

Oncogenic Long Noncoding RNAs in Prostate Cancer, Osteosarcoma, and Metastasis


Abstract
:1. Introduction
2. LncRNAs and Their Mechanism
3. Oncogenic lncRNAs in PC and OS Malignancy
3.1. Differentiation Antagonizing Non-Protein Coding RNA
3.2. Metastasis Associated Lung Adenocarcinoma Transcript 1
3.3. Nuclear Enriched Abundant Transcript 1
3.4. HOX Transcript Antisense RNA
3.5. Taurine Upregulated Gene 1
3.6. PC Associated Transcript 1
3.7. LncRNA-Activated by TGF-β
3.8. Plasmacytoma Variant Translocation 1
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, L.; Lu, B.; He, M.; Wang, Y.; Wang, Z.; Du, L. Prostate Cancer Incidence and Mortality: Global Status and Temporal Trends in 89 Countries From 2000 to 2019. Front. Public Health 2022, 10, 811044. [Google Scholar] [CrossRef] [PubMed]
- Valery, P.C.; Laversanne, M.; Bray, F. Bone cancer incidence by morphological subtype: A global assessment. Cancer Causes Control 2015, 26, 1127–1139. [Google Scholar] [CrossRef] [PubMed]
- Ghafouri-Fard, S.; Shirvani-Farsani, Z.; Hussen, B.M.; Taheri, M. The critical roles of lncRNAs in the development of osteosarcoma. Biomed. Pharmacother. 2021, 135, 111217. [Google Scholar] [CrossRef] [PubMed]
- Rojas, G.A.; Hubbard, A.K.; Diessner, B.J.; Ribeiro, K.B.; Spector, L.G. International trends in incidence of osteosarcoma (1988–2012). Int. J. Cancer 2021, 149, 1044–1053. [Google Scholar] [CrossRef]
- Singh, A.; Malvankar, S.; Ravi Kumar, Y.S.; Seervi, M.; Srivastava, R.K.; Verma, B. Role of various non-coding RNAs in EMT, cancer, and metastasis: Recent trends and future perspective. Adv. Cancer Biol.-Metastasis 2022, 4, 100039. [Google Scholar] [CrossRef]
- Nie, H.; Liao, Z.; Wang, Y.; Zhou, J.; He, X.; Ou, C. Exosomal long non-coding RNAs: Emerging players in cancer metastasis and potential diagnostic biomarkers for personalized oncology. Genes Dis. 2021, 8, 769–780. [Google Scholar] [CrossRef]
- Castaneda, M.; den Hollander, P.; Kuburich, N.A.; Rosen, J.M.; Mani, S.A. Mechanisms of cancer metastasis. Semin. Cancer Biol. 2022, 87, 17–31. [Google Scholar] [CrossRef]
- Tay, K.J.; Moul, J.W.; Armstrong, A.J. Management of Prostate Cancer in the Elderly. Clin. Geriatr. Med. 2016, 32, 113–132. [Google Scholar] [CrossRef]
- Li, Y.; Egranov, S.D.; Yang, L.; Lin, C. Molecular mechanisms of long noncoding RNAs-mediated cancer metastasis. Genes Chromosom. Cancer 2019, 58, 200–207. [Google Scholar] [CrossRef]
- Kahn, D.; Weiner, G.J.; Ben-Haim, S.; Boles Ponto, L.L.; Madsen, M.T.; Bushnell, D.L.; Watkins, G.L.; Argenyi, E.A.; Hichwa, R.D. Positron Emission Tomographic Measurement of Bone Marrow Blood Flow to the Pelvis and Lumbar Vertebrae in Young Normal Adults. Blood 1994, 83, 958–963. [Google Scholar] [CrossRef] [Green Version]
- Guise, T.A. The vicious cycle of bone metastases. J. Musculoskelet. Neuronal Interact. 2002, 2, 570–572. [Google Scholar] [PubMed]
- Guise, T.A.; Mohammad, K.S.; Clines, G.; Stebbins, E.G.; Wong, D.H.; Higgins, L.S.; Vessella, R.; Corey, E.; Padalecki, S.; Suva, L.; et al. Basic mechanisms responsible for osteolytic and osteoblastic bone metastases. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2006, 12, 6213s–6216s. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fizazi, K.; Yang, J.; Peleg, S.; Sikes, C.R.; Kreimann, E.L.; Daliani, D.; Olive, M.; Raymond, K.A.; Janus, T.J.; Logothetis, C.J.; et al. Prostate cancer cells-osteoblast interaction shifts expression of growth/survival-related genes in prostate cancer and reduces expression of osteoprotegerin in osteoblasts. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2003, 9, 2587–2597. [Google Scholar]
- Ottewell, P.D. The role of osteoblasts in bone metastasis. J. Bone Oncol. 2016, 5, 124–127. [Google Scholar] [CrossRef] [Green Version]
- Casimiro, S.; Ferreira, A.R.; Mansinho, A.; Alho, I.; Costa, L. Molecular Mechanisms of Bone Metastasis: Which Targets Came from the Bench to the Bedside? Int. J. Mol. Sci. 2016, 17, 1415. [Google Scholar] [CrossRef] [Green Version]
- David Roodman, G.; Silbermann, R. Mechanisms of osteolytic and osteoblastic skeletal lesions. BoneKEy Rep. 2015, 4, 753. [Google Scholar] [CrossRef] [Green Version]
