
Streptomyces Bioactive Metabolites Prevent Liver Cancer through Apoptosis, Inhibiting Oxidative Stress and Inflammatory Markers in Diethylnitrosamine-Induced Hepatocellular Carcinoma


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of the Bacterial Extract
2.2. Gas Chromatography–Mass Spectrometry (GC-MS) Analysis for the Chemical Constituents of the Streptomyces Extract
2.3. Determination of Cell Viability by MTT Assay
2.4. Animals and Chemicals
2.5. General Experimental Procedures
2.6. The Toxicity Test for the Natural Extract
2.7. Sampling and Biochemical Analysis
2.8. Oxidative Stress and Antioxidant Markers in Liver Homogenate
2.9. Analysis of Gene Expression: Quantitative PCR (RT-qPCR)
2.10. Histopathological Investigations
2.11. Immunohistochemistry Procedure of Paraffin Sections
2.12. Statistical Analysis
3. Results
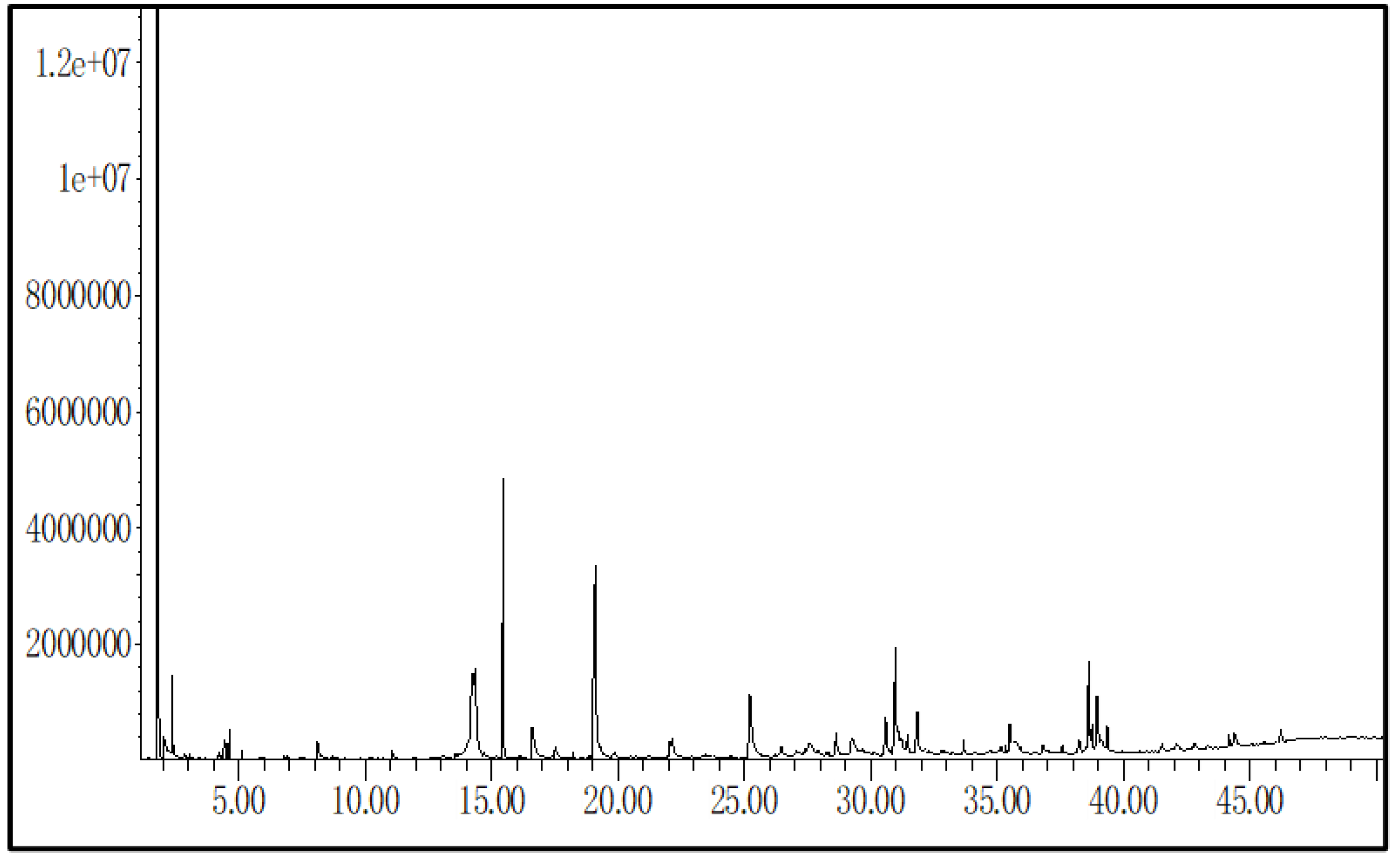
3.1. Gas Chromatography–Mass Spectrometry (GC–MS) Analysis of Streptomyces sp. (A16) Extract
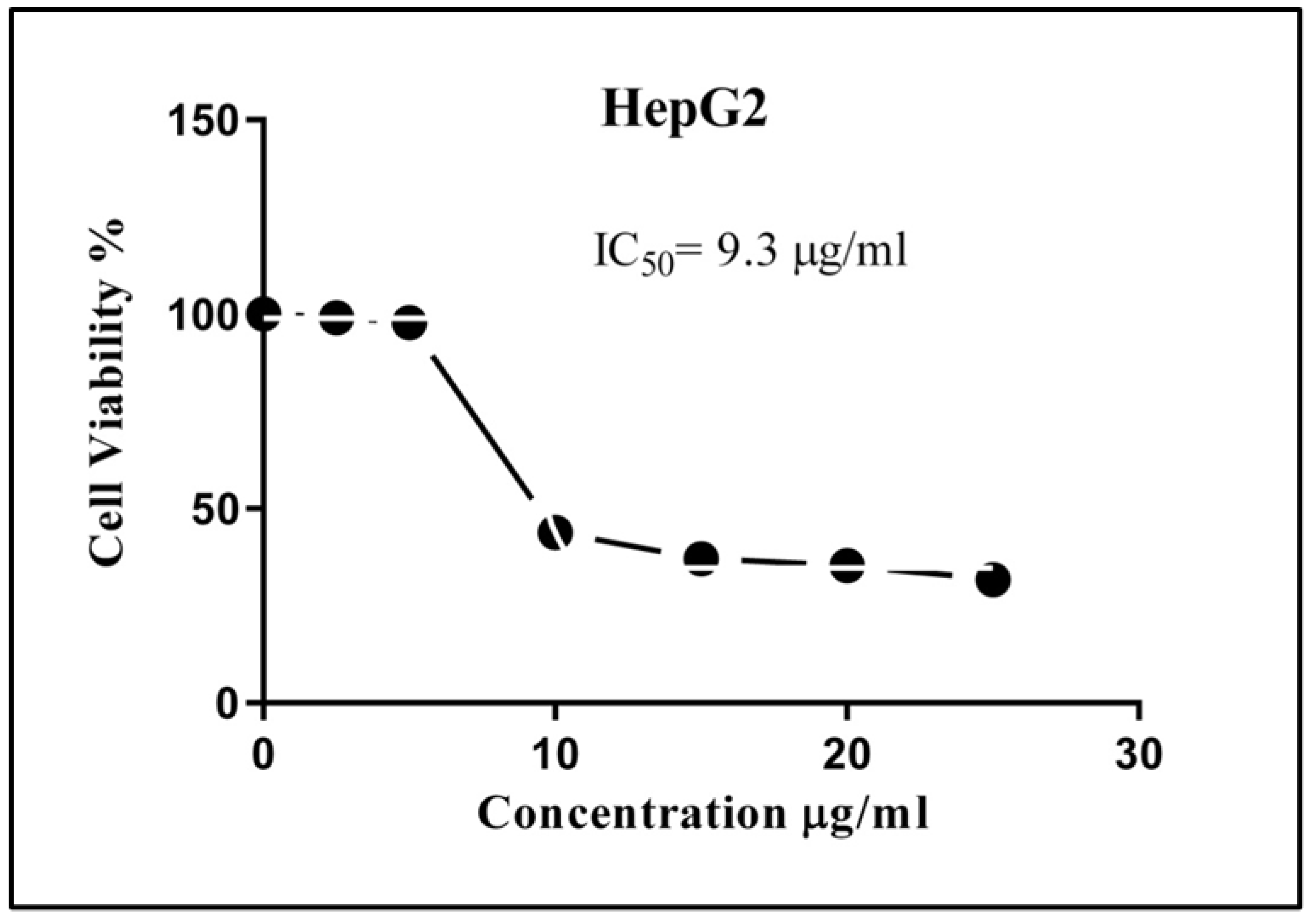
3.2. Antiproliferative Effects of Streptomyces sp. (A16) HepG2 Cells Viability
3.3. Acute Toxicity Test
3.4. The Impact of Streptomyces sp. (A16) Extract on the Architecture, Weight, and Serum AST and ALT Levels of the Liver Tissues in DEN-Induced Hepatocarcinogenesis
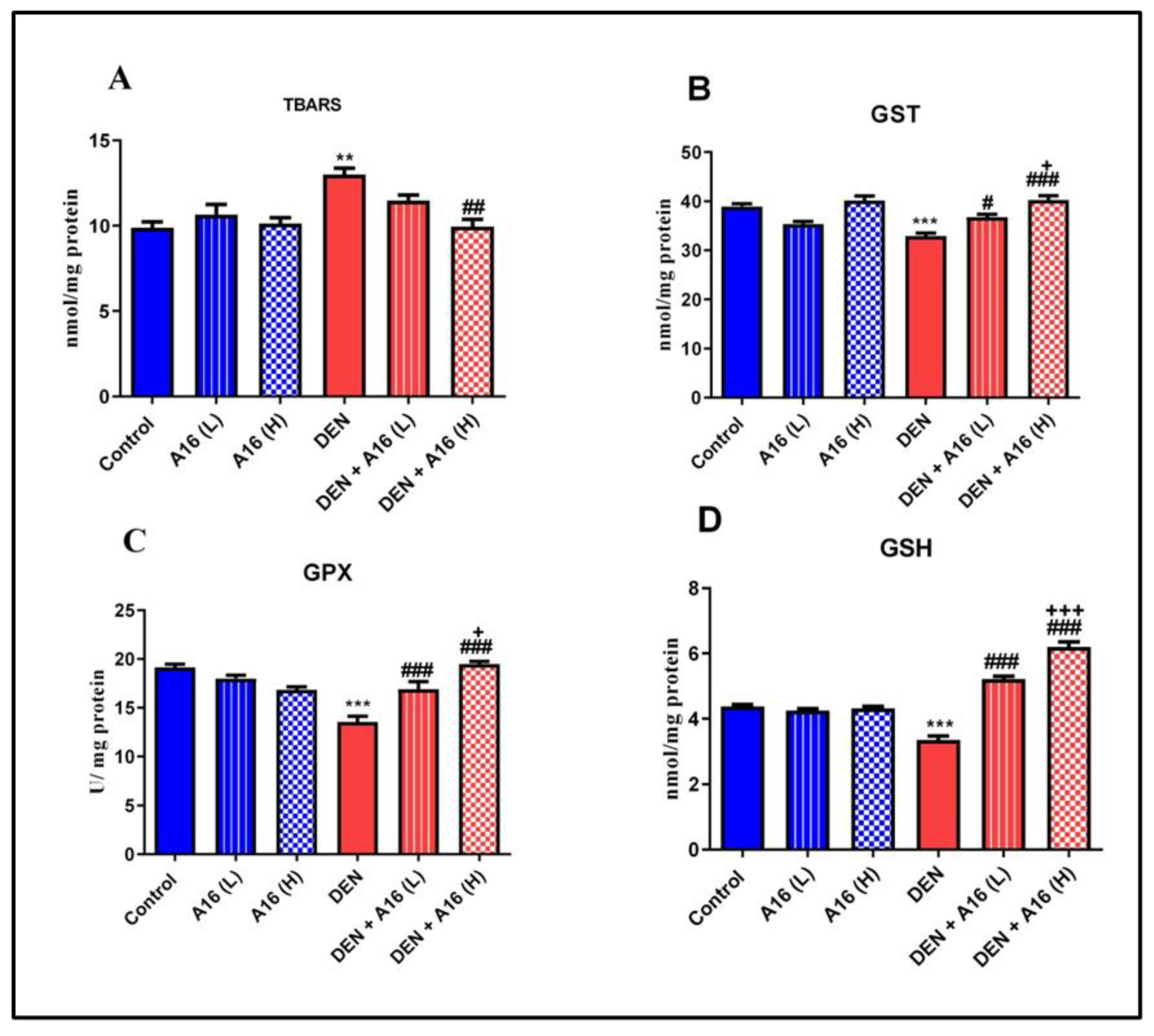
3.5. Effects of Streptomyces sp. Extract on Oxidative Stress and Antioxidant Defense Markers in DEN-Treated Mice
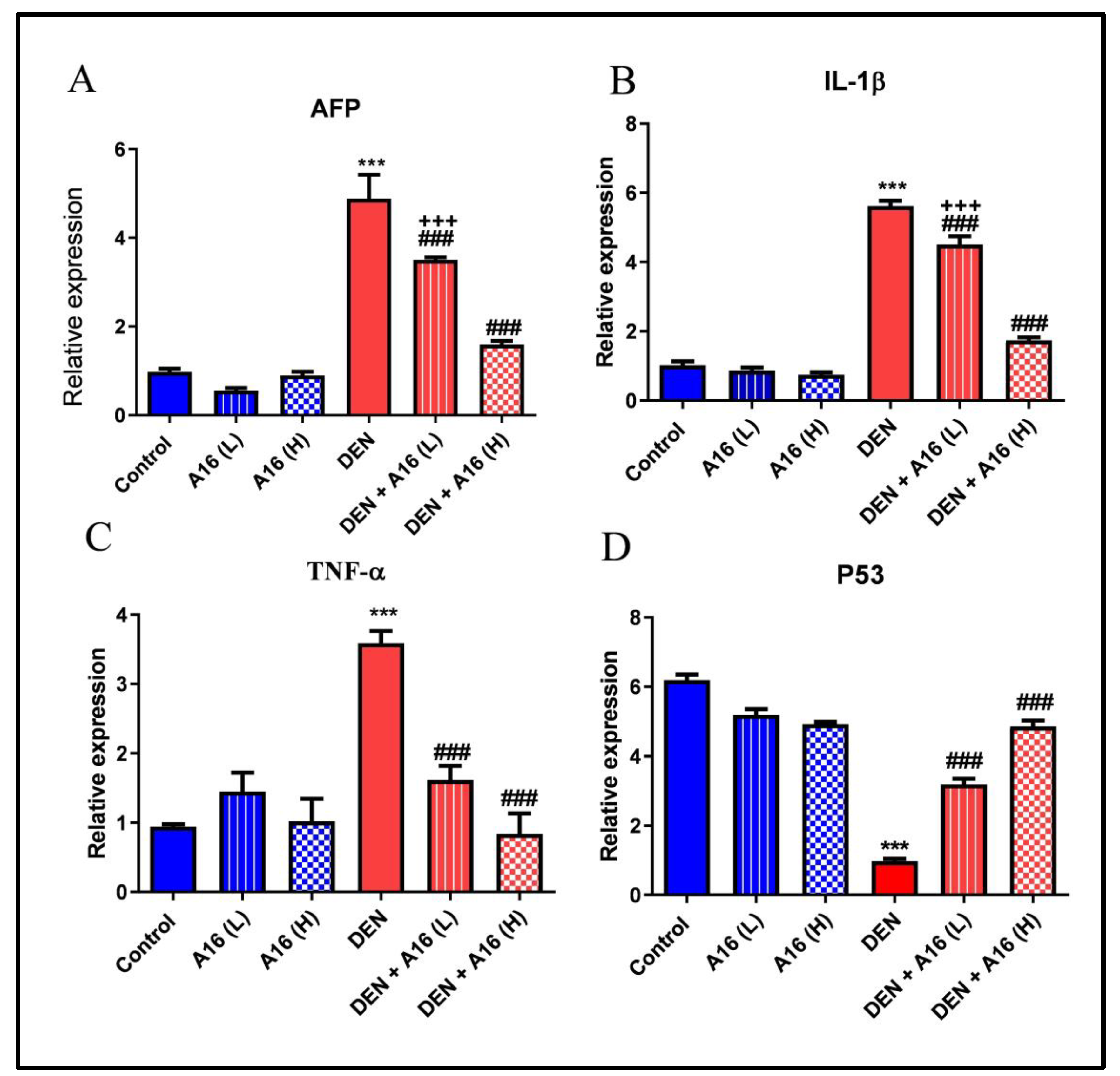
3.6. Effects of Streptomyces sp.(A16) Extract on mRNA Expressions of AFP, IL-1β, TNF-α and P53 in DEN-Induced Hepatocarcinogenesis
3.7. Effects of Streptomyces sp.(A16) Extract on Pathological Changes in DEN-Induced Hepatocarcinogenesis
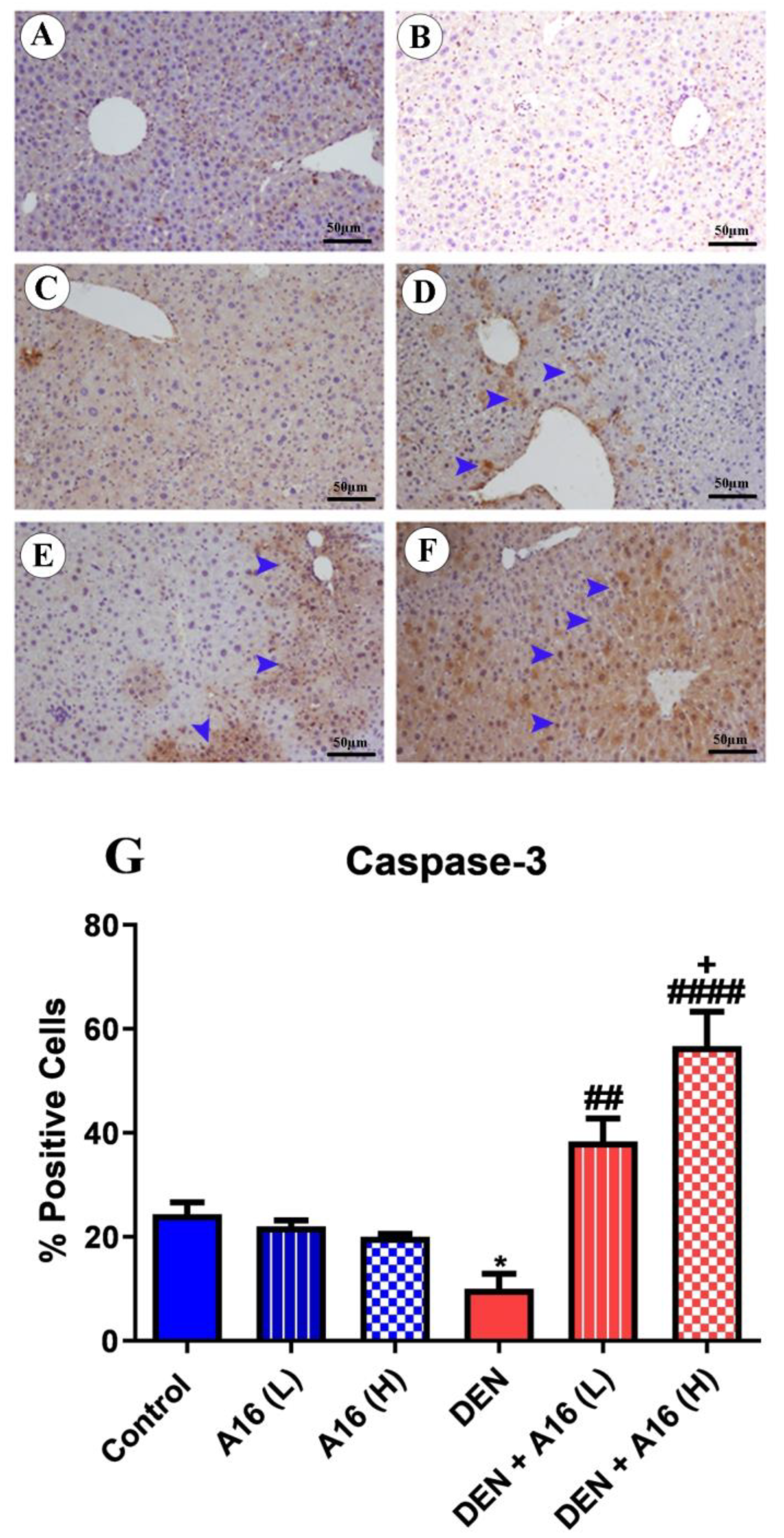
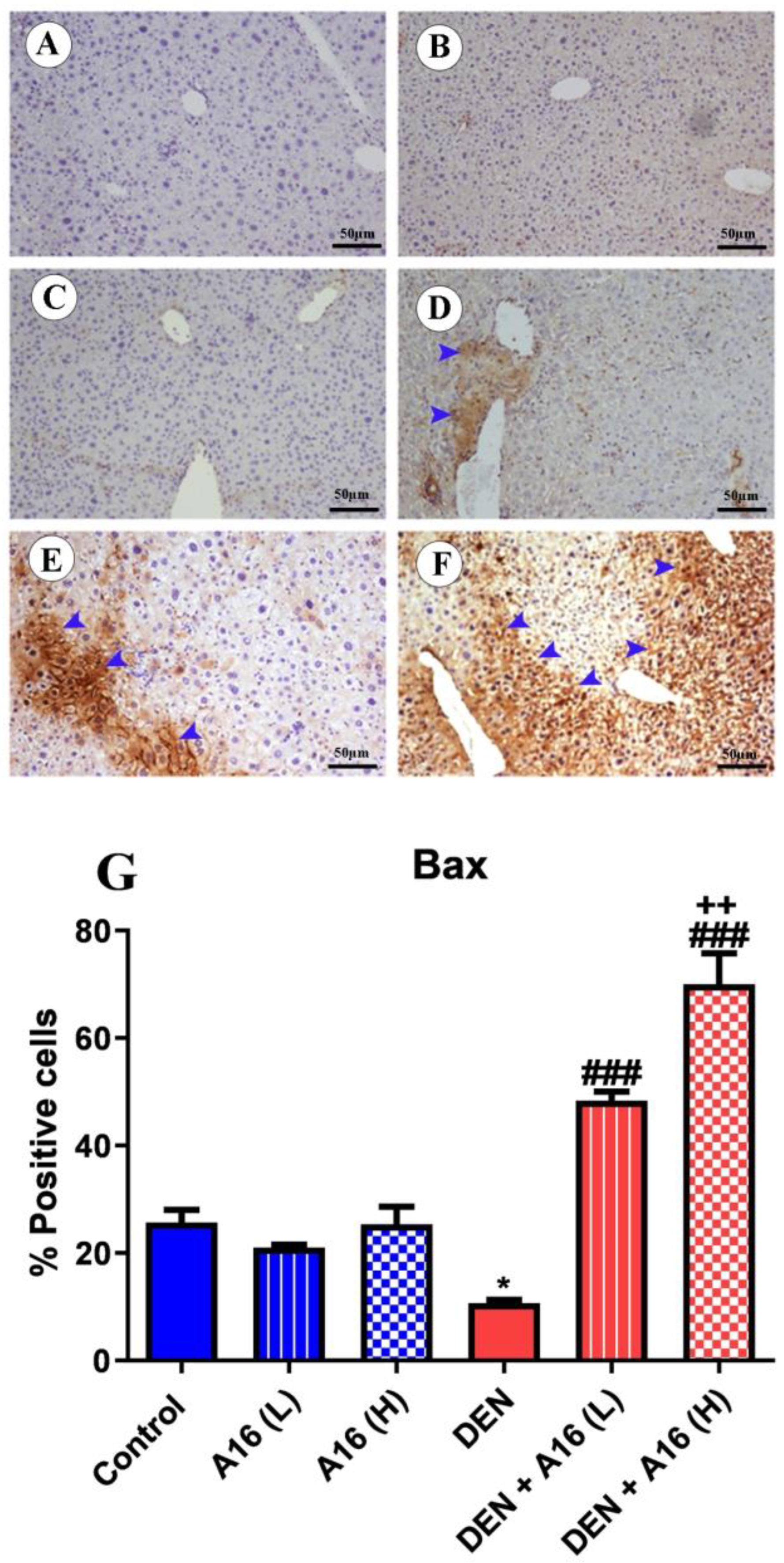
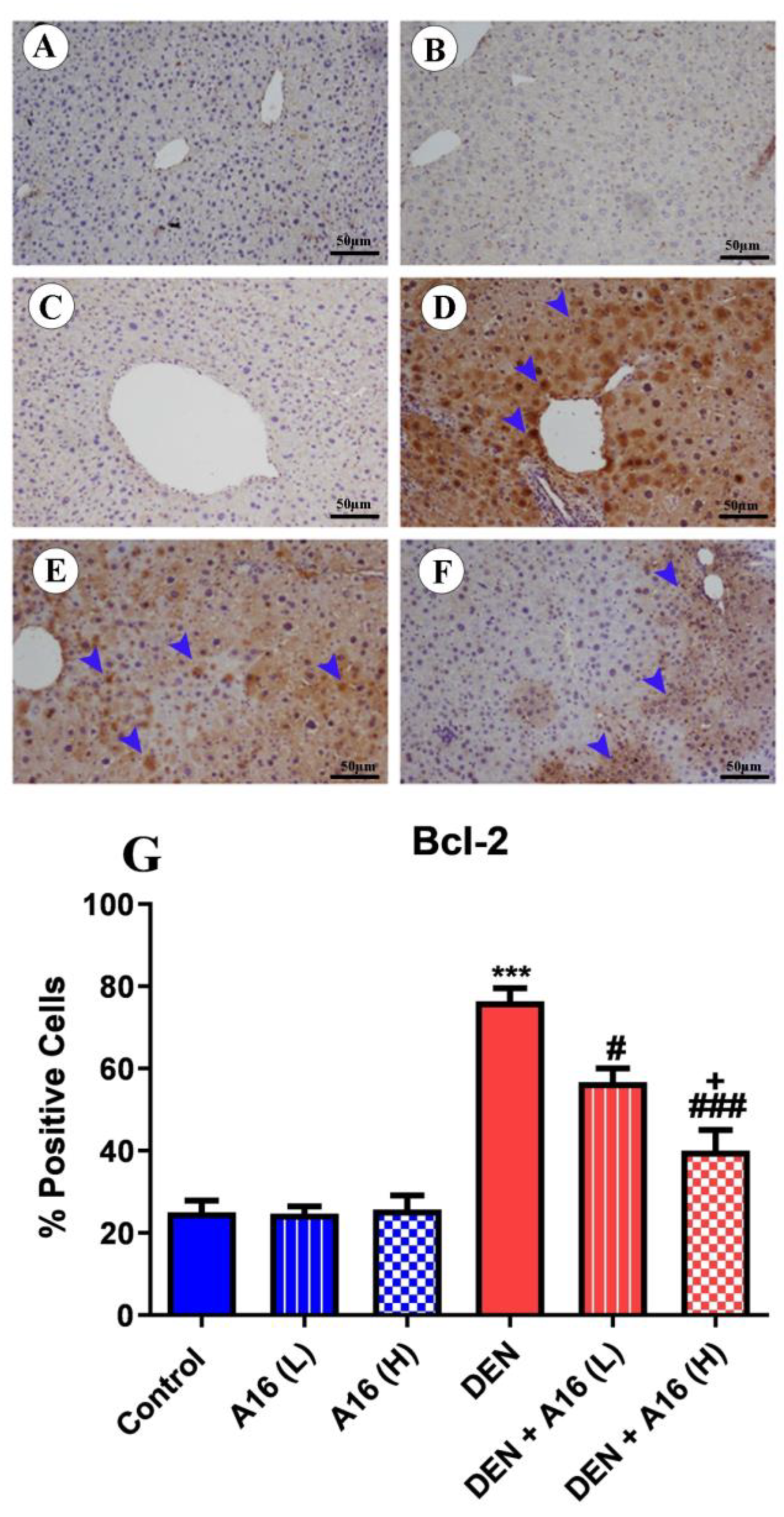
3.8. Immunohistochemical Analysis of Bcl2, Bax, and Caspase-3 after Streptomyces sp.(A16) Extract Treatment of DEN-Induced Hepatocarcinogenesis
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- EASL. EASL Clinical Practice Guidelines: Management of hepatocellular carcinoma. J. Hepatol. 2018, 69, 182–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Nakeep, S. Molecular and genetic markers in hepatocellular carcinoma: In silico analysis to clinical validation (current limitations and future promises). World J. Gastrointest. Pathophysiol. 2022, 13, 1. [Google Scholar] [CrossRef] [PubMed]
