
Newborns from Mothers Who Intensely Consumed Sucralose during Pregnancy Are Heavier and Exhibit Markers of Metabolic Alteration and Low-Grade Systemic Inflammation: A Cross-Sectional, Prospective Study

 ,
,  , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Demographic, Anthropometric, and Clinical Measurements
2.3. Design of Light or Heavy Sucralose-Consuming Groups
2.4. Umbilical Cord Blood Samples
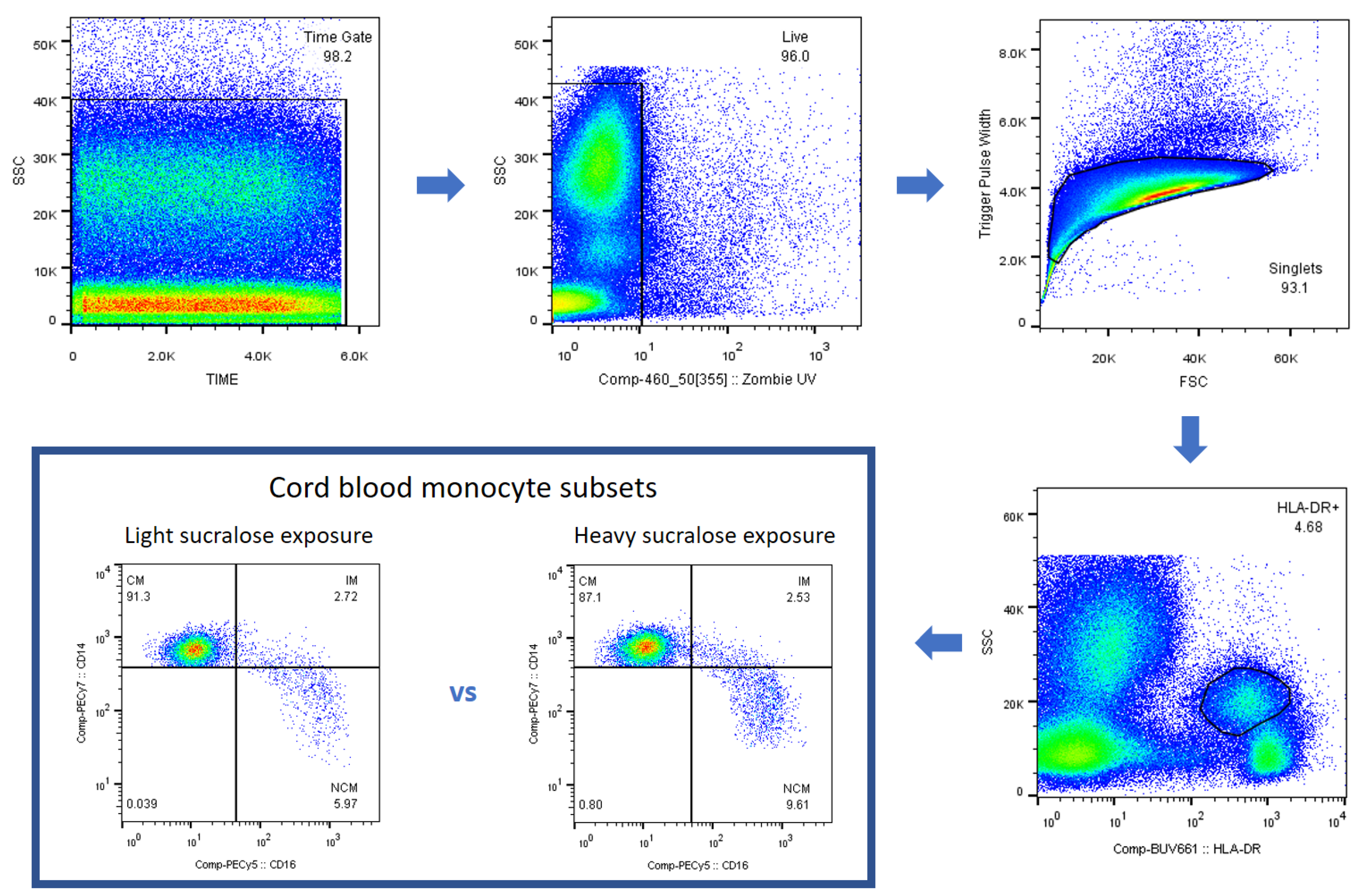
2.5. Immunostaining and Flow Cytometry for Monocyte Subsets
2.6. Gating Strategy for Monocyte Subsets
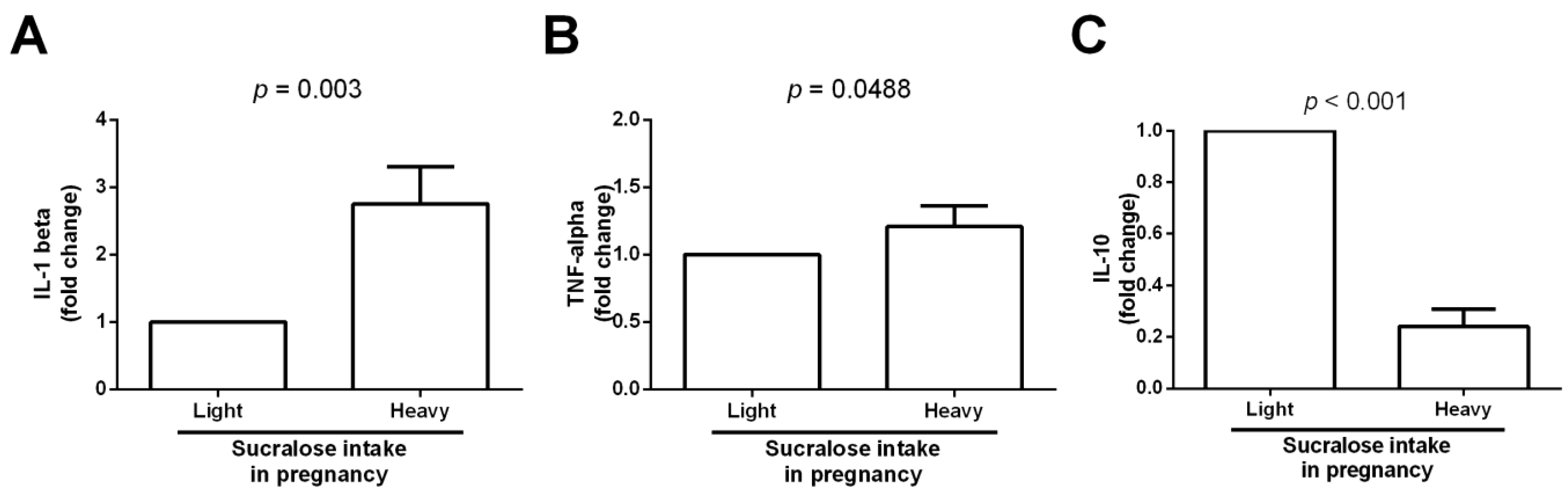
2.7. Umbilical Cord Specimens for IL-1 Beta, TNF-Alpha, and IL-10 mRNA Expression
2.8. Statistics
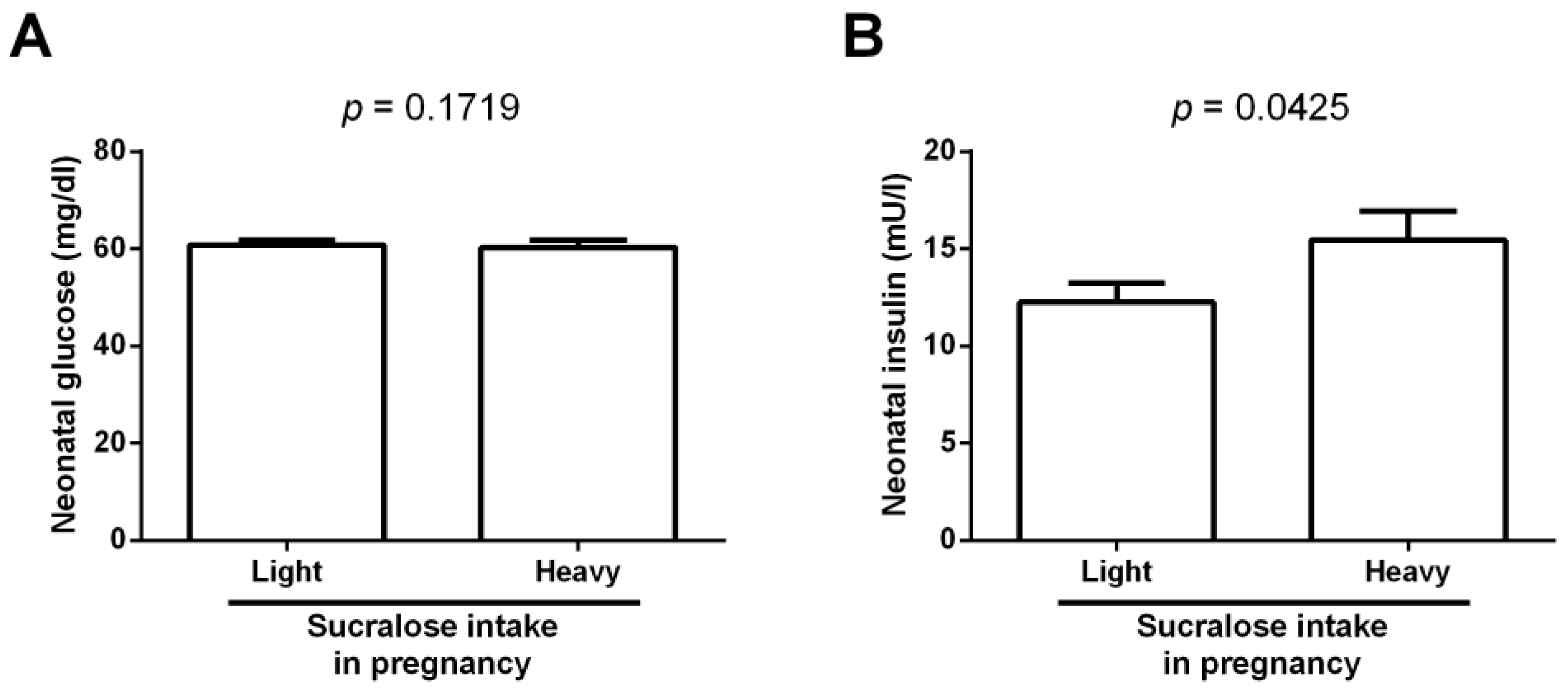
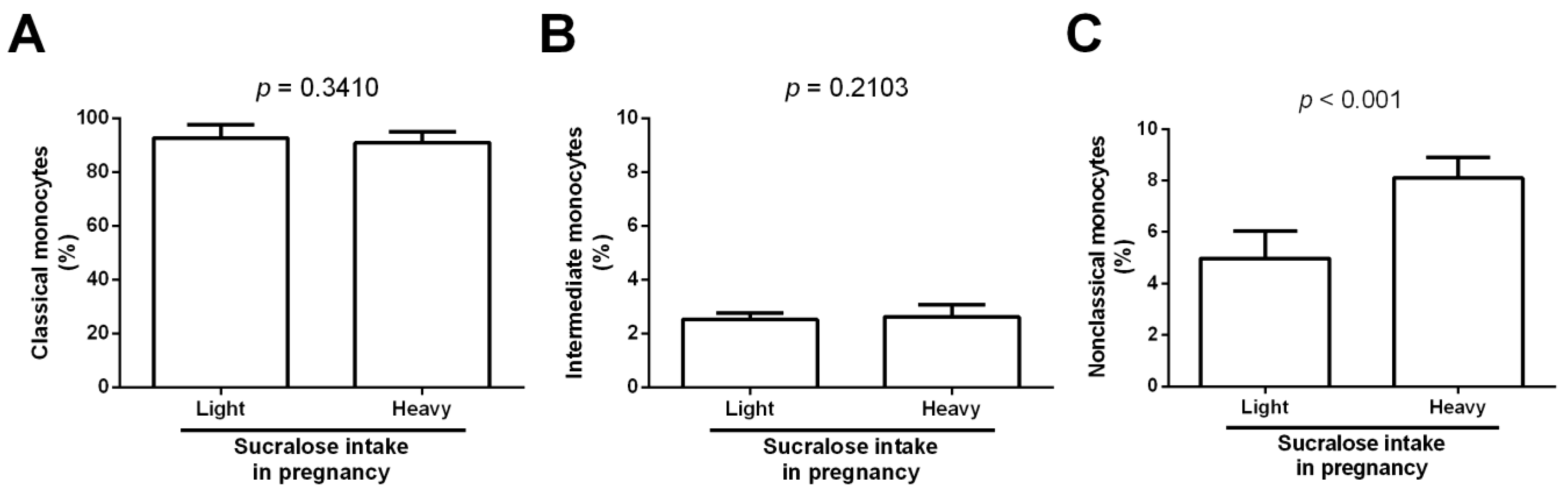
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kelly, T.; Yang, W.; Chen, C.S.; Reynolds, K.; He, J. Global burden of obesity in 2005 and projections to 2030. Int. J. Obes. 2008, 32, 1431–1437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, X.; Xu, Y.; Pan, X.; Xu, J.; Ding, Y.; Sun, X.; Song, X.; Ren, Y.; Shan, P.F. Global, regional, and national burden and trend of diabetes in 195 countries and territories: An analysis from 1990 to 2025. Sci. Rep. 2020, 10, 14790. [Google Scholar] [CrossRef]
- An, R. Prevalence and Trends of Adult Obesity in the US, 1999–2012. ISRN Obes. 2014, 2014, 185132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sylvetsky, A.C.; Jin, Y.; Clark, E.J.; Welsh, J.A.; Rother, K.I.; Talegawkar, S.A. Consumption of Low-Calorie Sweeteners among Children and Adults in the United States. J. Acad. Nutr. Diet. 2017, 117, 441–448. [Google Scholar] [CrossRef]
- Venegas Hargous, C.; Reyes, M.; Smith Taillie, L.; Gonzalez, C.G.; Corvalan, C. Consumption of non-nutritive sweeteners by pre-schoolers of the food and environment Chilean cohort (FECHIC) before the implementation of the Chilean food labelling and advertising law. Nutr. J. 2020, 19, 69. [Google Scholar] [CrossRef] [PubMed]
- Shum, B.; Georgia, S. The Effects of Non-Nutritive Sweetener Consumption in the Pediatric Populations: What We Know, What We Don't, and What We Need to Learn. Front. Endocrinol. 2021, 12, 625415. [Google Scholar] [CrossRef]
- Sylvetsky, A.C.; Rother, K.I. Trends in the consumption of low-calorie sweeteners. Physiol. Behav. 2016, 164, 446–450. [Google Scholar] [CrossRef] [Green Version]
