

Synergistic Power of Piceatannol and/or Vitamin D in Bleomycin-Induced Pulmonary Fibrosis In Vivo: A Preliminary Study

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Drugs and Chemicals
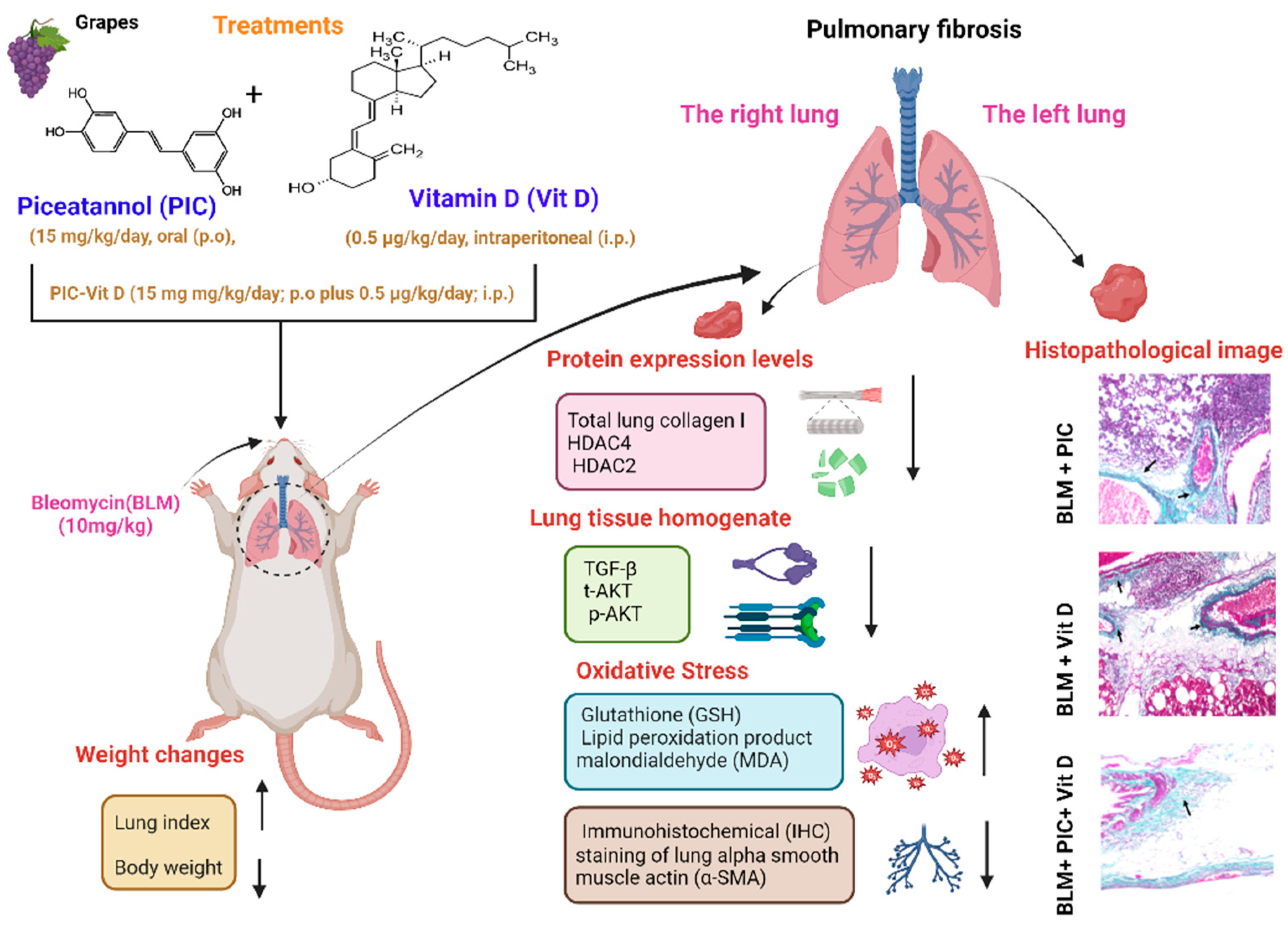
2.3. Experimental Design
2.4. Tissue Sampling
2.5. Tissue Homogenate Preparation
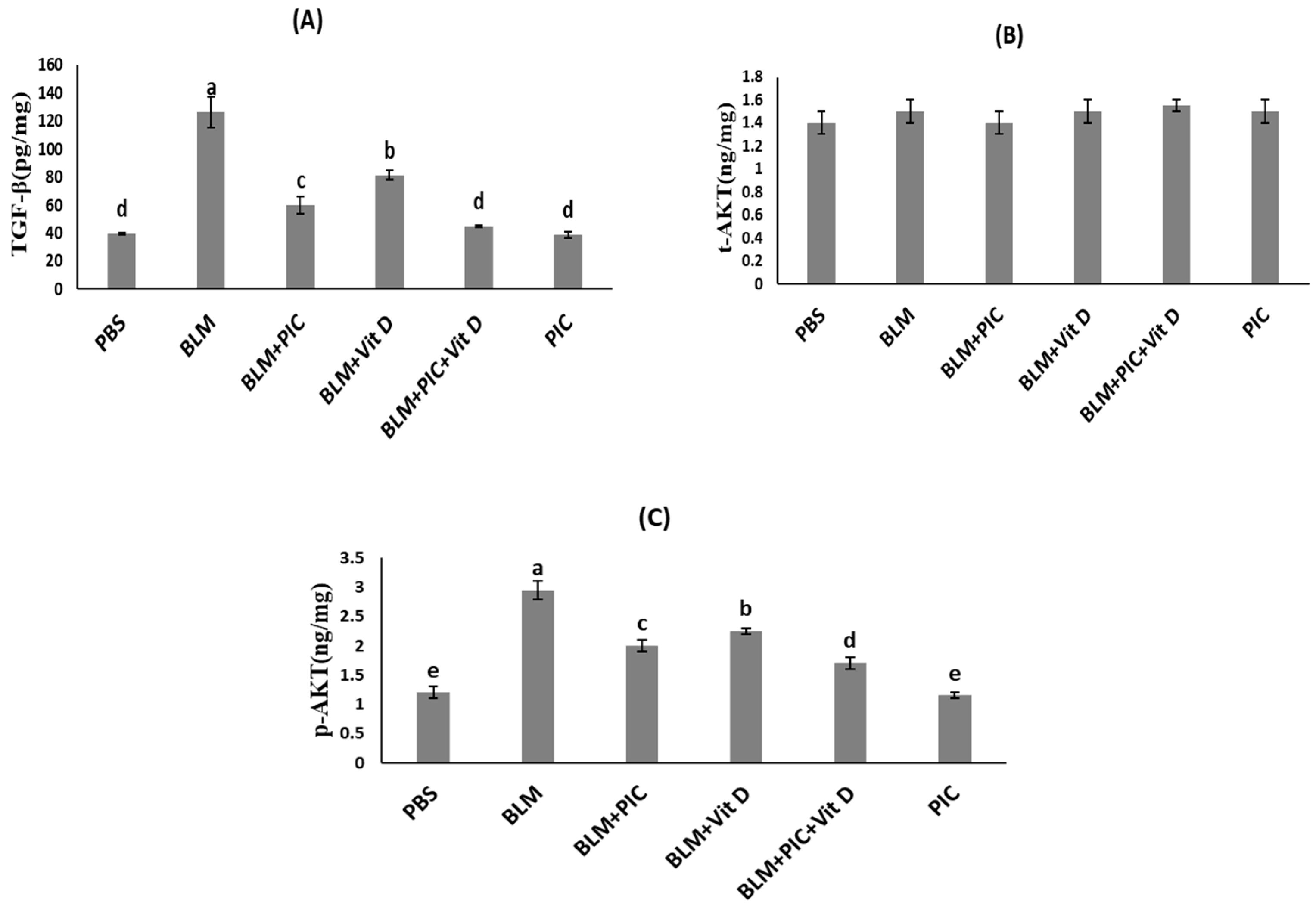
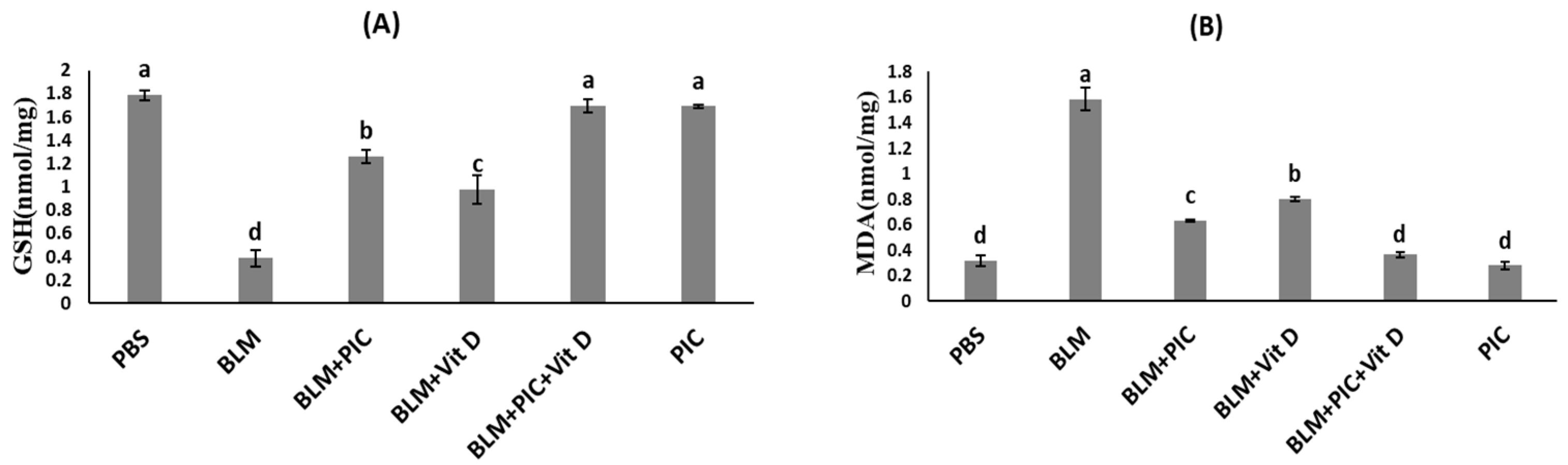
2.6. Measurements of Lung TGF-β, t-AKT, p-AKT, GSH, and MDA Levels Using ELISA Kits
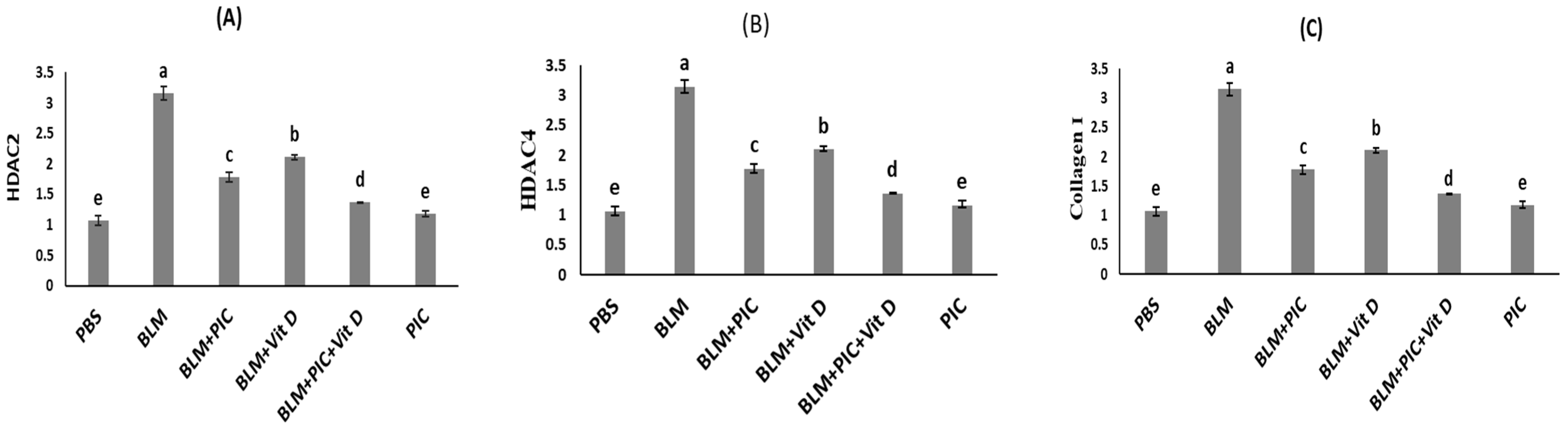
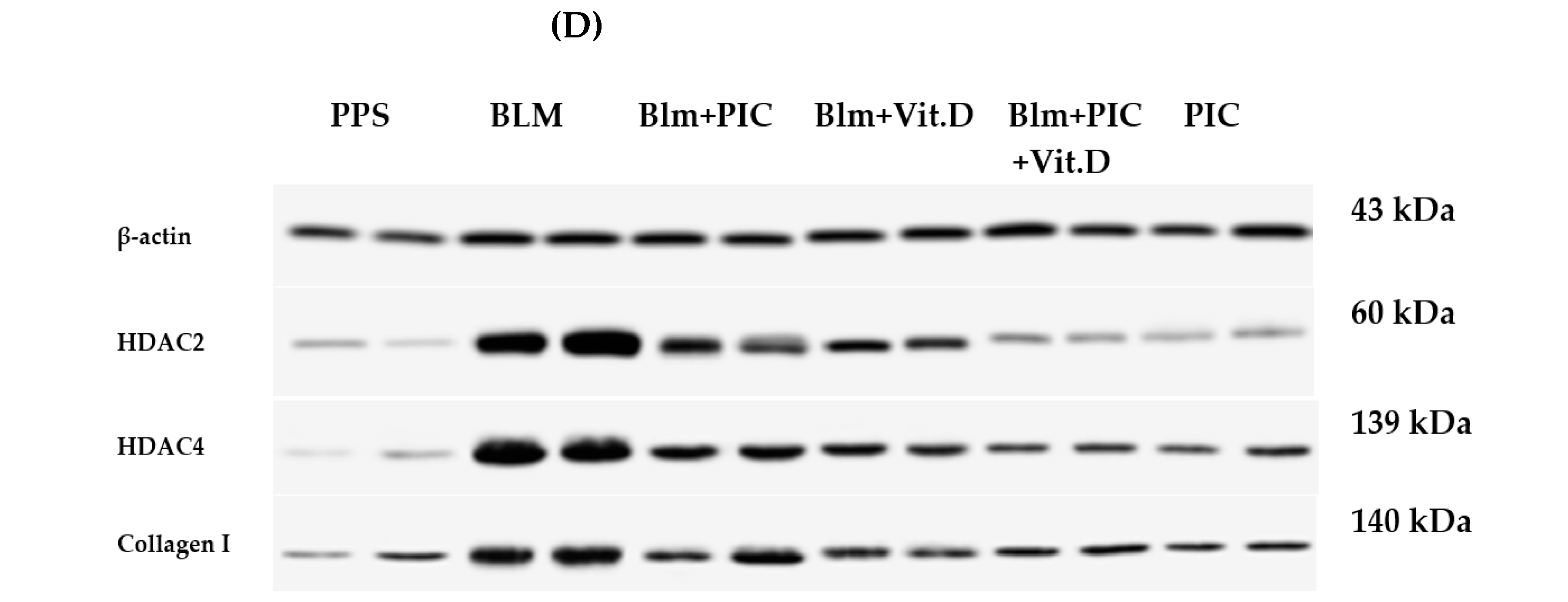
2.7. Western Blotting Analysis of HDAC2, HDAC4 and Collagen I
2.8. Immunohistochemical Staining and Image Analysis for α-SMA
2.9. Histopathological Examination of the Lung Tissues
2.10. Statistical Analysis
3. Results
3.1. Effects of Piceatannol, Vitamin D, and a Combination of the Two on Lung TGF-β, t-AKT, and p-AKT Concentrations
3.2. Effects of Piceatannol, Vitamin D, and a Combination of the Two on the Antioxidant Status of Lung Tissues
3.3. Effects of Piceatannol, Vitamin D, and a Combination of the Two on HDAC2, HDAC4, and Collagen I Concentrations in the Lungs
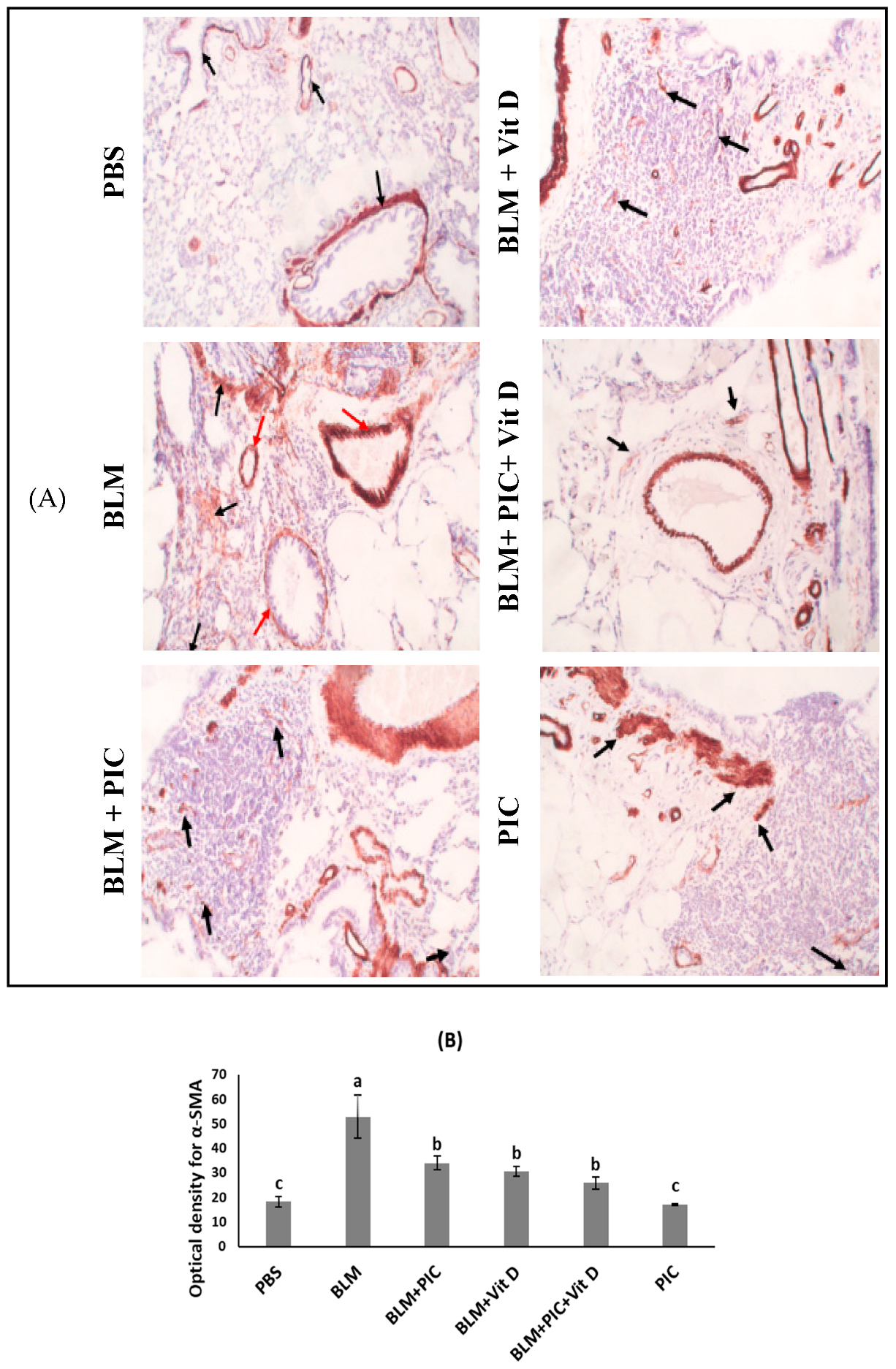
3.4. Effects of Piceatannol, Vitamin D, and a Combination of the Two on Immunohistochemical Staining for α-SMA in Lung Tissue
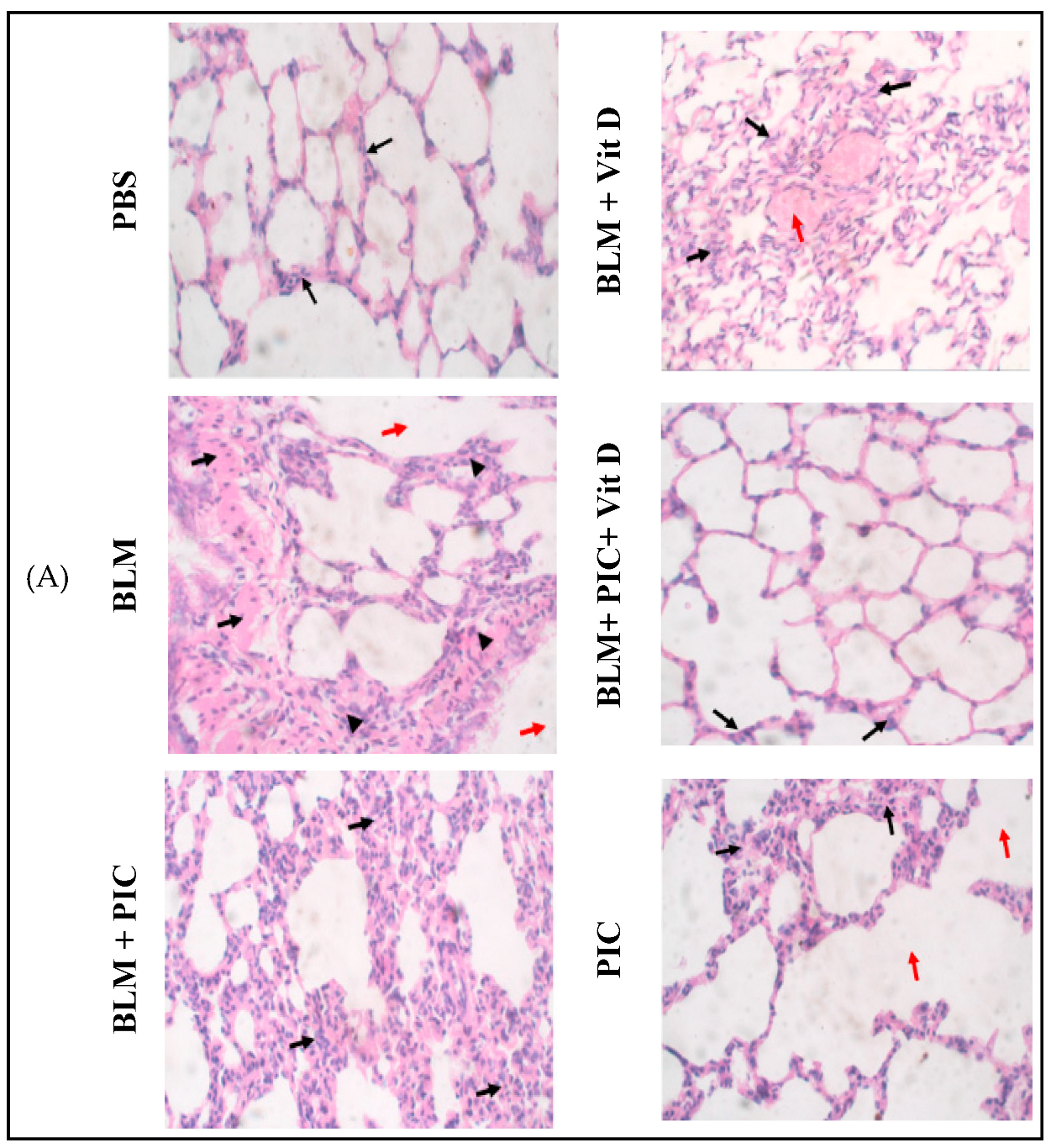
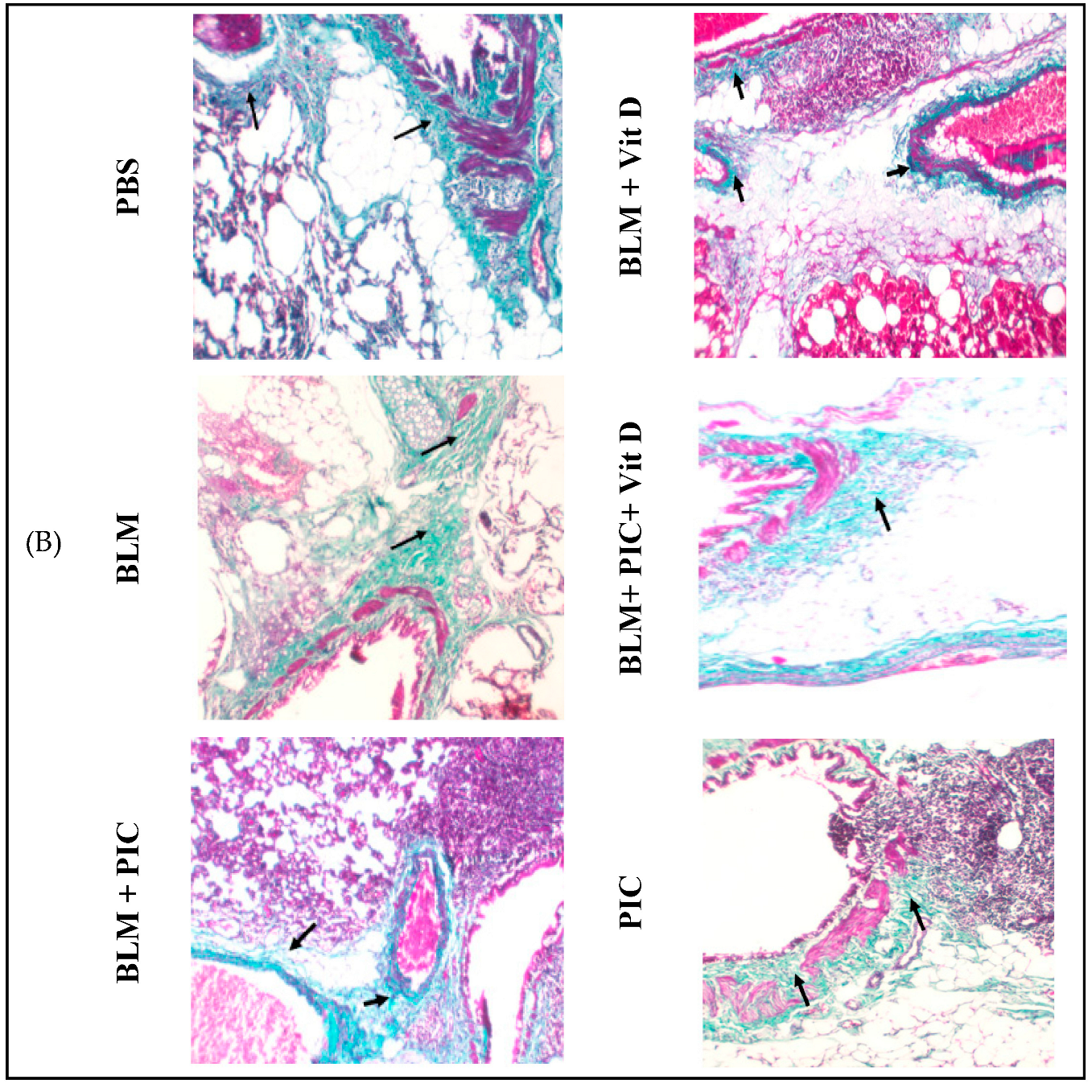
3.5. Effects of Piceatannol, Vitamin D, and a Combination of the Two on BLM-Induced Histopathological Changes
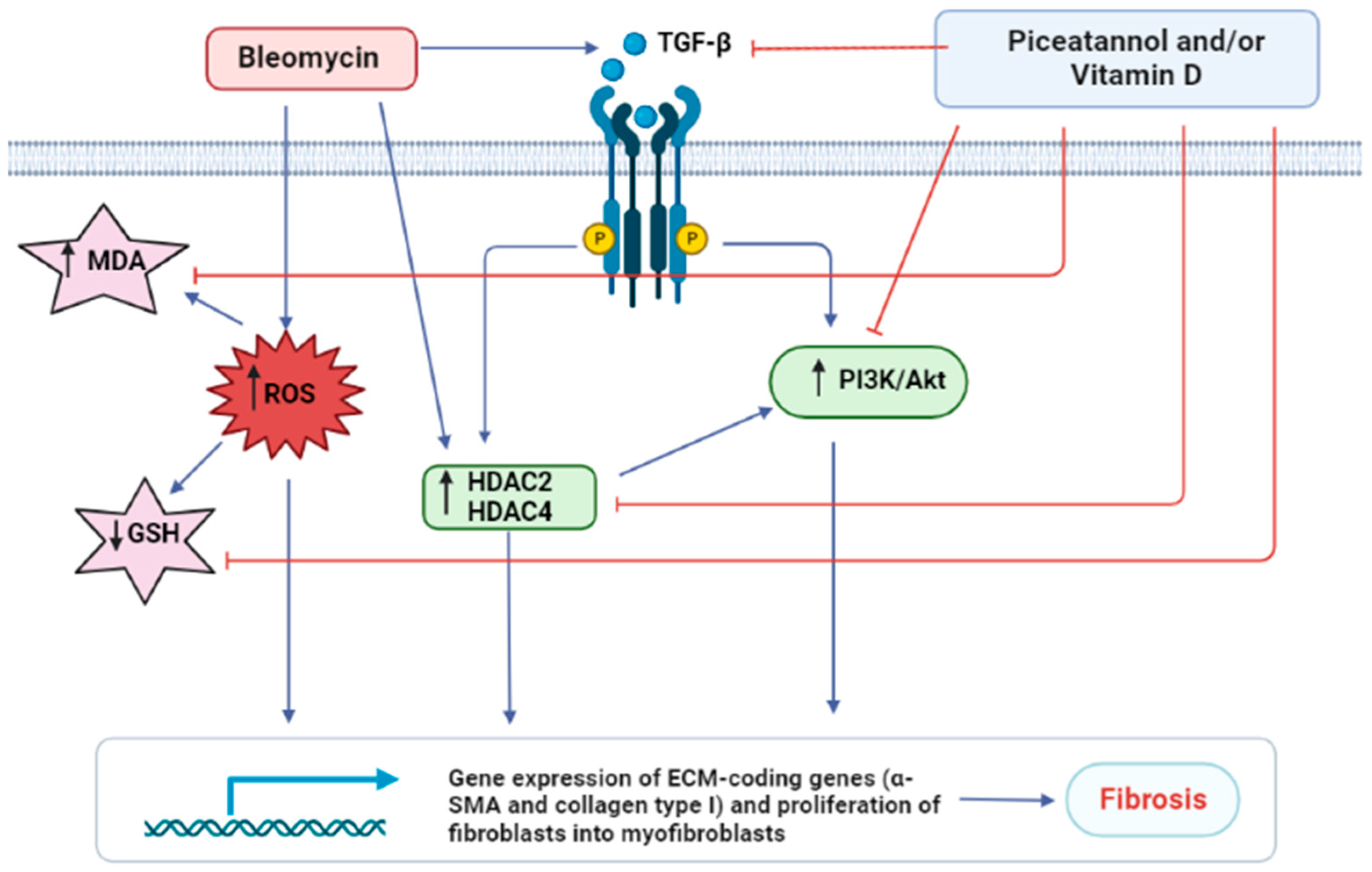
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sundarakrishnan, A.; Chen, Y.; Black, L.D.; Aldridge, B.B.; Kaplan, D.L. Engineered cell and tissue models of pulmonary fibrosis. Adv. Drug Deliv. Rev. 2018, 129, 78–94. [Google Scholar] [CrossRef] [PubMed]
- Yim, J.; Lim, H.H.; Kwon, Y. COVID-19 and pulmonary fibrosis: Therapeutics in clinical trials, repurposing, and potential development. Arch. Pharmacal. Res. 2021, 44, 499–513. [Google Scholar] [CrossRef] [PubMed]
- Kasthuri, R.S.; Glover, S.L.; Boles, J.; Mackman, N. Tissue factor and tissue factor pathway inhibitor as key regulators of global hemostasis: Measurement of their levels in coagulation assays. Semin. Thromb. Hemost. 2010, 36, 764–771. [Google Scholar] [CrossRef]
