
Hepatoprotective Efficacy of Cycloastragenol Alleviated the Progression of Liver Fibrosis in Carbon-Tetrachloride-Treated Mice

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Experimental Protocol
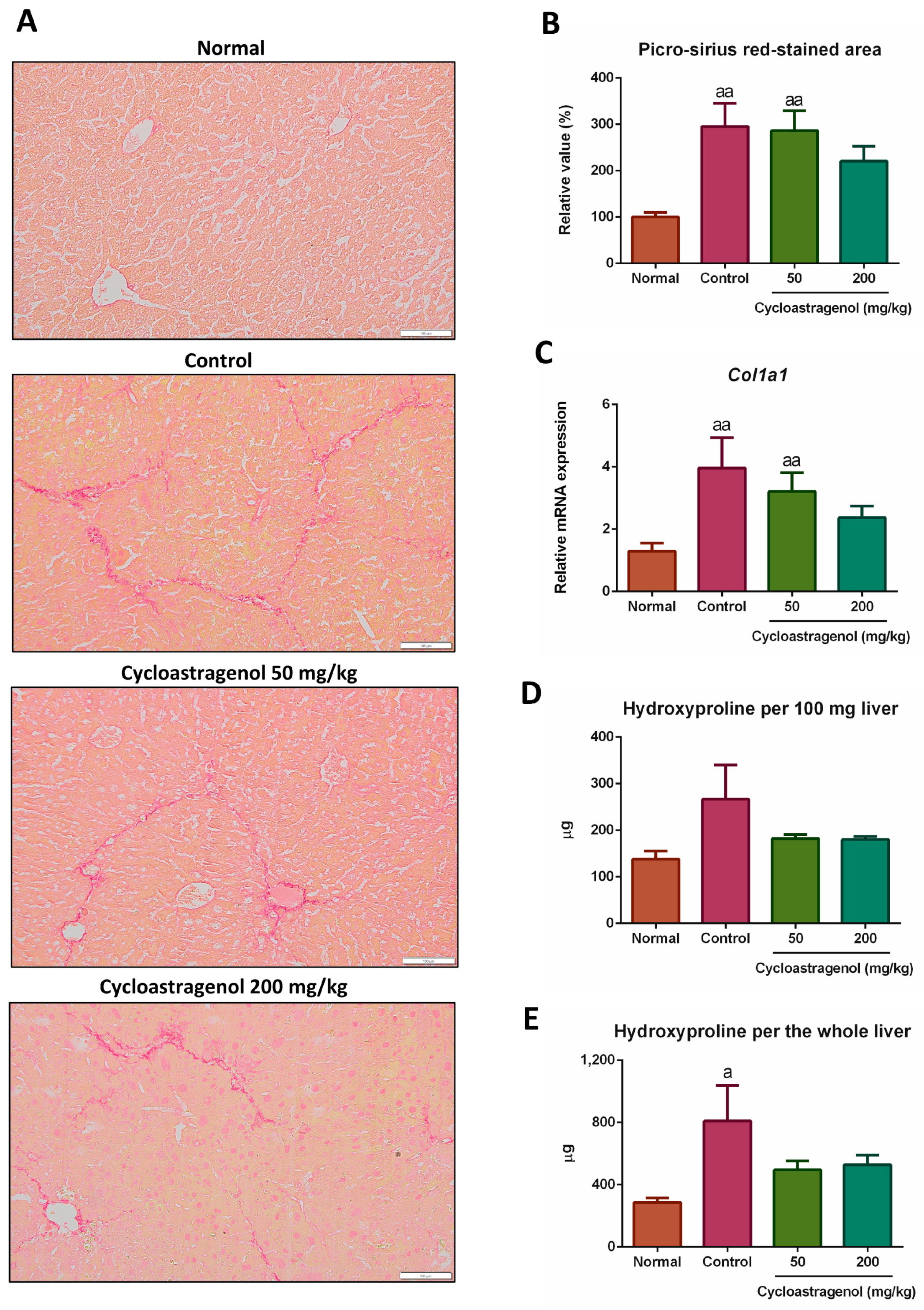
2.3. Histological Evaluations
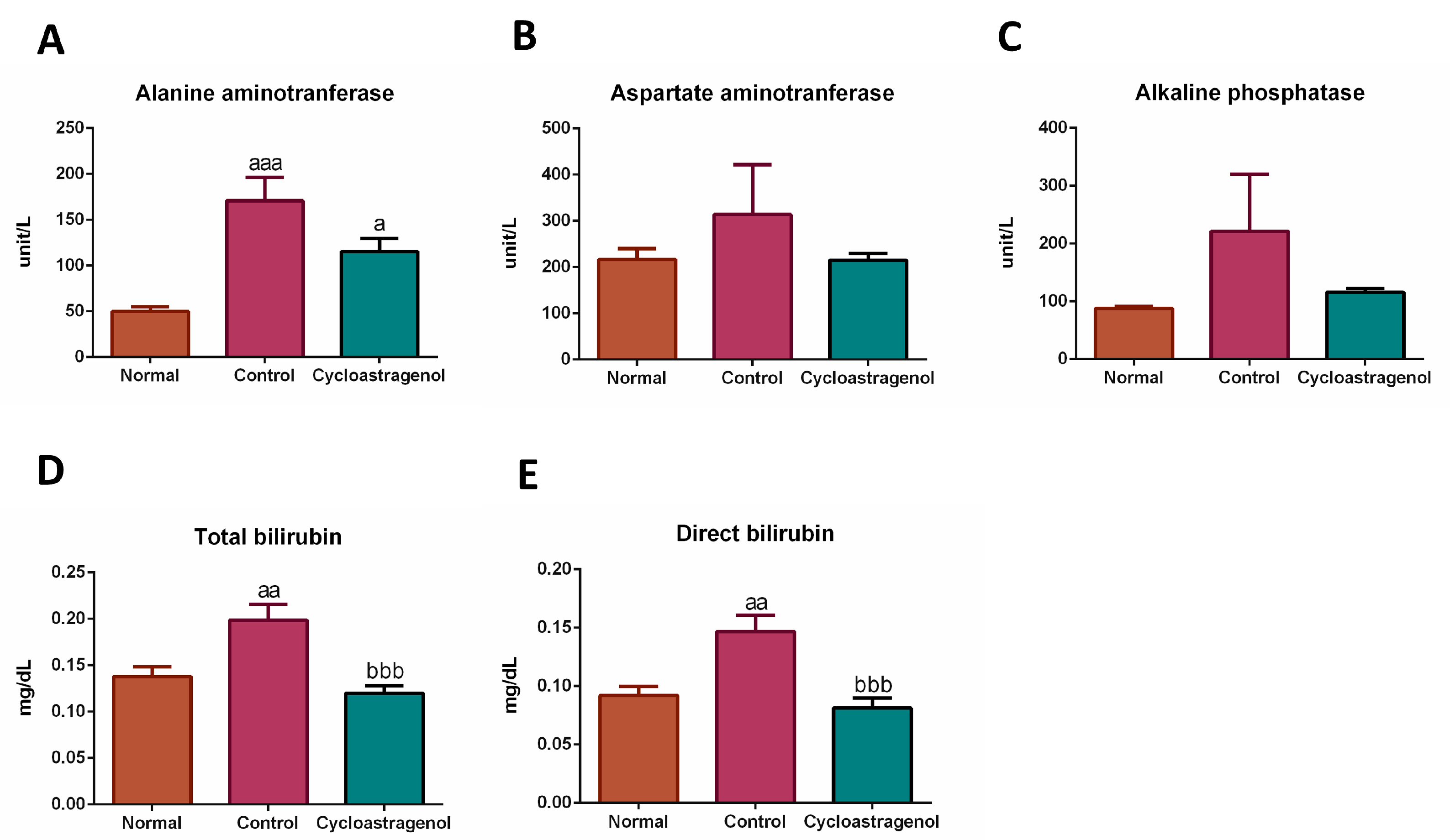
2.4. Serum Biomarker Measurements
2.5. Hepatic Hydroxyproline Assay
2.6. Total RNA and Genomic DNA Isolation
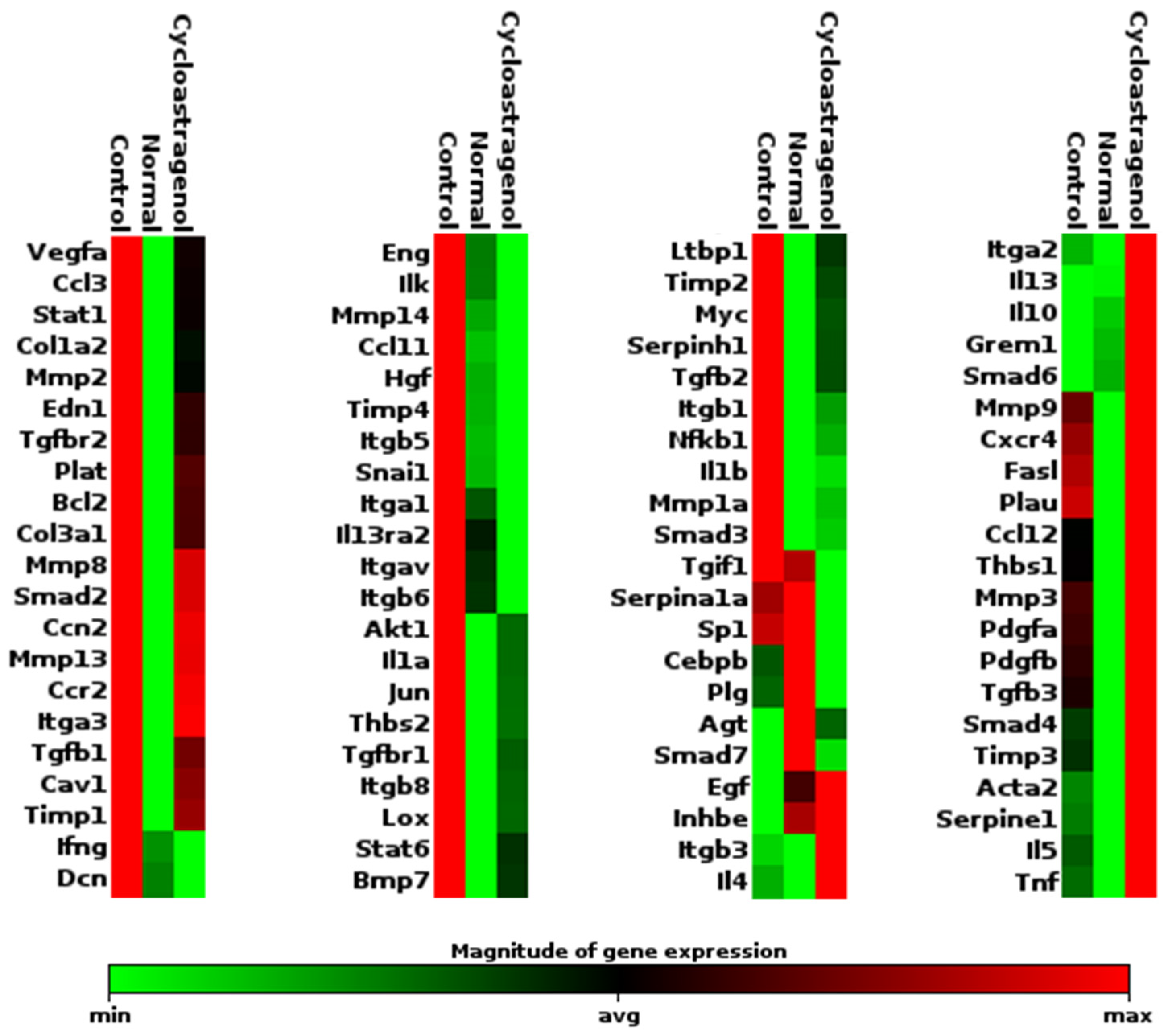
2.7. Evaluation of Gene Expression
2.8. Quantification of Telomere Length
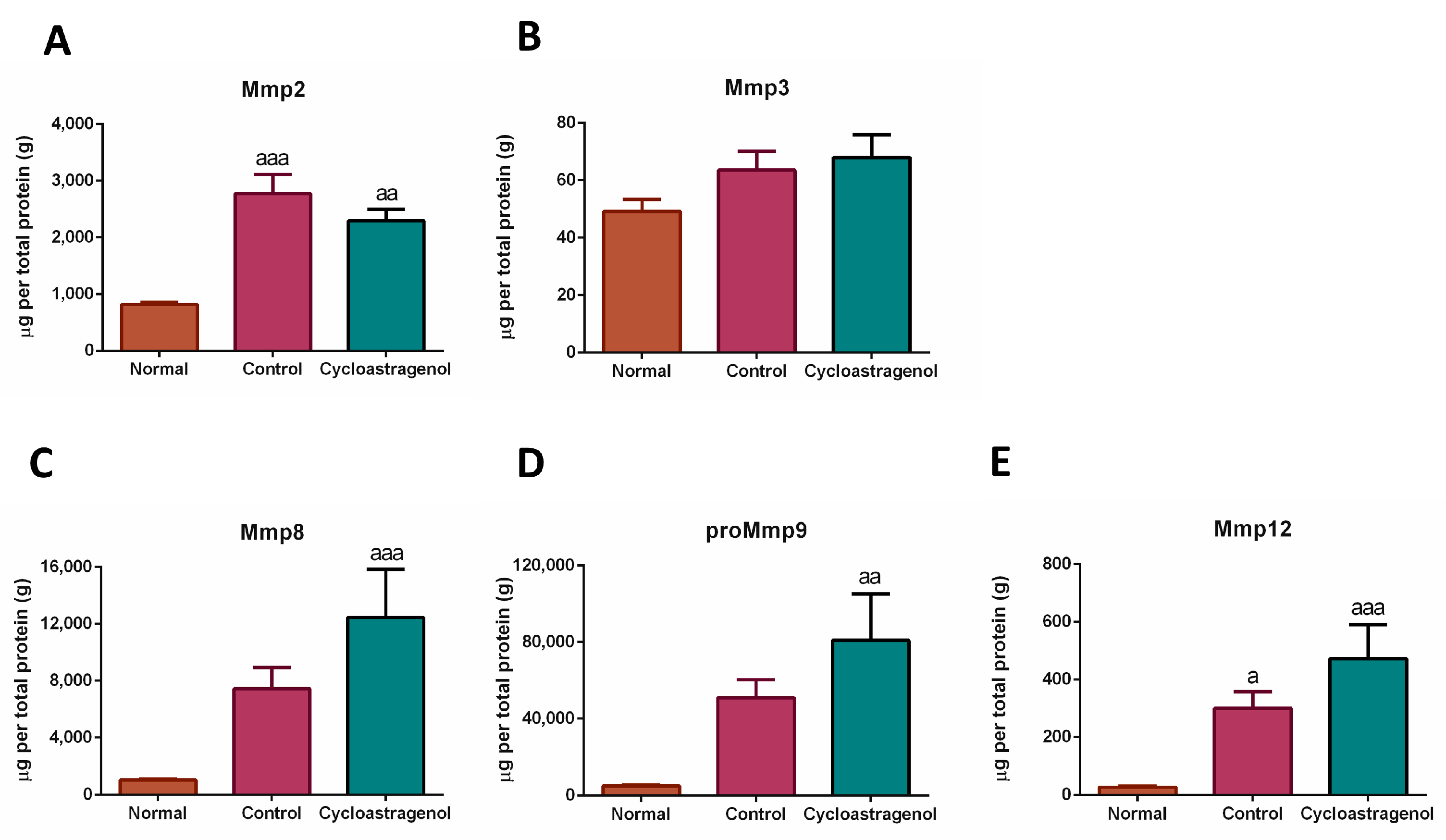
2.9. Evaluation of Protein Expression
2.10. Statistic
3. Results
3.1. Anti-Fibrotic Efficacy
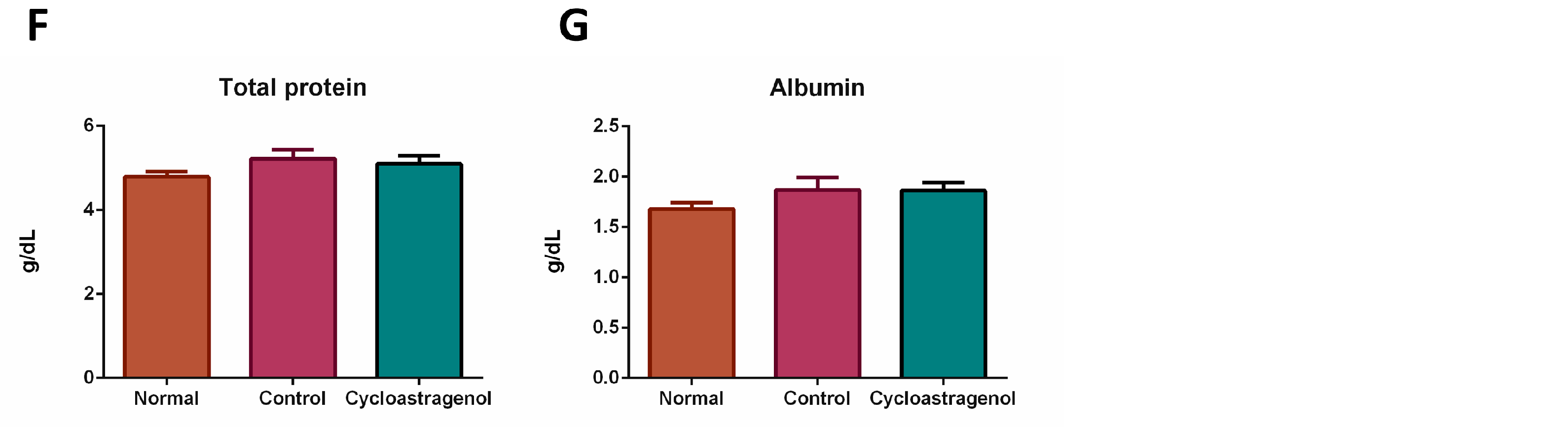
3.2. Hepatoprotective Efficacy
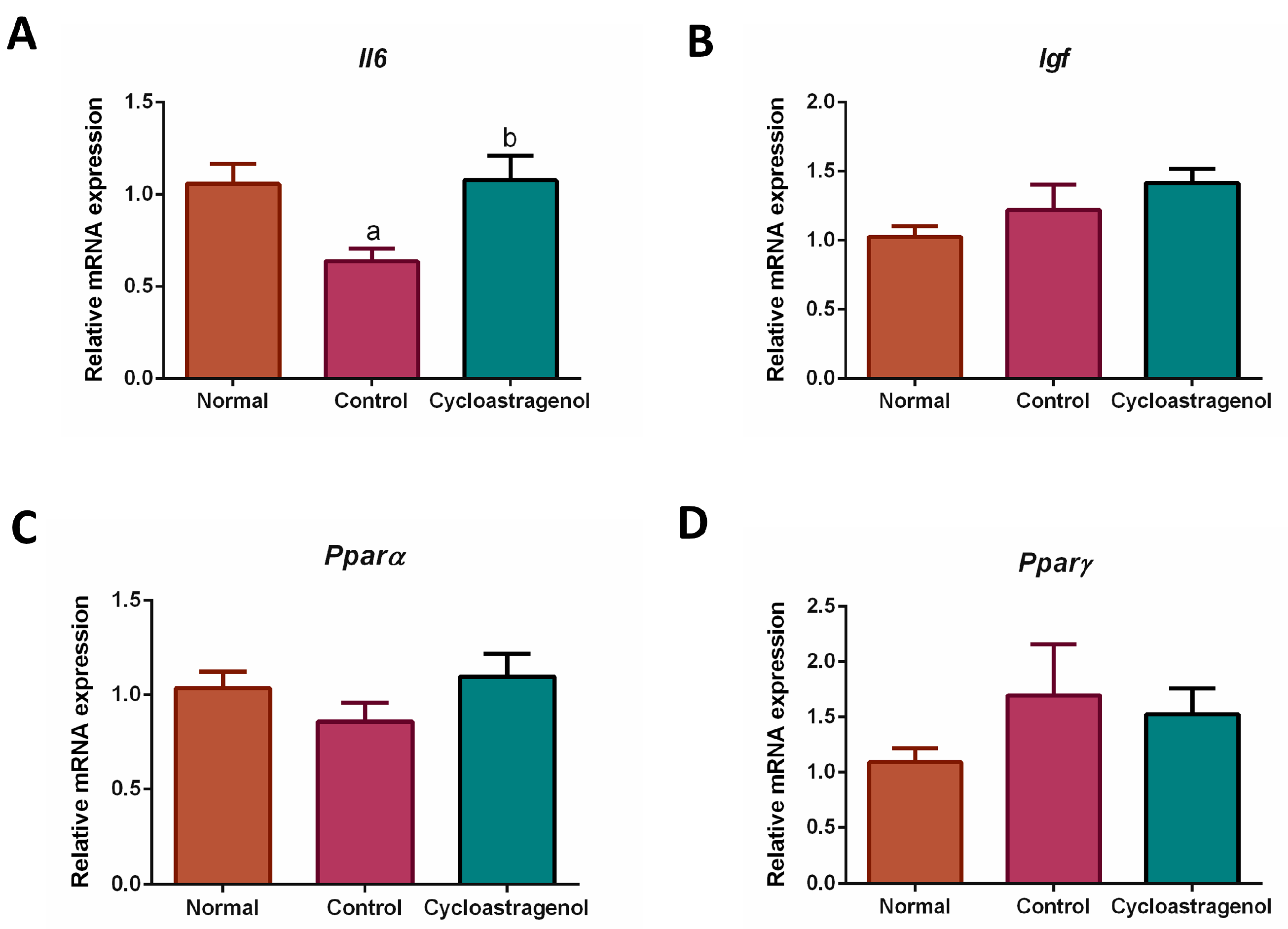
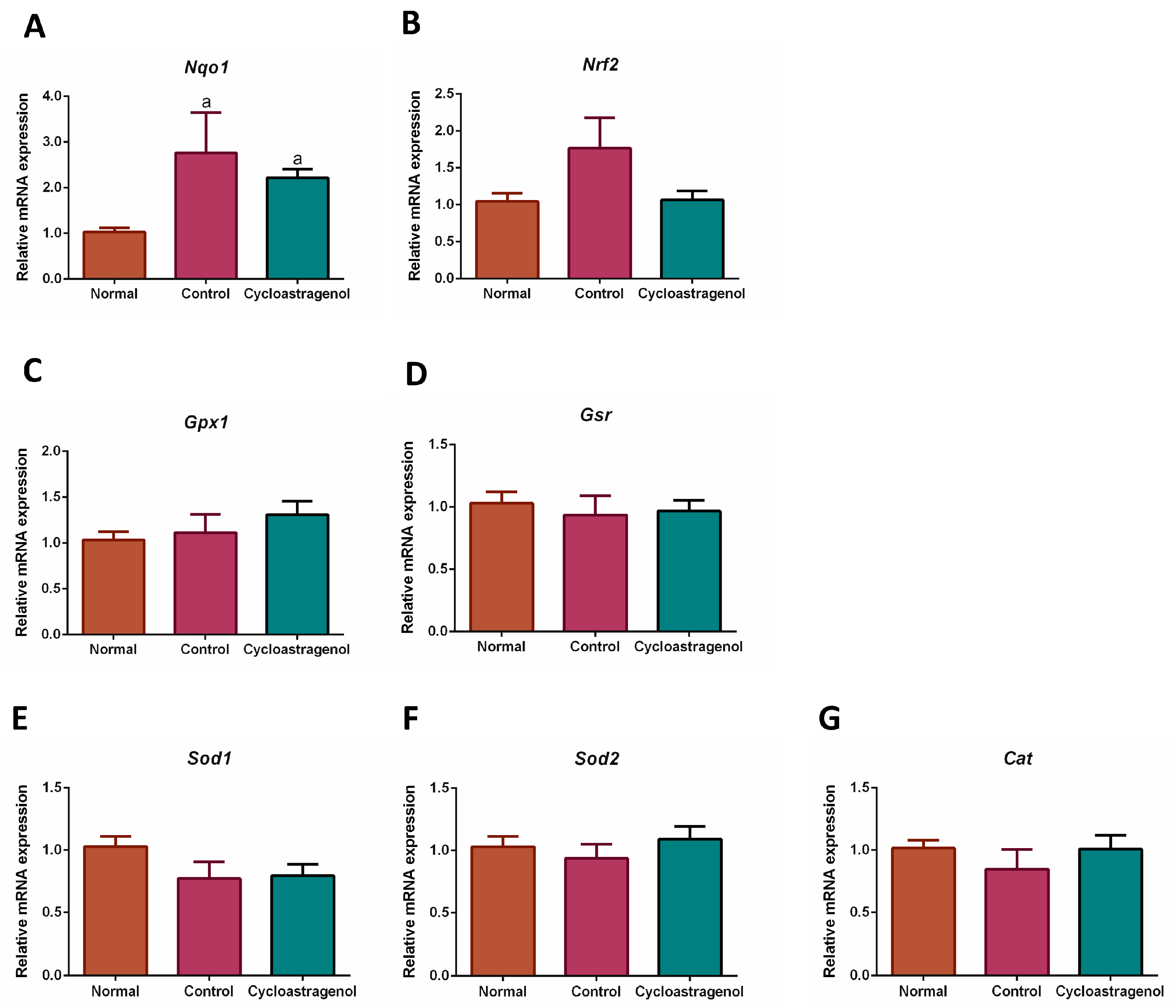
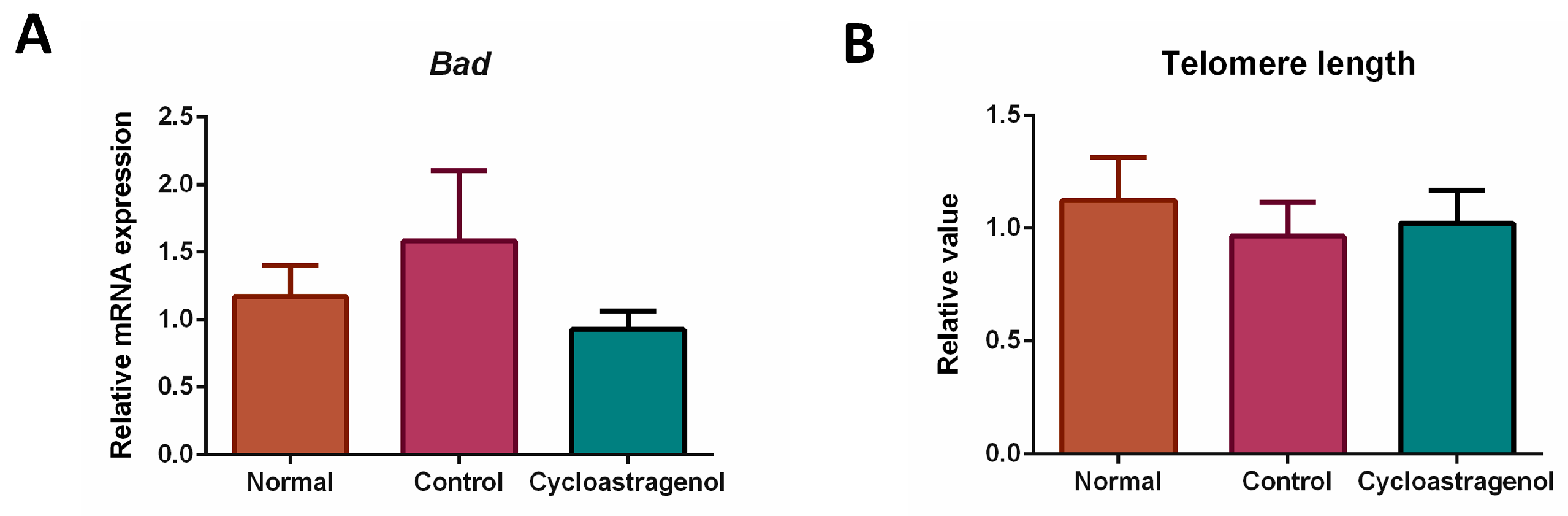
3.3. Anti-Inflammatory, Antioxidative, and Anti-Senescent Efficacy
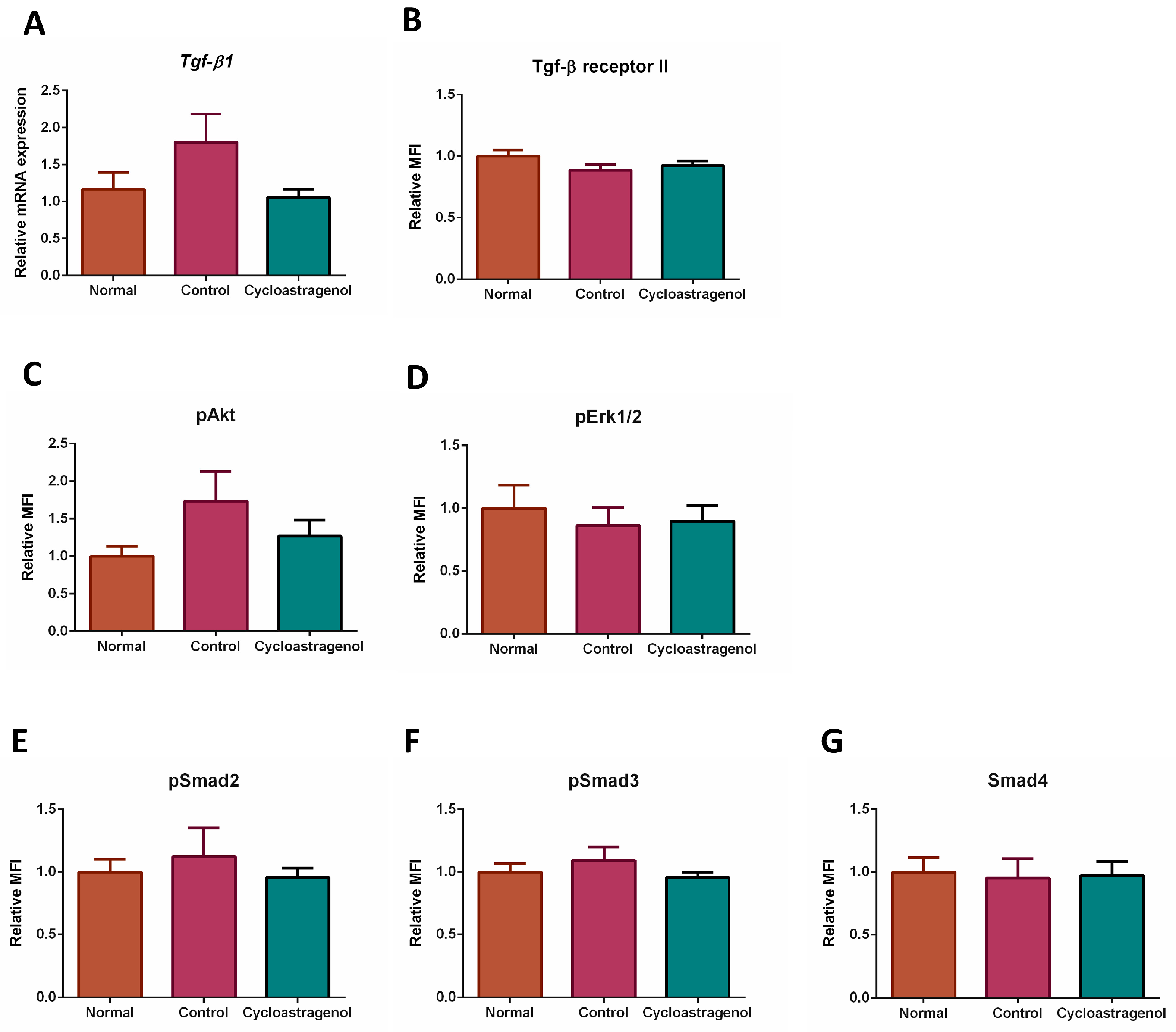
3.4. Effects on TGF-β1 Signaling Pathway
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- D’souza, S.; Lau, K.C.; Coffin, C.S.; Patel, T.R. Molecular Mechanisms of Viral Hepatitis Induced Hepatocellular Carcinoma. World J. Gastroenterol. 2020, 26, 5759–5783. [Google Scholar] [CrossRef] [PubMed]
- Lackner, C.; Tiniakos, D. Fibrosis and Alcohol-Related Liver Disease. J. Hepatol. 2019, 70, 294–304. [Google Scholar] [CrossRef] [PubMed]
- Kolaric, T.O.; Nincevic, V.; Kuna, L.; Duspara, K.; Bojanic, K.; Vukadin, S.; Raguz-Lucic, N.; Wu, G.Y.; Smolic, M. Drug-Induced Fatty Liver Disease: Pathogenesis and Treatment. J. Clin. Transl. Hepatol. 2021, 9, 731–737. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Chen, C.; Ziani, S.; Nelson, L.J.; Ávila, M.A.; Nevzorova, Y.A.; Cubero, F.J. Fibrotic Events in the Progression of Cholestatic Liver Disease. Cells 2021, 10, 1107. [Google Scholar] [CrossRef] [PubMed]