- Fuentealba Arevalo, F.; Espinoza Espinoza, J.; Salazar Ibacahe, C.; Duran Aguero, S. Consumption of non-caloric sweeteners among pregnant Chileans: A cross-sectional study. Nutr. Hosp. 2019, 36, 890–897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palatnik, A.; Moosreiner, A.; Olivier-Van Stichelen, S. Consumption of non-nutritive sweeteners during pregnancy. Am. J. Obstet. Gynecol. 2020, 223, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Archibald, A.J.; Dolinsky, V.W.; Azad, M.B. Early-Life Exposure to Non-Nutritive Sweeteners and the Developmental Origins of Childhood Obesity: Global Evidence from Human and Rodent Studies. Nutrients 2018, 10, 194. [Google Scholar] [CrossRef] [Green Version]
- Araujo, J.R.; Martel, F.; Keating, E. Exposure to non-nutritive sweeteners during pregnancy and lactation: Impact in programming of metabolic diseases in the progeny later in life. Reprod. Toxicol. 2014, 49, 196–201. [Google Scholar] [CrossRef] [PubMed]
- Cai, C.; Sivak, A.; Davenport, M.H. Effects of prenatal artificial sweeteners consumption on birth outcomes: A systematic review and meta-analysis. Public Health Nutr. 2021, 24, 5024–5033. [Google Scholar] [CrossRef]
- Azad, M.B.; Archibald, A.; Tomczyk, M.M.; Head, A.; Cheung, K.G.; de Souza, R.J.; Becker, A.B.; Mandhane, P.J.; Turvey, S.E.; Moraes, T.J.; et al. Nonnutritive sweetener consumption during pregnancy, adiposity, and adipocyte differentiation in offspring: Evidence from humans, mice, and cells. Int. J. Obes. 2020, 44, 2137–2148. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Guo, Z.; Chen, D.; Li, L.; Song, X.; Liu, T.; Jin, G.; Li, Y.; Liu, Y.; Ajiguli, A.; et al. Maternal sucralose intake alters gut microbiota of offspring and exacerbates hepatic steatosis in adulthood. Gut Microbes 2020, 11, 1043–1063. [Google Scholar] [CrossRef]
- Gomez-Arauz, A.Y.; Bueno-Hernandez, N.; Palomera, L.F.; Alcantara-Suarez, R.; De Leon, K.L.; Mendez-Garcia, L.A.; Carrero-Aguirre, M.; Manjarrez-Reyna, A.N.; Martinez-Reyes, C.P.; Esquivel-Velazquez, M.; et al. A Single 48 mg Sucralose Sip Unbalances Monocyte Subpopulations and Stimulates Insulin Secretion in Healthy Young Adults. J. Immunol. Res. 2019, 2019, 6105059. [Google Scholar] [CrossRef]
- Koelman, L.; Pivovarova-Ramich, O.; Pfeiffer, A.F.H.; Grune, T.; Aleksandrova, K. Cytokines for evaluation of chronic inflammatory status in ageing research: Reliability and phenotypic characterisation. Immun. Ageing 2019, 16, 11. [Google Scholar] [CrossRef] [Green Version]
- Khanna, D.; Khanna, S.; Khanna, P.; Kahar, P.; Patel, B.M. Obesity: A Chronic Low-Grade Inflammation and Its Markers. Cureus 2022, 14, e22711. [Google Scholar] [CrossRef] [PubMed]