- Buijs, J.T.; Rentsch, C.A.; van der Horst, G.; van Overveld, P.G.; Wetterwald, A.; Schwaninger, R.; Henriquez, N.V.; Ten Dijke, P.; Borovecki, F.; Markwalder, R.; et al. BMP7, a putative regulator of epithelial homeostasis in the human prostate, is a potent inhibitor of prostate cancer bone metastasis in vivo. Am. J. Pathol. 2007, 171, 1047–1057. [Google Scholar] [CrossRef] [Green Version]
- Chaffer, C.L.; Thompson, E.W.; Williams, E.D. Mesenchymal to epithelial transition in development and disease. Cells Tissues Organs 2007, 185, 7–19. [Google Scholar] [CrossRef]
- van der Pluijm, G. Epithelial plasticity, cancer stem cells and bone metastasis formation. Bone 2011, 48, 37–43. [Google Scholar] [CrossRef]
- Yu, X.; Yustein, J.T.; Xu, J. Research models and mesenchymal/epithelial plasticity of osteosarcoma. Cell Biosci. 2021, 11, 94. [Google Scholar] [CrossRef]
- Sannino, G.; Marchetto, A.; Kirchner, T.; Grünewald, T.G.P. Epithelial-to-Mesenchymal and Mesenchymal-to-Epithelial Transition in Mesenchymal Tumors: A Paradox in Sarcomas? Cancer Res. 2017, 77, 4556–4561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aird, J.; Baird, A.-M.; Lim, M.C.J.; McDermott, R.; Finn, S.P.; Gray, S.G. Carcinogenesis in prostate cancer: The role of long non-coding RNAs. Non-Coding RNA Res. 2018, 3, 29–38. [Google Scholar] [CrossRef]
- Mukhametov, U.; Lyulin, S.; Borzunov, D.; Sufianova, G.; Shumadalova, A.; Zhang, D.; Gareev, I. Functions of the bone morphogenetic protein signaling pathway through non-coding RNAs. Non-Coding RNA Res. 2022, 7, 178–183. [Google Scholar] [CrossRef] [PubMed]
- Sikora, M.; Marycz, K.; Smieszek, A. Small and Long Non-coding RNAs as Functional Regulators of Bone Homeostasis, Acting Alone or Cooperatively. Mol. Ther.-Nucleic Acids 2020, 21, 792–803. [Google Scholar] [CrossRef] [PubMed]
- Volders, P.J.; Anckaert, J.; Verheggen, K.; Nuytens, J.; Martens, L.; Mestdagh, P.; Vandesompele, J. LNCipedia 5: Towards a reference set of human long non-coding RNAs. Nucleic Acids Res. 2019, 47, D135–D139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Li, Z.; Liu, C.; Zou, D.; Li, Q.; Feng, C.; Jing, W.; Luo, S.; Zhang, Z.; Ma, L. LncRNAWiki 2.0: A knowledgebase of human long non-coding RNAs with enhanced curation model and database system. Nucleic Acids Res. 2022, 50, D190–D195. [Google Scholar] [CrossRef]
- Peng, S.; Cao, L.; He, S.; Zhong, Y.; Ma, H.; Zhang, Y.; Shuai, C. An Overview of Long Noncoding RNAs Involved in Bone Regeneration from Mesenchymal Stem Cells. Stem Cells Int. 2018, 2018, 8273648. [Google Scholar] [CrossRef]
- Anil, P.; Ghosh Dastidar, S.; Banerjee, S. Unravelling the role of long non-coding RNAs in prostate carcinoma. Adv. Cancer Biol.-Metastasis 2022, 6, 100067. [Google Scholar] [CrossRef]
- Wang, D.; Zhang, W.; Zhang, C.; Wang, L.; Chen, H.; Xu, J. Exosomal non-coding RNAs have a significant effect on tumor metastasis. Mol. Ther.-Nucleic Acids 2022, 29, 16–35. [Google Scholar] [CrossRef]
- Tay, Y.; Rinn, J.; Pandolfi, P.P. The multilayered complexity of ceRNA crosstalk and competition. Nature 2014, 505, 344–352. [Google Scholar] [CrossRef] [Green Version]
- Sebastian-delaCruz, M.; Gonzalez-Moro, I.; Olazagoitia-Garmendia, A.; Castellanos-Rubio, A.; Santin, I. The Role of lncRNAs in Gene Expression Regulation through mRNA Stabilization. Non-Coding RNA 2021, 7, 3. [Google Scholar] [CrossRef]
- Zhang, Y.; Liao, G.; Bai, J.; Zhang, X.; Xu, L.; Deng, C.; Yan, M.; Xie, A.; Luo, T.; Long, Z.; et al. Identifying Cancer Driver lncRNAs Bridged by Functional Effectors through Integrating Multi-omics Data in Human Cancers. Mol. Ther.-Nucleic Acids 2019, 17, 362–373. [Google Scholar] [CrossRef] [Green Version]
- Mu, H.; Zhang, S.; Yao, Z.; Liu, Y.; Lin, K.; Zhao, Z.; Zhu, Y. The diagnostic and prognostic value of exosome-derived long non-coding RNAs in cancer patients: A meta-analysis. Clin. Exp. Med. 2020, 20, 339–348. [Google Scholar] [CrossRef]
- Jiang, M.C.; Ni, J.J.; Cui, W.Y.; Wang, B.Y.; Zhuo, W. Emerging roles of lncRNA in cancer and therapeutic opportunities. Am. J. Cancer Res. 2019, 9, 1354–1366. [Google Scholar]