- Qu, W.; Ma, T.; Cai, J.; Zhang, X.; Zhang, P.; She, Z.; Wan, F.; Li, H. Liver Fibrosis and MAFLD: From Molecular Aspects to Novel Pharmacological Strategies. Front. Med. 2021, 8, 761538. [Google Scholar] [CrossRef] [PubMed]
- Roehlen, N.; Crouchet, E.; Baumert, T.F. Liver Fibrosis: Mechanistic Concepts and Therapeutic Perspectives. Cells 2020, 9, 875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kisseleva, T.; Brenner, D. Molecular and Cellular Mechanisms of Liver Fibrosis and Its Regression. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 151–166. [Google Scholar] [CrossRef]
- Duarte, S.; Baber, J.; Fujii, T.; Coito, A.J. Matrix Metalloproteinases in Liver Injury, Repair and Fibrosis. Matrix Biol. 2015, 44–46, 147–156. [Google Scholar] [CrossRef]
- Hoare, M.; Das, T.; Alexander, G. Ageing, Telomeres, Senescence, and Liver Injury. J. Hepatol. 2010, 53, 950–961. [Google Scholar] [CrossRef] [Green Version]
- Dewidar, B.; Meyer, C.; Dooley, S.; Meindl-Beinker, A.N. TGF-β in Hepatic Stellate Cell Activation and Liver Fibrogenesis-Updated 2019. Cells 2019, 8, 1419. [Google Scholar] [CrossRef]
- Yu, Y.; Zhou, L.; Yang, Y.; Liu, Y. Cycloastragenol: An Exciting Novel Candidate for Age-Associated Diseases. Exp. Ther. Med. 2018, 16, 2175–2182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yongping, M.; Zhang, X.; Xuewei, L.; Fan, W.; Chen, J.; Zhang, H.; Chen, G.; Liu, C.; Liu, P. Astragaloside Prevents BDL-Induced Liver Fibrosis through Inhibition of Notch Signaling Activation. J. Ethnopharmacol. 