- Zajkowska, M.; Mroczko, B. Chemokines in Primary Liver Cancer. Int. J. Mol. Sci. 2022, 23, 8846. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, N.; Friedman, S.L.; Goossens, N.; Hoshida, Y. Risk factors and prevention of hepatocellular carcinoma in the era of precision medicine. J. Hepatol. 2018, 68, 526–549. [Google Scholar] [CrossRef] [Green Version]
- Poustchi, H.; Sepanlou, S.; Esmaili, S.; Mehrabi, N.; Ansarymoghadam, A. Hepatocellular carcinoma in the world and the middle East. Middle East J. Dig. Dis. 2010, 2, 31–41. [Google Scholar]
- Brown, Z.J.; Heinrich, B.; Greten, T.F. Mouse models of hepatocellular carcinoma: An overview and highlights for immunotherapy research. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 536–554. [Google Scholar] [CrossRef] [PubMed]
- Márquez-Quiroga, L.V.; Arellanes-Robledo, J.; Vásquez-Garzón, V.R.; Villa-Treviño, S.; Muriel, P. Models of nonalcoholic steatohepatitis potentiated by chemical inducers leading to hepatocellular carcinoma. Biochem. Pharmacol. 2022, 195, 114845. [Google Scholar] [CrossRef]
- Verna, L.; Whysner, J.; Williams, G.M. N-nitrosodiethylamine mechanistic data and risk assessment: Bioactivation, DNA-adduct formation, mutagenicity, and tumor initiation. Pharmacol. Ther. 1996, 71, 57–81. [Google Scholar] [CrossRef]
- Xu, X.; Lei, Y.; Chen, L.; Zhou, H.; Liu, H.; Jiang, J.; Yang, Y.; Wu, B. Phosphorylation of NF-κBp65 drives inflammation-mediated hepatocellular carcinogenesis and is a novel therapeutic target. J. Exp. Clin. Cancer Res. 2021, 40, 253. [Google Scholar] [CrossRef]
- Naugler, W.E.; Sakurai, T.; Kim, S.; Maeda, S.; Kim, K.; Elsharkawy, A.M.; Karin, M. Gender disparity in liver cancer due to sex differences in MyD88-dependent IL-6 production. Science 2007, 317, 121–124. [Google Scholar] [CrossRef] [Green Version]
- Carneiro, B.A.; El-Deiry, W.S. Targeting apoptosis in cancer therapy. Nat. Rev. Clin. Oncol. 2020, 17, 395–417. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Letai, A.; Sarosiek, K. Regulation of apoptosis in health and disease: The balancing act of BCL-2 family proteins. Nat. Rev. Mol. Cell Biol. 2019, 20, 175–193. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.W.; Talati, C.; Kim, R. Hepatocellular carcinoma (HCC): Beyond sorafenib—Chemotherapy. J. Gastrointest. Oncol. 2017, 8, 256. [Google Scholar] [CrossRef] [Green Version]
