

Antimicrobial and Antiviral Compounds of Phlomis viscosa Poiret

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Plant Material
2.2. Identification of Plant Extract Components
2.3. Materials and Bacterial Strains
2.4. Biofilm Formation Estimation
2.5. Animal Cells and Viruses
2.6. Antiviral Activity
2.7. Statistical Analysis
3. Results and Discussion
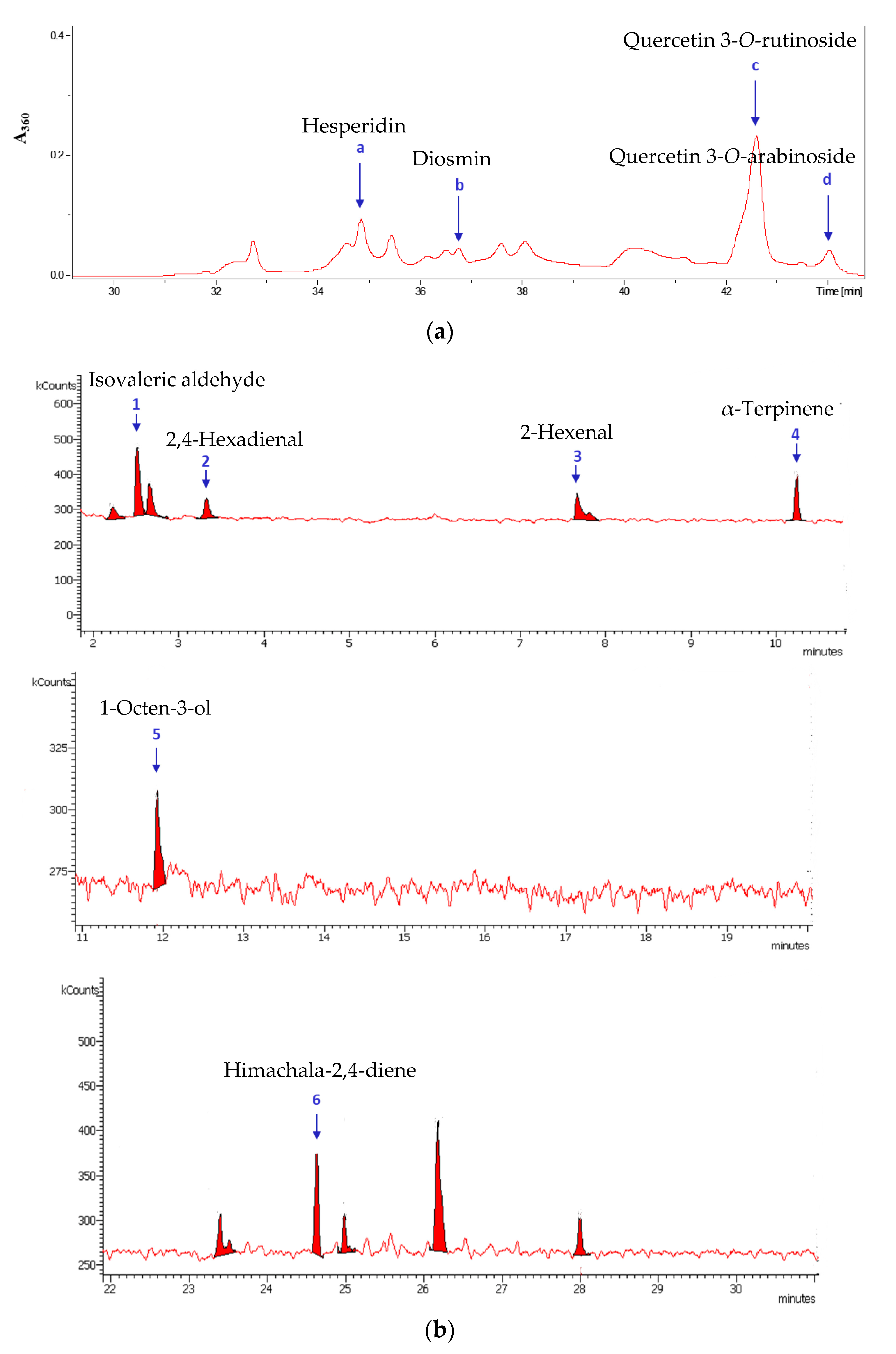
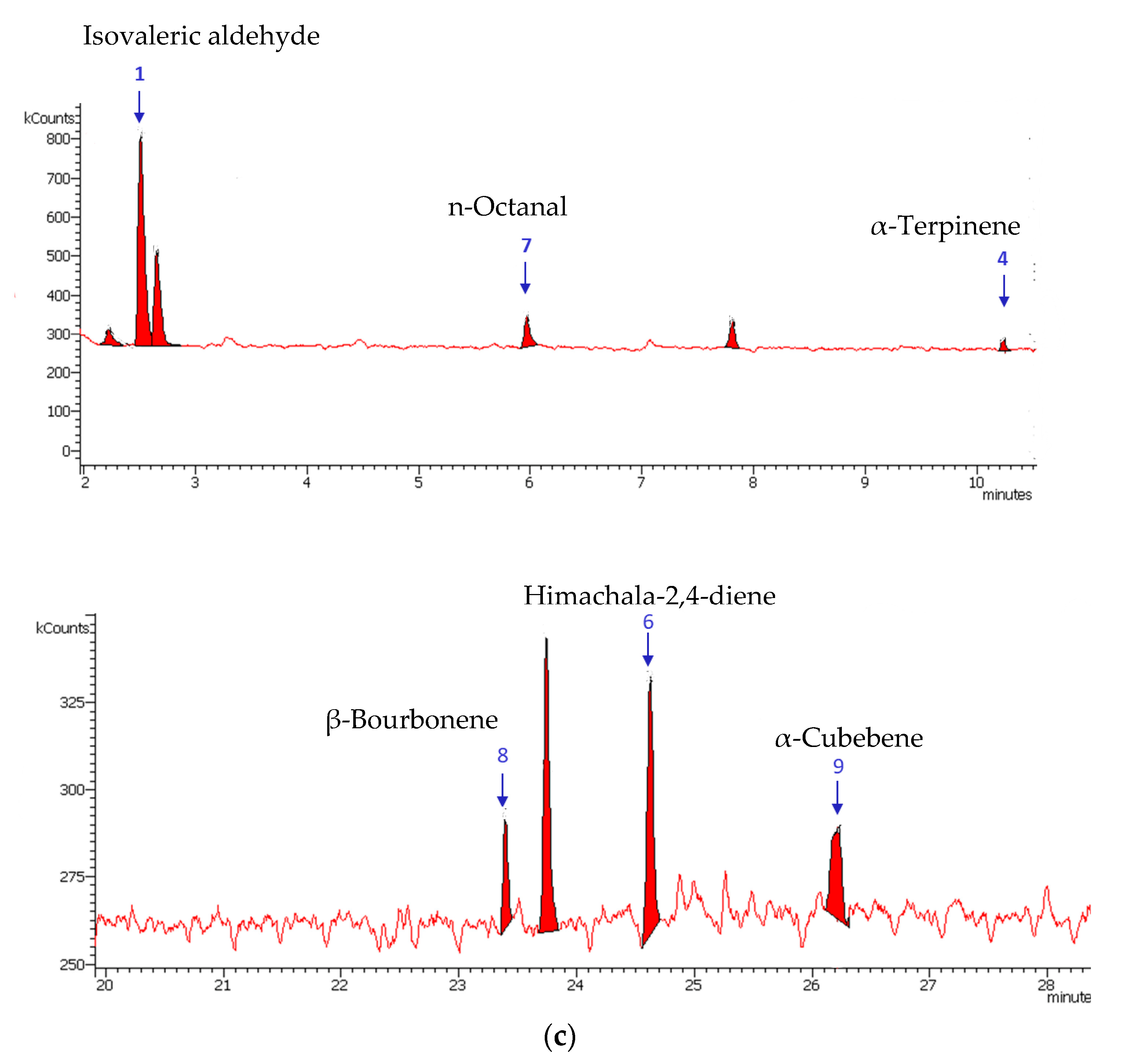
3.1. Identification of Plant Extract Components
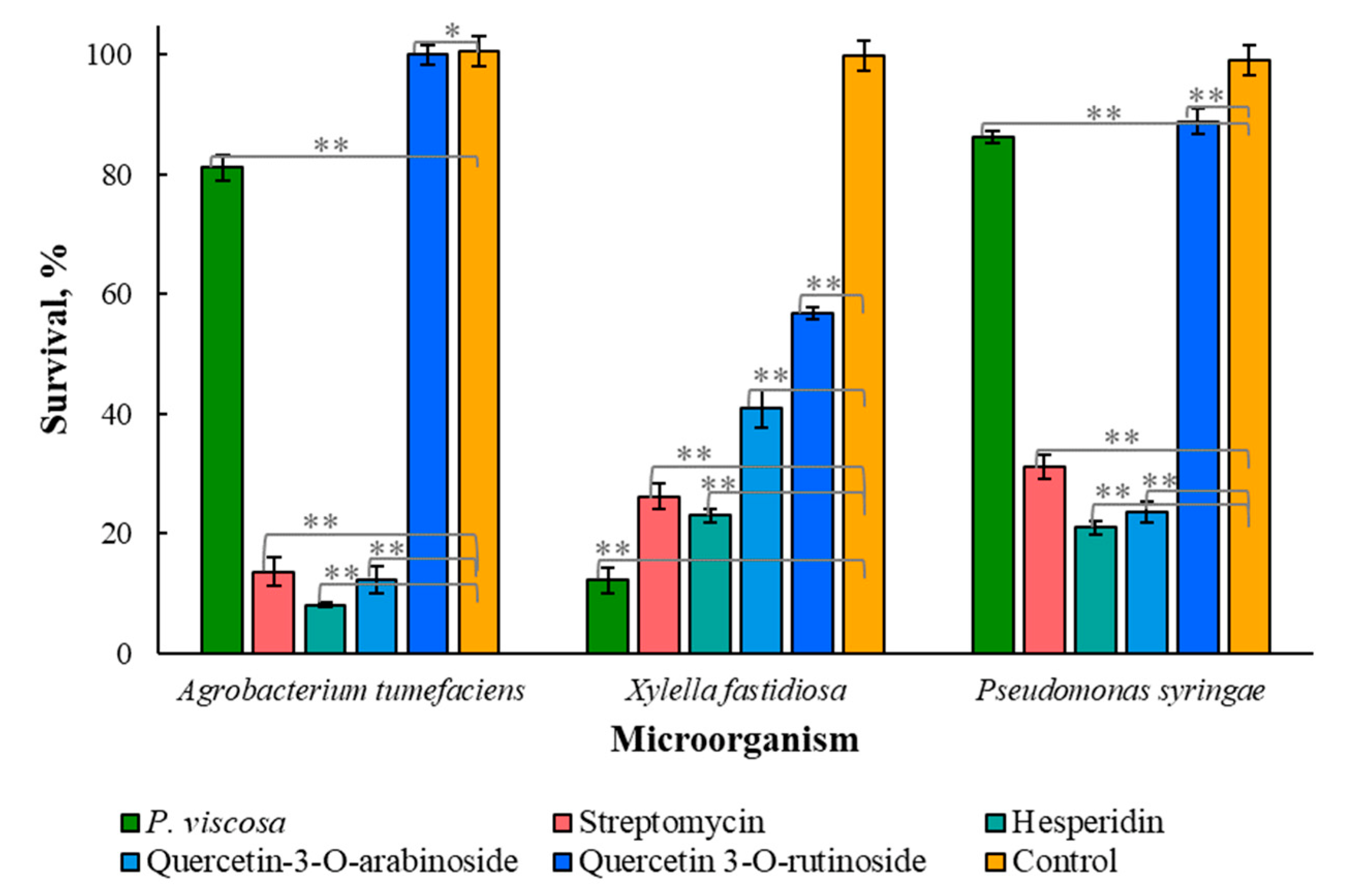
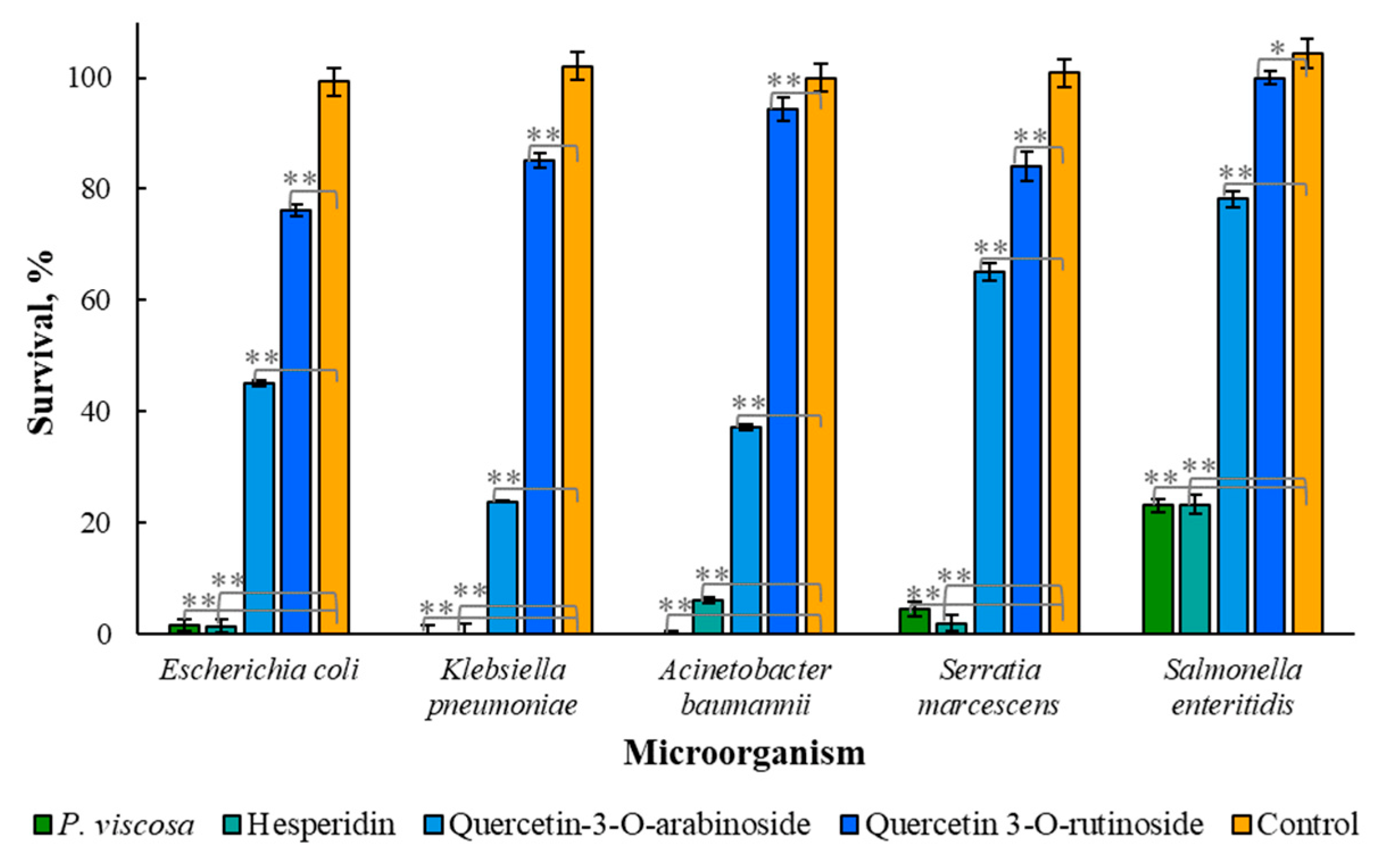
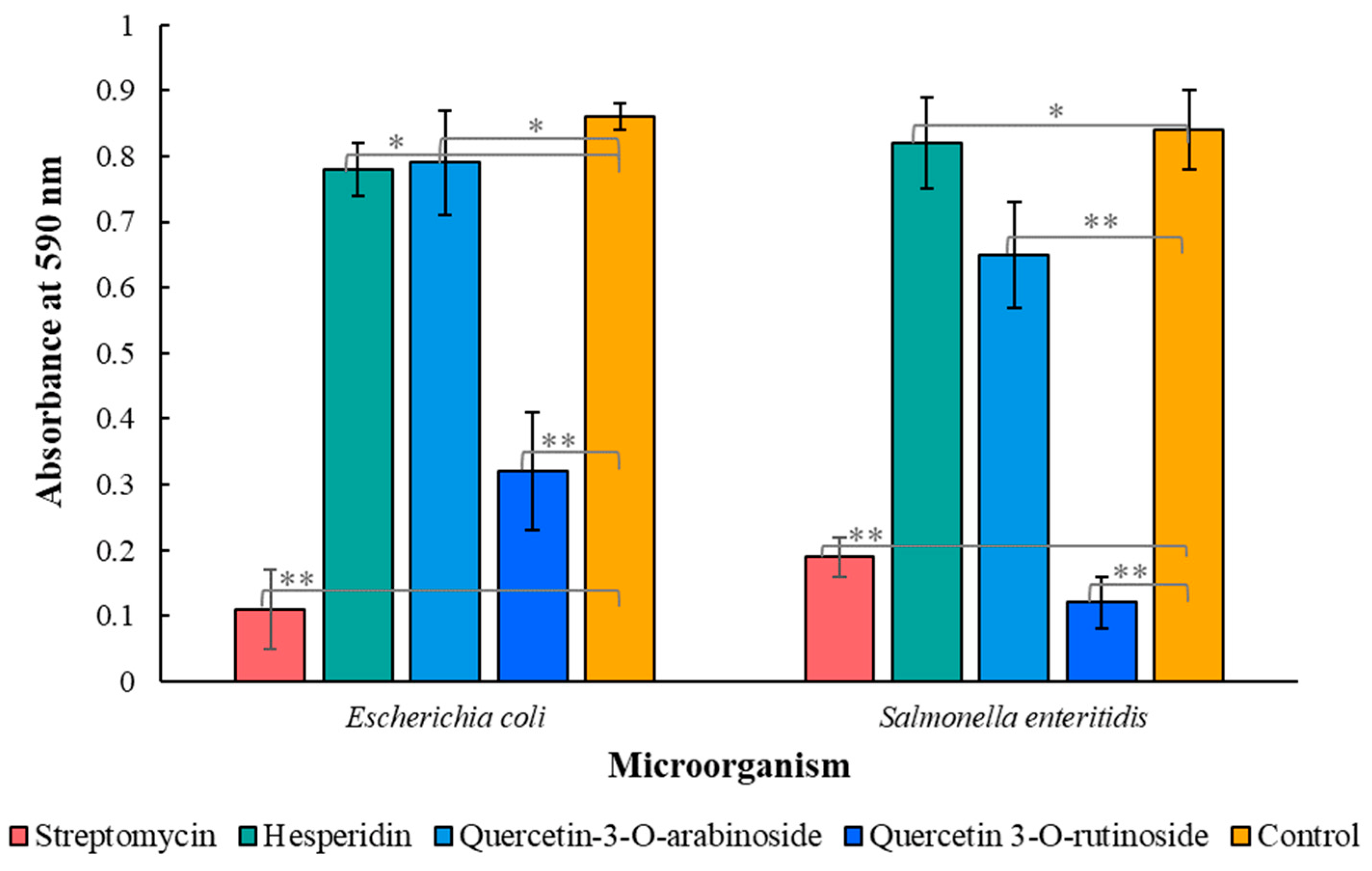
3.2. Antimicrobial Properties of Phytochemicals from P. viscosa
3.3. Toxicity of Phytochemicals from P. viscosa against Eukaryotic Cells
3.4. Antiviral Activity of Phytochemicals from P. viscosa
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mathiesen, C.; Scheen, A.-C.; Lindqvist, C. Phylogeny and biogeography of the lamioid genus Phlomis (Lamiaceae). Kew Bull. 2011, 66, 83–99. [Google Scholar] [CrossRef]
- Limem-Ben Amor, I.; Boubaker, J.; Ben Sgaier, M.; Skandrani, I.; Bhouri, W.; Neffati, A.; Kilani, S.; Bouhlel, I.; Ghedira, K.; Chekir-Ghedira, L. Phytochemistry and biological activities of Phlomis species. J. Ethnopharmacol. 2009, 125, 183–202. [Google Scholar] [CrossRef] [PubMed]