2015, 169, 200–209. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Tong, X.; Ren, S.; Wang, X.; Chen, J.; Mu, Y.; Sun, M.; Chen, G.; Zhang, H.; Liu, P. Synergistic Anti-Liver Fibrosis Actions of Total Astragalus Saponins and Glycyrrhizic Acid via TGF-β1/Smads Signaling Pathway Modulation. J. Ethnopharmacol. 2016, 190, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Kilkenny, C.; Browne, W.; Cuthill, I.C.; Emerson, M.; Altman, D.G. NC3Rs Reporting Guidelines Working Group Animal Research: Reporting in Vivo Experiments: The ARRIVE Guidelines. Br. J. Pharmacol. 2010, 160, 1577–1579. [Google Scholar] [CrossRef]
- Lattouf, R.; Younes, R.; Lutomski, D.; Naaman, N.; Godeau, G.; Senni, K.; Changotade, S. Picrosirius Red Staining: A Useful Tool to Appraise Collagen Networks in Normal and Pathological Tissues. J. Histochem. Cytochem. 2014, 62, 751–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suvarna, S.K.; Layton, C.; Bancroft, J.D. (Eds.) Bancroft’s Theory and Practice of Histological Techniques, 8th ed.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 126–138. [Google Scholar]
- Kim, Y.O.; Popov, Y.; Schuppan, D. Optimized Mouse Models for Liver Fibrosis. Methods Mol. Biol. 2017, 1559, 279–296. [Google Scholar] [CrossRef] [PubMed]
- Zarrinpar, A.; Busuttil, R.W. Liver Transplantation: Past, Present and Future. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 434–440. [Google Scholar] [CrossRef]
- Caligiuri, A.; Gentilini, A.; Pastore, M.; Gitto, S.; Marra, F. Cellular and Molecular Mechanisms Underlying Liver Fibrosis Regression. Cells 2021, 10, 2759. [Google Scholar] [CrossRef]