- Jia, J.; Li, F.; Tang, X.S.; Xu, S.; Gao, Y.; Shi, Q.; Guo, W.; Wang, X.; He, D.; Guo, P. Long noncoding RNA DANCR promotes invasion of prostate cancer through epigenetically silencing expression of TIMP2/3. Oncotarget 2016, 7, 37868–37881. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.Y.; Chen, C. lncRNA-DANCR Promotes Taxol Resistance of Prostate Cancer Cells through Modulating the miR-33b-5p-LDHA Axis. Dis. Mrk. 2022, 2022, 9516774. [Google Scholar] [CrossRef]
- Deng, H.; Zhu, B.; Dong, Z.; Jiang, H.; Zhao, X.; Wu, S. miR-214-5p targeted by LncRNA DANCR mediates TGF-β signaling pathway to accelerate proliferation, migration and inhibit apoptosis of prostate cancer cells. Am. J. Transl. Res. 2021, 13, 2224–2240. [Google Scholar] [PubMed]
- Jiang, N.; Wang, X.; Xie, X.; Liao, Y.; Liu, N.; Liu, J.; Miao, N.; Shen, J.; Peng, T. lncRNA DANCR promotes tumor progression and cancer stemness features in osteosarcoma by upregulating AXL via miR-33a-5p inhibition. Cancer Lett. 2017, 405, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Bi, Y.; Guo, S.; Xu, X.; Kong, P.; Cui, H.; Yan, T.; Ma, Y.; Cheng, Y.; Chen, Y.; Liu, X.; et al. Decreased ZNF750 promotes angiogenesis in a paracrine manner via activating DANCR/miR-4707-3p/FOXC2 axis in esophageal squamous cell carcinoma. Cell Death Dis. 2020, 11, 296. [Google Scholar] [CrossRef] [PubMed]
- Ren, S.; Liu, Y.; Xu, W.; Sun, Y.; Lu, J.; Wang, F.; Wei, M.; Shen, J.; Hou, J.; Gao, X.; et al. Long noncoding RNA MALAT-1 is a new potential therapeutic target for castration resistant prostate cancer. J. Urol. 2013, 190, 2278–2287. [Google Scholar] [CrossRef]
- Sebastian, A.; Hum, N.R.; Hudson, B.D.; Loots, G.G. Cancer-Osteoblast Interaction Reduces Sost Expression in Osteoblasts and Up-Regulates lncRNA MALAT1 in Prostate Cancer. Microarrays 2015, 4, 503–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Huang, W.; Sun, W.; Zheng, B.; Wang, C.; Luo, Z.; Wang, J.; Yan, W. LncRNA MALAT1 Promotes Cancer Metastasis in Osteosarcoma via Activation of the PI3K-Akt Signaling Pathway. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2018, 51, 1313–1326. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Zhang, T.; Chen, B. Long Non-Coding RNA Metastasis-Associated Lung Adenocarcinoma Transcript 1 (MALAT1) Promotes Proliferation and Metastasis of Osteosarcoma Cells by Targeting c-Met and SOX4 via miR-34a/c-5p and miR-449a/b. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2019, 25, 1410–1422. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Piao, C.D.; Ding, J.; Li, Z.W. LncRNA MALAT1 facilitates lung metastasis of osteosarcomas through miR-202 sponging. Sci. Rep. 2020, 10, 12757. [Google Scholar] [CrossRef] [PubMed]
- Wen, S.; Wei, Y.; Zen, C.; Xiong, W.; Niu, Y.; Zhao, Y. Long non-coding RNA NEAT1 promotes bone metastasis of prostate cancer through N6-methyladenosine. Mol. Cancer 2020, 19, 171. [Google Scholar] [CrossRef]
- Zhao, W.; Zhu, X.; Jin, Q.; Lin, B.; Ji, R. The lncRNA NEAT1/miRNA-766-5p/E2F3 Regulatory Axis Promotes Prostate Cancer Progression. J. Oncol. 2022, 2022, 1866972. [Google Scholar] [CrossRef]
- Chen, Y.; Li, J.; Xiao, J.K.; Xiao, L.; Xu, B.W.; Li, C. The lncRNA NEAT1 promotes the epithelial-mesenchymal transition and metastasis of osteosarcoma cells by sponging miR-483 to upregulate STAT3 expression. Cancer Cell Int. 2021, 21, 90. [Google Scholar] [CrossRef]
- Li, P.; Huang, R.; Huang, T.; Cheng, S.; Chen, Y.; Wang, Z. Long non-coding RNA NEAT1 promotes proliferation, migration and invasion of human osteosarcoma cells. Int. J. Med. Sci. 2018, 15, 1227–1234. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Liu, N.; Gao, Y.; Quan, Z.; Hao, Y.; Yu, C.; Li, L.; Yuan, M.; Niu, L.; Luo, C.; et al. Long noncoding RNA HOTAIR regulates the invasion and metastasis of prostate cancer by targeting hepaCAM. Br. J. Cancer 2021, 124, 247–258. [Google Scholar] [CrossRef]