- Yarmolinsky, L.; Budovsky, A.; Ben-Shabat, S.; Khalfin, B.; Gorelick, J.; Bishitz, Y.; Miloslavski, R.; Yarmolinsky, L. Recent Updates on the Phytochemistry and Pharmacological Properties of Phlomis viscosa Poiret. Rejuvenation Res. 2019, 22, 282–288. [Google Scholar] [CrossRef] [PubMed]
- Yarmolinsky, L.L.; Budovsky, A.; Yarmolinsky, L.L.; Khalfin, B.; Glukhman, V.; Ben-Shabat, S. Effect of bioactive phytochemicals from Phlomis viscosa poiret on wound healing. Plants 2019, 8, 609. [Google Scholar] [CrossRef]
- Sengupta, S.; Chattopadhyay, M.K.; Grossart, H.-P. The multifaceted roles of antibiotics and antibiotic resistance in nature. Front. Microbiol. 2013, 4, 47. [Google Scholar] [CrossRef]
- Hochma, E.; Yarmolinsky, L.; Khalfin, B.; Nisnevitch, M.; Ben-Shabat, S.; Nakonechny, F. Antimicrobial Effect of Phytochemicals from Edible Plants. Processes 2021, 9, 2089. [Google Scholar] [CrossRef]
- Mulat, M.; Pandita, A.; Khan, F. Medicinal Plant Compounds for Combating the Multi-drug Resistant Pathogenic Bacteria: A Review. Curr. Pharm. Biotechnol. 2019, 20, 183–196. [Google Scholar] [CrossRef]
- Friesen, M.L. Social Evolution and Cheating in Plant Pathogens. Annu. Rev. Phytopathol. 2020, 58, 55–75. [Google Scholar] [CrossRef]
- Asghari, S.; Harighi, B.; Ashengroph, M.; Clement, C.; Aziz, A.; Esmaeel, Q.; Ait Barka, E. Induction of systemic resistance to Agrobacterium tumefaciens by endophytic bacteria in grapevine. Plant Pathol. 2020, 69, 827–837. [Google Scholar] [CrossRef]