- Shen, C.Y.; Jiang, J.G.; Yang, L.; Wang, D.W.; Zhu, W. Anti-Ageing Active Ingredients from Herbs and Nutraceuticals Used in Traditional Chinese Medicine: Pharmacological Mechanisms and Implications for Drug Discovery. Br. J. Pharmacol. 2017, 174, 1395–1425. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.D.; Lu, J.; Yan, J.; Zhang, H. Research Progress of Preparation Technology and Pharmacological Effect of Cy-cloastragenol. Chin. J. New Drugs 2016, 16, 1872–1875. [Google Scholar]
- Scholten, D.; Trebicka, J.; Liedtke, C.; Weiskirchen, R. The Carbon Tetrachloride Model in Mice. Lab. Anim. 2015, 49, 4–11. [Google Scholar] [CrossRef] [PubMed]
- Weber, L.W.D.; Boll, M.; Stampfl, A. Hepatotoxicity and Mechanism of Action of Haloalkanes: Carbon Tetrachloride as a Toxicological Model. Crit. Rev. Toxicol. 2003, 33, 105–136. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Duan, Q.; Wu, R.; Harris, E.N.; Su, Q. Pathophysiological Communication between Hepatocytes and Non-Parenchymal Cells in Liver Injury from NAFLD to Liver Fibrosis. Adv. Drug Deliv. Rev. 2021, 176, 113869. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, R.; Haag, M.; Schaeffeler, E.; Nies, A.T.; Zanger, U.M.; Schwab, M. Systemic Regulation of Bilirubin Homeostasis: Potential Benefits of Hyperbilirubinemia. Hepatology 2018, 67, 1609–1619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naim, A.; Pan, Q.; Baig, M.S. Matrix Metalloproteinases (MMPs) in Liver Diseases. J. Clin. Exp. Hepatol. 2017, 7, 367–372. [Google Scholar] [CrossRef] [PubMed]
- Préaux, A.M.; Mallat, A.; Nhieu, J.T.; D’Ortho, M.P.; Hembry, R.M.; Mavier, P. Matrix Metalloproteinase-2 Activation in Human Hepatic Fibrosis Regulation by Cell-Matrix Interactions. Hepatology 1999, 30, 944–950. [Google Scholar] [CrossRef]
