends Cogn. Sci. 1998, 2, 436–447. [Google Scholar] [CrossRef] [PubMed]
- Dresel, S.; Krause, J.; Krause, K.H.; LaFougere, C.; Brinkbäumer, K.; Kung, H.F.; Hahn, K.; Tatsch, K. Attention Deficit Hyperactivity Disorder: Binding of [99mTc]TRODAT-1 to the Dopamine Transporter before and after Methylphenidate Treatment. Eur. J. Nucl. Med. 2000, 27, 1518–1524. [Google Scholar] [CrossRef]
- Purves-Tyson, T.D.; Owens, S.J.; Rothmond, D.A.; Halliday, G.M.; Double, K.L.; Stevens, J.; McCrossin, T.; Shannon Weickert, C. Putative Presynaptic Dopamine Dysregulation in Schizophrenia Is Supported by Molecular Evidence from Post-Mortem Human Midbrain. Transl. Psychiatry 2017, 7, e1003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palermo, G.; Ceravolo, R. Molecular Imaging of the Dopamine Transporter. Cells 2019, 8, 872. [Google Scholar] [CrossRef] [Green Version]
- Salatino-Oliveira, A.; Rohde, L.A.; Hutz, M.H. The Dopamine Transporter Role in Psychiatric Phenotypes. Am. J. Med. Genet. Part B Neuropsychiatr. Genet. 2018, 177, 211–231. [Google Scholar] [CrossRef] [Green Version]
- Kourosh-Arami, M.; Komaki, A.; Zarrindast, M.-R. Dopamine as a Potential Target for Learning and Memory: Contribution to Related Neurological Disorders. CNS Neurol. Disord.—Drug Targets 2022, 21, 1. [Google Scholar] [CrossRef]
- Madras, B.K.; Miller, G.M.; Fischman, A.J. The Dopamine Transporter and Attention-Deficit/Hyperactivity Disorder. Biol. Psychiatry 2005, 57, 1397–1409. [Google Scholar] [CrossRef]
- Ramos, B.P.; Arnsten, A.F.T. Adrenergic Pharmacology and Cognition: Focus on the Prefrontal Cortex. Pharmacol. Ther. 2007, 113, 523–536. [Google Scholar] [CrossRef] [Green Version]
- Ranjbar-Slamloo, Y.; Fazlali, Z. Dopamine and Noradrenaline in the Brain; Overlapping or Dissociate Functions? Front. Mol. Neurosci. 2020, 12, 334. [Google Scholar] [CrossRef]
- Xing, B.; Li, Y.C.; Gao, W.J. Norepinephrine versus Dopamine and Their Interaction in Modulating Synaptic Function in the Prefrontal Cortex. Brain Res. 2016, 1641, 217–233. [Google Scholar] [CrossRef] [Green Version]
- Plichta, M.M.; Vasic, N.; Wolf, R.C.; Lesch, K.-P.; Brummer, D.; Jacob, C.; Fallgatter, A.J.; Grön, G. Neural Hyporesponsiveness and Hyperresponsiveness during Immediate and Delayed Reward Processing in Adult Attention-Deficit/Hyperactivity Disorder. Biol. Psychiatry 2009, 65, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Devilbiss, D.M.; Berridge, C.W. Low-Dose Methylphenidate Actions on Tonic and Phasic Locus Coeruleus Discharge. J. Pharmacol. Exp. Ther. 2006, 319, 1327–1335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnsten, A.F.T.; Pliszka, S.R. Catecholamine Influences on Prefrontal Cortical Function: Relevance to Treatment of Attention Deficit/Hyperactivity Disorder and Related Disorders. Pharmacol. Biochem. Behav. 2011, 99, 211–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chernysheva, M.; Sych, Y.; Fomins, A.; Luis, J.; Warren, A.; Lewis, C.; Serratosa Capdevila, L.; Boehringer, R.; Amadei, E.A.; Grewe, B.F.; et al. Striatum-Projecting Prefrontal Cortex Neurons Support Working Memory Maintenance. bioRxiv 2021. [Google Scholar] [CrossRef]