- Mansfield, J.; Genin, S.; Magori, S.; Citovsky, V.; Sriariyanum, M.; Ronald, P.; Dow, M.; Verdier, V.; Beer, S.V.; Machado, M.A.; et al. Top 10 plant pathogenic bacteria in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 614–629. [Google Scholar] [CrossRef] [Green Version]
- Whitelaw-Weckert, M.A.; Whitelaw, E.S.; Rogiers, S.Y.; Quirk, L.; Clark, A.C.; Huang, C.X. Bacterial inflorescence rot of grapevine caused by Pseudomonas syringae pv. syringae. Plant Pathol. 2011, 60, 325–337. [Google Scholar] [CrossRef]
- Orton, T.J. Chapter 17—Breeding Methods for Outcrossing Plant Species: III. Asexual Propagation. In Horticultural Plant Breeding; Orton, T.J., Ed.; Academic Press, Imprint of Elsevier: London, UK, 2020; pp. 309–326. ISBN 978-0-12-815396-3. [Google Scholar]
- Yarmolinsky, L.; Bronstein, M.; Gorelick, J. Review: Inhibition of bacterial quorum sensing by plant extracts. Isr. J. Plant Sci. 2015, 62, 294–297. [Google Scholar] [CrossRef]
- Gao, Z.; Zhong, W.; Chen, K.; Tang, P.; Guo, J. Chemical composition and anti-biofilm activity of essential oil from Citrus medica L. var. sarcodactylis Swingle against Listeria monocytogenes. Ind. Crops Prod. 2020, 144, 112036. [Google Scholar] [CrossRef]
- Field, H.J.; Biswas, S.; Mohammad, I.T. Herpesvirus latency and therapy--from a veterinary perspective. Antivir. Res. 2006, 71, 127–133. [Google Scholar] [CrossRef]
- Field, A.K.; Biron, K.K. “The end of innocence” revisited: Resistance of herpesviruses to antiviral drugs. Clin. Microbiol. Rev. 1994, 7, 1–13. [Google Scholar] [CrossRef]