- Khan, S.; Ahirwar, K.; Jena, G. Anti-fibrotic effects of valproic acid: Role of HDAC inhibition and associated mechanisms. Epigenomics 2016, 8, 1087–1101. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.M.; Nepali, K.; Liou, J.P. Idiopathic Pulmonary Fibrosis: Current Status, Recent Progress, and Emerging Targets. J. Med. Chem. 2017, 60, 527–553. [Google Scholar] [CrossRef]
- Wynn, T.A. Integrating mechanisms of pulmonary fibrosis. J. Exp. Med. 2011, 208, 1339–1350. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.Y.; Piao, Z.H.; Jin, L.; Kim, J.H.; Kim, G.R.; Ryu, Y.; Lin, M.Q.; Kim, H.S.; Kee, H.J.; Jeong, M.H. Piceatannol Attenuates Renal Fibrosis Induced by Unilateral Ureteral Obstruction via Downregulation of Histone Deacetylase 4/5 or p38-MAPK Signaling. PLoS ONE 2016, 11, e0167340. [Google Scholar] [CrossRef]
- Lyu, X.; Hu, M.; Peng, J.; Zhang, X.; Sanders, Y.Y. HDAC inhibitors as antifibrotic drugs in cardiac and pulmonary fibrosis. Ther. Adv. Chronic Dis. 2019, 10, 2040622319862697. [Google Scholar] [CrossRef]
- Pang, M.; Zhuang, S. Histone Deacetylase: A Potential Therapeutic Target for Fibrotic Disorders. J. Pharmacol. Exp. Ther. 2010, 335, 266–272. [Google Scholar] [CrossRef]
- Gong, P.; Wang, Y.; Jing, Y. Apoptosis induction by histone deacetylase inhibitors in cancer cells: Role of Ku70. Int. J. Mol. Sci. 2019, 20, 1601. [Google Scholar] [CrossRef]
- Hung, W.-L.; Hsiao, Y.-T.; Chiou, Y.-S.; Nagabhushanam, K.; Ho, C.-T.; Pan, M.-H. Hepatoprotective effect of piceatannol against carbon tetrachloride-induced liver fibrosis in mice. Food Funct. 2021, 12, 11229–11240. [Google Scholar] [CrossRef] [PubMed]
- Kukreja, A.; Wadhwa, N.; Tiwari, A. Therapeutic role of resveratrol and piceatannol in disease prevention. J. Blood Disord. Transfus. 2014, 5, 9. [Google Scholar] [CrossRef]
- Cordova-Gomez, M.; Galano, A.; Alvarez-Idaboy, J.R. Piceatannol, a better peroxyl radical scavenger than resveratrol. RSC Adv. 2013, 3, 20209–20218. [Google Scholar] [CrossRef]