- Zhang, A.; Zhao, J.C.; Kim, J.; Fong, K.W.; Yang, Y.A.; Chakravarti, D.; Mo, Y.Y.; Yu, J. LncRNA HOTAIR Enhances the Androgen-Receptor-Mediated Transcriptional Program and Drives Castration-Resistant Prostate Cancer. Cell Rep. 2015, 13, 209–221. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.J.; Zhou, X.H.; Zhou, Y.; Wang, Y.G.; Qian, B.Z.; He, A.N.; Shen, Z.; Hu, H.Y.; Yao, Y. Bufalin suppresses the migration and invasion of prostate cancer cells through HOTAIR, the sponge of miR-520b. Acta Pharmacol. Sin. 2019, 40, 1228–1236. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Qu, X.L.; Liu, J.; Lu, J.; Zhou, Z.Y. HOTAIR promotes osteosarcoma development by sponging miR-217 and targeting ZEB1. J. Cell. Physiol. 2019, 234, 6173–6181. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Min, J. Role of Long Noncoding RNA HOTAIR in the Growth and Apoptosis of Osteosarcoma Cell MG-63. BioMed Res. Int. 2016, 2016, 5757641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, J.; Dou, D.; Zhang, T.; Wang, B. HOTAIR Promotes Cisplatin Resistance of Osteosarcoma Cells by Regulating Cell Proliferation, Invasion, and Apoptosis via miR-106a-5p/STAT3 Axis. Cell Transplant. 2020, 29, 963689720948447. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Jia, H.H.; Xu, Y.Q.; Zhou, X.; Zhao, X.H.; Wang, Y.F.; Song, X.; Zhu, Z.Y.; Sun, T.; Dou, Y.; et al. Paracrine and epigenetic control of CAF-induced metastasis: The role of HOTAIR stimulated by TGF-ß1 secretion. Mol. Cancer 2018, 17, 5. [Google Scholar] [CrossRef]
- Yang, B.; Tang, X.; Wang, Z.; Sun, D.; Wei, X.; Ding, Y. TUG1 promotes prostate cancer progression by acting as a ceRNA of miR-26a. Biosci. Rep. 2018, 38, BSR20180677. [Google Scholar] [CrossRef] [Green Version]
- Xiu, D.; Liu, L.; Cheng, M.; Sun, X.; Ma, X. Knockdown of lncRNA TUG1 Enhances Radiosensitivity of Prostate Cancer via the TUG1/miR-139-5p/SMC1A Axis. OncoTargets Ther. 2020, 13, 2319–2331. [Google Scholar] [CrossRef] [Green Version]
- Hao, S.D.; Ma, J.X.; Liu, Y.; Liu, P.J.; Qin, Y. Long non-coding TUG1 accelerates prostate cancer progression through regulating miR-128-3p/YES1 axis. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 619–632. [Google Scholar] [CrossRef]
- Sheng, K.; Li, Y. LncRNA TUG1 promotes the development of osteosarcoma through RUNX2. Exp. Ther. Med. 2019, 18, 3002–3008. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.; Hu, T.; Xu, Y. Anticancer potential of TUG1 knockdown in cisplatin-resistant osteosarcoma through inhibition of MET/Akt signalling. J. Drug Target. 2020, 28, 204–211. [Google Scholar] [CrossRef]
- Yu, X.; Hu, L.; Li, S.; Shen, J.; Wang, D.; Xu, R.; Yang, H. Long non-coding RNA Taurine upregulated gene 1 promotes osteosarcoma cell metastasis by mediating HIF-1α via miR-143-5p. Cell Death Dis. 2019, 10, 280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, G.; Liu, K.; Du, X. Long Non-Coding RNA TUG1 Promotes Proliferation and Inhibits Apoptosis of Osteosarcoma Cells by Sponging miR-132-3p and Upregulating SOX4 Expression. Yonsei Med. J. 2018, 59, 226–235. [Google Scholar] [CrossRef] [PubMed]
- Shang, Z.; Yu, J.; Sun, L.; Tian, J.; Zhu, S.; Zhang, B.; Dong, Q.; Jiang, N.; Flores-Morales, A.; Chang, C.; et al. LncRNA PCAT1 activates AKT and NF-κB signaling in castration-resistant prostate cancer by regulating the PHLPP/FKBP51/IKKα complex. Nucleic Acids Res. 2019, 47, 4211–4225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Zhang, Y.; Mao, Y.; Ma, X. The lncRNA PCAT1 is correlated with poor prognosis and promotes cell proliferation, invasion, migration and EMT in osteosarcoma. OncoTargets Ther. 2018, 11, 629–638. [Google Scholar] [CrossRef] [Green Version]
- Chang, L.; Jia, D.L.; Cao, C.S.; Wei, H.; Li, Z.Q. LncRNA PCAT-1 promotes the progression of osteosarcoma via miR-508-3p/ZEB1 axis. Eur. Rev. Med. Pharmacol. Sci. 2021, 25, 2517–2527. [Google Scholar] [CrossRef]
- Xu, S.; Yi, X.M.; Tang, C.P.; Ge, J.P.; Zhang, Z.Y.; Zhou, W.Q. Long non-coding RNA ATB promotes growth and epithelial-mesenchymal transition and predicts poor prognosis in human prostate carcinoma. Oncol. Rep. 2016, 36, 10–22. [Google Scholar] [CrossRef] [Green Version]
- Han, F.; Wang, C.; Wang, Y.; Zhang, L. Long noncoding RNA ATB promotes osteosarcoma cell proliferation, migration and invasion by suppressing miR-200s. Am. J. Cancer Res. 2017, 7, 770–783. [Google Scholar]
- Wu, H.; Tian, X.; Zhu, C. Knockdown of lncRNA PVT1 inhibits prostate cancer progression in vitro and in vivo by the suppression of KIF23 through stimulating miR-15a-5p. Cancer Cell Int. 2020, 20, 283. [Google Scholar] [CrossRef]
- Liu, J.; Li, Y.; Zhang, Q.; Lv, C.; Wang, M.; Jiao, Y.; Wang, C. PVT1 Expression Is a Predictor for Poor Survival of Prostate Cancer Patients. Technol. Cancer Res. Treat. 2021, 20, 1533033820971610. [Google Scholar] [CrossRef]