- Sioka, C.; Kyritsis, A.P. Central and peripheral nervous system toxicity of common chemotherapeutic agents. Cancer Chemother. Pharmacol. 2009, 63, 761–767. [Google Scholar] [CrossRef]
- Donald, L.; Pipite, A.; Subramani, R.; Owen, J.; Keyzers, R.A.; Taufa, T. Streptomyces: Still the biggest producer of new natural secondary metabolites, a current perspective. Microbiol. Res. 2022, 13, 418–465. [Google Scholar] [CrossRef]
- Elsayed, T.R.; Galil, D.F.; Sedik, M.Z.; Hassan, H.; Sadik, M.W. Antimicrobial and anticancer activities of actinomycetes isolated from Egyptian soils. Int. J. Curr. Microbiol. Appl. Sci. 2020, 9, 2020. [Google Scholar] [CrossRef]
- Balachandran, C.; Sangeetha, B.; Duraipandiyan, V.; Raj, M.K.; Ignacimuthu, S.; Al-Dhabi, N.; Balakrishna, K.; Parthasarathy, K.; Arulmozhi, N.; Arasu, M.V. A flavonoid isolated from Streptomyces sp.(ERINLG-4) induces apoptosis in human lung cancer A549 cells through p53 and cytochrome c release caspase dependant pathway. Chem. Biol. Interact. 2014, 224, 24–35. [Google Scholar] [CrossRef]
- Kouroshnia, A.; Zeinali, S.; Irani, S.; Sadeghi, A. Induction of apoptosis and cell cycle arrest in colorectal cancer cells by novel anticancer metabolites of Streptomyces sp. 801. Cancer Cell Int. 2022, 22, 235. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.-Y.; Lin, Y.-S.; Shih, S.-P.; Lee, S.-B.; El-Shazly, M.; Chang, K.-M.; Yang, Y.-C.S.; Lee, Y.-L.; Lu, M.-C. The anti-proliferative activity of secondary metabolite from the marine streptomyces sp. against prostate cancer cells. Life 2021, 11, 1414. [Google Scholar] [CrossRef]
- Shirling, E.T.; Gottlieb, D. Methods for characterization of Streptomyces species. Int. J. Syst. Bacteriol. 1966, 16, 313–340. [Google Scholar] [CrossRef] [Green Version]
- Arai, T.; Tamotsu, F.; Masa, H.; Akihiro, M.; Yuzuru, M. Culture Media for Actinomycetes; The Society for Actinomycetes Japan: Tokyo, Japan, 1975; pp. 1–20. [Google Scholar]
- Azmi, M.N.; Sian, T.A.; Suhaimi, M.; Kamarudin, M.N.A.; Din, M.F.M.; Nafiah, M.A.; Thomas, N.F.; Kadir, H.A.; Awang, K. Synthesis of Indolostilbenes via FeCl3-promoted Oxidative Cyclisation and their Biological Effects on NG108-15 Cell Viability and H2O2-induced Cytotoxicity. J. Phys. Sci. 2021, 32, 69–89. [Google Scholar] [CrossRef]
- Lim, J.Y.; Liu, C.; Hu, K.Q.; Smith, D.E.; Wu, D.; Lamon-Fava, S.; Ausman, L.M.; Wang, X.D. Xanthophyll β-Cryptoxanthin Inhibits Highly Refined Carbohydrate Diet–Promoted Hepatocellular Carcinoma Progression in Mice. Mol. Nutr. Food Res. 2020, 64, 1900949. [Google Scholar] [CrossRef] [PubMed]