- Terra, H.; Bruinsma, B.; de Kloet, S.F.; van der Roest, M.; Pattij, T.; Mansvelder, H.D. Prefrontal Cortical Projection Neurons Targeting Dorsomedial Striatum Control Behavioral Inhibition. Curr. Biol. 2020, 30, 4188–4200.e5. [Google Scholar] [CrossRef]
- Wang, F.; Yang, J.; Pan, F.; Ho, R.C.; Huang, J.H. Editorial: Neurotransmitters and Emotions. Front. Psychol. 2020, 11, 21. [Google Scholar] [CrossRef] [Green Version]
- Battaglia, S.; Cardellicchio, P.; Di Fazio, C.; Nazzi, C.; Fracasso, A.; Borgomaneri, S. The Influence of Vicarious Fear-Learning in “Infecting” Reactive Action Inhibition. Front. Behav. Neurosci. 2022, 16, 946263. [Google Scholar] [CrossRef]
- Battaglia, S.; Cardellicchio, P.; Di Fazio, C.; Nazzi, C.; Fracasso, A.; Borgomaneri, S. Stopping in (e)Motion: Reactive Action Inhibition When Facing Valence-Independent Emotional Stimuli. Front. Behav. Neurosci. 2022, 16, 998714. [Google Scholar] [CrossRef]
- Antshel, K.M.; Hargrave, T.M.; Simonescu, M.; Kaul, P.; Hendricks, K.; Faraone, S.V. Advances in Understanding and Treating ADHD. BMC Med. 2011, 9, 72. [Google Scholar] [CrossRef] [PubMed]
- Mechler, K.; Banaschewski, T.; Hohmann, S.; Häge, A. Evidence-Based Pharmacological Treatment Options for ADHD in Children and Adolescents. Pharmacol. Ther. 2022, 230, 107940. [Google Scholar] [CrossRef] [PubMed]
- Castells, X.; Blanco-Silvente, L.; Cunill, R. Amphetamines for Attention Deficit Hyperactivity Disorder (ADHD) in Adults. Cochrane Database Syst. Rev. 2018, 8, CD007813. [Google Scholar] [CrossRef] [PubMed]
- Faraone, S.V. The Pharmacology of Amphetamine and Methylphenidate: Relevance to the Neurobiology of Attention-Deficit/Hyperactivity Disorder and Other Psychiatric Comorbidities. Neurosci. Biobehav. Rev. 2018, 87, 255–270. [Google Scholar] [CrossRef]
- Budur, K.; Mathews, M.; Adetunji, B.; Mathews, M.; Mahmud, J. Non-Stimulant Treatment for Attention Deficit Hyperactivity Disorder. Psychiatry 2005, 2, 44. [Google Scholar] [PubMed]
- Higgins, G.A.; Silenieks, L.B.; MacMillan, C.; Thevarkunnel, S.; Parachikova, A.I.; Mombereau, C.; Lindgren, H.; Bastlund, J.F. Characterization of Amphetamine, Methylphenidate, Nicotine, and Atomoxetine on Measures of Attention, Impulsive Action, and Motivation in the Rat: Implications for Translational Research. Front. Pharmacol. 2020, 11, 427. [Google Scholar] [CrossRef]
- Nishitomi, K.; Yano, K.; Kobayashi, M.; Jino, K.; Kano, T.; Horiguchi, N.; Shinohara, S.; Hasegawa, M. Systemic Administration of Guanfacine Improves Food-Motivated Impulsive Choice Behavior Primarily via Direct Stimulation of Postsynaptic A2A-Adrenergic Receptors in Rats. Behav. Brain Res. 2018, 345, 21–29. [Google Scholar] [CrossRef]
- Kawaura, K.; Karasawa, J.-I.; Chaki, S.; Hikichi, H. Stimulation of Postsynapse Adrenergic A2A Receptor Improves Attention/Cognition Performance in an Animal Model of Attention Deficit Hyperactivity Disorder. Behav. Brain Res. 2014, 270, 349–356. [Google Scholar] [CrossRef]