- Sun, F.; Wu, K.; Yao, Z.; Mu, X.; Zheng, Z.; Sun, M.; Wang, Y.; Liu, Z.; Zhu, Y. Long Noncoding RNA PVT1 Promotes Prostate Cancer Metastasis by Increasing NOP2 Expression via Targeting Tumor Suppressor MicroRNAs. OncoTargets Ther. 2020, 13, 6755–6765. [Google Scholar] [CrossRef]
- Yan, M.; Pan, X.F.; Liu, Y.; Zhao, S.; Gong, W.Q.; Liu, W. Long noncoding RNA PVT1 promotes metastasis via miR-484 sponging in osteosarcoma cells. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 2229–2238. [Google Scholar] [CrossRef] [PubMed]
- Xun, C.; Jiang, D.; Tian, Z.; Yunus, A.; Chen, J. Long noncoding RNA plasmacytoma variant translocation gene 1 promotes epithelial-mesenchymal transition in osteosarcoma. J. Clin. Lab. Anal. 2021, 35, e23587. [Google Scholar] [CrossRef] [PubMed]
- Smolle, M.A.; Bauernhofer, T.; Pummer, K.; Calin, G.A.; Pichler, M. Current Insights into Long Non-Coding RNAs (LncRNAs) in Prostate Cancer. Int. J. Mol. Sci. 2017, 18, 473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Chen, L. An integrated analysis of the competing endogenous RNA network and co-expression network revealed seven hub long non-coding RNAs in osteoarthritis. Bone Jt. Res. 2020, 9, 90–98. [Google Scholar] [CrossRef]
- Chen, R.; Wang, G.; Zheng, Y.; Hua, Y.; Cai, Z. Long non-coding RNAs in osteosarcoma. Oncotarget 2017, 8, 20462–20475. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Gu, J.; Zhang, X.; Yang, J.; Zhang, X.; Fang, X. Long Non-coding RNA DANCR in Cancer: Roles, Mechanisms, and Implications. Front. Cell Dev. Biol. 2021, 9, 753706. [Google Scholar] [CrossRef]
- Li, Z.X.; Zhu, Q.N.; Zhang, H.B.; Hu, Y.; Wang, G.; Zhu, Y.S. MALAT1: A potential biomarker in cancer. Cancer Manag. Res. 2018, 10, 6757–6768. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Hao, X.; Yin, M.; Xu, T.; Guo, F. Long non-coding RNA in osteogenesis: A new world to be explored. Bone Jt. Res. 2019, 8, 73–80. [Google Scholar] [CrossRef]
- Deng, J.; Tang, J.; Wang, G.; Zhu, Y.S. Long Non-Coding RNA as Potential Biomarker for Prostate Cancer: Is It Making a Difference? Int. J. Environ. Res. Public Health 2017, 14, 270. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Li, Z.; Zheng, H.; Chan, M.T.; Wu, W.K. NEAT1: A novel cancer-related long non-coding RNA. Cell Prolif. 2017, 50, e12329. [Google Scholar] [CrossRef] [Green Version]
- Hajjari, M.; Salavaty, A. HOTAIR: An oncogenic long non-coding RNA in different cancers. Cancer Biol. Med. 2015, 12, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Shen, J.; Chan, M.T.; Wu, W.K. TUG1: A pivotal oncogenic long non-coding RNA of human cancers. Cell Prolif. 2016, 49, 471–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghafouri-Fard, S.; Dashti, S.; Taheri, M. PCAT1: An oncogenic lncRNA in diverse cancers and a putative therapeutic target. Exp. Mol. Pathol. 2020, 114, 104429. [Google Scholar] [CrossRef]
- Tang, F.; Xu, Y.; Wang, H.; Bian, E.; Zhao, B. LncRNA-ATB in cancers: What do we know so far? Mol. Biol. Rep. 2020, 47, 4077–4086. [Google Scholar] [CrossRef] [PubMed]
- Onagoruwa, O.T.; Pal, G.; Ochu, C.; Ogunwobi, O.O. Oncogenic Role of PVT1 and Therapeutic Implications. Front. Oncol. 2020, 10, 17. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Zhou, R.; Wu, Y.; Liu, Y.; Su, W.; Xiong, W.; Zeng, Z. PVT1 Promotes Cancer Progression via MicroRNAs. Front. Oncol. 2019, 9, 609. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
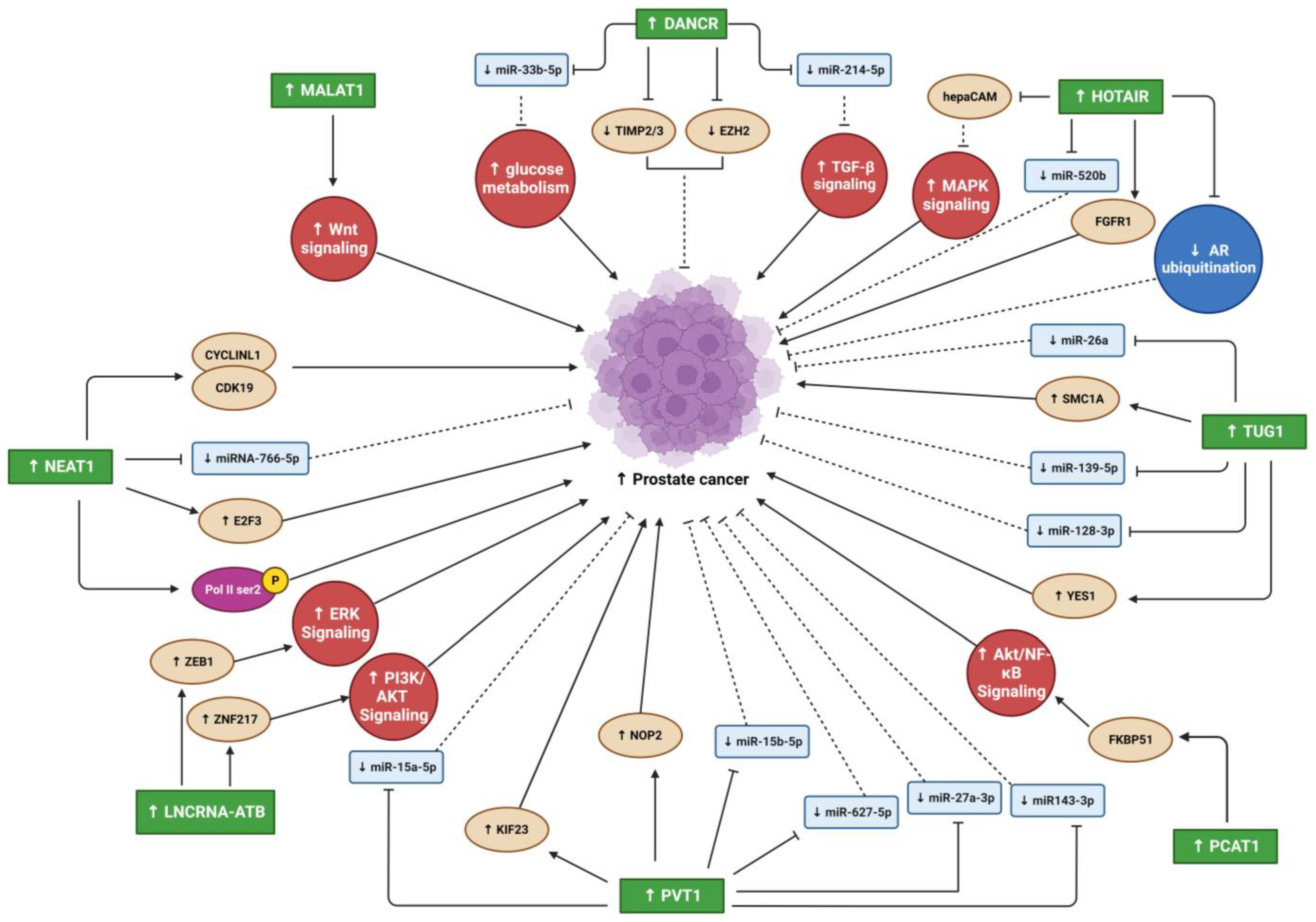{kind=link}
{kind=link}
| LncRNA | Cancer Type | Tumor Samples (Human Tissues, Cell Lines, and Animal Models) | LncRNA Overexpression Downstream Effect | Affected Pathway | LncRNA Upregulation Outcomes | LncRNA Downregulation Outcomes | References |
|---|---|---|---|---|---|---|---|
| DANCR | PC metastasis | 150 human PC samples; three PC cell lines (CWR22Rv1, PC3, and C4-2B); xenograft tumor model (nude mice). | Decreased TIMP2/3 expression synergistic with EZH2 by silencing their promoters. | - | Promotes invasion and migration in vitro; promotes metastasis in vivo. | TIMP2/3 upregulation; Enzalutamide treatment inhibits invasion and migration in vitro; androgen-AR suppresses invasion and migration in vivo. | [35] |