- Kuczmarski, R.J.; Ogden, C.L.; Guo, S.S.; Grummer-Strawn, L.M.; Flegal, K.M.; Mei, Z.; Wei, R.; Curtin, L.R.; Roche, A.F.; Johnson, C.L. 2000 CDC Growth Charts for the United States: Methods and Development; National Center for Health Statistics: Hyattsville, MD, USA, 2002; Volume 11, pp. 1–190. ISBN 0-8406-0575-7. [Google Scholar]
- Mendoza-Martinez, V.M.; Zavala-Solares, M.R.; Espinosa-Flores, A.J.; Leon-Barrera, K.L.; Alcantara-Suarez, R.; Carrillo-Ruiz, J.D.; Escobedo, G.; Roldan-Valadez, E.; Esquivel-Velazquez, M.; Melendez-Mier, G.; et al. Is a Non-Caloric Sweetener-Free Diet Good to Treat Functional Gastrointestinal Disorder Symptoms? A Randomized Controlled Trial. Nutrients 2022, 14, 1095. [Google Scholar] [CrossRef]
- Maslova, E.; Strom, M.; Olsen, S.F.; Halldorsson, T.I. Consumption of artificially-sweetened soft drinks in pregnancy and risk of child asthma and allergic rhinitis. PLoS ONE 2013, 8, e57261. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Li, X.; Wu, Y.; Su, Q.; Qin, L.; Ma, J. The Associations between Maternal Serum Aspartame and Sucralose and Metabolic Health during Pregnancy. Nutrients 2022, 14, 5001. [Google Scholar] [CrossRef] [PubMed]
- Ayoob, K.T. Consumption of non-nutritive sweeteners during pregnancy. Am. J. Obstet. Gynecol. 2020, 223, 952–953. [Google Scholar] [CrossRef]
- Azad, M.B.; Sharma, A.K.; de Souza, R.J.; Dolinsky, V.W.; Becker, A.B.; Mandhane, P.J.; Turvey, S.E.; Subbarao, P.; Lefebvre, D.L.; Sears, M.R.; et al. Association Between Artificially Sweetened Beverage Consumption During Pregnancy and Infant Body Mass Index. JAMA Pediatr. 2016, 170, 662–670. [Google Scholar] [CrossRef]
- Briones-Avila, L.S.; Moranchel-Hernandez, M.A.; Moreno-Riolobos, D.; Silva Pereira, T.S.; Ortega Regules, A.E.; Villasenor Lopez, K.; Islas Romero, L.M. Analysis of Caloric and Noncaloric Sweeteners Present in Dairy Products Aimed at the School Market and Their Possible Effects on Health. Nutrients 2021, 13, 2994. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.T.; Lin, C.H.; Pai, H.L.; Chen, Y.C.; Cheng, K.P.; Kuo, H.Y.; Li, C.H.; Ou, H.Y. Sucralose, a Non-nutritive Artificial Sweetener Exacerbates High Fat Diet-Induced Hepatic Steatosis Through Taste Receptor Type 1 Member 3. Front. Nutr. 2022, 9, 823723. [Google Scholar] [CrossRef] [PubMed]