- Siller-López, F.; Sandoval, A.; Salgado, S.; Salazar, A.; Bueno, M.; Garcia, J.; Vera, J.; Gálvez, J.; Hernández, I.; Ramos, M.; et al. Treatment with Human Metalloproteinase-8 Gene Delivery Ameliorates Experimental Rat Liver Cirrhosis. Gastroenterology 2004, 126, 1122–1133. [Google Scholar] [CrossRef]
- Wu, Y.; Lu, S.; Huang, X.; Liu, Y.; Huang, K.; Liu, Z.; Xu, W.; Zhu, W.; Hou, J.; Liu, H.; et al. Targeting CIAPs Attenuates CCl4-Induced Liver Fibrosis by Increasing MMP9 Expression Derived from Neutrophils. Life Sci. 2022, 289, 120235. [Google Scholar] [CrossRef]
- Pellicoro, A.; Aucott, R.L.; Ramachandran, P.; Robson, A.J.; Fallowfield, J.A.; Snowdon, V.K.; Hartland, S.N.; Vernon, M.; Duffield, J.S.; Benyon, R.C.; et al. Elastin Accumulation Is Regulated at the Level of Degradation by Macrophage Metalloelastase (MMP-12) during Experimental Liver Fibrosis. Hepatology 2012, 55, 1965–1975. [Google Scholar] [CrossRef]
- Naseem, S.; Hussain, T.; Manzoor, S. Interleukin-6: A Promising Cytokine to Support Liver Regeneration and Adaptive Immunity in Liver Pathologies. Cytokine Growth Factor Rev. 2018, 39, 36–45. [Google Scholar] [CrossRef]
- Nasir, G.A.; Mohsin, S.; Khan, M.; Shams, S.; Ali, G.; Khan, S.N.; Riazuddin, S. Mesenchymal Stem Cells and Interleukin-6 Attenuate Liver Fibrosis in Mice. J. Transl. Med. 2013, 11, 78. [Google Scholar] [CrossRef]
- Aleksunes, L.M.; Goedken, M.; Manautou, J.E. Up-Regulation of NAD(P)H Quinone Oxidoreductase 1 during Human Liver Injury. World J. Gastroenterol. 2006, 12, 1937–1940. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Wu, J.; Li, J.; Liu, Y.; Zheng, X.; Du, M.; Zhou, L.; Yang, Y.; Luo, S.; Hu, W.; et al. Cycloastragenol Prevents Age-Related Bone Loss: Evidence in d-Galactose-Treated and Aged Rats. Biomed. Pharmacother. 2020, 128, 110304. [Google Scholar] [CrossRef] [PubMed]