- Skevaki, C.L.; Galani, I.E.; Pararas, M.V.; Giannopoulou, K.P.; Tsakris, A. Treatment of viral conjunctivitis with antiviral drugs. Drugs 2011, 71, 331–347. [Google Scholar] [CrossRef]
- Ben-Shabat, S.; Yarmolinsky, L.; Porat, D.; Dahan, A. Antiviral effect of phytochemicals from medicinal plants: Applications and drug delivery strategies. Drug Deliv. Transl. Res. 2020, 10, 354–367. [Google Scholar] [CrossRef]
- Dekić, M.S.; Radulović, N.S.; Stojanović, N.M.; Randjelović, P.J.; Stojanović-Radić, Z.Z.; Najman, S.; Stojanović, S. Spasmolytic, antimicrobial and cytotoxic activities of 5-phenylpentyl isothiocyanate, a new glucosinolate autolysis product from horseradish (Armoracia rusticana P. Gaertn., B. Mey. & Scherb., Brassicaceae). Food Chem. 2017, 232, 329–339. [Google Scholar] [CrossRef]
- Staszowska-Karkut, M.; Materska, M. Phenolic Composition, Mineral Content, and Beneficial Bioactivities of Leaf Extracts from Black Currant (Ribes nigrum L.), Raspberry (Rubus idaeus), and Aronia (Aronia melanocarpa). Nutrients 2020, 12, 463. [Google Scholar] [CrossRef]
- Calis, I.; Kirmizibekmez, H.; Beutler, J.A.; Donmez, A.A.; Yalcin, F.N.; Kilic, E.; Ozalp, M.; Ruedi, P.; Tasdemir, D. Secondary metabolites of Phlomis viscosa and their biological activities. Turk. J. Chem. 2005, 29, 71–81. [Google Scholar]
- Khalil, A.; Dababneh, B.F. Inhibition of Phytopathogenic Fungi by Extracts from Medicinal Plants in Jordan. J. Biol. Sci. 2007, 7, 579–581. [Google Scholar] [CrossRef] [Green Version]