- Seyed, M.A.; Jantan, I.; Bukhari, S.N.A.; Vijayaraghavan, K. A comprehensive review on the chemotherapeutic potential of piceatannol for cancer treatment, with mechanistic insights. J. Agric. Food Chem. 2016, 64, 725–737. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.; Feng, Z.; Tong, M.; Xu, J.; Zheng, G.; Shen, L.; Shang, P.; Zhang, Y.; Liu, H. Piceatannol inhibits the IL-1β-induced inflammatory response in human osteoarthritic chondrocytes and ameliorates osteoarthritis in mice by activating Nrf2. Food Funct. 2017, 8, 3926–3937. [Google Scholar] [CrossRef]
- Ma, D.; Peng, L. Vitamin D and pulmonary fibrosis: A review of molecular mechanisms. Int. J. Clin. Exp. Pathol. 2019, 12, 3171. [Google Scholar]
- Hansdottir, S.; Monick, M.M. Vitamin D Effects on Lung Immunity and Respiratory Diseases. In Vitamins & Hormones; Elsevier: Amsterdam, The Netherlands, 2011; pp. 217–237. [Google Scholar]
- Papagni, R.; Pellegrino, C.; Di Gennaro, F.; Patti, G.; Ricciardi, A.; Novara, R.; Cotugno, S.; Musso, M.; Guido, G.; Ronga, L. Impact of vitamin D in prophylaxis and treatment in tuberculosis patients. Int. J. Mol. Sci. 2022, 23, 3860. [Google Scholar] [CrossRef]
- Patti, G.; Pellegrino, C.; Ricciardi, A.; Novara, R.; Cotugno, S.; Papagni, R.; Guido, G.; Totaro, V.; De Iaco, G.; Romanelli, F. Potential role of vitamins A, B, C, D and E in TB treatment and prevention: A narrative review. Antibiotics 2021, 10, 1354. [Google Scholar] [CrossRef]
- Hijona, E.; Aguirre, L.; Pérez-Matute, P.; Villanueva-Millán, M.; Mosqueda-Solis, A.; Hasnaoui, M.; Nepveu, F.; Senard, J.; Bujanda, L.; Aldamiz-Echevarría, L. Limited beneficial effects of piceatannol supplementation on obesity complications in the obese Zucker rat: Gut microbiota, metabolic, endocrine, and cardiac aspects. J. Physiol. Biochem. 2016, 72, 567–582. [Google Scholar] [CrossRef]
- Llarena, M.; Andrade, F.; Hasnaoui, M.; Portillo, M.P.; Pérez-Matute, P.; Arbones-Mainar, J.M.; Hijona, E.; Villanueva-Millán, M.J.; Aguirre, L.; Carpéné, C. Potential renoprotective effects of piceatannol in ameliorating the early-stage nephropathy associated with obesity in obese Zucker rats. J. Physiol. Biochem. 2016, 72, 555–566. [Google Scholar] [CrossRef]
- Saleh, M.A.; Antar, S.A.; Hazem, R.M.; El-Azab, M.F. Pirfenidone and vitamin D ameliorate cardiac fibrosis induced by doxorubicin in Ehrlich ascites carcinoma bearing mice: Modulation of monocyte chemoattractant protein-1 and Jun N-terminal kinase-1 pathways. Pharmaceuticals 2020, 13, 348. [Google Scholar] [CrossRef] [PubMed]