- McCracken, J.M.; Chalise, P.; Briley, S.M.; Dennis, K.L.; Jiang, L.; Duncan, F.E.; Pritchard, M.T. C57BL/6 Substrains Exhibit Different Responses to Acute Carbon Tetrachloride Exposure: Implications for Work Involving Transgenic Mice. Gene Expr. 2017, 17, 187–205. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Huang, C.; Wu, Q.; Su, Y.; Wang, X.; Xuan, Z.; Wang, Y.; Xu, F.; Ge, C. Sini San Ameliorates CCl4-Induced Liver Fibrosis in Mice by Inhibiting AKT-Mediated Hepatocyte Apoptosis. J. Ethnopharmacol. 2022, 303, 115965. [Google Scholar] [CrossRef]
- Gu, M.; Zhang, S.; Zhao, Y.; Huang, J.; Wang, Y.; Li, Y.; Fan, S.; Yang, L.; Ji, G.; Tong, Q.; et al. Cycloastragenol Improves Hepatic Steatosis by Activating Farnesoid X Receptor Signalling. Pharmacol. Res. 2017, 121, 22–32. [Google Scholar] [CrossRef]
- Schwabl, P.; Hambruch, E.; Seeland, B.A.; Hayden, H.; Wagner, M.; Garnys, L.; Strobel, B.; Schubert, T.-L.; Riedl, F.; Mitteregger, D.; et al. The FXR Agonist PX20606 Ameliorates Portal Hypertension by Targeting Vascular Remodelling and Sinusoidal Dysfunction. J. Hepatol. 2017, 66, 724–733. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Ge, C.; Zhou, J.; Guo, Y.; Cui, S.; Huang, N.; Yan, T.; Cao, L.; Che, Y.; Zheng, Q.; et al. Noncanonical Farnesoid X Receptor Signaling Inhibits Apoptosis and Impedes Liver Fibrosis. EBioMedicine 2018, 37, 322–333. [Google Scholar] [CrossRef] [Green Version]