- Lizunkova, P.; Enuwosa, E.; Chichger, H. Activation of the sweet taste receptor T1R3 by sucralose attenuates VEGF-induced vasculogenesis in a cell model of the retinal microvascular endothelium. Graefes Arch. Clin. Exp. Ophthalmol. 2019, 257, 71–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pepino, M.Y.; Tiemann, C.D.; Patterson, B.W.; Wice, B.M.; Klein, S. Sucralose affects glycemic and hormonal responses to an oral glucose load. Diabetes Care 2013, 36, 2530–2535. [Google Scholar] [CrossRef] [Green Version]
- Lertrit, A.; Srimachai, S.; Saetung, S.; Chanprasertyothin, S.; Chailurkit, L.O.; Areevut, C.; Katekao, P.; Ongphiphadhanakul, B.; Sriphrapradang, C. Effects of sucralose on insulin and glucagon-like peptide-1 secretion in healthy subjects: A randomized, double-blind, placebo-controlled trial. Nutrition 2018, 55–56, 125–130. [Google Scholar] [CrossRef]
- Ahmad, S.Y.; Friel, J.K.; Mackay, D.S. Effect of sucralose and aspartame on glucose metabolism and gut hormones. Nutr. Rev. 2020, 78, 725–746. [Google Scholar] [CrossRef]
- Nakagawa, Y.; Nagasawa, M.; Yamada, S.; Hara, A.; Mogami, H.; Nikolaev, V.O.; Lohse, M.J.; Shigemura, N.; Ninomiya, Y.; Kojima, I. Sweet taste receptor expressed in pancreatic beta-cells activates the calcium and cyclic AMP signaling systems and stimulates insulin secretion. PLoS ONE 2009, 4, e5106. [Google Scholar] [CrossRef] [Green Version]
- Rosales-Gomez, C.A.; Martinez-Carrillo, B.E.; Resendiz-Albor, A.A.; Ramirez-Duran, N.; Valdes-Ramos, R.; Mondragon-Velasquez, T.; Escoto-Herrera, J.A. Chronic Consumption of Sweeteners and Its Effect on Glycaemia, Cytokines, Hormones, and Lymphocytes of GALT in CD1 Mice. Biomed. Res. Int. 2018, 2018, 1345282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romo-Romo, A.; Aguilar-Salinas, C.A.; Brito-Cordova, G.X.; Gomez-Diaz, R.A.; Almeda-Valdes, P. Sucralose decreases insulin sensitivity in healthy subjects: A randomized controlled trial. Am. J. Clin. Nutr. 2018, 108, 485–491. [Google Scholar] [CrossRef] [PubMed]
- Bueno-Hernandez, N.; Esquivel-Velazquez, M.; Alcantara-Suarez, R.; Gomez-Arauz, A.Y.; Espinosa-Flores, A.J.; de Leon-Barrera, K.L.; Mendoza-Martinez, V.M.; Sanchez Medina, G.A.; Leon-Hernandez, M.; Ruiz-Barranco, A.; et al. Chronic sucralose consumption induces elevation of serum insulin in young healthy adults: A randomized, double blind, controlled trial. Nutr. J. 2020, 19, 32. [Google Scholar] [CrossRef] [Green Version]
- Dalenberg, J.R.; Patel, B.P.; Denis, R.; Veldhuizen, M.G.; Nakamura, Y.; Vinke, P.C.; Luquet, S.; Small, D.M. Short-Term Consumption of Sucralose with, but Not without, Carbohydrate Impairs Neural and Metabolic Sensitivity to Sugar in Humans. Cell Metab. 2020, 31, 493–502.e497. [Google Scholar] [CrossRef]