- Uehara, T.; Pogribny, I.P.; Rusyn, I. The DEN and CCl4-Induced Mouse Model of Fibrosis and Inflammation-Associated Hepatocellular Carcinoma. Curr. Protoc. 2021, 1, e211. [Google Scholar] [CrossRef] [PubMed]
- Haque, M.U.; Rahman, M.A.; Haque, M.A.; Sarker, A.K.; Islam, M.A.U. Antimicrobial and anticancer activities of ethyl acetate extract of co-culture of Streptomyces sp. ANAM-5 and AIAH-10 Isolated from Mangrove Forest of Sundarbans, Bangladesh. J. Appl. Pharm. Sci. 2016, 6, 051–055. [Google Scholar] [CrossRef] [Green Version]
- Li, S.-Q.; Wang, D.-M.; Shu, Y.-J.; Wan, X.-D.; Xu, Z.-S.; Li, E.-Z. Proper heat shock pretreatment reduces acute liver injury induced by carbon tetrachloride and accelerates liver repair in mice. J. Toxicol. Pathol. 2013, 26, 365–373. [Google Scholar] [CrossRef] [Green Version]
- Krishnan, K.; Mani, A.; Jasmine, S. Cytotoxic activity of bioactive compound 1, 2-benzene dicarboxylic acid, mono 2-ethylhexyl ester extracted from a marine derived Streptomyces sp. VITSJK8. Int. J. Mol. Cell. Med. 2014, 3, 246. [Google Scholar]
- Al-Enazi, N.M.; Abdel-Raouf, N.; Alharbi, R.M.; Sholkamy, E.N. Metabolic Profiling of Streptomyces sp. Strain ess_amH1 Isolated from Apis mellifera yemintica’s Gut Microbiome, and Its Anticancer Activity against Breast Cancer (MCF7) and Hepatocarcinoma (HepG2) Cell Lines, as Well as Antimicrobial Activity. Appl. Sci. 2022, 12, 12257. [Google Scholar] [CrossRef]
- Tawfek, N.S.; Al Azhary, D.B.; Hussien, B.K.A.; Abd Elgeleel, D.M. Effects of Cassia fistula and Ficus carica leaf extracts on hepatocarcinogenesis in rats. Middle East J. Appl. Sci. 2015, 5, 462–479. [Google Scholar]
- Tisdale, M.J. Metabolic abnormalities in cachexia and anorexia. Nutrition 2000, 16, 1013–1014. [Google Scholar] [CrossRef]
- Wang, J.; Chu, H.; Wang, Z.; Wang, X.; Liu, X.; Song, Z.; Liu, F. In vivo study revealed pro-tumorigenic effect of CMTM3 in hepatocellular carcinoma involving the regulation of peroxisome proliferator-activated receptor gamma (PPARγ). Cell. Oncol. 2022, 46, 49–64. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Yi, J.-K.; Chen, J.; Huang, H.; Wu, L.; Yin, X.; Wang, J. Boron attenuated diethylnitrosamine induced hepatocellular carcinoma in C3H/HeN mice via alteration of oxidative stress and apoptotic pathway. J. Trace Elem. Med. Biol. 2022, 74, 127052. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Hamid, N.M.; Abass, S.A.; Mohamed, A.A.; Hamid, D.M. Herbal management of hepatocellular carcinoma through cutting the pathways of the common risk factors. Biomed. Pharmacother. 2018, 107, 1246–1258. [Google Scholar] [CrossRef]