- Carino, A.; Cipriani, S.; Marchianò, S.; Biagioli, M.; Santorelli, C.; Donini, A.; Zampella, A.; Monti, M.C.; Fiorucci, S. BAR502, a Dual FXR and GPBAR1 Agonist, Promotes Browning of White Adipose Tissue and Reverses Liver Steatosis and Fibrosis. Sci. Rep. 2017, 7, 42801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, G.; Chen, W.; Wang, P.; Zhan, T.; Zheng, W.; Gu, Z.; Wang, X.; Ji, X.; Sun, Y. Inhibition of NLRP3 Inflammasome-Mediated Pyroptosis in Macrophage by Cycloastragenol Contributes to Amelioration of Imiquimod-Induced Psoriasis-Like Skin Inflammation in Mice. Int. Immunopharmacol. 2019, 74, 105682. [Google Scholar] [CrossRef] [PubMed]
- Luangmonkong, T.; Pransin, C.; Nopphalee, L.; Meechai, S.; Chunya, S.; Rattanavaraha, A.; Kaewnoppharat, N.; Khuituan, T.; Bunyakiat, S.; Parichatikanond, W. Carbon Tetrachloride-Induced Acute Liver Toxicity: Selecting Dosage and Biomarkers for Evaluating Hepatoprotective Drugs in ICR Outbred Mice. Pharm. Sci. Asia 2022, 49, 534–542. [Google Scholar] [CrossRef]
- Tuttle, A.H.; Philip, V.M.; Chesler, E.J.; Mogil, J.S. Comparing Phenotypic Variation between Inbred and Outbred Mice. Nat. Methods 2018, 15, 994–996. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Forward Primers (5′–3′) | Reverse Primers (5′–3′) |
|---|---|---|
| Bad | CTCCGAAGGATGAGCGATGAG | CTCCGAAGGATGAGCGATGAG |
| Cat | GGAGGCGGGAACCCAATAG | GGAGGCGGGAACCCAATAG |
| Col1a1 | TGACTGGAAGAGCGGAGAGT | ATCCATCGGTCATGCTCTCT |
| Gapdh | ACAGTCCATGCCATCACTGC | GATCCACGACGGACACATTG |
| Gpx1 | CCACCGTGTATGCCTTCTCC | AGAGAGACGCGACATTCTCAAT |
| Gsr | CACGGCTATGCAACATTCGC | GTGTGGAGCGGTAAACTTTTTC |
| Igf | TCGTGGGATGGGTGCTTT | TGAAGACAGTAGGGAAGAGACAAG |
| Il6 | TCCATCCAGTTGCCTTCT | TAAGCCTCCGACTTGTGAA |