- Halasa, B.C.; Sylvetsky, A.C.; Conway, E.M.; Shouppe, E.L.; Walter, M.F.; Walter, P.J.; Cai, H.; Hui, L.; Rother, K.I. Non-Nutritive Sweeteners in Human Amniotic Fluid and Cord Blood: Evidence of Transplacental Fetal Exposure. Am. J. Perinatol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Knight, J.; Jiang, J.; Wood, K.D.; Holmes, R.P.; Assimos, D.G. Oxalate and sucralose absorption in idiopathic calcium oxalate stone formers. Urology 2011, 78, 475–479.e13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soffritti, M.; Padovani, M.; Tibaldi, E.E.A.; Falcioni, L.; Manservisi, F.; Lauriola, M.; Bua, L.; Manservigi, M.; Belpoggi, F. Sucralose administered in feed, beginning prenatally through lifespan, induces hematopoietic neoplasias in male swiss mice. Int. J. Occup. Environ. Health 2016, 22, 7–17. [Google Scholar] [CrossRef] [Green Version]
- Wood, S.G.; John, B.A.; Hawkins, D.R. The pharmacokinetics and metabolism of sucralose in the dog. Food Chem. Toxicol. 2000, 38 (Suppl. S2), S99–S106. [Google Scholar] [CrossRef]
- Kapellos, T.S.; Bonaguro, L.; Gemund, I.; Reusch, N.; Saglam, A.; Hinkley, E.R.; Schultze, J.L. Human Monocyte Subsets and Phenotypes in Major Chronic Inflammatory Diseases. Front. Immunol. 2019, 10, 2035. [Google Scholar] [CrossRef] [Green Version]
- Devevre, E.F.; Renovato-Martins, M.; Clement, K.; Sautes-Fridman, C.; Cremer, I.; Poitou, C. Profiling of the three circulating monocyte subpopulations in human obesity. J. Immunol. 2015, 194, 3917–3923. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, R.; Kanti Barman, P.; Kumar Thatoi, P.; Tripathy, R.; Kumar Das, B.; Ravindran, B. Non-Classical monocytes display inflammatory features: Validation in Sepsis and Systemic Lupus Erythematous. Sci. Rep. 2015, 5, 13886. [Google Scholar] [CrossRef] [Green Version]
- Ong, S.M.; Teng, K.; Newell, E.; Chen, H.; Chen, J.; Loy, T.; Yeo, T.W.; Fink, K.; Wong, S.C. A Novel, Five-Marker Alternative to CD16-CD14 Gating to Identify the Three Human Monocyte Subsets. Front. Immunol. 2019, 10, 1761. [Google Scholar] [CrossRef] [Green Version]
- Black, A.R.; Black, J.D.; Azizkhan-Clifford, J. Sp1 and kruppel-like factor family of transcription factors in cell growth regulation and cancer. J. Cell Physiol. 2001, 188, 143–160. [Google Scholar] [CrossRef]