- Yarmolinsky, L.; Huleihel, M.; Zaccai, M.; Ben-Shabat, S. Potent antiviral flavone glycosides from Ficus benjamina leaves. Fitoterapia 2012, 83, 362–367. [Google Scholar] [CrossRef] [PubMed]
- Vasavi, H.S.; Arun, A.B.; Rekha, P.D. Anti-quorum sensing activity of Psidium guajava L. flavonoids against Chromobacterium violaceum and Pseudomonas aeruginosa PAO1. Microbiol. Immunol. 2014, 58, 286–293. [Google Scholar] [CrossRef] [PubMed]
- Yarmolinsky, L.; Zaccai, M.; Ben-Shabat, S.; Mills, D.; Huleihel, M. Antiviral activity of ethanol extracts of Ficus binjamina and Lilium candidum in vitro. New Biotechnol. 2009, 26, 307–313. [Google Scholar] [CrossRef]
- Joo, K.-M.; Kim, S.; Koo, Y.J.; Lee, M.; Lee, S.-H.; Choi, D.; Lim, K.-M. Development and validation of UPLC method for WST-1 cell viability assay and its application to MCTT HCETM eye irritation test for colorful substances. Toxicol. In Vitro Int. J. Publ. Assoc. BIBRA 2019, 60, 412–419. [Google Scholar] [CrossRef]
- Berrani, A.; Marmouzi, I.; Bouyahya, A.; Kharbach, M.; El Hamdani, M.; El Jemli, M.; Lrhorfi, A.; Zouarhi, M.; Faouzi, M.E.A.; Bengueddour, R. Phenolic Compound Analysis and Pharmacological Screening of Vitex agnus-castus Functional Parts. BioMed Res. Int. 2021, 2021, 6695311. [Google Scholar] [CrossRef]