| Nqo1 | AGGATGGGAGGTACTCGAATC | TGCTAGAGATGACTCGGAAGG |
| Nrf2 | CTGAACTCCTGGACGGGACTA | CGGTGGGTCTCCGTAAATGG |
| Pparα | CACTTGCTCACTACTGTCCTT | GATGCTGGTATCGGCTCAA |
| Pparγ | GGTGCTCCAGAAGATGACAGA | TCAGCGGGTGGGACTTTC |
| Sod1 | AACCAGTTGTGTTGTCAGGAC | CCACCATGTTTCTTAGAGTGAGG |
| Sod2 | TGGACAAACCTGAGCCCTAAG | CCCAAAGTCACGCTTGATAGC |
| Tgf-β1 | GGTTCATGTCATGGATGGTGC | TGACGTCACTGGAGTTGTACGG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luangmonkong, T.; Puphancharoensuk, P.; Tongsongsang, V.; Olinga, P.; Parichatikanond, W. Hepatoprotective Efficacy of Cycloastragenol Alleviated the Progression of Liver Fibrosis in Carbon-Tetrachloride-Treated Mice. Biomedicines 2023, 11, 231. https://doi.org/10.3390/biomedicines11010231
Luangmonkong T, Puphancharoensuk P, Tongsongsang V, Olinga P, Parichatikanond W. Hepatoprotective Efficacy of Cycloastragenol Alleviated the Progression of Liver Fibrosis in Carbon-Tetrachloride-Treated Mice. Biomedicines. 2023; 11(1):231. https://doi.org/10.3390/biomedicines11010231
Chicago/Turabian StyleLuangmonkong, Theerut, Pittaya Puphancharoensuk, Varisara Tongsongsang, Peter Olinga, and Warisara Parichatikanond. 2023. "Hepatoprotective Efficacy of Cycloastragenol Alleviated the Progression of Liver Fibrosis in Carbon-Tetrachloride-Treated Mice" Biomedicines 11, no. 1: 231. https://doi.org/10.3390/biomedicines11010231
APA StyleLuangmonkong, T., Puphancharoensuk, P., Tongsongsang, V., Olinga, P., & Parichatikanond, W. (2023). Hepatoprotective Efficacy of Cycloastragenol Alleviated the Progression of Liver Fibrosis in Carbon-Tetrachloride-Treated Mice. Biomedicines, 11(1), 231. https://doi.org/10.3390/biomedicines11010231