- Resendes, K.K.; Rosmarin, A.G. Sp1 control of gene expression in myeloid cells. Crit. Rev. Eukaryot. Gene Expr. 2004, 14, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Masubuchi, Y.; Nakagawa, Y.; Medina, J.; Nagasawa, M.; Kojima, I.; Rasenick, M.M.; Inagaki, T.; Shibata, H. T1R3 homomeric sweet taste receptor regulates adipogenesis through Galphas-mediated microtubules disassembly and Rho activation in 3T3-L1 cells. PLoS ONE 2017, 12, e0176841. [Google Scholar] [CrossRef]
- Daniels, M.J.; Brough, D. Unconventional Pathways of Secretion Contribute to Inflammation. Int. J. Mol. Sci. 2017, 18, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, P.; Hui, X.; Zheng, Q.; Gao, Y.; Jin, L.; Jiang, W.; Zhou, C.; Liu, T.; Huang, Y.; Liu, Q.; et al. Mitochondrial uncoupling protein 1 antagonizes atherosclerosis by blocking NLRP3 inflammasome-dependent interleukin-1beta production. Sci. Adv. 2021, 7, eabl4024. [Google Scholar] [CrossRef] [PubMed]
- Alzamil, H. Elevated Serum TNF-alpha Is Related to Obesity in Type 2 Diabetes Mellitus and Is Associated with Glycemic Control and Insulin Resistance. J. Obes. 2020, 2020, 5076858. [Google Scholar] [CrossRef] [Green Version]
- Mendez-Garcia, L.A.; Trejo-Millan, F.; Martinez-Reyes, C.P.; Manjarrez-Reyna, A.N.; Esquivel-Velazquez, M.; Melendez-Mier, G.; Islas-Andrade, S.; Rojas-Bernabe, A.; Kzhyshkowska, J.; Escobedo, G. Infliximab ameliorates tumor necrosis factor-alpha-induced insulin resistance by attenuating PTP1B activation in 3T3L1 adipocytes in vitro. Scand. J. Immunol. 2018, 88, e12716. [Google Scholar] [CrossRef]
- Mohallem, R.; Aryal, U.K. Regulators of TNFalpha mediated insulin resistance elucidated by quantitative proteomics. Sci. Rep. 2020, 10, 20878. [Google Scholar] [CrossRef]
- Khan, M.A.; Ashoor, G.A.; Shamma, T.; Alanazi, F.; Altuhami, A.; Kazmi, S.; Ahmed, H.A.; Mohammed Assiri, A.; Clemens Broering, D. IL-10 Mediated Immunomodulation Limits Subepithelial Fibrosis and Repairs Airway Epithelium in Rejecting Airway Allografts. Cells 2021, 10, 1248. [Google Scholar] [CrossRef] [PubMed]
- Saraiva, M.; Vieira, P.; O’Garra, A. Biology and therapeutic potential of interleukin-10. J. Exp. Med. 2020, 217, e20190418. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Gao, T.; Li, Y.; Xie, Y.; Zeng, S.; Tai, C.; Feng, Y.; Shen, P.; Wang, B. A long-term anti-inflammation markedly alleviated high-fat diet-induced obesity by repeated administrations of overexpressing IL10 human umbilical cord-derived mesenchymal stromal cells. Stem Cell Res. Ther. 2022, 13, 259. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.H.; Park, H.S.; Kim, K.S.; Choi, W.H.; Ahn, C.W.; Kim, B.T.; Kim, S.M.; Lee, S.Y.; Ahn, S.M.; Kim, Y.K.; et al. Effect of weight loss on some serum cytokines in human obesity: Increase in IL-10 after weight loss. J. Nutr. Biochem. 2008, 19, 371–375. [Google Scholar] [CrossRef] [PubMed]