- Adebayo, O.A.; Akinloye, O.; Adaramoye, O.A. Cerium oxide nanoparticles attenuate oxidative stress and inflammation in the liver of diethylnitrosamine-treated mice. Biol. Trace Elem. Res. 2020, 193, 214–225. [Google Scholar] [CrossRef]
- El-Nekeety, A.A.; Salman, A.S.; Hathout, A.S.; Sabry, B.A.; Abdel-Aziem, S.H.; Hassan, N.S.; Abdel-Wahhab, M.A. Evaluation of the bioactive extract of actinomyces isolated from the Egyptian environment against aflatoxin B1-induce cytotoxicity, genotoxicity and oxidative stress in the liver of rats. Food Chem. Toxicol. 2017, 105, 241–255. [Google Scholar] [CrossRef] [PubMed]
- Rashwan, H.M.; Mohammed, H.E.; El-Nekeety, A.A.; Hamza, Z.K.; Abdel-Aziem, S.H.; Hassan, N.S.; Abdel-Wahhab, M.A. Bioactive phytochemicals from Salvia officinalis attenuate cadmium-induced oxidative damage and genotoxicity in rats. Environ. Sci. Pollut. Res. Int. 2021, 28, 68498–68512. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.B.; Le, X.-T.T.; Nguyen, H.H.; Vo, T.T.; Le, M.K.; Nguyen, N.T.; Do-Nguyen, T.M.; Truong-Nguyen, C.M.; Nguyen, B.-S.T. Diagnostic Value of hTERT mRNA and in Combination With AFP, AFP-L3%, Des-γ-carboxyprothrombin for Screening of Hepatocellular Carcinoma in Liver Cirrhosis Patients HBV or HCV-Related. Cancer Inform. 2022, 21, 11769351221100730. [Google Scholar] [CrossRef]
- Lee, T.K.-W.; Guan, X.-Y.; Ma, S. Cancer stem cells in hepatocellular carcinoma—From origin to clinical implications. Nat. Rev. Gastroenterol. Hepatol. 2022, 19, 26–44. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Turpin, C.; Qiu, G.; Xu, M.; Lee, E.; Hinds, T.D., Jr.; Peterson, M.L.; Spear, B.T. Zinc fingers and homeoboxes 2 is required for diethylnitrosamine-induced liver tumor formation in C57BL/6 mice. Hepatol. Commun. 2022, 6, 3550–3562. [Google Scholar] [CrossRef]