- Álamo, C.; López-Muñoz, F.; Sánchez-García, J. Mechanism of Action of Guanfacine: A Postsynaptic Differential Approach to the Treatment of Attention Deficit Hyperactivity Disorder (Adhd). Actas Esp. Psiquiatr. 2016, 44, 107–112. [Google Scholar]
- Ramos, B.P.; Stark, D.; Verduzco, L.; Van Dyck, C.H.; Arnsten, A.F.T. A2A-Adrenoceptor Stimulation Improves Prefrontal Cortical Regulation of Behavior through Inhibition of CAMP Signaling in Aging Animals. Learn. Mem. 2006, 13, 770–776. [Google Scholar] [CrossRef] [Green Version]
- Arnsten, A.F.T. Guanfacine’s Mechanism of Action in Treating Prefrontal Cortical Disorders: Successful Translation across Species. Neurobiol. Learn. Mem. 2020, 176, 107327. [Google Scholar] [CrossRef]
- Sagvolden, T. The Alpha-2A Adrenoceptor Agonist Guanfacine Improves Sustained Attention and Reduces Overactivity and Impulsiveness in an Animal Model of Attention-Deficit/Hyperactivity Disorder (ADHD). Behav. Brain Funct. 2006, 2, 41. [Google Scholar] [CrossRef] [PubMed]
- Adinolfi, A.; Zelli, S.; Leo, D.; Carbone, C.; Mus, L.; Illiano, P.; Alleva, E.; Gainetdinov, R.R.; Adriani, W. Behavioral Characterization of DAT-KO Rats and Evidence of Asocial-like Phenotypes in DAT-HET Rats: The Potential Involvement of Norepinephrine System. Behav. Brain Res. 2019, 359, 516–527. [Google Scholar] [CrossRef] [PubMed]
- Mallien, A.S.; Becker, L.; Pfeiffer, N.; Terraneo, F.; Hahn, M.; Middelman, A.; Palme, R.; Creutzberg, K.C.; Begni, V.; Riva, M.A.; et al. Dopamine Transporter Knockout Rats Show Impaired Wellbeing in a Multimodal Severity Assessment Approach. Front. Behav. Neurosci. 2022, 16, 254. [Google Scholar] [CrossRef] [PubMed]
- Ralph, R.J.; Paulus, M.P.; Fumagalli, F.; Caron, M.G.; Geyer, M.A. Prepulse Inhibition Deficits and Perseverative Motor Patterns in Dopamine Transporter Knock-Out Mice: Differential Effects of D1 and D2 Receptor Antagonists. J. Neurosci. 2001, 21, 305–313. [Google Scholar] [CrossRef] [Green Version]
- Yamashita, M.; Fukushima, S.; Shen, H.; Hall, F.S.; Uhl, G.R.; Numachi, Y.; Kobayashi, H.; Sora, I. Norepinephrine Transporter Blockade Can Normalize the Prepulse Inhibition Deficits Found in Dopamine Transporter Knockout Mice. Neuropsychopharmacology 2006, 31, 2132–2139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bubser, M.; Koch, M. Prepulse Inhibition of the Acoustic Startle Response of Rats Is Reduced by 6-Hydroxydopamine Lesions of the Medial Prefrontal Cortex. Psychopharmacology 1994, 113, 487–492. [Google Scholar] [CrossRef]
- Gómez-Nieto, R.; Hormigo, S.; López, D.E. Prepulse Inhibition of the Auditory Startle Reflex Assessment as a Hallmark of Brainstem Sensorimotor Gating Mechanisms. Brain Sci. 2020, 10, 639. [Google Scholar] [CrossRef]
- Snyder, S.M.; Hall, J.R. A Meta-Analysis of Quantitative EEG Power Associated with Attention-Deficit Hyperactivity Disorder. J. Clin. Neurophysiol. 2006, 23, 441–456. [Google Scholar] [CrossRef]
- Loo, S.K.; Bilder, R.M.; Cho, A.L.; Sturm, A.; Cowen, J.; Walshaw, P.; Levitt, J.; Del’Homme, M.; Piacentini, J.; McGough, J.J.; et al. Effects of D-Methylphenidate, Guanfacine, and Their Combination on Electroencephalogram Resting State Spectral Power in Attention-Deficit/Hyperactivity Disorder. J. Am. Acad. Child Adolesc. Psychiatry 2016, 55, 674–682. [Google Scholar] [CrossRef] [Green Version]