| PC | 30 human prostate samples; four PC cell lines (PC3, DU145, LN96, and OPCT-1). | Promotes Taxol resistance in both tissues and cell lines; sponges miR-33b-5p. | Increases glucose metabolism. | Promotes glycolysis in Taxol-resistant cell lines. | Inhibits proliferation and migration in vitro; increased Taxol sensitivity in vitro. | [36] | |
| PC | 53 human PC samples; four PC cell lines (DU145, 22Rv1, RC-92a, and PC-3M). | Sponges miR-214-5p. | Increases TGF-β signaling. | Promotes proliferation and migration in vitro; decreases apoptosis in vitro. | Decreases proliferation and migration in vitro; increases apoptosis in vitro. | [37] | |
| OS metastasis | 34 human OS samples; five OS cell lines (MG-63, U2OS, Saos2, HOS, and 143B); xenograft tumor model (male BALB/C nude mice). | Sponges miR-33a-5p; increases AXL expression. | Increases PI3K-AKT signaling. | Promotes proliferation, migration, and invasion in vitro; promotes tumor progression and lung metastasis in vivo; enhances the stem cells characteristics of OS cells. | - | [38] | |
| Metastasis | 612 ESCC patients; eight ESCC cell lines (KYSE140, KYSE150, KYSE180, KYSE410, KYSE510, KYSE450, Colo680N, and ECA109); xenograft tumor model (BALB/c nude female mice). | Sponges miR4707-3p; increases FOXC2 expression. | Enhances FOXC2 signaling. | Induces tumor angiogenesis. | Decreases FOXC2 expression. | [39] | |
| MALAT1 | PC metastasis | Human CRPC tissue; two PC cell lines (22RV1 and LNCaP-AI); xenograft tumor model (castrated male Sprague–Dawley athymic nude rats). | - | - | Increases Gleason score, tumor progression, and PC castration resistance. | Reduces cell growth, invasion, migration, and formation of cancer cell colonies in vitro; promotes cell cycle arrest and apoptosis rates in vitro; delays tumor growth and reduces metastasis in vivo. | [40] |
| PC metastasis | One PC cell line (PC3); one OS cell line (UMR-106); primary osteoblasts; xenograft tumor model (wildtype C57BL/6 and SostKO mice (C57BL/6 background)). | - | Wnt signaling. | Promotes PC-associated bone metastasis. | - | [41] | |
| OS metastasis | 68 human OS samples; five OS cell lines (MG63, U2OS, Saos-2, SOSP-9607, and SW1353); xenograft tumor model (male BALB/C nude mice). | Sponges miR-129-5p; increases RET expression. | PI3K-AKT signaling. | Increases cell proliferation, migration, and invasion in vitro; promotes tumor growth in vivo; correlates with tumor progression and metastasis in OS tissues; mediates tumor stem cell characteristics. | - | [42] | |
| OS metastasis | 76 human OS samples; four OS cell lines (MNNG/HOS, Saos-2, U2OS, and MG-63). | Sponges miR-34a/c-5p and miR-449a/b; increases c-Met and SOX4 expression. | - | Induces OS cell proliferation and metastasis. | Decreases OS cell proliferation, migration, and invasion. | [43] | |
| OS metastasis | 32 OS tissues from patients without metastasis; 24 OS tissues from patients with lung metastases; four OS cell lines (KRIB, Saos-2, MG63, and U2OS). | Sponges miR-202. | - | Enhances OS-associated lung metastasis. | Decreases OS cell invasiveness. | [44] | |
| NEAT1 | PC metastasis | Primary patient-derived PC and metastatic bone tissues; patient-derived xenograft model (male athymic nude mice). | Enhances the binding of CCNL1 to CDK19; promotes Pol II Ser2 phosphorylation. | - | Induces PC cell metastasis to lung and bone; reduces xenograft model survival. | Decreases Pol II Ser2p levels in the RUNX2 promoter. | [45] |
| PC | 50 human PC samples; four PC cell lines (PC3, P4E6, LNCaP, and DU145). | Sponges miR-766-5p; activates E2F3. | - | Increases proliferation, migration, and invasion. | Decreases proliferation, invasion, and migration in vitro; stimulates apoptosis and cell cycle arrest in vitro. | [46] | |
| OS metastasis | 20 human OS samples; three OS cell lines (U2OS, MG-63 and Saos2); xenograft tumor model (female BALB/C nude mice). | Sponges miR-483; activates STAT3 expression. | - | Enhances migration, invasion, and EMT in vitro. | Reduces OS-associated metastases in vivo; suppresses OS MET at metastasis sites. | [47] | |
| OS metastasis | 19 human OS samples; one OS cell line (U2OS). | - | - | - | Suppresses OS proliferation, migration, and invasion in vitro. | [48] | |