- Temsamani, H.; Krisa, S.; Decossas-Mendoza, M.; Lambert, O.; Mérillon, J.-M.; Richard, T. Piceatannol and other wine stilbenes: A pool of inhibitors against α-synuclein aggregation and cytotoxicity. Nutrients 2016, 8, 367. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, T.; Yang, P.-C. Western blot: Technique, theory, and trouble shooting. N. Am. J. Med. Sci. 2012, 4, 429–434. [Google Scholar] [PubMed]
- Elaidy, S.M.; Essawy, S.S. The antifibrotic effects of alveolar macrophages 5-HT2C receptors blockade on bleomycin-induced pulmonary fibrosis in rats. Pharmacol. Rep. 2016, 68, 1244–1253. [Google Scholar] [CrossRef] [PubMed]
- El-Gamal, M.A.; Zaitone, S.A.; Moustafa, Y.M. Role of irbesartan in protection against pulmonary toxicity induced by bleomycin in rats. IOSR J. Pharm. 2013, 3, 38–47. [Google Scholar] [CrossRef]
- Meyerholz, D.K.; Beck, A.P. Histopathologic Evaluation and Scoring of Viral Lung Infection. Methods Mol. Biol. 2020, 2099, 205–220. [Google Scholar] [PubMed]
- Liu, B.; Li, R.; Zhang, J.; Meng, C.; Zhang, J.; Song, X.; Lv, C. MicroRNA-708-3p as a potential therapeutic target via the ADAM17-GATA/STAT3 axis in idiopathic pulmonary fibrosis. Exp. Mol. Med. 2018, 50, e465. [Google Scholar] [CrossRef]
- George, P.M.; Wells, A.U.; Jenkins, R.G. Pulmonary fibrosis and COVID-19: The potential role for antifibrotic therapy. Lancet Respir. Med. 2020, 8, 807–815. [Google Scholar] [CrossRef]
- Alsayed, H.A.; Mohammad, H.M.F.; Khalil, C.M.; El-Kherbetawy, M.K.; Elaidy, S.M. Autophagy modulation by irbesartan mitigates the pulmonary fibrotic alterations in bleomycin challenged rats: Comparative study with rapamycin. Life Sci. 2022, 303, 120662. [Google Scholar] [CrossRef]
- Leask, A.; Abraham, D.J. TGF-β signaling and the fibrotic response. FASEB J. 2004, 18, 816–827. [Google Scholar] [CrossRef]
- Roberts, A.B.; Piek, E.; Böttinger, E.P.; Ashcroft, G.; Mitchell, J.B.; Flanders, K.C. Is Smad3 a Major Player in Signal Transduction Pathways Leading to Fibrogenesis? Chest 2001, 120, S43–S47. [Google Scholar] [CrossRef]
- Yue, X.; Shan, B.; Lasky, J.A. TGF-β: Titan of Lung Fibrogenesis. Curr. Enzym. Inhib. 2010, 6, 24187529. [Google Scholar] [CrossRef]