- Xu, S.-H.; Luo, H.-X.; Huang, B.-J.; Yu, L.; Luo, S.-J.; Hu, H.; Li, Y.; Lin, X.-T.; Cao, Z.-R.; Deng, Y.-J. Therapeutic Effect of Catgut Implantation at Acupoint in a Mouse Model of Hepatocellular Carcinoma by Suppressing Immune Escape. Evid. Based Complement. Altern. Med. 2022, 2022, 5572869. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, N.A.; Anwar, H.M.; Moghazy, A.M.; El Malah, T.; Ragab, W.M.; El-Aal, A.; Hassan, R.A.; Saleh, N.A.; Eldosoki, D.E. Heme oxygenase–1 Expression in Liver and Colon of Rats Exposed to Oxidative stress and Dysplasia by a Carcinogen Diethylnitrosamine and the Possible Therapeutic Effects of Probiotic Versus Pyridazine Derivative and Chemotherapy. Egypt. J. Chem. 2022, 65, 1–2. [Google Scholar] [CrossRef]
- Bishayee, A. The role of inflammation and liver cancer. Adv. Exp. Med. Biol. 2014, 816, 401–435. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Liu, N.-N.; Li, J.-R.; Wang, M.-X.; Tan, J.-L.; Dong, B.; Lan, P.; Zhao, L.-M.; Peng, Z.-G.; Jiang, J.-D. Bicyclol ameliorates advanced liver diseases in murine models via inhibiting the IL-6/STAT3 signaling pathway. Biomed. Pharmacother. 2022, 150, 113083. [Google Scholar] [CrossRef] [PubMed]
- Muhammed, A.; Fulgenzi, C.A.M.; Dharmapuri, S.; Pinter, M.; Balcar, L.; Scheiner, B.; Marron, T.U.; Jun, T.; Saeed, A.; Hildebrand, H. The systemic inflammatory response identifies patients with adverse clinical outcome from immunotherapy in hepatocellular carcinoma. Cancers 2022, 14, 186. [Google Scholar] [CrossRef]
- Arvanitakis, K.; Koletsa, T.; Mitroulis, I.; Germanidis, G. Tumor-Associated Macrophages in Hepatocellular Carcinoma Pathogenesis, Prognosis and Therapy. Cancers 2022, 14, 226. [Google Scholar] [CrossRef]
- Kim, M.E.; Na, J.Y.; Lee, J.S. Anti-inflammatory effects of trans-cinnamaldehyde on lipopolysaccharide-stimulated macrophage activation via MAPKs pathway regulation. Immunopharmacol. Immunotoxicol. 2018, 40, 219–224. [Google Scholar] [CrossRef]
- Gaikwad, N.; Nanduri, S.; Madhavi, Y.V. Cinnamamide: An insight into the pharmacological advances and structure-activity relationships. Eur. J. Med. Chem. 2019, 181, 111561. [Google Scholar] [CrossRef]
- Balthazar, J.D.; Soosaimanickam, M.P.; Emmanuel, C.; Krishnaraj, T.; Sheikh, A.; Alghafis, S.F.; Ibrahim, H.-I.M. 8-Hydroxyquinoline a natural chelating agent from Streptomyces spp. inhibits A549 lung cancer cell lines via BCL2/STAT3 regulating pathways. World J. Microbiol. Biotechnol. 2022, 38, 1–12. [Google Scholar] [CrossRef]
- Farghadani, R.; Naidu, R. Curcumin as an enhancer of therapeutic efficiency of chemotherapy drugs in breast cancer. Int. J. Mol. Sci. 2022, 23, 2144. [Google Scholar] [CrossRef]