- Simmonds, M.; Llewellyn, A.; Owen, C.G.; Woolacott, N. Predicting adult obesity from childhood obesity: A systematic review and meta-analysis. Obes. Rev. 2016, 17, 95–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbasi, A.; Juszczyk, D.; van Jaarsveld, C.H.M.; Gulliford, M.C. Body Mass Index and Incident Type 1 and Type 2 Diabetes in Children and Young Adults: A Retrospective Cohort Study. J. Endocr. Soc. 2017, 1, 524–537. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sucralose Intake in Pregnancy | ||||
|---|---|---|---|---|
| Characteristics | Light | Heavy | p Value | |
| Mothers | Number of participants | 205 | 87 | - |
| Age (years) | 25.9 ± 5.3 | 26.1 ± 3.3 | 0.4675 | |
| SBP (mmHg) | 106.7 ± 11.5 | 106.9 ± 10.3 | 0.4869 | |
| DBP (mmHg) | 70.4 ± 9.7 | 70.6 ± 9.0 | 0.4827 | |
| Pregestational BMI (kg/m2) | 27.2 ± 4.3 | 27.4 ± 4.6 | 0.4591 | |
| Previous pregnancies | 1.6 ± 0.8 | 1.7 ± 0.8 | 0.3945 | |
| Age of pregnancy (weeks) | 39.1 ± 1.2 | 39.2 ± 1.3 | 0.4411 | |
| OGTT blood levels at 0′ (mg/dL) | 89.9 ± 6.1 | 90.9 ± 5.6 | 0.3545 | |
| OGTT blood levels at 30′ (mg/dL) | 122.3 ± 10.2 | 123.5 ± 9.3 | 0.3936 | |
| OGTT blood levels at 60′ (mg/dL) | 101.6 ± 31.1 | 99.0 ± 17.3 | 0.4101 | |
| OGTT blood levels at 90′ (mg/dL) | 93.0 ± 12.2 | 94.2 ± 13.7 | 0.4196 | |
| OGTT blood levels at 120′ (mg/dL) | 97.4 ± 14.6 | 97.1 ± 14.3 | 0.4818 | |
| Serum insulin concentration (mU/L) | 10.2 ± 4.2 | 13.2 ± 4.5 | 0.0618 | |
| Serum sucralose concentration (ng/mL) | 3.1 ± 1.4 | 25.4 ± 4.2 | <0.0001 * | |
| Number of SCP eaten or drunk per week | 2.0 ± 1.6 | 23.3 ± 1.5 | <0.0001 * | |
| Type of SCP more often consumed | Yogurt, diet sodas, candy, baked goods, gelatin | Yogurt, diet sodas, candy, baked goods, jams, gelatin | - | |
| Neonates | Sex (f/m) | 109/96 | 48/39 | 0.7981 |
| Birth weight (kg) | 2.8 ± 0.1 | 3.2 ± 0.6 | 0.0005 * | |
| Neonates above the 95th percentile (n) | 15 | 13 | 0.0470 * | |
| Height (cm) | 48.6 ± 2.4 | 50.1 ± 2.0 | 0.1044 | |
| Capurro index (weeks) | 39.3 ± 1.6 | 39.3 ± 1.5 | 0.9326 | |
| Apgar score | 8.9 ± 0.3 | 8.7 ± 0.4 | 0.2221 | |
| Maternal Variables | β | SE | T | p Value |
|---|---|---|---|---|