- Korfei, M.; Mahavadi, P.; Guenther, A. Targeting Histone Deacetylases in Idiopathic Pulmonary Fibrosis: A Future Therapeutic Option. Cells 2022, 11, 1626. [Google Scholar] [CrossRef]
- Lu, Y.; Azad, N.; Wang, L.; Iyer, A.K.; Castranova, V.; Jiang, B.H.; Rojanasakul, Y. Phosphatidylinositol-3-kinase/akt regulates bleomycin-induced fibroblast proliferation and collagen production. Am. J. Respir. Cell Mol. Biol. 2010, 42, 432–441. [Google Scholar] [CrossRef]
- Spassov, S.G.; Donus, R.; Ihle, P.M.; Engelstaedter, H.; Hoetzel, A.; Faller, S. Hydrogen Sulfide Prevents Formation of Reactive Oxygen Species through PI3K/Akt Signaling and Limits Ventilator-Induced Lung Injury. Oxidative Med. Cell. Longev. 2017, 2017, 3715037. [Google Scholar] [CrossRef]
- Jiménez-Uribe, A.P.; Gómez-Sierra, T.; Aparicio-Trejo, O.E.; Orozco-Ibarra, M.; Pedraza-Chaverri, J. Backstage players of fibrosis: NOX4, mTOR, HDAC, and S1P; companions of TGF-β. Cell Signal. 2021, 87, 110123. [Google Scholar] [CrossRef]
- Sivashanmugam, K.; Kandasamy, M.; Subbiah, R.; Ravikumar, V. Repurposing of histone deacetylase inhibitors: A promising strategy to combat pulmonary fibrosis promoted by TGF-β signaling in COVID-19 survivors. Life Sci. 2021, 266, 118883. [Google Scholar]
- Samah, M.; El-Aidy, A.E.-R.; Tawfik, M.K.; Ewais, M.M.S. Evaluation of the antifibrotic effect of fenofibrate and rosiglitazone on bleomycin-induced pulmonary fibrosis in rats. Eur. J. Pharmacol. 2012, 689, 186–193. [Google Scholar] [CrossRef]
- Zhu, W.; Ding, Q.; Wang, L.; Xu, G.; Diao, Y.; Qu, S.; Chen, S.; Shi, Y. Vitamin D3 alleviates pulmonary fibrosis by regulating the MAPK pathway via targeting PSAT1 expression in vivo and in vitro. Int. Immunopharmacol. 2021, 101, 108212. [Google Scholar] [CrossRef] [PubMed]
- Xiong, C.; Guan, Y.; Zhou, X.; Liu, L.; Zhuang, M.A.; Zhang, W.; Zhang, Y.; Masucci, M.V.; Bayliss, G.; Zhao, T.C.; et al. Selective inhibition of class IIa histone deacetylases alleviate renal fibrosis. FASEB J. 2019, 33, 8249–8262. [Google Scholar] [CrossRef] [PubMed]
- Sanders, Y.Y.; Hagood, J.S.; Liu, H.; Zhang, W.; Ambalavanan, N.; Thannickal, V.J. Histone deacetylase inhibition promotes fibroblast apoptosis and ameliorates pulmonary fibrosis in mice. Eur. Respir. J. 2014, 43, 1448–1458. [Google Scholar] [CrossRef] [PubMed]