- Green, D.R. The mitochondrial pathway of apoptosis Part II: The BCL-2 protein family. Cold Spring Harb. Perspect. Biol. 2022, 14, a041046. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′-3′) | Reverse Primer (5′-3′) |
|---|---|---|
| AFP | CCAGGAAGTCTGTTTCACAGAAG | CAAAAGGCTCACACCAAAGAG |
| P53 | TGAAACGCCGACCTATCCTTA | GGCACAAACACGAACCTCAAA |
| IL-1β | CTATGGCAACTGTCCCTGAA | GGCTTGGAAGCAATCCTT |
| TNF-α | GCTTGGTGGTTTGCTACGAC | ACTGAACTTCGGGGTGATTG |
| GAPDH | AAGGTGGAAGAATGGGAGTT | GGAAAGCTGTGGCGTGAT |
| No. | Detected Compound | Formula | Retention Time (min) | Peak Area | Abundance (%) |
|---|---|---|---|---|---|
| 1 | Ethyl Acetate | C4H8O2 | 1.80 | 86,851,429 | 44.307 |
| 2 | Propanoic acid, ethyl ester | C5H10O2 | 2.37 | 2,576,397 | 1.314 |
| 3 | p-Xylene | C8H10 | 4.65 | 1,201,182 | 0.613 |
| 4 | 1,3,5-Cyclooctatriene | C8H10 | 5.18 | 345,979 | 0.176 |
| 5 | Benzaldehyde | C7H6O | 6.80 | 194,101 | 0.099 |
| 5 | 5-Methyl-2-furaldehyde | C6H6O2 | 6.95 | 321,237 | 0.164 |
| 6 | 2-Methyl-5-hexanone oxime | C7H15NO | 8.15 | 2,498,550 | 1.275 |
| 7 | Corylon | C6H8O2 | 8.64 | 230,347 | 0.118 |
| 8 | Phenylethyl Alcohol | C8H10O | 11.05 | 366,472 | 0.187 |
| 9 | Larixic acid | C6H6O3 | 11.20 | 331,418 | 0.169 |
| 10 | Pyranone | C6H8O4 | 11.95 | 150,614 | 0.077 |
| 11 | 2,4,4-Trimethyl-1-pentyl methylphosphonofluoridate | C9H20FO2P | 13.62 | 460,008 | 0.235 |
| 12 | Pyrrole-2-carboxylic acid | C5H5NO2 | 14.20 | 6,297,782 | 3.213 |
| 14 | trans-Cinnamaldehyde | C9H8O | 15.43 | 14,191,887 | 7.240 |
| 15 | Pyrrole-2-carboxamide | C5H6N2O | 16.59 | 14,191,887 | 7.240 |
| 16 | Benzamide | C7H7NO | 17.55 | 1,141,967 | 0.583 |
| 17 | α-ylangene | C15H24 | 18.23 | 257,653 | 0.131 |
| 18 | Benzeneacetamide | C8H9NO | 19.12 | 22,818,275 | 11.641 |
| 19 | Phenylpropanamide | C9H11NO | 22.06 | 1,628,020 | 0.831 |
| 20 | Cinnamamide | C9H9NO | 25.23 | 10,206,505 | 5.207 |
| 21 | Uric acid | C5H4N4O3 | 26.45 | 1,488,722 | 0.759 |
| 22 | 3-(Hydroxymethyl)-5-methoxyphenol | C8H10O3 | 28.65 | 1,955,728 | 0.998 |
| 23 | Cyclo(leucyloprolyl) | C11H18N2O2 | 30.93 | 8,920,233 | 4.551 |
| 24 | Chiapin B | C19H26O6 | 31.82 | 3,618,392 | 1.846 |
| 25 | cis-13-Eicosenoic acid | C20H38O2 | 33.68 | 880,571 | 0.449 |
| 26 | 2,5-Piperazinedione, 3,6-bis(2-methylpropyl | C12H22N2O2 | 35.50 | 1,801,612 | 0.919 |
| 27 | (Z)-9-Octadecenamide | C18H35NO | 38.61 | 5,237,385 | 2.672 |
| 28 | Dihydroergotamine | C33H37N5O5 | 38.95 | 4,316,002 | 2.202 |
| 29 | cis-5,8,11,14,17-Eicosapentaenoic acid | C20H30O2 | 39.36 | 1,542,283 | 0.787 |