- Sevindik, M.; Rasul, A.; Hussain, G.; Anwar, H.; Zahoor, M.K.; Sarfraz, I.; Kamran, K.S.; Akgul, H.; Akata, I.; Selamoglu, Z. Determination of anti-oxidative, anti-microbial activity and heavy metal contents of Leucoagaricus leucothites. Pak. J. Pharm. Sci. 2018, 31, 2163–2168. [Google Scholar]
- Cohen, J.I. 137—Introduction to Herpesviridae. In Blaser Douglas, and Bennett’s Principles and Practice of Infectious Diseases, 8th ed.; Bennett, J.E., Dolin, R., Martin, J., Eds.; W.B. Saunders: Philadelphia, PA, USA, 2015; p. 1707–1712.e1. ISBN 978-1-4557-4801-3. [Google Scholar]
- Liu, Z.-H.; Niu, F.-J.; Xie, Y.-X.; Xie, S.-M.; Liu, Y.-N.; Yang, Y.-Y.; Zhou, C.-Z.; Wan, X.-H. A review: Natural polysaccharides from medicinal plants and microorganisms and their anti-herpetic mechanism. Biomed. Pharmacother. 2020, 129, 110469. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Compound | Molecular Formula | Retention Time, min | Probability of Identification (%) | Origin |
|---|---|---|---|---|---|
| a | Hesperidin | C28H34O15 | 34.85 | 95.6 | Leaves and Flowers |
| b | Diosmin | C28H32O15 | 36.76 | 98.7 | Leaves |
| c | Quercetin 3-O-rutinoside | C27H30O16 | 42.67 | 96.4 | Leaves |
| d | Quercetin 3-O-arabinoside | C20H18O11 | 44.06 | 95.9 | Leaves |
| No | Compound | Molecular Formula | Retention Time, min | Probability of Identification (%) | Origin |
|---|---|---|---|---|---|
| 1 | Isovaleric aldehyde | C5H10O | 2.50 | 84 | Leaves |
| 2 | 2,4-Hexadienal | C6H8O | 3.33 | 83.5 | Leaves |
| 3 | 2-Hexenal | C6H10O | 7.66 | 87.2 | Leaves |