- Ota, C.; Yamada, M.; Fujino, N.; Motohashi, H.; Tando, Y.; Takei, Y.; Suzuki, T.; Takahashi, T.; Kamata, S.; Makiguchi, T.; et al. Histone deacetylase inhibitor restores surfactant protein-C expression in alveolar-epithelial type II cells and attenuates bleomycin-induced pulmonary fibrosis in vivo. Exp. Lung Res. 2015, 41, 422–434. [Google Scholar] [CrossRef]
- Chen, L.; Alam, A.; Pac-Soo, A.; Chen, Q.; Shang, Y.; Zhao, H.; Yao, S.; Ma, D. Pretreatment with valproic acid alleviates pulmonary fibrosis through epithelial–mesenchymal transition inhibition in vitro and in vivo. Lab. Investig. 2021, 101, 1166–1175. [Google Scholar] [CrossRef] [PubMed]
- Williams, S.M.; Golden-Mason, L.; Ferguson, B.S.; Schuetze, K.B.; Cavasin, M.A.; Demos-Davies, K.; Yeager, M.E.; Stenmark, K.R.; McKinsey, T.A. Class I HDACs regulate angiotensin II-dependent cardiac fibrosis via fibroblasts and circulating fibrocytes. J. Mol. Cell. Cardiol. 2014, 67, 112–125. [Google Scholar] [CrossRef] [PubMed]
- Parbin, S.; Kar, S.; Shilpi, A.; Sengupta, D.; Deb, M.; Rath, S.K.; Patra, S.K. Histone Deacetylases: A Saga of Perturbed Acetylation Homeostasis in Cancer. J. Histochem. Cytochem. 2013, 62, 11–33. [Google Scholar] [CrossRef] [PubMed]
- Korfei, M.; Skwarna, S.; Henneke, I.; MacKenzie, B.; Klymenko, O.; Saito, S.; Ruppert, C.; von der Beck, D.; Mahavadi, P.; Klepetko, W.; et al. Aberrant expression and activity of histone deacetylases in sporadic idiopathic pulmonary fibrosis. Thorax 2015, 70, 1022–1032. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Zhang, Y.; Tu, T.; Schmull, S.; Han, Y.; Wang, W.; Li, H. Dual inhibition of HDAC and tyrosine kinase signaling pathways with CUDC-907 attenuates TGFβ1 induced lung and tumor fibrosis. Cell Death Dis. 2020, 11, 765. [Google Scholar] [CrossRef]
- Li, M.; Zheng, Y.; Yuan, H.; Liu, Y.; Wen, X. Effects of dynamic changes in histone acetylation and deacetylase activity on pulmonary fibrosis. Int. Immunopharmacol. 2017, 52, 272–280. [Google Scholar] [CrossRef]
- Guo, W.; Shan, B.; Klingsberg, R.C.; Qin, X.; Lasky, J.A. Abrogation of TGF-β1-induced fibroblast-myofibroblast differentiation by histone deacetylase inhibition. Am. J. Physiol. Lung Cell. Mol. Physiol. 2009, 297, L864–L870. [Google Scholar] [CrossRef]