| Parameters/Groups | Control | A16 (L) | A16 (H) | DEN | DEN+ A16 (L) | DEN + A16 (H) |
|---|---|---|---|---|---|---|
| Body weight(g) week32 | 43 ± 1.6 | 43.6 ± 1.6 | 43.5 ± 3 | 38.8 ± 0.8 *** | 39.6 ± 1.6 ### | 38.9 ± 0.6 ## |
| Body weight(g) week36 | 44.4 ± 1.6 | 43.8 ± 1.9 | 44.8 ± 1.9 | 41.0 ± 0.9 *** | 43.0 ± 2.0 ### | 43.6 ± 0.6 ## |
| Absolute liver weight (g) | 2.3 ± 0.06 | 2.3 ± 0.1 | 2.1 ± 0.1 | 3.3 ± 0.08 ** | 2.9 ± 0.08 ## | 2.7 ± 0.08 ### |
| AST (U/L) | 106.4 ± 1.11 | 112.6 ± 2.72 | 112.8 ± 0.44 | 227.9 ± 5.74 *** | 116.7 ± 0.88 ### | 109.0 ± 2.17 ### |
| ALT (U/L) | 37.35 ± 0.5 | 40.51 ± 0.46 | 42.32 ± 0.43 | 125.5 ± 1.5 *** | 48.76 ± 0.53 ### | 41.00 ± 1.11 ### |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alhawsawi, S.M.; Mohany, M.; Baabbad, A.A.; Almoutiri, N.D.; Maodaa, S.N.; Al-shaebi, E.M.; Yaseen, K.N.; Wadaan, M.A.M.; Hozzein, W.N. Streptomyces Bioactive Metabolites Prevent Liver Cancer through Apoptosis, Inhibiting Oxidative Stress and Inflammatory Markers in Diethylnitrosamine-Induced Hepatocellular Carcinoma. Biomedicines 2023, 11, 1054. https://doi.org/10.3390/biomedicines11041054
Alhawsawi SM, Mohany M, Baabbad AA, Almoutiri ND, Maodaa SN, Al-shaebi EM, Yaseen KN, Wadaan MAM, Hozzein WN. Streptomyces Bioactive Metabolites Prevent Liver Cancer through Apoptosis, Inhibiting Oxidative Stress and Inflammatory Markers in Diethylnitrosamine-Induced Hepatocellular Carcinoma. Biomedicines. 2023; 11(4):1054. https://doi.org/10.3390/biomedicines11041054
Chicago/Turabian StyleAlhawsawi, Sana M., Mohamed Mohany, Almohannad A. Baabbad, Nawaf D. Almoutiri, Saleh N. Maodaa, Esam M. Al-shaebi, Khadijah N. Yaseen, Mohammed A. M. Wadaan, and Wael N. Hozzein. 2023. "Streptomyces Bioactive Metabolites Prevent Liver Cancer through Apoptosis, Inhibiting Oxidative Stress and Inflammatory Markers in Diethylnitrosamine-Induced Hepatocellular Carcinoma" Biomedicines 11, no. 4: 1054. https://doi.org/10.3390/biomedicines11041054
APA StyleAlhawsawi, S. M., Mohany, M., Baabbad, A. A., Almoutiri, N. D., Maodaa, S. N., Al-shaebi, E. M., Yaseen, K. N., Wadaan, M. A. M., & Hozzein, W. N. (2023). Streptomyces Bioactive Metabolites Prevent Liver Cancer through Apoptosis, Inhibiting Oxidative Stress and Inflammatory Markers in Diethylnitrosamine-Induced Hepatocellular Carcinoma. Biomedicines, 11(4), 1054. https://doi.org/10.3390/biomedicines11041054