| 4 | α-Terpinene | C10H16 | 10.24 | 86.5 | Leaves and Flowers |
| 5 | 1-Octen-3-ol | C8H16O | 11.92 | 87.3 | Leaves |
| 6 | Himachala-2,4-diene | C15H24 | 24.63 | 85.3 | Leaves and Flowers |
| 7 | n-Octanal | C8H16O | 5.97 | 87.3 | Flowers |
| 8 | β-Bourbonene | C15H24 | 23.74 | 89.2 | Flowers |
| 9 | α-Cubebene | C15H24 | 26.21 | 87.3 | Leaves and Flowers |
| Extract or Compound | Concentration (µg/mL) | Cell Count (% of Control) | Viability (% of Control) |
|---|---|---|---|
| P. viscosa | 1 | 101.5 ± 1.4 | 102.8 ± 2.5 |
| 10 | 100.7 ± 1.4 | 100.5 ± 0.9 | |
| 100 | 99.0 ± 1.9 | 100.9 ± 2.3 | |
| 500 | 100.9 ± 1.9 | 99.9 ± 4.1 | |
| Diosmin | 1 | 101.9 ± 1.6 | 101.6 ± 1.8 |
| 10 | 101.2 ± 1.3 | 102.1 ± 2.2 | |
| 100 | 100.1 ± 1.1 | 100.7 ± 1.3 | |
| 200 | 72.2 ± 3.1 | 70.9 ± 2.6 | |
| Hesperidin | 1 | 101.5 ± 1.4 | 100.5 ± 1.6 |
| 10 | 98.9 ± 1.8 | 99.2 ± 1.5 | |
| 50 | 87.1 ± 3.1 | 88.0 ± 2.2 | |
| 100 | 75.3 ± 3.1 | 74.8 ± 2.5 | |
| 200 | 71.4 ± 1.1 | 71.9 ± 1.6 | |
| Quercetin 3-O-arabinoside | 1 | 102.2 ± 1.5 | 102.7 ± 1.8 |
| 10 | 97.3 ± 1.2 | 97.6 ± 1.4 | |
| 50 | 94.2 ± 2.5 | 94.7 ± 2.3 | |
| 100 | 89.7 ± 1.2 | 89.2 ± 1.4 | |
| 200 | 84.8 ± 1.1 | 84.1 ± 2.0 | |
| Quercetin 3-O-rutinoside | 1 | 103.2 ± 1.9 | 103.6 ± 1.2 |
| 10 | 102.2 ± 1.2 | 101.9 ± 1.7 | |
| 100 | 100.4 ± 1.5 | 100.7 ± 1.7 | |
| 200 | 100.7 ± 1.3 | 100.4 ± 2.0 | |
| 1000 | 100.1 ± 2.2 | 100.2 ± 1.8 |
| Compound | Fraction of Plaque Forming Units | ||
|---|---|---|---|