- Khalil, W.; Xia, H.; Bodempudi, V.; Kahm, J.; Hergert, P.; Smith, K.; Peterson, M.; Parker, M.; Herrera, J.; Bitterman, P.B. Pathologic regulation of collagen I by an aberrant protein phosphatase 2A/histone deacetylase C4/microRNA-29 signal axis in idiopathic pulmonary fibrosis fibroblasts. Am. J. Respir. Cell Mol. Biol. 2015, 53, 391–399. [Google Scholar] [CrossRef]
- Krämer, O.H. HDAC2: A critical factor in health and disease. Trends Pharmacol. Sci. 2009, 30, 647–655. [Google Scholar] [CrossRef] [PubMed]
- Estornut, C.; Milara, J.; Bayarri, M.A.; Belhadj, N.; Cortijo, J. Targeting Oxidative Stress as a Therapeutic Approach for Idiopathic Pulmonary Fibrosis. Front. Pharmacol. 2021, 12, 794997. [Google Scholar] [CrossRef]
- Otoupalova, E.; Smith, S.; Cheng, G.; Thannickal, V.J. Oxidative Stress in Pulmonary Fibrosis. Compr. Physiol. 2020, 10, 509–547. [Google Scholar] [PubMed]
- Tsubouchi, K.; Araya, J.; Yoshida, M.; Sakamoto, T.; Koumura, T.; Minagawa, S.; Hara, H.; Hosaka, Y.; Ichikawa, A.; Saito, N.; et al. Involvement of GPx4-Regulated Lipid Peroxidation in Idiopathic Pulmonary Fibrosis Pathogenesis. J. Immunol. 2019, 203, 2076–2087. [Google Scholar] [CrossRef]
- Beeh, K.M.; Beier, J.; Haas, I.C.; Kornmann, O.; Micke, P.; Buhl, R. Glutathione deficiency of the lower respiratory tract in patients with idiopathic pulmonary fibrosis. Eur. Respir. J. 2002, 19, 1119–1123. [Google Scholar] [CrossRef] [PubMed]
- Della Latta, V.; Cecchettini, A.; Del Ry, S.; Morales, M.A. Bleomycin in the setting of lung fibrosis induction: From biological mechanisms to counteractions. Pharmacol. Res. 2015, 97, 122–130. [Google Scholar] [CrossRef]
- Piceatannol Mediated Modulation of Oxidative Stress and Regeneration in the Liver of Endotoxemic Mice. J. Med. Food 2019, 22, 594–601. [CrossRef]
- Da Costa, F.P.; Puty, B.; Nogueira, L.S.; Mitre, G.P.; Santos, S.M.d.; Teixeira, B.J.B.; Kataoka, M.S.d.S.; Martins, M.D.; Barboza, C.A.G.; Monteiro, M.C.; et al. Piceatannol Increases Antioxidant Defense and Reduces Cell Death in Human Periodontal Ligament Fibroblast under Oxidative Stress. Antioxidants 2020, 9, 16. [Google Scholar] [CrossRef]