| HOTAIR | PC metastasis | 70 human PC samples; three PC cell lines (RWPE-1, PC3 and DU145). | Suppresses the hepaCAM gene | Activates MAPK signaling. | Increases PC cell invasion and metastasis. | Reduces invasiveness and metastasis in vitro. | [49] |
| PC | Human PC samples; chromatin immunoprecipitation sequencing (ChIP-seq) data of PC cell lines (LNCaP); three PC cell lines (LNCaP, LAPC4 and C4-2B) | Prevents MDM2 from binding to AR protein NTD | Inhibits AR ubiquitination | Activates AR without the presence of androgen; promotes many AR genes similar to those induced by androgen; increases LNCaP cell proliferation and invasion. | Decreases PC cell proliferation and invasion in CRPC cell line (C4-2B). | [50] | |
| PC Metastasis | 40 serum samples from PC patients with bone metastasis; 40 serum samples from PC patients without bone metastasis; 40 PC patients primary tissues and bone metastatic tissues; two PC cell lines (PC3 and DU145). | Sponges miR-520b; activates FGFR1 expression | - | Promotes migration and invasion of (PC3 and DU145) cells; promotes PC-associated bone metastasis | - | [51] | |
| OS | Four OS cell lines (U2OS, MG-63, Saos2, and SW1353). | Sponges miR-217; activates ZEB1 expression. | - | Promotes OS progression. | Inhibits OS progression. | [52] | |
| OS | One OS cell line (MG-63). | - | Activates TGF-β, BCL2, but inhibits TP53, and TNF-α signaling | Promotes proliferation of MG-63 cells; inhibits apoptosis of MG-63 cells. | Downregulation of TGF-β and BCL2 expression; upregulation of TP53 and TNF-α expression. | [53] | |
| OS | 60 human OS samples; three OS cell lines (Saos2, MG-63, and U2OS); three DDP-resistant OS cell lines (Saos2/DDP, MG-63/DDP, and U2OS/DDP). | Sponges miR-106a-5p; activates STAT3 expression. | - | Increases cell proliferation and invasion; reduces apoptosis; promotes DDP-resistance. | Decreases DDP resistance in Saos2/DDP, MG-63/DDP, and U2OS/DDP cells; inhibits cell proliferation and invasion in DDP-resistant cells; promotes apoptosis in DDP-resistant cells. | [54] | |
| Metastasis | Two breast cancer cell lines (MDA-MB-231 and MCF-7); CAFs from four invasive breast cancer patients. | Stimulates TGF-β1 secretion. | Activates CDK5 signaling. | Induces EMT and metastasis. | Inhibits CAF-induced tumor progression and metastasis. | [55] | |
| TUG1 | PC metastasis | 86 human PC samples; four PC cell lines (DU145, PC3, LNCaP, and 22RV1). | Sponges miR-26a. | - | Promotes PC progression and metastasis in vitro. | Induces apoptosis in vitro. | [56] |
| PC | 50 human PC samples; four PC cell lines (LNCaP, 22RV1, PC3, and DU145); xenograft tumor model (male nude mice). | Sponges miR-139-5p; SMC1A overexpression. | - | - | Reduces cell proliferation; enhances apoptosis; enhances radiosensitivity in vivo and in vitro. | [57] | |
| PC | 30 human PC samples; two PC cell lines (PC3 and DU145); xenograft tumor model (male BALB/C mice). | Sponges miR-128-3p; increases YES1 expression. | - | - | Suppresses cell proliferation, EMT, migration, and invasion in vitro; increases apoptosis in vitro; reduces tumor growth in vivo. | [58] | |
| OS | 40 human OS samples; two OS cell lines (MG-63 and U2OS). | Activates RUNX2 expression. | - | Poor prognosis and low survival in patients; increases proliferation, migration, and invasion in vitro. | Decreases proliferation, migration, and invasion in vitro. | [59] | |
| OS | Two OS cell lines (MG-63 and Saos2); Two OS DDP-resistant cell lines (Saos2/DDP and MG-63/DDP); xenograft tumor model (male BALB/C nude mice). | - | TUG1 knockdown inhibited MET/AKT signaling. | - | Reduces OS DDP resistance in vitro; increases apoptosis in vitro; decreases DDP resistance in vivo. | [60] | |
| OS metastasis | Human OS tissues; five OS cell lines (143B, HOS, MG-63, Saos2, and U2OS); CAFs from human OS tissues; xenograft tumor model (male nude mice). | Sponges miR143-5p; activates HIF-1α expression. | Activates HIF-1α signaling. | Poor prognosis in OS patients; promotes OS invasion and angiogenesis in vitro. | Decreases tumor growth, peritoneal spreading, and metastasis in vivo. | [61] | |
| OS | 22 human OS samples; four OS cell lines (U2OS, MG-63, Saos2, and 143B). | Sponges miR-132-3p; increases SOX4 expression. | - | - | Suppresses proliferation in vitro; induces apoptosis in vitro. | [62] | |