| Age | −0.165 | 1.124 | −0.132 | 0.880 |
| SBP | 0.878 | 0.679 | 1.030 | 0.772 |
| DBP | 0.896 | 1.826 | 0.636 | 0.523 |
| Pregestational BMI | 1.857 | 3.751 | 0.347 | 0.712 |
| Previous pregnancies | −0.647 | 0.902 | −0.651 | 0.503 |
| Age of pregnancy (weeks) | 0.023 | 0.018 | 0.347 | 0.764 |
| Serum insulin concentration (mU/L) | 8.250 | 3.720 | 2.328 | 0.061 |
| Serum sucralose concentration (ng/mL) | 0.376 | 0.571 | 0.714 | 0.520 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aguayo-Guerrero, J.A.; Méndez-García, L.A.; Manjarrez-Reyna, A.N.; Esquivel-Velázquez, M.; León-Cabrera, S.; Meléndez, G.; Zambrano, E.; Ramos-Martínez, E.; Fragoso, J.M.; Briones-Garduño, J.C.; et al. Newborns from Mothers Who Intensely Consumed Sucralose during Pregnancy Are Heavier and Exhibit Markers of Metabolic Alteration and Low-Grade Systemic Inflammation: A Cross-Sectional, Prospective Study. Biomedicines 2023, 11, 650. https://doi.org/10.3390/biomedicines11030650
Aguayo-Guerrero JA, Méndez-García LA, Manjarrez-Reyna AN, Esquivel-Velázquez M, León-Cabrera S, Meléndez G, Zambrano E, Ramos-Martínez E, Fragoso JM, Briones-Garduño JC, et al. Newborns from Mothers Who Intensely Consumed Sucralose during Pregnancy Are Heavier and Exhibit Markers of Metabolic Alteration and Low-Grade Systemic Inflammation: A Cross-Sectional, Prospective Study. Biomedicines. 2023; 11(3):650. https://doi.org/10.3390/biomedicines11030650
Chicago/Turabian StyleAguayo-Guerrero, José Alfredo, Lucía Angélica Méndez-García, Aarón Noe Manjarrez-Reyna, Marcela Esquivel-Velázquez, Sonia León-Cabrera, Guillermo Meléndez, Elena Zambrano, Espiridión Ramos-Martínez, José Manuel Fragoso, Juan Carlos Briones-Garduño, and et al. 2023. "Newborns from Mothers Who Intensely Consumed Sucralose during Pregnancy Are Heavier and Exhibit Markers of Metabolic Alteration and Low-Grade Systemic Inflammation: A Cross-Sectional, Prospective Study" Biomedicines 11, no. 3: 650. https://doi.org/10.3390/biomedicines11030650
APA StyleAguayo-Guerrero, J. A., Méndez-García, L. A., Manjarrez-Reyna, A. N., Esquivel-Velázquez, M., León-Cabrera, S., Meléndez, G., Zambrano, E., Ramos-Martínez, E., Fragoso, J. M., Briones-Garduño, J. C., & Escobedo, G. (2023). Newborns from Mothers Who Intensely Consumed Sucralose during Pregnancy Are Heavier and Exhibit Markers of Metabolic Alteration and Low-Grade Systemic Inflammation: A Cross-Sectional, Prospective Study. Biomedicines, 11(3), 650. https://doi.org/10.3390/biomedicines11030650