- Yamini, P.; Ray, R.S.; Chopra, K. Vitamin D3 attenuates cognitive deficits and neuroinflammatory responses in ICV-STZ induced sporadic Alzheimer’s disease. Inflammopharmacology 2018, 26, 39–55. [Google Scholar] [CrossRef]
- Sheng, H.; Lin, G.; Zhao, S.; Li, W.; Zhang, Z.; Zhang, W.; Yun, L.; Yan, X.; Hu, H. Antifibrotic Mechanism of Piceatannol in Bleomycin-Induced Pulmonary Fibrosis in Mice. Front. Pharmacol. 2022, 13, 35747752. [Google Scholar] [CrossRef]
- Tieyuan, Z.; Ying, Z.; Xinghua, Z.; Huimin, W.; Huagang, L. Piceatannol-mediated JAK2/STAT3 signaling pathway inhibition contributes to the alleviation of oxidative injury and collagen synthesis during pulmonary fibrosis. Int. Immunopharmacol. 2022, 111, 109107. [Google Scholar] [CrossRef] [PubMed]
- Li, S.-R.; Tan, Z.-X.; Chen, Y.-H.; Hu, B.; Zhang, C.; Wang, H.; Zhao, H.; Xu, D.-X. Vitamin D deficiency exacerbates bleomycin-induced pulmonary fibrosis partially through aggravating TGF-β/Smad2/3-mediated epithelial-mesenchymal transition. Respir. Res. 2019, 20, 266. [Google Scholar] [CrossRef] [PubMed]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ezz Eldeen, N.; Moustafa, Y.M.; Alwaili, M.A.; Alrehaili, A.A.; Khodeer, D.M. Synergistic Power of Piceatannol and/or Vitamin D in Bleomycin-Induced Pulmonary Fibrosis In Vivo: A Preliminary Study. Biomedicines 2023, 11, 2647. https://doi.org/10.3390/biomedicines11102647
Ezz Eldeen N, Moustafa YM, Alwaili MA, Alrehaili AA, Khodeer DM. Synergistic Power of Piceatannol and/or Vitamin D in Bleomycin-Induced Pulmonary Fibrosis In Vivo: A Preliminary Study. Biomedicines. 2023; 11(10):2647. https://doi.org/10.3390/biomedicines11102647
Chicago/Turabian StyleEzz Eldeen, Nehal, Yasser M. Moustafa, Maha Abdullah Alwaili, Amani A. Alrehaili, and Dina M. Khodeer. 2023. "Synergistic Power of Piceatannol and/or Vitamin D in Bleomycin-Induced Pulmonary Fibrosis In Vivo: A Preliminary Study" Biomedicines 11, no. 10: 2647. https://doi.org/10.3390/biomedicines11102647
APA StyleEzz Eldeen, N., Moustafa, Y. M., Alwaili, M. A., Alrehaili, A. A., & Khodeer, D. M. (2023). Synergistic Power of Piceatannol and/or Vitamin D in Bleomycin-Induced Pulmonary Fibrosis In Vivo: A Preliminary Study. Biomedicines, 11(10), 2647. https://doi.org/10.3390/biomedicines11102647