| PCAT1 | PC | Human PC tissues (ADPC and CRPC); Two PC cell lines (LNCaP and C4-2); xenograft tumor model (male nude mice). | Directly binds FKBP51. | Activates AKT/NF-κB signaling in CRPC. | Promotes CRPC progression. | Decreases AKT and NF-κB signaling; suppresses CRPC progression in vivo and in vitro. | [63] |
| OS metastasis | 30 human OS samples; four OS cell lines (LM7, KHOS, MG-63, and U2OS). | - | - | Correlates with poor prognosis and metastasis development in OS patients; promotes cell proliferation, invasion, and migration in vitro; reduces apoptosis rate in vitro; increases CDH2 and VIM expression in vitro; decreases CDH1 expression in vitro. | Decreases cell proliferation, invasion, and migration in vitro; increases apoptosis in vitro; decreases CDH2 and VIM expression in vitro; increases CDH1 expression in vitro. | [64] | |
| OS metastasis | 49 human OS samples; two OS cell lines (MG-63 and U2OS). | Sponges miR-508-3p; activates ZEB1 expression. | - | Indicates poor prognosis in OS patients; promotes cell proliferation, invasion, migration, and metastasis in vitro. | Reduces cell proliferation, invasion, migration, and metastasis in vitro. | [65] | |
| LncRNA-ATB | PC | 57 human PC samples; two PC cell lines (DU145 and PC3). | Activates ZEB1 and ZNF217 expression. | Activates ERK and PI3K/AKT signaling. | Promotes EMT by reducing CDH1 and ZO-1 expression; promotes CDH2 and VIM expression. | Reduces cell proliferation. | [66] |
| OS | 60 human OS samples; four OS cell lines (HOS, MG-63, Saos2, and U2OS); xenograft tumor model (athymic BALB/C nude mice). | Sponges miR-200s; activates ZEB1 and ZEB2 expression. | - | Associated with poor prognosis and metastasis in OS patients; promotes cell proliferation, migration, and invasion; promotes tumor growth in vivo. | Reduces cell proliferation, migration, and invasion. | [67] | |
| PVT1 | PC | 25 human PC samples; two PC cell lines (DU145 and 22RV1); xenograft tumor model (male BALB/C nude mice). | Sponges miR-15a-5p; induces KIF23 activity. | - | Promotes cell proliferation, invasion, and migration in vitro; promotes PC progression in tissues and in vivo. | Inhibits cell proliferation, invasion, and migration in vitro; increases apoptosis in vitro; suppresses tumorigenesis in vivo. | [68] |
| PC | PVT1 RNA-Seq and clinical data for 498 PC patients from the TCGA-PRAD database. | - | - | Poor prognosis and survival. | - | [69] | |
| PC metastasis | Four PC cell lines (VCaP, PC3, DU145, and 22RV1); xenograft tumor model (transgenic adenocarcinomas of mouse prostate (TRAMP) and ProbCre/Ptenfl/fl mice). | Sponges miR-15b-5p, miR-27a-3p, miR143-3p and miR-627-5p; activates NOP2 expression. | - | Promotes progression and metastasis in vivo; decreases survival time in vivo; increases migration and invasion in vitro. | Inhibits progression and metastasis in vivo; increases survival time in vivo; inhibits migration and invasion in vitro. | [70] | |
| OS metastasis | 48 human OS tissues; four OS cell lines (HOS, MG-63, 143B, and U2OS). | Sponges miR-486. | - | Promotes migration, invasion, and metastasis in vitro. | Inhibits migration, invasion, and metastasis in vitro. | [71] | |
| OS | 78 human OS samples; four OS cell lines (MG-63, SW1353, Saos2, and U2OS). | - | - | Poor prognosis. | Inhibits cell proliferation, invasion, and migration in vitro; decreases CDH2, SNAI1, and VIM expression in vitro; increases CDH1 expression in vitro. | [72] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Shehri, A.; Bakhashab, S. Oncogenic Long Noncoding RNAs in Prostate Cancer, Osteosarcoma, and Metastasis. Biomedicines 2023, 11, 633. https://doi.org/10.3390/biomedicines11020633
Al-Shehri A, Bakhashab S. Oncogenic Long Noncoding RNAs in Prostate Cancer, Osteosarcoma, and Metastasis. Biomedicines. 2023; 11(2):633. https://doi.org/10.3390/biomedicines11020633
Chicago/Turabian StyleAl-Shehri, Aishah, and Sherin Bakhashab. 2023. "Oncogenic Long Noncoding RNAs in Prostate Cancer, Osteosarcoma, and Metastasis" Biomedicines 11, no. 2: 633. https://doi.org/10.3390/biomedicines11020633
APA StyleAl-Shehri, A., & Bakhashab, S. (2023). Oncogenic Long Noncoding RNAs in Prostate Cancer, Osteosarcoma, and Metastasis. Biomedicines, 11(2), 633. https://doi.org/10.3390/biomedicines11020633