| HSV-1 | HSV-1, Mutant | VZV | |
| Untreated control | 1 | 1 | 1 |
| Diosmin | 1 | 1 | 0 |
| Hesperidin | 1 | 1 | 0 |
| Quercetin 3-O-arabinoside | 0.35 ± 0.05 | 0.36 ± 0.03 | 0.11 ± 0.07 |
| Quercetin 3-O-rutinoside | 0.61 ± 0.08 | 1 | 1 |
| Acyclovir | 0.12 ± 0.02 | 1 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yarmolinsky, L.; Nakonechny, F.; Budovsky, A.; Zeigerman, H.; Khalfin, B.; Sharon, E.; Yarmolinsky, L.; Ben-Shabat, S.; Nisnevitch, M. Antimicrobial and Antiviral Compounds of Phlomis viscosa Poiret. Biomedicines 2023, 11, 441. https://doi.org/10.3390/biomedicines11020441
Yarmolinsky L, Nakonechny F, Budovsky A, Zeigerman H, Khalfin B, Sharon E, Yarmolinsky L, Ben-Shabat S, Nisnevitch M. Antimicrobial and Antiviral Compounds of Phlomis viscosa Poiret. Biomedicines. 2023; 11(2):441. https://doi.org/10.3390/biomedicines11020441
Chicago/Turabian StyleYarmolinsky, Ludmila, Faina Nakonechny, Arie Budovsky, Haim Zeigerman, Boris Khalfin, Eyal Sharon, Leonid Yarmolinsky, Shimon Ben-Shabat, and Marina Nisnevitch. 2023. "Antimicrobial and Antiviral Compounds of Phlomis viscosa Poiret" Biomedicines 11, no. 2: 441. https://doi.org/10.3390/biomedicines11020441
APA StyleYarmolinsky, L., Nakonechny, F., Budovsky, A., Zeigerman, H., Khalfin, B., Sharon, E., Yarmolinsky, L., Ben-Shabat, S., & Nisnevitch, M. (2023). Antimicrobial and Antiviral Compounds of Phlomis viscosa Poiret. Biomedicines, 11(2), 441. https://doi.org/10.3390/biomedicines11020441