- Michelini, G.; Lenartowicz, A.; Vera, J.D.; Bilder, R.M.; McGough, J.J.; McCracken, J.T.; Loo, S.K. Electrophysiological and Clinical Predictors of Methylphenidate, Guanfacine, and Combined Treatment Outcomes in Children with Attention-Deficit/Hyperactivity Disorder. J. Am. Acad. Child Adolesc. Psychiatry 2022, in press. [CrossRef] [PubMed]
- Arnsten, A.F.T.; Jin, L.E. Guanfacine for the Treatment of Cognitive Disorders: A Century of Discoveries at Yale. Yale J. Biol. Med. 2012, 85, 45. [Google Scholar] [PubMed]
- Arnsten, A.F.T. Catecholamine Influences on Dorsolateral Prefrontal Cortical Networks. Biol. Psychiatry 2011, 69, e89–e99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palva, S.; Palva, J.M. Functional Roles of Alpha-Band Phase Synchronization in Local and Large-Scale Cortical Networks. Front. Psychol. 2011, 2, 204. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Adamek, J.H.; Crocetti, D.; Mostofsky, S.H.; Ewen, J.B. Dissociation in Neural Correlates of Hyperactive/Impulsive vs. Inattentive Symptoms in Attention-Deficit/Hyperactivity Disorder. Front. Neurosci. 2022, 16, 893239. [Google Scholar] [CrossRef]
- Sallee, F.R.; Lyne, A.; Wigal, T.; McGough, J.J. Long-Term Safety and Efficacy of Guanfacine Extended Release in Children and Adolescents with Attention-Deficit/Hyperactivity Disorder. J. Child Adolesc. Psychopharmacol. 2009, 19, 215–226. [Google Scholar] [CrossRef] [Green Version]
- Iwanami, A.; Saito, K.; Fujiwara, M.; Okutsu, D.; Ichikawa, H. Safety and Efficacy of Guanfacine Extended-Release in Adults with Attention-Deficit/Hyperactivity Disorder: An Open-Label, Long-Term, Phase 3 Extension Study. BMC Psychiatry 2020, 20, 485. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Volnova, A.; Kurzina, N.; Belskaya, A.; Gromova, A.; Pelevin, A.; Ptukha, M.; Fesenko, Z.; Ignashchenkova, A.; Gainetdinov, R.R. Noradrenergic Modulation of Learned and Innate Behaviors in Dopamine Transporter Knockout Rats by Guanfacine. Biomedicines 2023, 11, 222. https://doi.org/10.3390/biomedicines11010222
Volnova A, Kurzina N, Belskaya A, Gromova A, Pelevin A, Ptukha M, Fesenko Z, Ignashchenkova A, Gainetdinov RR. Noradrenergic Modulation of Learned and Innate Behaviors in Dopamine Transporter Knockout Rats by Guanfacine. Biomedicines. 2023; 11(1):222. https://doi.org/10.3390/biomedicines11010222
Chicago/Turabian StyleVolnova, Anna, Natalia Kurzina, Anastasia Belskaya, Arina Gromova, Arseniy Pelevin, Maria Ptukha, Zoia Fesenko, Alla Ignashchenkova, and Raul R. Gainetdinov. 2023. "Noradrenergic Modulation of Learned and Innate Behaviors in Dopamine Transporter Knockout Rats by Guanfacine" Biomedicines 11, no. 1: 222. https://doi.org/10.3390/biomedicines11010222
APA StyleVolnova, A., Kurzina, N., Belskaya, A., Gromova, A., Pelevin, A., Ptukha, M., Fesenko, Z., Ignashchenkova, A., & Gainetdinov, R. R. (2023). Noradrenergic Modulation of Learned and Innate Behaviors in Dopamine Transporter Knockout Rats by Guanfacine. Biomedicines, 11(1), 222. https://doi.org/10.3390/biomedicines11010222